















































 Биология
БиологияПохожие презентации:








. Обмен гликогена. Особенности обмена углеводов в различных органах и тканях. Лекция №6")

Глюконеогенез. Обмен гликогена. Регуляция и патология углеводного обмена
1.
2.
ПЛАН ЛЕКЦИИ1. Основные этапы (схема и реакции) глюконеогенеза.
2. Пути включения в глюконеогенез лактата, глицерола
и аминокислот.
3. Этапы синтеза гликогена. Особенности
мобилизации гликогена в печени и мышцах
4. Субстратная регуляция углеводного обмена.
5. Гормональная регуляция углеводного обмена.
6. Гипогликемия и гипергликемия (виды, причины,
механизмы, последствия).
7. Нарушения углеводного обмена.
3.
Глюконеогенез –– процесс синтеза глюкозы извеществ неуглеводной природы.
Основная функция:
поддержание уровня глюкозы в крови в период
длительного
голодания
и
интенсивных
физических нагрузок.
4.
Процесс протекает в основном в печени,менее интенсивно в корковом веществе
почек, а также в слизистой оболочке
кишечника.
5.
Первичные субстраты глюконеогенезаЛактат - продукт анаэробного гликолиза.
Образуется при любых состояниях организма в
эритроцитах и работающих мышцах.
Глицерол – высвобождается при гидролизе
жиров в жировой ткани в период голодания или при
длительной физической нагрузке.
Аминокислоты
образуются
распада мышечных белков.
в
результате
6.
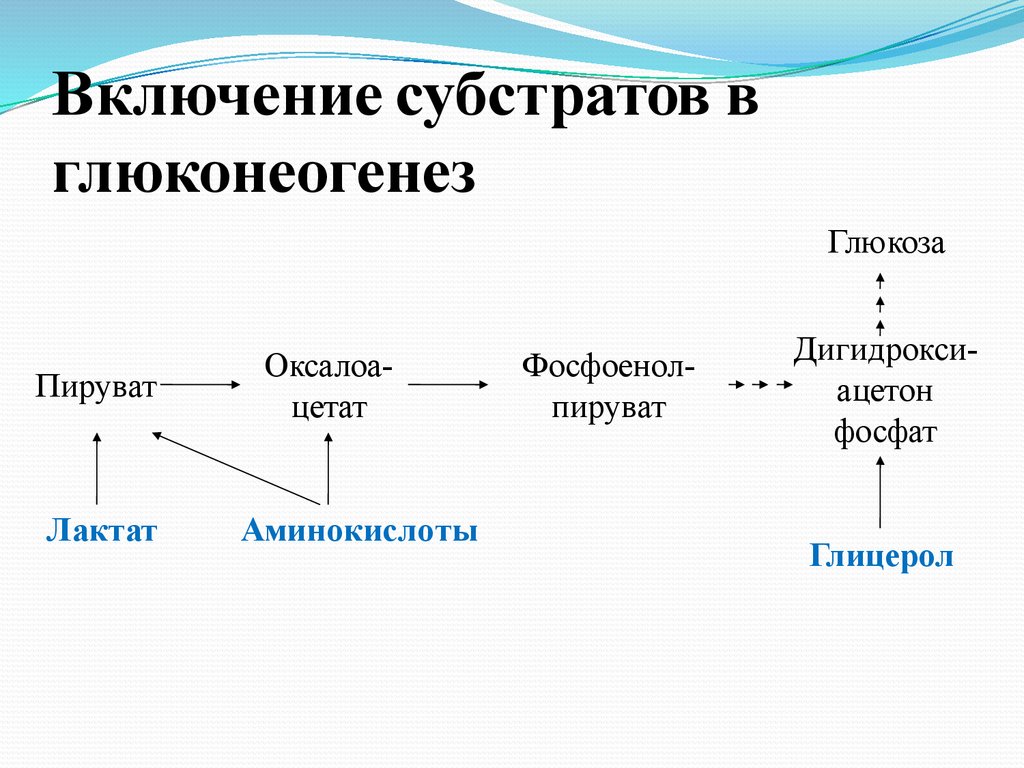
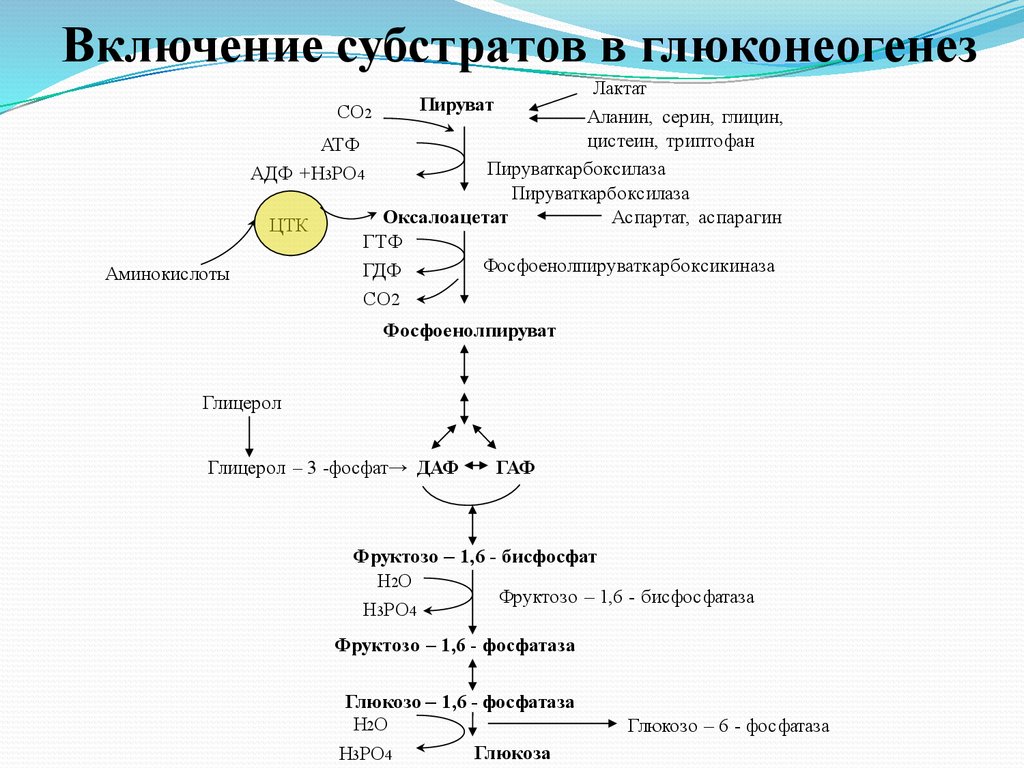
Включение субстратов вглюконеогенез
Глюкоза
Пируват
Лактат
Оксалоацетат
Аминокислоты
Фосфоенолпируват
Дигидроксиацетон
фосфат
Глицерол
7.
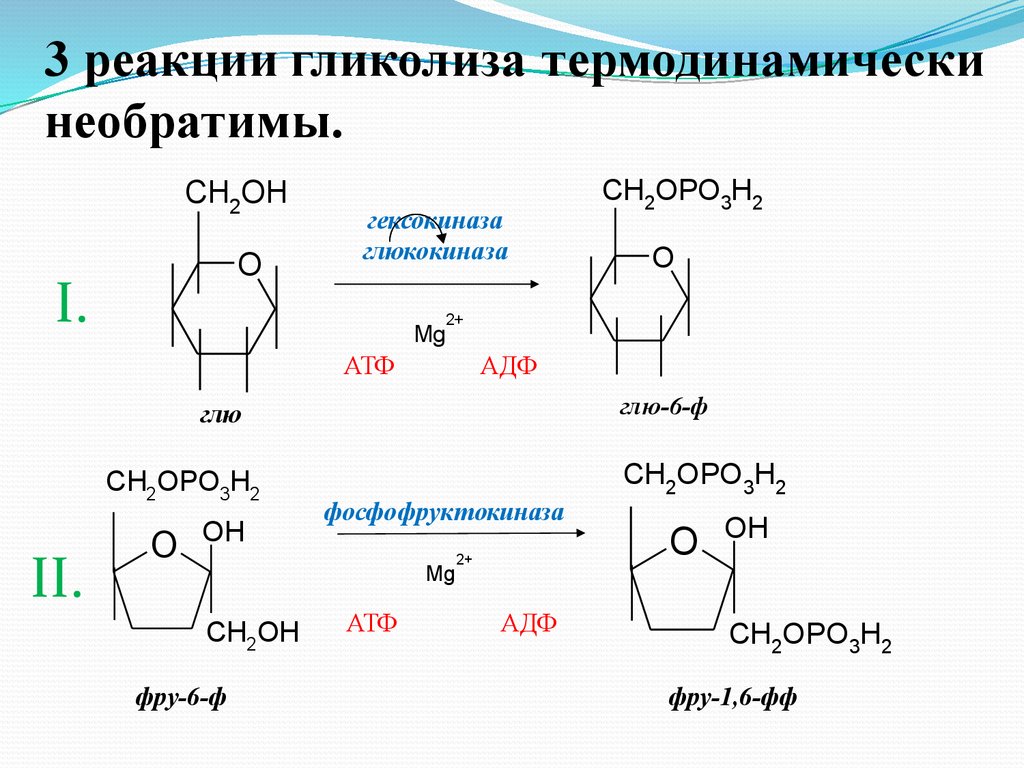
3 реакции гликолиза термодинамическинеобратимы.
CH2OH
O
I.
гексокиназа
глюкокиназа
O
2+
Mg
АТФ
АДФ
глю-6-ф
глю
CH2OPO3H2
II.
CH2OPO3H2
O
OH
фосфофруктокиназа
Mg
CH2OH
фру-6-ф
АТФ
CH2OPO3H2
O
2+
АДФ
OH
CH2OPO3H2
фру-1,6-фф
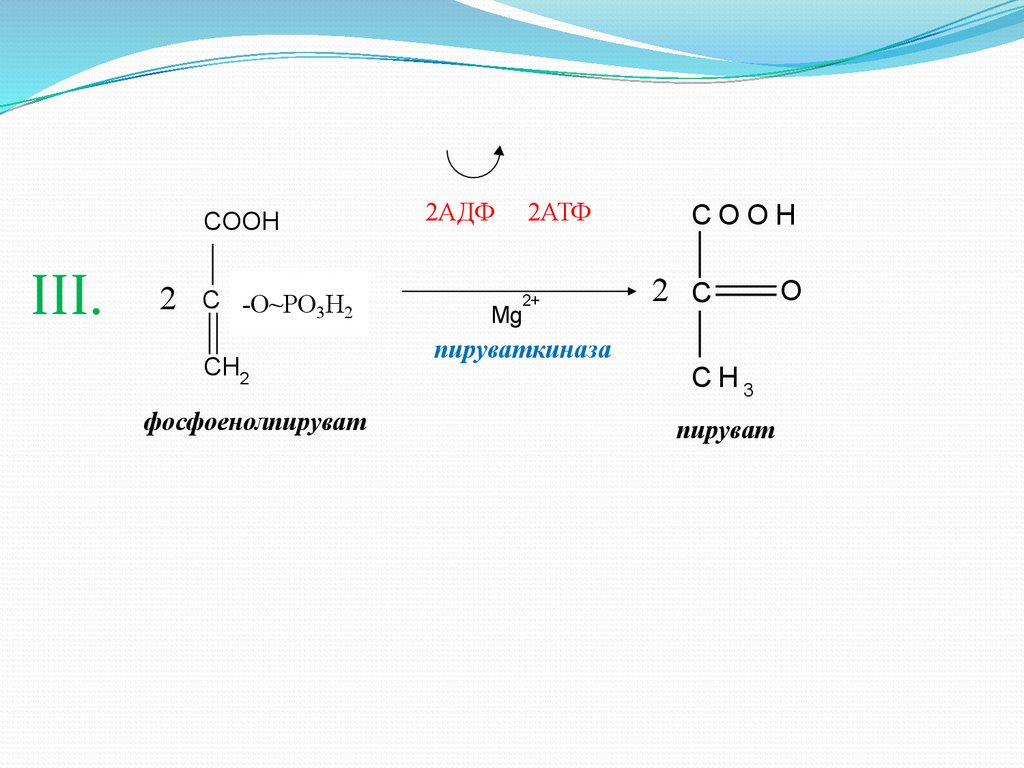
8.
COOHIII.
2
C -О~PO
OPO3H
H
3 22
CH2
фосфоенолпируват
2АДФ
2АТФ
2+
Mg
COOH
2 C
пируваткиназа
CH3
пируват
O
9.
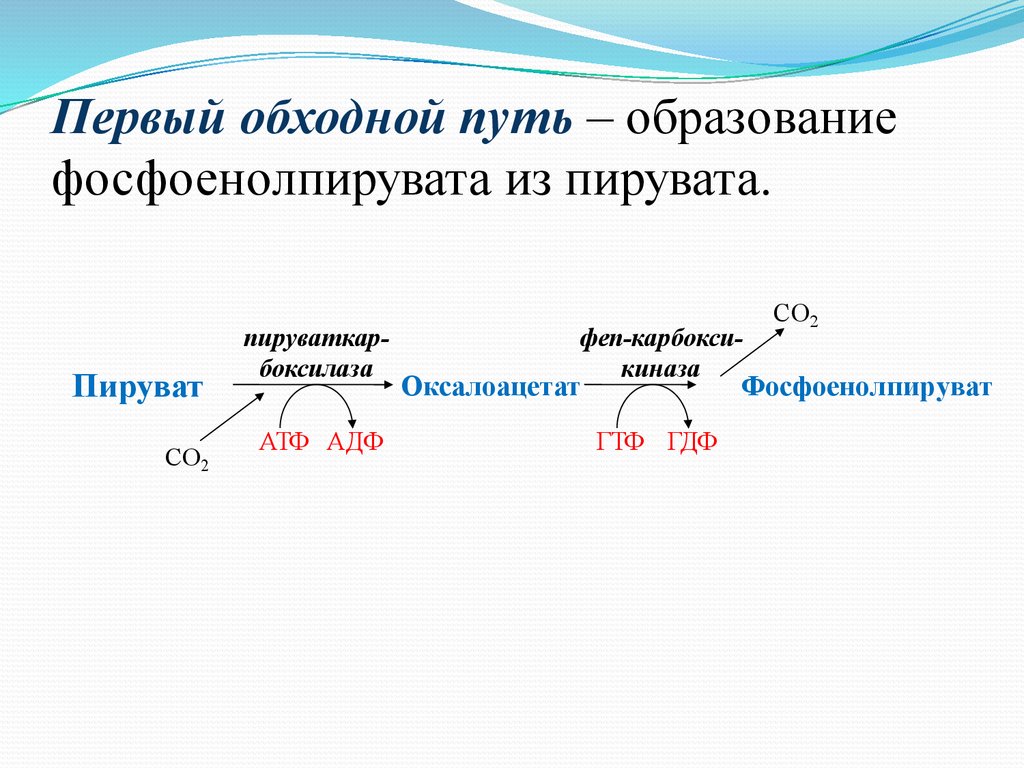
Первый обходной путь – образованиефосфоенолпирувата из пирувата.
Пируват
CO2
пируваткарбоксилаза
АТФ АДФ
феп-карбоксикиназа
Оксалоацетат
CO2
Фосфоенолпируват
ГТФ ГДФ
10.
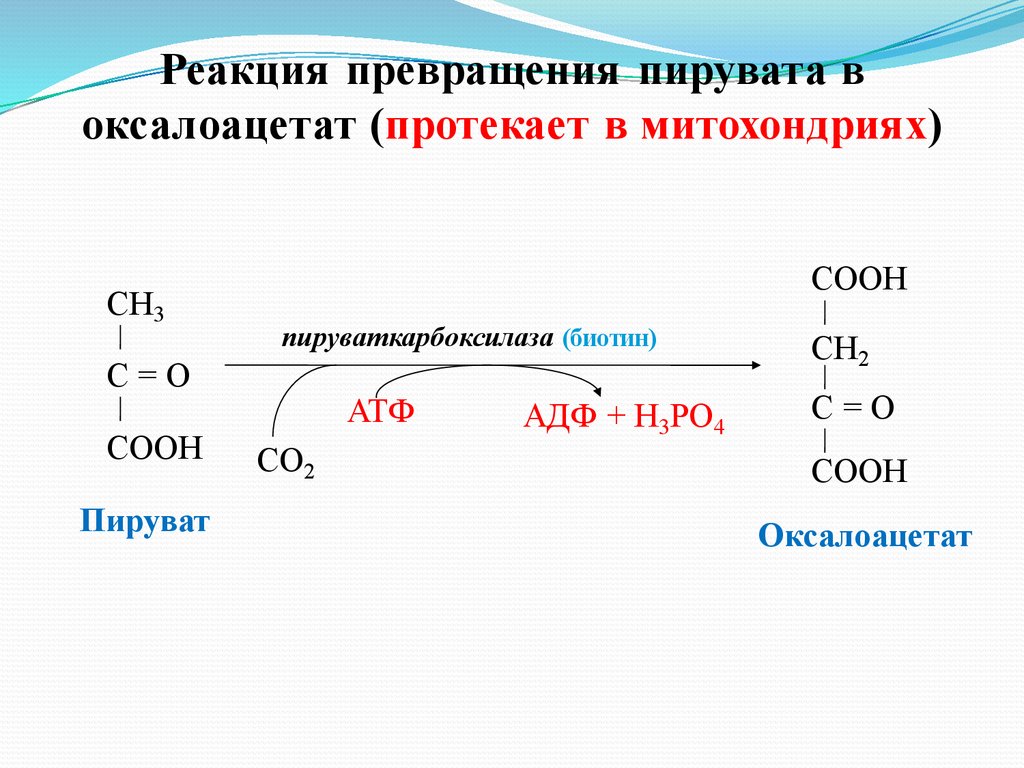
Реакция превращения пирувата воксалоацетат (протекает в митохондриях)
СН3
СООН
пируваткарбоксилаза (биотин)
С=О
СООН
Пируват
АТФ
СО2
АДФ + Н3РО4
СН2
С=О
СООН
Оксалоацетат
11.
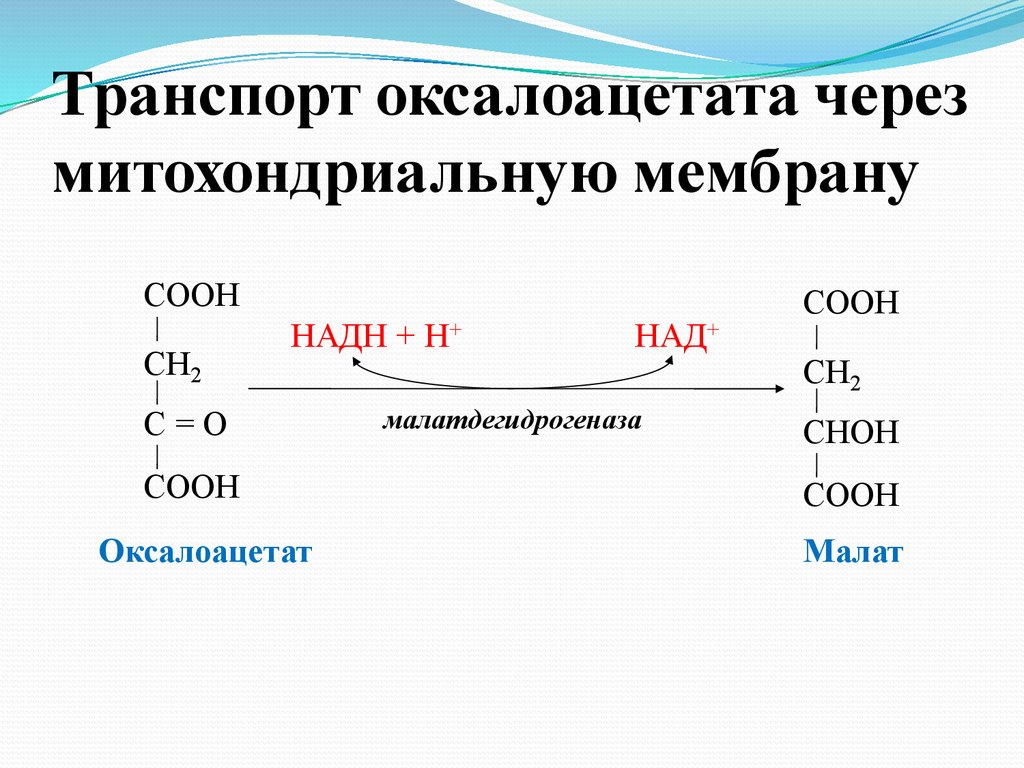
Транспорт оксалоацетата черезмитохондриальную мембрану
СООН
СН2
НАДН + Н+
С=О
СООН
Оксалоацетат
НАД+
малатдегидрогеназа
СООН
СН2
СHOH
СООН
Малат
12.
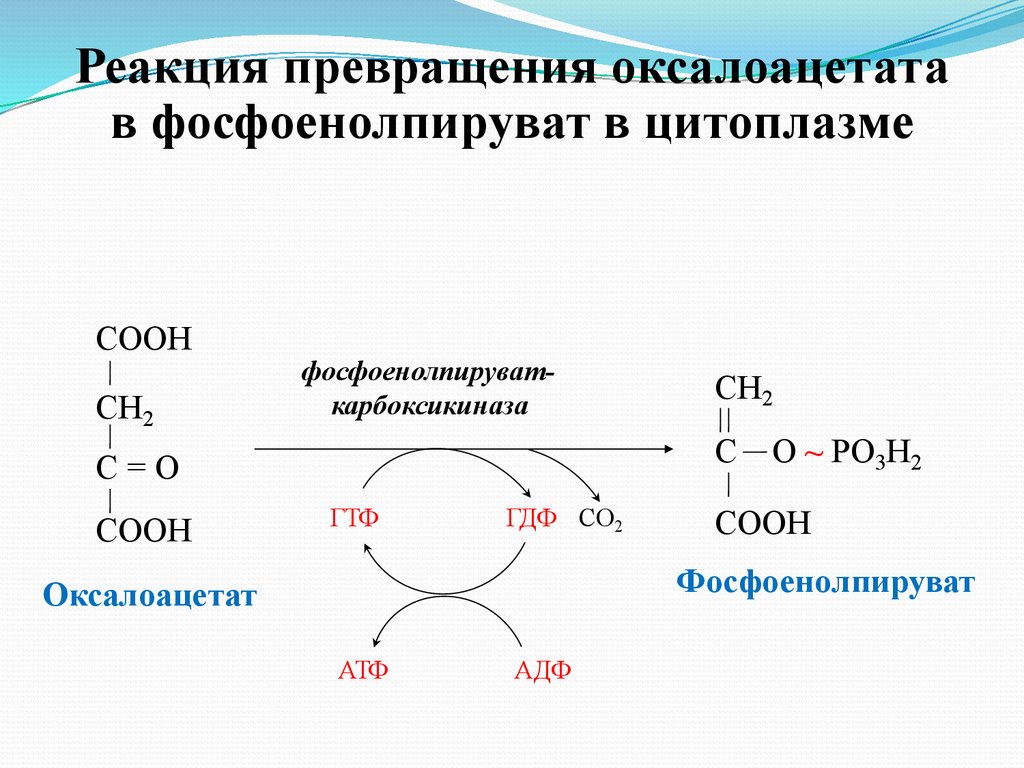
Реакция превращения оксалоацетатав фосфоенолпируват в цитоплазме
СООН
СН2
фосфоенолпируваткарбоксикиназа
С
С=О
СООН
СН2
ГТФ
ГДФ СО2
О ~ PO3Н2
СООН
Фосфоенолпируват
Оксалоацетат
АТФ
АДФ
13.
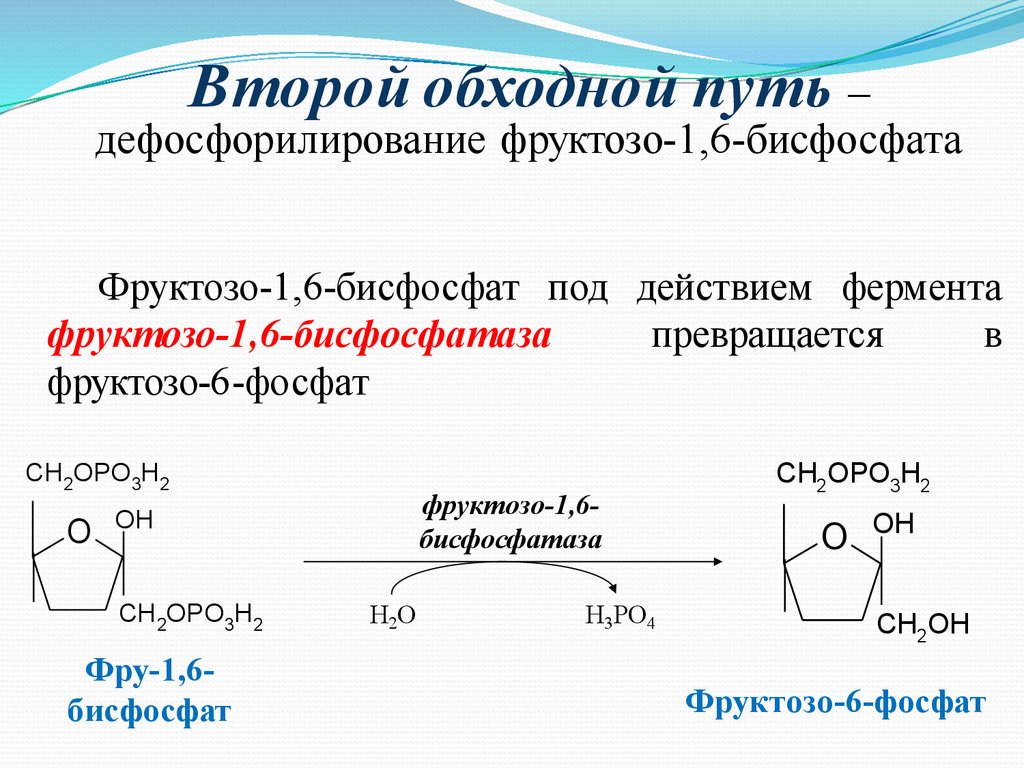
Второй обходной путь –дефосфорилирование фруктозо-1,6-биcфосфата
Фруктозо-1,6-бисфосфат под действием фермента
фруктозо-1,6-бисфосфатаза
превращается
в
фруктозо-6-фосфат
CH2OPO3H2
O
фруктозо-1,6бисфосфатаза
OH
CH2OPO3H2
Фру-1,6бисфосфат
Н2О
Н3РО4
CH2OPO3H2
O
OH
CH2OH
Фруктозо-6-фосфат
14.
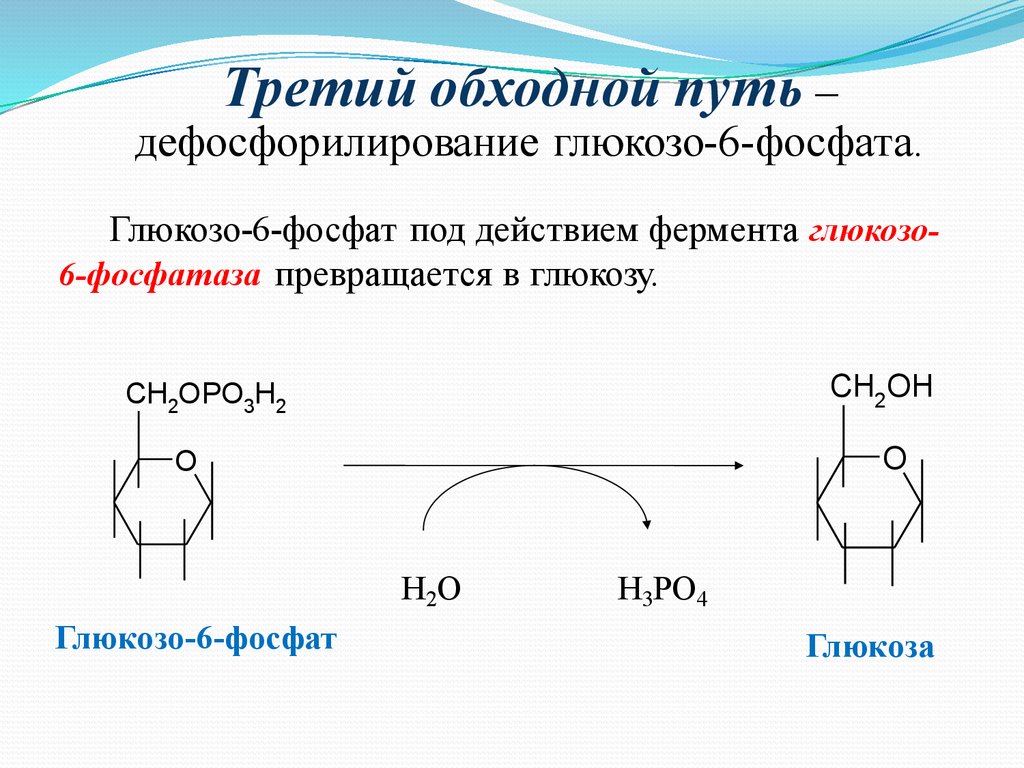
Третий обходной путь –дефосфорилирование глюкозо-6-фосфата.
Глюкозо-6-фосфат под действием фермента глюкозо6-фосфатаза превращается в глюкозу.
CH2OH
CH2OPO3H2
O
O
Н2О
Глюкозо-6-фосфат
Н3РО4
Глюкоза
15.
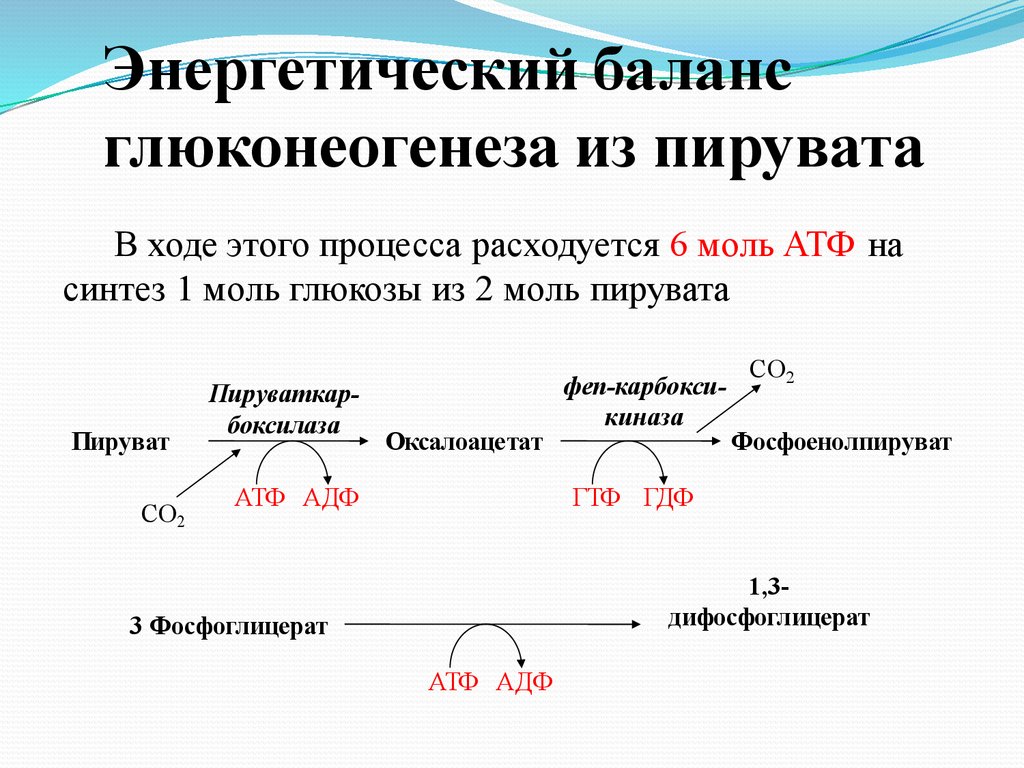
Энергетический балансглюконеогенеза из пирувата
В ходе этого процесса расходуется 6 моль АТФ на
синтез 1 моль глюкозы из 2 моль пирувата
Пируват
CO2
Пируваткарбоксилаза
Оксалоацетат
АТФ АДФ
феп-карбоксикиназа
CO2
Фосфоенолпируват
ГТФ ГДФ
1,3дифосфоглицерат
3 Фосфоглицерат
АТФ АДФ
16.
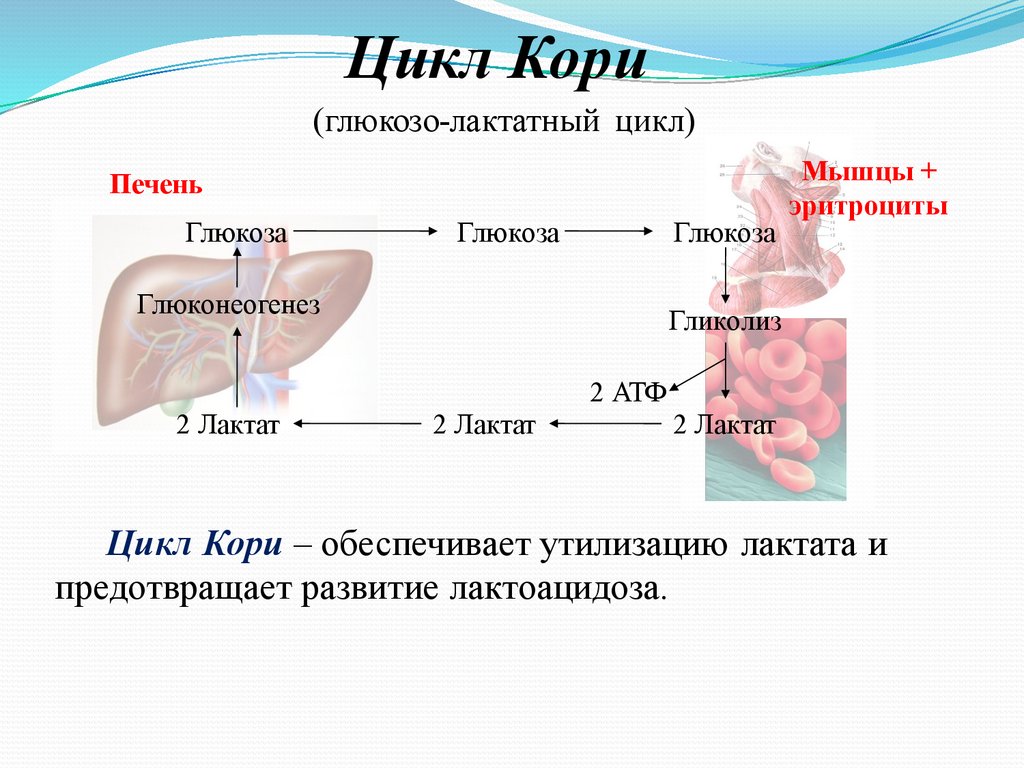
Цикл Кори(глюкозо-лактатный цикл)
Печень
Глюкоза
Глюкоза
Глюкоза
Глюконеогенез
Мышцы +
эритроциты
Гликолиз
2 АТФ
2 Лактат
2 Лактат
2 Лактат
Цикл Кори – обеспечивает утилизацию лактата и
предотвращает развитие лактоацидоза.
17.
Нарушение метаболизма прилактоацидозе
Лактат
Глюкоза
Пируват
1.
2.
Ацетил-КоА
СО2, Н2О
1. – Нарушение использования пирувата в глюконеогенезе
2. – Нарушение окисления пирувата
18.
Глюкозо-аланиновый циклПечень
Мышца
ОПК
СО2
Н2О
Глюкоза
Глюкоза
Пируват
Пируват
Энергия
Аланин
ОПК
СО2
Аланин
Н 2О
Глюкозо-аланиновый цикл осуществляет транспорт
аминного азота из мышц в печень и предотвращает
лактоацидоз.
Энергия
19.
Включение субстратов в глюконеогенезЛактат
СО2
Аланин, серин, глицин,
цистеин, триптофан
АТФ
Пируваткарбоксилаза
АДФ +Н3РО4
Пируваткарбоксилаза
Оксалоацетат
Аспартат, аспарагин
ЦТК
ГТФ
Фосфоенолпируваткарбоксикиназа
ГДФ
СО2
Пируват
Аминокислоты
Фосфоенолпируват
Глицерол
Глицерол – 3 -фосфат→ ДАФ
ГАФ
Фруктозо – 1,6 - бисфосфат
Н2О
Фруктозо – 1,6 - бисфосфатаза
Н3РО4
Фруктозо – 1,6 - фосфатаза
Глюкозо – 1,6 - фосфатаза
Н2О
Глюкоза
Н3РО4
Глюкозо – 6 - фосфатаза
20.
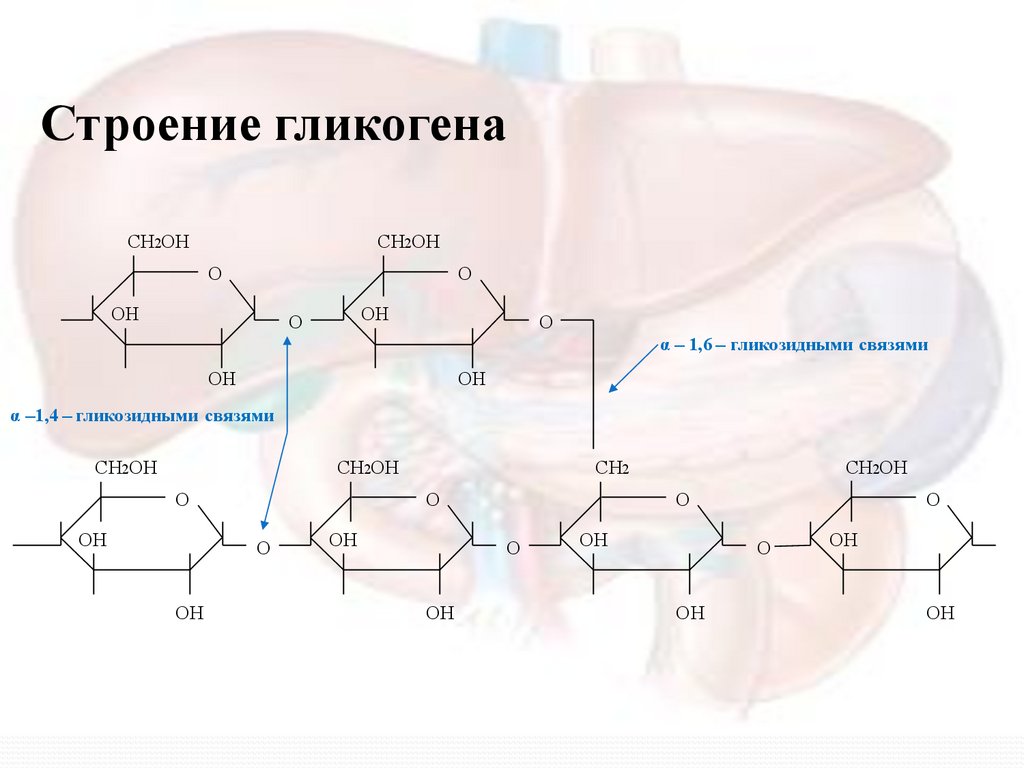
Строение гликогенаСН2ОН
СН2ОН
О
О
ОН
ОН
О
О
α – 1,6 – гликозидными связями
ОН
ОН
α –1,4 – гликозидными связями
СН2ОН
СН2ОН
О
ОН
О
О
ОН
СН2
ОН
О
О
ОН
СН2ОН
ОН
О
О
ОН
ОН
ОН
21.
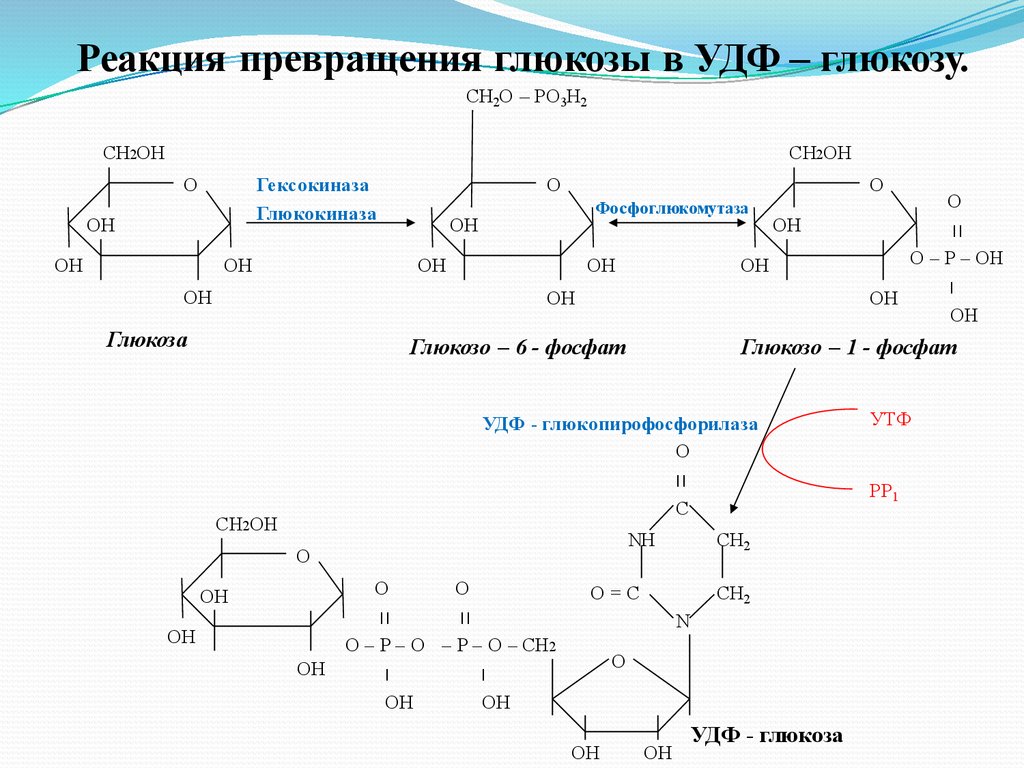
Синтез гликогена (гликогеногенез) ускоряетсяв период пищеварения, когда глюкоза
активно поступает из крови в ткани и
фосфорилируется, превращаясь в глюкозо –
6 – фосфат. Затем глюкозо – 6- фосфат
превращается фосфоглюкомутазой в
глюкозо – 1 – фосфат, из которой под
действием ( УДФ) – глюкопирофосфорилазы
и при участии ( УТФ) образуется УДФ –
глюкоза.
22.
Это соединение является субстратомгликогенсинтетазы. Фермент переносит
остаток глюкозы на олигосахарид,
состоящий из 6-10 остатков глюкозы и
представляющий собой праймер (затравку),
присоединяя её α –1,4 – гликозидной
связью. Поскольку праймер редуцирующим
концом соединён с ОН-группой остатка
тирозина белка гликогенина, то
гликогенсинтетаза последовательно
присоединяет глюкозу к нередуцирующему
концу.
23.
Когда количество мономеров всинтезирующемся полисахариде достигает
11-12 моносахаридных остатков, фермент
ветвления переносит фрагмент,
содержащий 6-8 мономеров, от конца
молекулы ближе к её середине и
присоединяет его α –1,6 – гликозидной
связью.
В итоге образуется сильно разветвлённый
гомополисахарид, который может содержать
от 6 тыс. до 1 млн остатков глюкозы.
24.
Синтезгликогена
(гликогенез).
Гликоген
25.
Синтез гликогена идёт с большой затратойэнергии.
Для присоединения 1 остатка глюкозы к
растущей цепи гликогена требуется 2 молекулы
АТФ.
26.
Мобилизация гликогена( гликогенолиз) ускоряется при
повышении потребности организма в
глюкозе.
Гликоген печени распадается в основном в
интервалах между приёмами пищи.
Гликогенолиз в печени и мышцах
ускоряется во время физической работы.
27.
Фермент гликогенфосфорилазарасщепляет α –1,4 – гликозидные связи при
участии Н3РО4 последовательно отщепляет
остатки глюкозы от нередуцирующих
концов молекулы гликогена и
фосфорилирует их с образованием глюкозо
– 1-фосфата. Это приводит к укорочению
ветвей.
28.
Когда количество остатков глюкозы вветвях гликогена достигает 4, то
фермент олигосахаридтрансфераза
расщепляет α –1,4 – гликозидную связь и
переносит фрагмент, состоящий из 3
мономеров, к концу более длинной
цепи.
Фермент α –1,6 – гликозидаза
гидролилизует α –1,6 – гликозидную
связь в точке ветвления и отщепляет
молекулу глюкозы.
29.
При мобилизации гликогена восновном образуется глюкозо – 1фосфат и небольшое количество
свободной глюкозы.
Глюкозо -1-фосфат при участии
фермента фосфоглюкомутаза
превращается в глюкозо -6-фосфат.
30.
Реакция превращения глюкозы в УДФ – глюкозу.СН2О – РО3Н2
СН2ОН
СН2ОН
Гексокиназа
Глюкокиназа
О
ОН
ОН
ОН
О
О
Фосфоглюкомутаза
ОН
ОН
ОН
ОН
ОН
ОН
ОН
Глюкоза
Глюкозо – 6 - фосфат
Глюкозо – 1 - фосфат
УДФ - глюкопирофосфорилаза
О
׀׀
C
СН2ОН
NH
О
ОН
ОН
ОН
О
О
׀׀
׀׀
О – Р – О – Р – О – СН2
׀
׀
ОН
ОН
CH2
O=C
CH2
N
О
ОН
О
׀׀
О – Р – ОН
׀
ОН
ОН
ОН
УДФ - глюкоза
УТФ
РР1
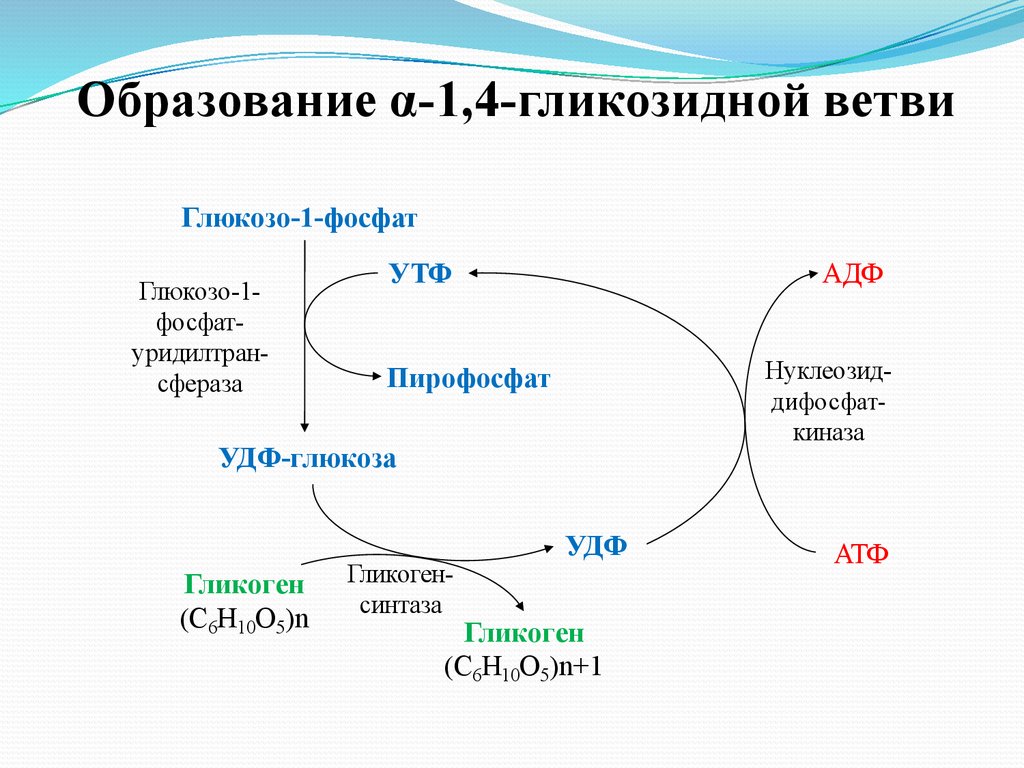
31.
Образование α-1,4-гликозидной ветвиГлюкозо-1-фосфат
Глюкозо-1фосфатуридилтрансфераза
УТФ
АДФ
Нуклеозиддифосфаткиназа
Пирофосфат
УДФ-глюкоза
Гликоген
(С6Н10О5)n
Гликогенсинтаза
УДФ
Гликоген
(С6Н10О5)n+1
АТФ
32.
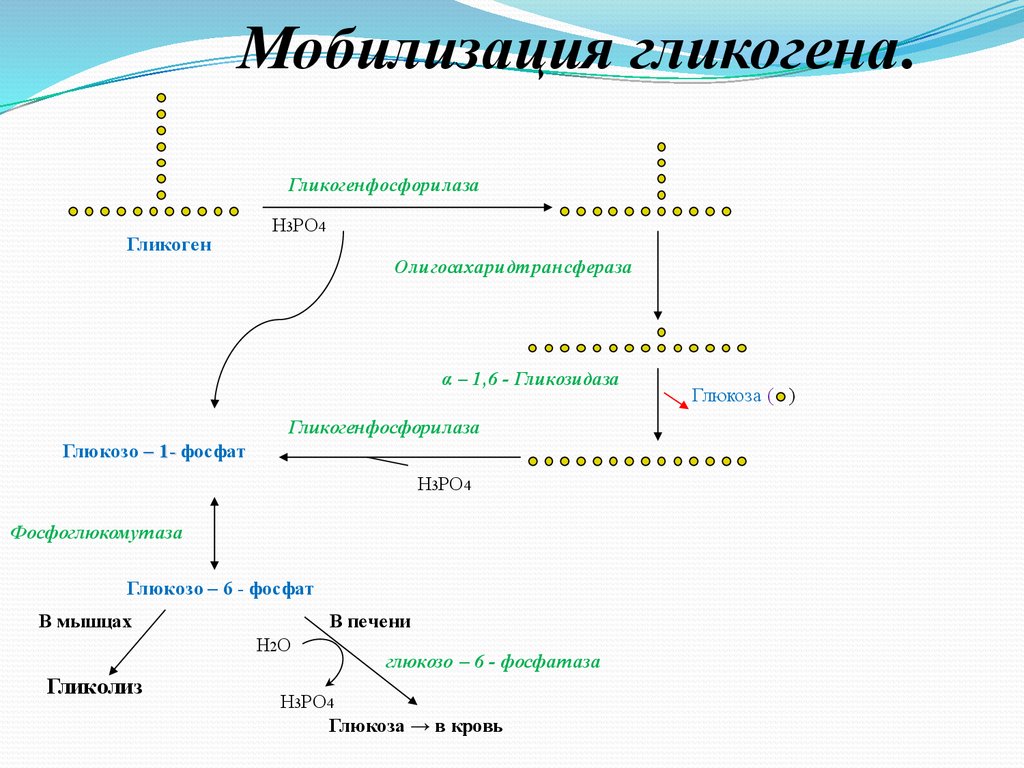
Мобилизация гликогена.Гликогенфосфорилаза
Гликоген
Н3РО4
Олигосахаридтрансфераза
α – 1,6 - Гликозидаза
Гликогенфосфорилаза
Глюкозо – 1- фосфат
Н3РО4
Фосфоглюкомутаза
Глюкозо – 6 - фосфат
В мышцах
В печени
Н2О
Гликолиз
глюкозо – 6 - фосфатаза
Н3РО4
Глюкоза → в кровь
Глюкоза ( )
33.
Мобилизация гликогена в печени и мышцах идётодинаково до образования глюкозо-6-фосфата.
В печени под действием глюкозо-6-фосфатазы
глюкозо-6-фосфат превращается в свободную
глюкозу, которая поступает в кровь и
доставляется в периферические ткани.
34.
Следовательно, мобилизация гликогена впечени обеспечивает сохранение нормального
уровня глюкозы в крови и снабжение глюкозой
других тканей.
Запасы гликогена в печени при голодании
исчерпываются в течении суток.
35.
Биологическое значение обменагликогена
I. Синтез и распад гликогена протекают по разным
метаболическим путям;
II. Печень запасает глюкозу в виде гликогена для
поддержания постоянной концентрации глюкозы в крови;
III. Функция
мышечного
гликогена
заключается
в
освобождении глюкозо-6-фосфата, потребляемого в самой
мышце для окисления и использования энергии;
IV. Синтез гликогена – процесс эндергонический;
V. Распад гликогена до глюкозо-6-фосфата не требует
энергии;
36.
37.
Регуляция углеводного обмена(по механизмам, локализации)
Внутриклеточная
(субстратная)
Внеклеточная
Путем изменения
концентрации
субстрата в крови:
глюкозы,
лактата,
аланина
Гормональная
38.
Внутриклеточные рецепторы.Субстраты и промежуточные продукты метаболических
реакций играют важную роль в регуляции соотношения между
гликолизом и глюконеогенезом.
Рассмотрим, как регулируется само вступление остатков
глюкозы на путь гликолиза. Вовлечение глюкозных остатков в
процесс гликолиза обеспечивает важная реакция и эта реакция
контролируется регуляторным ферментом. Реакция катализируется
гексокиназой и активность этого фермента ингибируется глюкозо-6фосфатом.
Когда концентрация глюкозо-6-фосфата в клетке сильно возрастает,
т.е. когда он образуется быстрее, чем потребляется, наступает
ингибирование – гексокиназа под действием глюкозо-6-фосфата
выключается и дальнейшего фосфорилирования глюкозы не
происходит до тех пор, пока избыток глюкозо-6-фосфата не будет
использован.
39.
В печени преобладает другой фермент – глюкокиназа,которая не ингибируется глюкозо-6-фосфатом. Поэтому в печени,
способной хранить большие количества гликогена, избыточная
глюкоза крови может фосфорилироваться в глюкозо-6-фосфат,
который через глюкозо-1-фосфат превращается в гликоген.
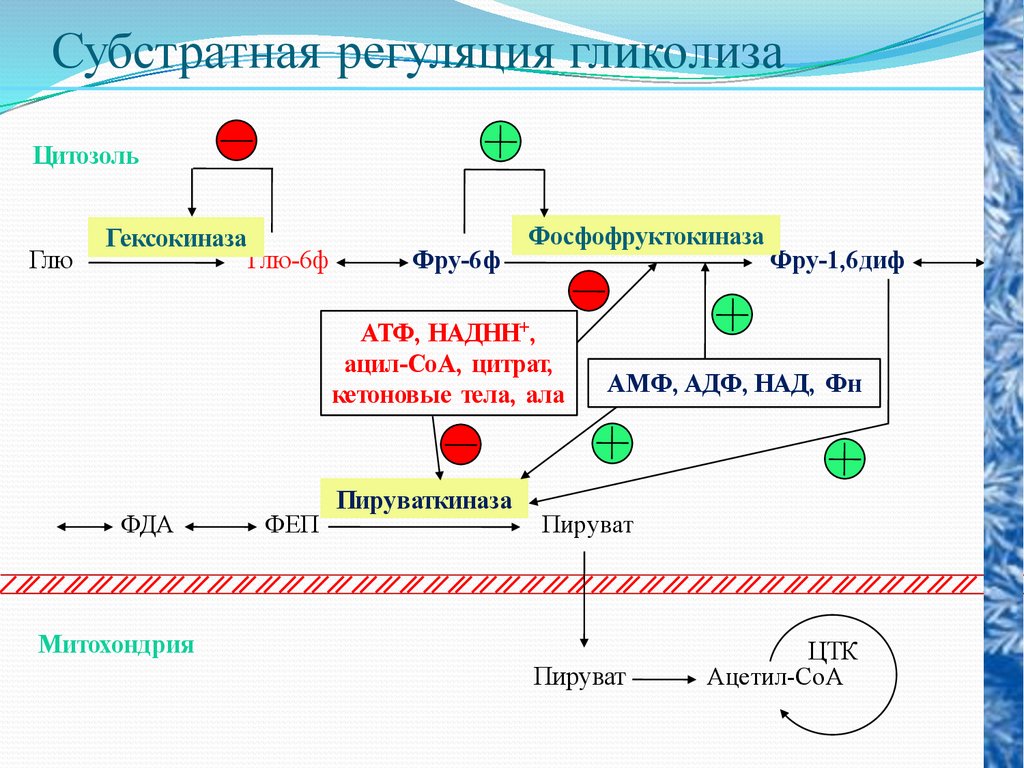
Кроме гексокиназной реакции в гликолизе имеются еще два
главных регулируемых этапа: это фосфофруктокиназная и
пируваткиназная реакция.
В
скелетных
мышцах
активность
фосфофруктокиназы
определяется концентрациями субстратов этого фермента (АТФ и
фруктозо-6-фосфата) и его продуктов (АДФ и фруктозо-1,6дифосфата), а также цитрат.
Главными отрицательными модуляторами фосфофруктокиназой
являются АТФ и цитрат.
Главными положительными модуляторами являются АМФ и
фруктозо-1,6-дифосфат.
40.
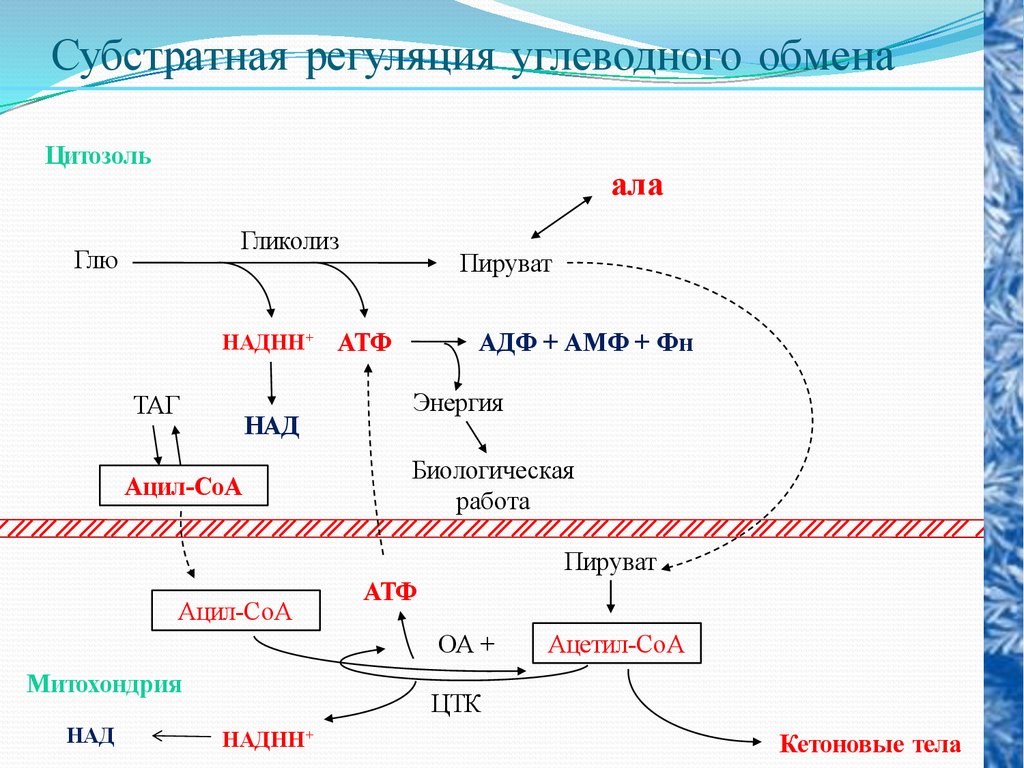
Субстратная регуляция углеводного обменаЦитозоль
ала
Гликолиз
Глю
НАДНН+
ТАГ
НАД
Ацил-СоА
Пируват
АТФ
АДФ + АМФ + Фн
Энергия
Биологическая
работа
Пируват
Ацил-СоА
АТФ
ОА +
Митохондрия
НАД
Ацетил-СоА
ЦТК
НАДНН+
Кетоновые тела
41.
Субстратная регуляция гликолизаЦитозоль
Глю
Гексокиназа
Глю-6ф
Фру-6ф
Фосфофруктокиназа
АТФ, НАДНН+,
ацил-СоА, цитрат,
кетоновые тела, ала
ФДА
ФЕП
Пируваткиназа
Фру-1,6диф
АМФ, АДФ, НАД, Фн
Пируват
Митохондрия
Пируват
ЦТК
Ацетил-СоА
42.
Регуляция гликолиза.Три
стадии
катаболизма
углеводов
обеспечивают получение энергии: гликолиз, цикл
лимонной кислоты и окислительноефосфорилирование.
При уменьшении содержания АТФ и Рн, это
приводит к возрастанию скорости переноса электронов и
окислительного фосфорилирования. Одновременно
повышается скорость окисления пирувата через цикл
лимонной кислоты, т.е. усиливается приток электронов в
дыхательную цепь. Это приводит к увеличению скорости
гликолиза, тем самым обеспечивается образование
пирувата. Затем наступает момент, когда отношение
АТФ/АДФ+Рн возвращается к обычному высокому
уровню. Теперь перенос электронов и окислительное
фосфорилирование замедляются. Цикл лимонной
кислоты замедляется, т.к. АТФ и цитрат являются
ингибитором фосфофруктокиназы и пируваткиназы.
43.
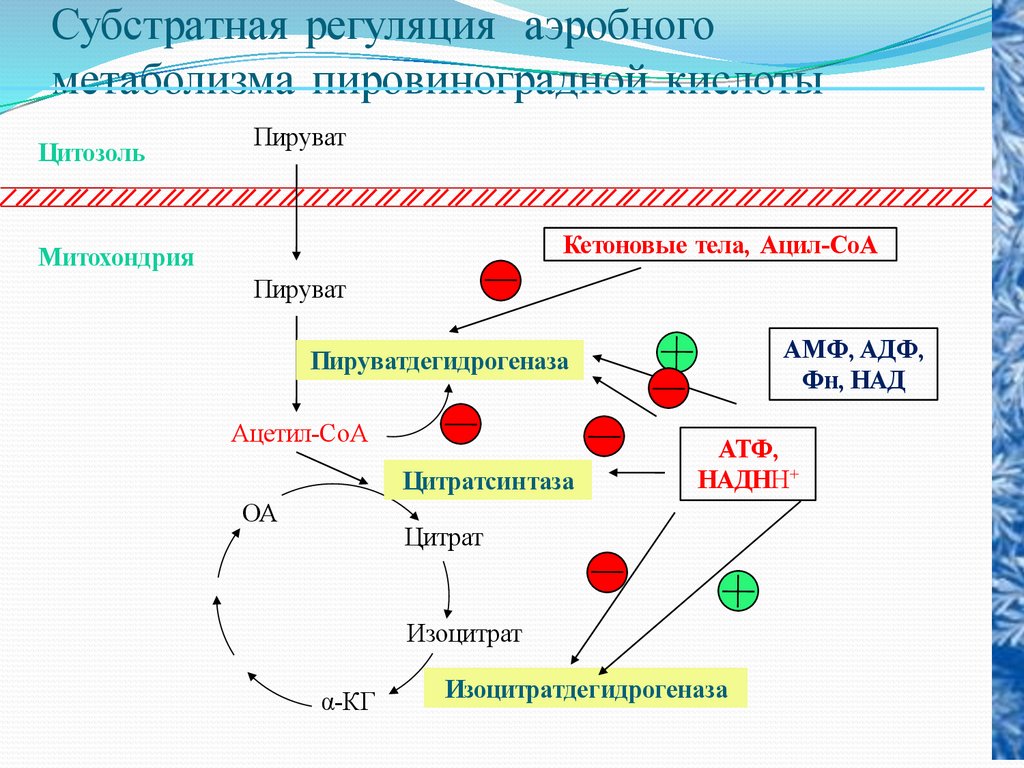
Субстратная регуляция аэробногометаболизма пировиноградной кислоты
Цитозоль
Пируват
Кетоновые тела, Ацил-СоА
Митохондрия
Пируват
АМФ, АДФ,
Фн, НАД
Пируватдегидрогеназа
Ацетил-СоА
Цитратсинтаза
ОА
АТФ,
НАДНН+
Цитрат
Изоцитрат
α-КГ
Изоцитратдегидрогеназа
44.
Регуляция пируватдегидрогеназного комплекса.Превращение пирувата в Ацетил-КоА происходит при
участии пируватдегидрогеназного комплекса, содержащего 5
коферментов: НАД, ФАД, НSКоА, ТДФ, липоевую кислоту. В основе
процессалежит реакция окислительного декарбоксилированя.
Образование ацетил-КоА из пирувата – это ключевой
необратимый этап метаболизма, потому что животные неспособны к
превращению ацетил-КоА в глюкозу.
Окислительной декарбоксилирование пирувата в ацетилКоА создает возможность превращения атомов глюкозы по 2 путям:
1) окисление до СО2 в ЦТК с одновременным регенерированием
энергии; 2) включение в липиды. Это позволяет считать, что
активность пируватдегидрогеназного комплекса должна строго
регулироваться.
45.
Активность ПДК (пируватдегидрогеназного комплекса)регулируется 3 путями:
1. Ингибирование продуктами реакции. Ацетил-КоА и
НАДН ингибируют превращение .
2. Регуляция нуклеотидами по принципу обратной
связи.
Пируватдегидрогеназный комплекс ингибируется ГТФ
и активируется АМФ.
3. Регуляция путем ковалентной модификации.
Комплекс теряет ферментативную активность, когда
специфический остаток серина
пируватдегидрогеназного комплекса фосфорилируется
АТФ. Фосфорилирование усиливается при высоких
соотношениях АТФ\АДФ, ацетил-КоА\СоА, НАДН\НАД+
и ингибируется пируватом.
46.
Регуляция ЦТК.Скорость функционирования ЦТК зависит от потребности клетки в
АТФ. Важной регуляторной реакцией цикла является синтез цитрата из
ацетил-КоА и оксалоацетата под действием цитратсинтетазы. АТФ –
аллостерический ингибитор цитратсинтетазы.
Вторая регуляторная реакция – это реакция, катализируемая изоцитратдегидрогеназой. Фермент аллостерически активируется АДФ, НАДН
ингибирует изоцитрат-дегидрогеназу.
Третьей регуляторной реакцией является реакция, катализируемая –
кетоглутаратдегидрогеназой.
Активация пируваткарбоксилазы.
Активность пируваткарбоксилазы зависит от присутствия
ацетил-КоА.
Пир может превращаться в ацетил-КоА, а также подвергаться
карбоксилированию с образование оксалоацетата или ЩУК. Высокое
содержание ацетил-КоА приводит к образованию количества
оксалоацетата. Если имеет место избыток АТФ, то оксалоацетат
потребляется в процессе глюконеогенеза.
47.
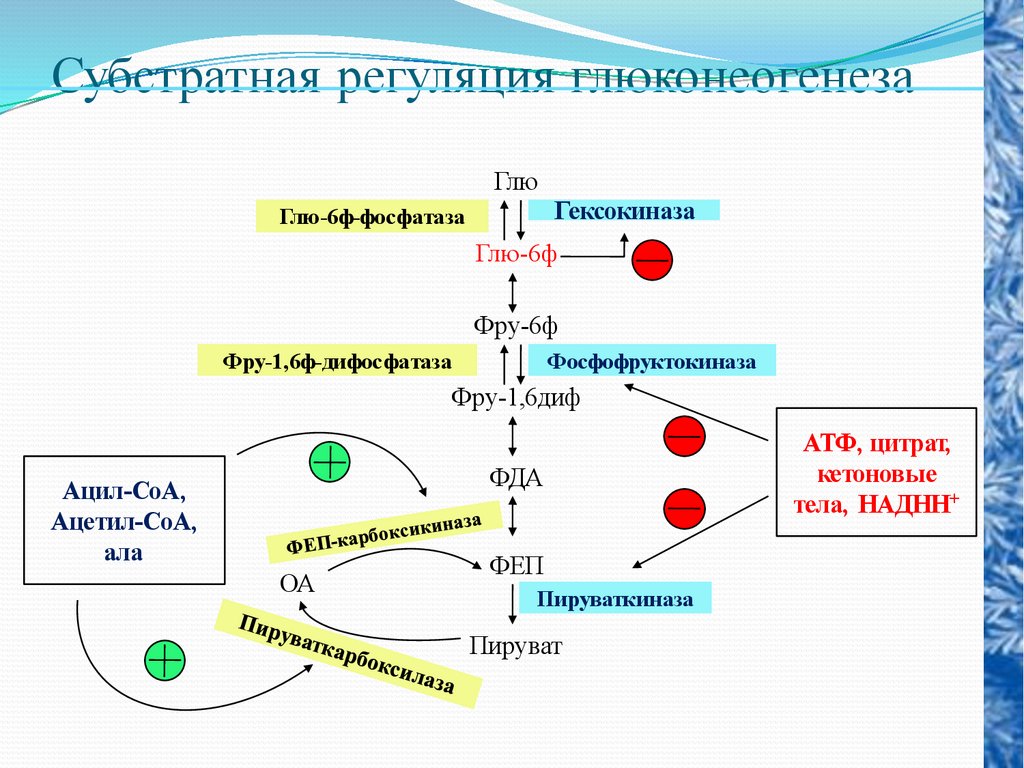
Субстратная регуляция глюконеогенезаГлю
Гексокиназа
Глю-6ф-фосфатаза
Глю-6ф
Фру-6ф
Фру-1,6ф-дифосфатаза
Фосфофруктокиназа
Фру-1,6диф
ФДА
Ацил-СоА,
Ацетил-СоА,
ала
ОА
ФЕП
Пируваткиназа
Пируват
АТФ, цитрат,
кетоновые
тела, НАДНН+
48.
Регуляция глюконеогенеза осуществляется восновном через реакции синтеза фосфоенолпирувата
и
глюкозо-6-фосфата.
Для
первой
реакции,катализируемой пируваткарбоксилазой, ал
лостерическим
модулятором
этого
фермента
является ацетил- СоА, при отсутствии которого
фермент почти полностью лишен активности.
Когда
в
клетке
накапливается
митохондриальный ацетил-СоА, биосинтез глюкозы
из пирувата усиливается, одновременно накопление
ацетил-СоА
замедляет
окислительное
декарбоксилирование
пирувата,
что
также
способствует превращению последнего в глюкозу.
49.
Вторая реакция катализируется фруктозо1,6-дифосфатазой, которая ингибируетсяАМФ и активируется АТФ. При низкой
концентрации АМФ и высоком уровне АТФ
происходит стимуляция глюконеогенеза. И
наоборот, когда отношение концентрации
АТФ к АМФ мало, в клетке происходит
расщепление глюкозы. Таким образом,
количество глюкозы в крови, оттекающей от
печени, зависит в основном от двух
взаимосвязанных процессов: гликолиза и
глюконеогенеза, которые в свою очередь
регулируются ключевыми ферментами.
50.
Судьба глюкозо-6-фосфата — вступит ли он вгликолиз
или
пентозофосфатный
путь
—
определяется потребностями клетки в данный
момент, а также концентрацией NADP+ в цитозоле.
Без наличия акцептора электронов первая реакция
пентозофосфатного пути (катализируемая глюкозо6-фосфатдегидрогеназой) не будет идти. Когда
клетка быстро переводит NADPH в NADP+ в
биосинтетических восстановительных реакциях,
уровень NADP+ поднимается, аллостерически
стимулируя глюкозо-6-фосфатдегидрогензазу и тем
самым увеличивая ток глюкозо-6-фосфата через
пентозофосфатный путь. Когда потребление NADPH
замедляется, уровень NADP+ снижается, и глюкозо6-фосфат утилизируется гликолитически
51.
Регуляция осуществляется на уровне двухферментов: гликогенсинтетазы,
участвующей в синтезе гликогена, и
фосфорилазы, катализирующей
расщепление гликогена. Основным
механизмом регуляции активности этих
ферментов является их ковалентная
модификация путем фосфорилированиядефосфорилирования.
52.
Распад гликогена в мышцах инициируетадреналин, который связывается со своим
рецептором и активирует аденилатциклазу.
Аденилатциклаза начинает синтезировать
циклический АМФ. Циклический АМФ
запускает целый каскад реакций, которые в
конечном итоге приводят к активации
фосфорилазы. Гликогенфосфорилаза
катализирует распад гликогена. В печени
распад гликогена стимулируется глюкагоном.
53.
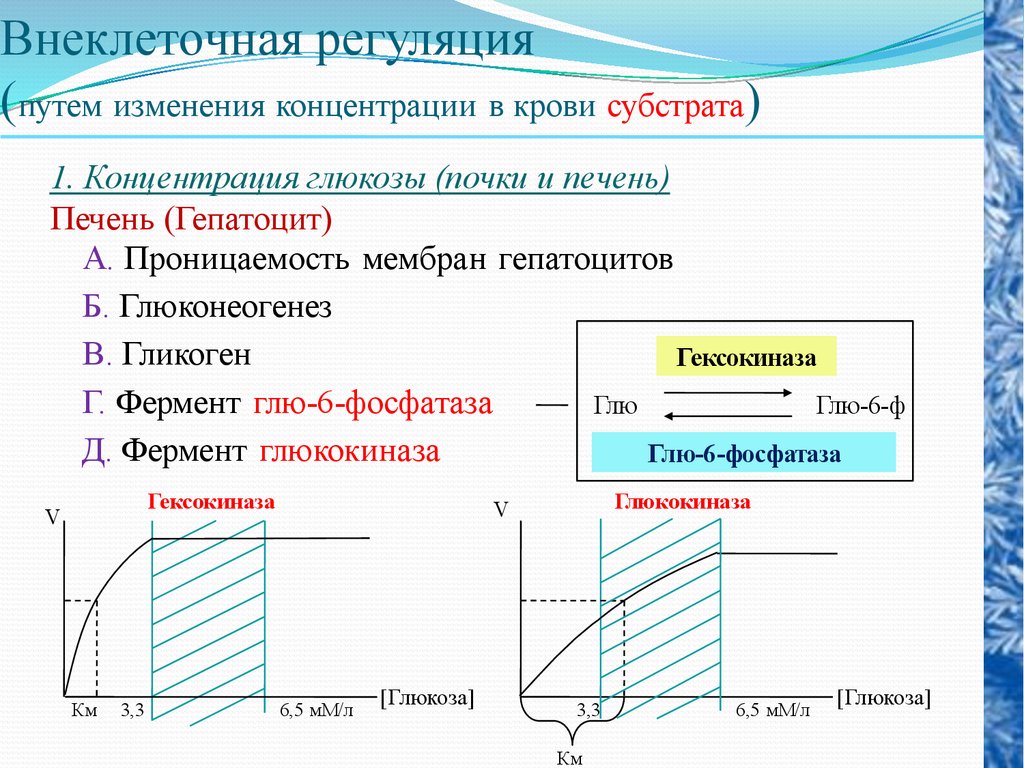
Внеклеточная регуляция(путем изменения концентрации в крови субстрата)
1. Концентрация глюкозы (почки и печень)
Печень (Гепатоцит)
А. Проницаемость мембран гепатоцитов
Б. Глюконеогенез
В. Гликоген
Гексокиназа
Г. Фермент глю-6-фосфатаза
Глю
Глю-6-ф
Д. Фермент глюкокиназа
Глю-6-фосфатаза
Гексокиназа
V
Км
3,3
Глюкокиназа
V
6,5 мМ/л
[Глюкоза]
3,3
Км
6,5 мМ/л
[Глюкоза]
54.
Одним из интегральных показателей внутренней среды,отражающим обмен в организме углеводов, белков и жиров,
является концентрация в крови глюкозы. Она является не только
источником энергии для синтеза жиров и белков, но и субстратом
для их синтеза. В печени происходит новообразование углеводов
из жирных кислот и аминокислот. Нормальное функционирование
клеток нервной системы, поперечнополосатых и гладких мышц,
для которых глюкоза является важнейшим энергосубстратом,
возможно при условии, что приток к ним глюкозы обеспечит их
энергетические потребности. Это достигается при содержании в
литре крови у человека в среднем 1 г (0,8—1,2 г) глюкозы. При
нормальном уровне содержания глюкозы в крови происходит
образование гликогена в печени и мышцах, синтез жиров, ее
потребление клетками мозга, мышцами и другими тканями. В
условиях гипергликемии избыточное количество глюкозы
удаляется из крови через почки, увеличивается синтез гликогена.
При гипогликемии усиливается гликогенолиз под влиянием
адреналина и глюкагона.
55.
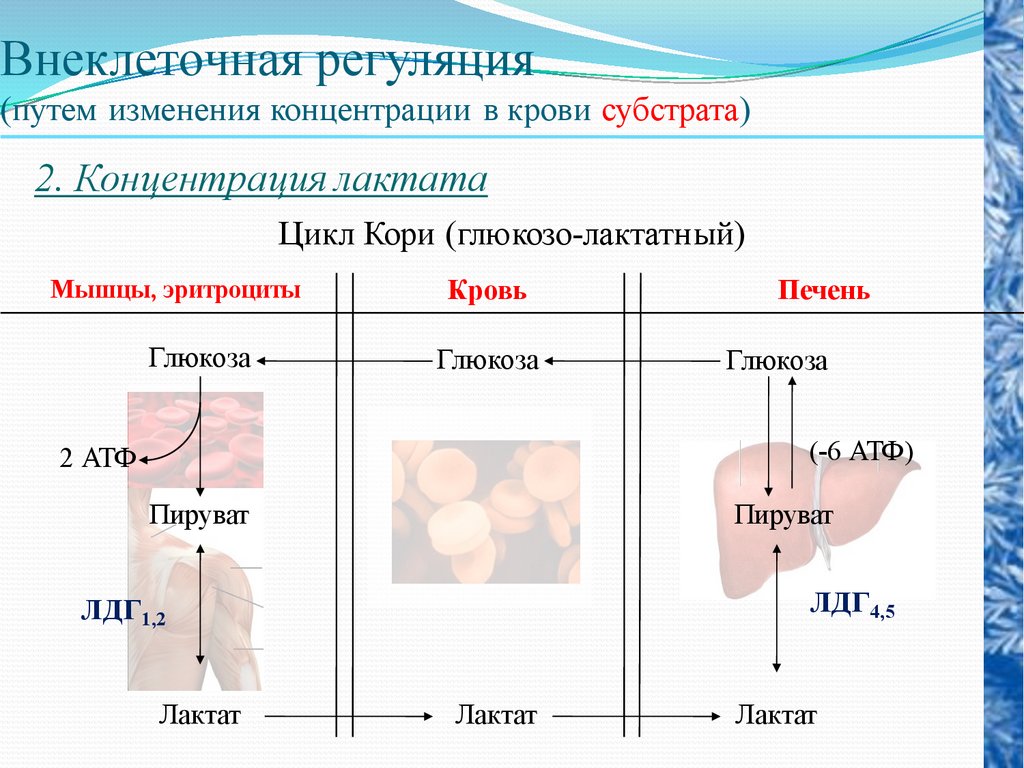
Внеклеточная регуляция(путем изменения концентрации в крови субстрата)
2. Концентрация лактата
Цикл Кори (глюкозо-лактатный)
Мышцы, эритроциты
Глюкоза
Кровь
Глюкоза
Печень
Глюкоза
(-6 АТФ)
2 АТФ
Пируват
Пируват
ЛДГ4,5
ЛДГ1,2
Лактат
Лактат
Лактат
56.
Глюкозо-лактатный цикл не требуетналичие кислорода, функционирует
всегда, обеспечивает: 1) утилизацию
лактата, образующегося в анаэробных
условиях (скелетные мышцы,
эритроциты), что предотвращает
лактоацидоз; 2) синтез глюкозы
(печень).
57.
Внеклеточная регуляция(путем изменения концентрации в крови субстрата)
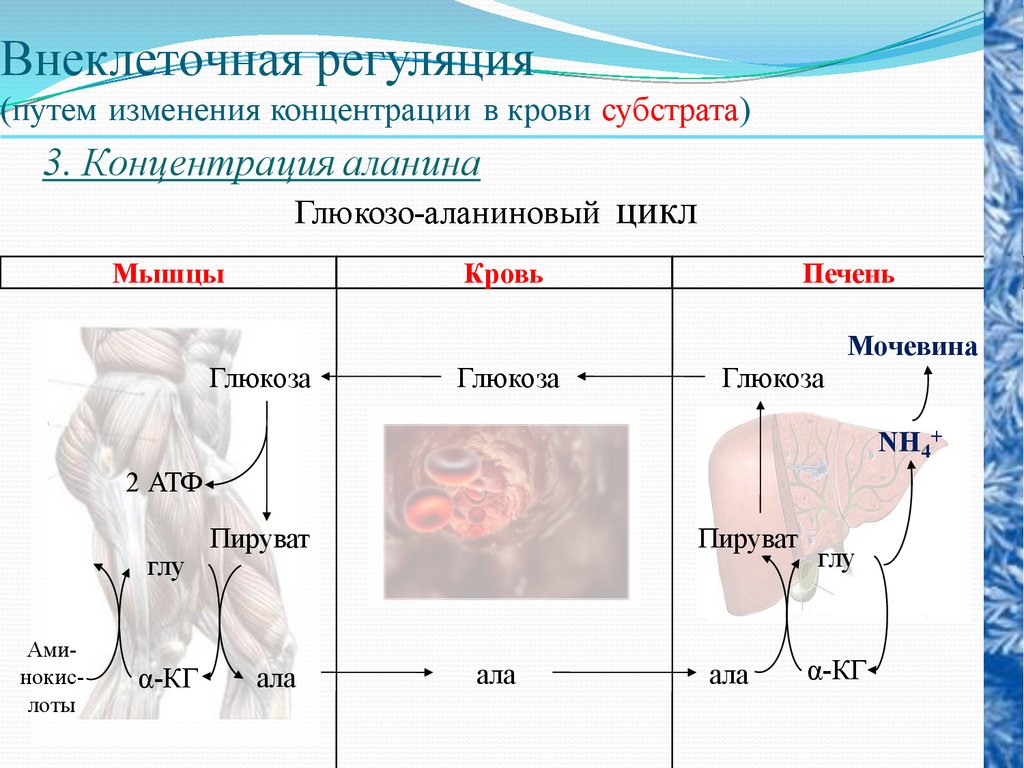
3. Концентрация аланина
Глюкозо-аланиновый цикл
Мышцы
Кровь
Печень
Мочевина
Глюкоза
Глюкоза
Глюкоза
NH4+
2 АТФ
глу
Аминокислоты
α-КГ
Пируват
ала
Пируват
ала
ала
глу
α-КГ
58.
Формируется глюкозо-аланиновый цикл, вкотором аланин мышц переносится в печень
и используется в процессе печеночного
глюконеогенеза и в то же время доставляет в
печень аминогруппы, удаляемые в виде
мочевины. Углеродный скелет аминокислот,
подвергшихся деградации и включившихся
в цикл трикарбоновых кислот в мышечной
ткани, превращается главным образом в
глутамин
и
пируват.
Глюкозоаланиновый цикл функционирует в
мышцах при голодании.
59.
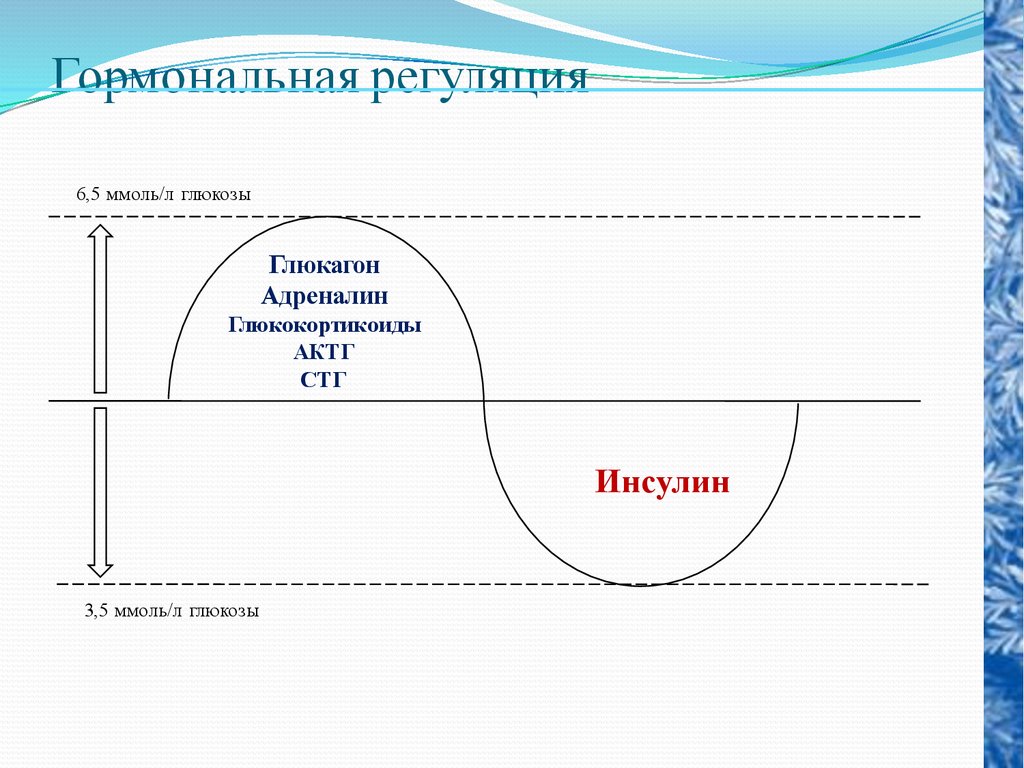
Гормональная регуляция6,5 ммоль/л глюкозы
Глюкагон
Адреналин
Глюкокортикоиды
АКТГ
СТГ
Инсулин
3,5 ммоль/л глюкозы
60.
Инсулин - гормон поджелудочной железы, обеспечиваетснижение сахара в крови путем активации ферментов,
ответственных за использование глюкозы клетками
организма.
Глюкоза крови при участии инсулина поступает в клетки
организма, в результате чего уровень ее в крови
снижается (гипогликемический эффект). В клетках
глюкоза превращается в глюкозо-6-фосфорный эфир (Г6-Ф), который подвергается распаду или по пути
гликолиза, или при аэробных условиях (пентозный
цикл). При гликолизе из промежуточных продуктов
может быть образован глицерин и небольшое количество
ацетил-КоА, который поступает в цикл Кребса. В
пентозном цикле глюкоза полностью окисляется с
выделением большого количества СО2 (из одной
молекулы глюкозы образуется 6 молекул СО2) и ряда
промежуточных соединений, из которых могут быть
синтезированы жирные кислоты.
61.
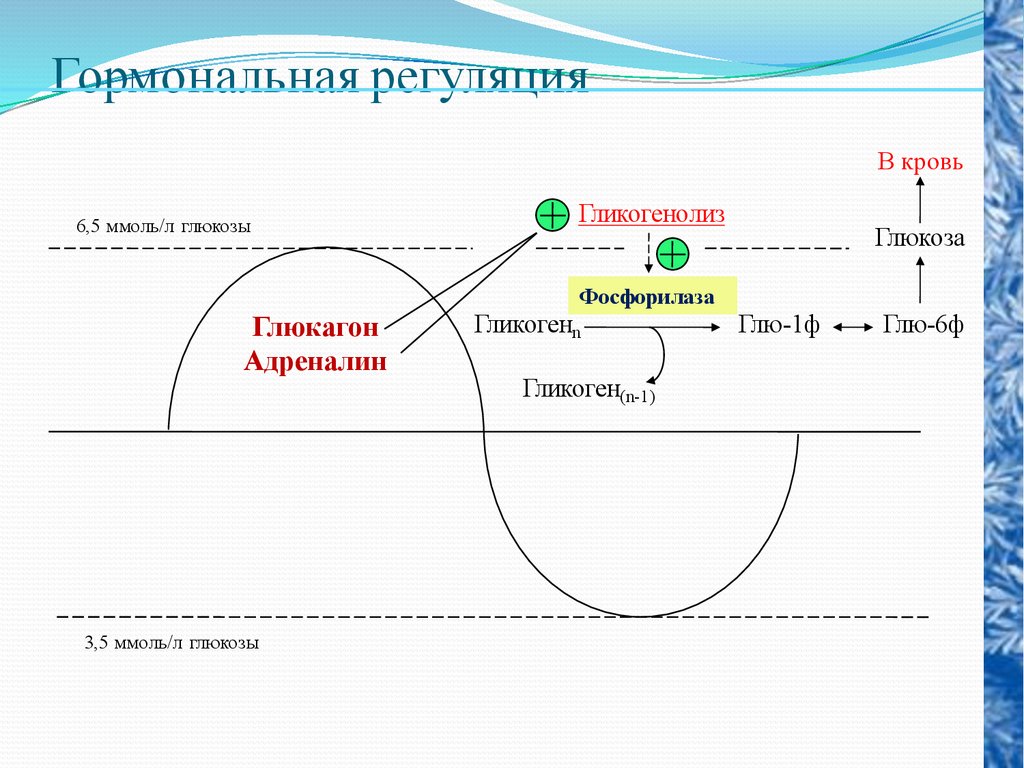
Из других гормонов, участвующих в регуляции уровня сахара вкрови, интерес представляет адреналин - гормон мозгового
слоя надпочечников. Адреналин повышает уровень сахара
путем активации распада гликогена (фермента фосфорилазы)
до глюкозы и выброса ее в кровь. Помимо этого, адреналин
умеренно активирует гликолиз. При этом происходит
образование
большего
количества
ацетил-КоА
и,
соответственно, большего количества энергии.
Глюкагон - гормон поджелудочной железы, действует
аналогично адреналину.
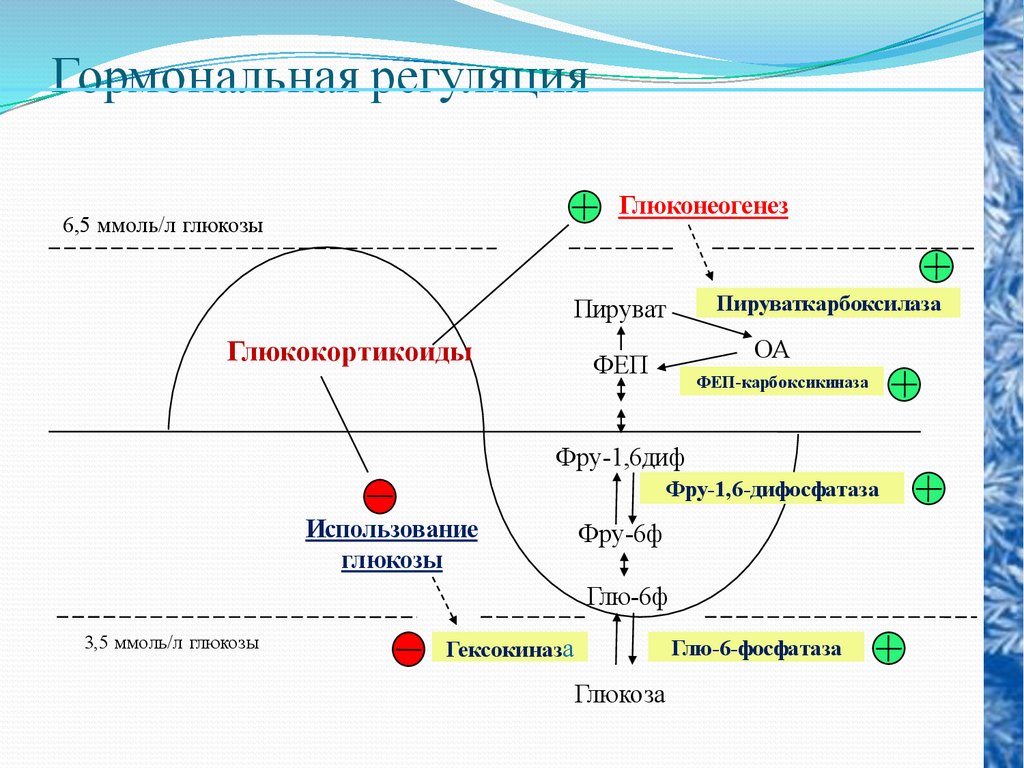
Глюкокортикоиды
гормоны
коры надпочечников,
активируют процесс превращения жиров и белков в углеводы глюконеогенез.
Адренокортикотропный гормон (АКТГ), вырабатываемый в
гипофизе, стимулирует выработку глюкокортикоидов, т. е.
опосредованно способствует повышению уровня сахара в крови
путем активации глюконеогенеза. Аналогичным путем
повышает сахар крови и соматотропный гормон.
62.
Только инсулин способствует снижению уровнясахара в крови, тогда как остальные гормоны
вызывают его повышение.
Адреналин и другие гормоны обеспечивают
распад запасной формы углеводов - гликогена - до
глюкозы и поступление ее в кровь.
Инсулин же способствует использованию этой
глюкозы клетками организма.
Из других регуляторных механизмов необходимо
выделить печень, в клетках которой происходят
процессы распада и синтеза гликогена. Поэтому
кровь, протекающая через печень, или насыщается
глюкозой при ее недостатке в крови, или уровень
сахара в крови снижается при ее избытке.
63.
Гормональная регуляцияВ кровь
6,5 ммоль/л глюкозы
Гликогенолиз
Глюкоза
Фосфорилаза
Глюкагон
Адреналин
3,5 ммоль/л глюкозы
Гликогенn
Гликоген(n-1)
Глю-1ф
Глю-6ф
64.
Гормональная регуляцияГлюконеогенез
6,5 ммоль/л глюкозы
Пируваткарбоксилаза
Пируват
Глюкокортикоиды
ОА
ФЕП
ФЕП-карбоксикиназа
Фру-1,6диф
Фру-1,6-дифосфатаза
Использование
глюкозы
Фру-6ф
Глю-6ф
3,5 ммоль/л глюкозы
Гексокиназа
Глю-6-фосфатаза
Глюкоза
65.
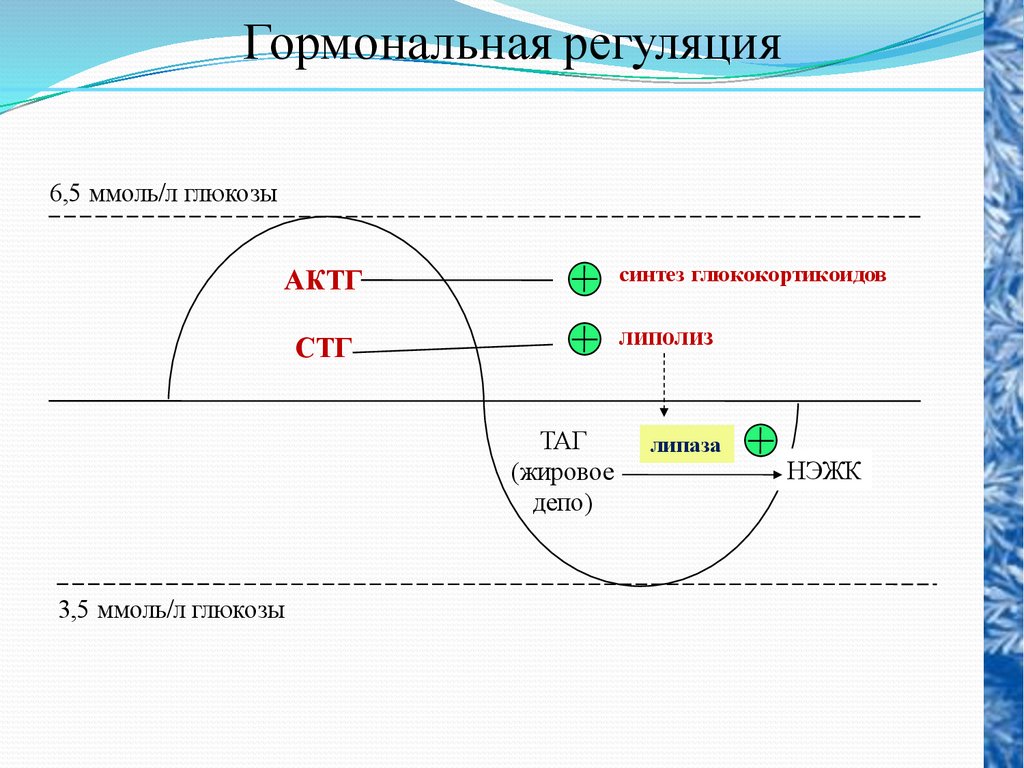
Гормональная регуляция6,5 ммоль/л глюкозы
синтез глюкокортикоидов
АКТГ
липолиз
СТГ
ТАГ
(жировое
депо)
3,5 ммоль/л глюкозы
липаза
НЭЖК
66.
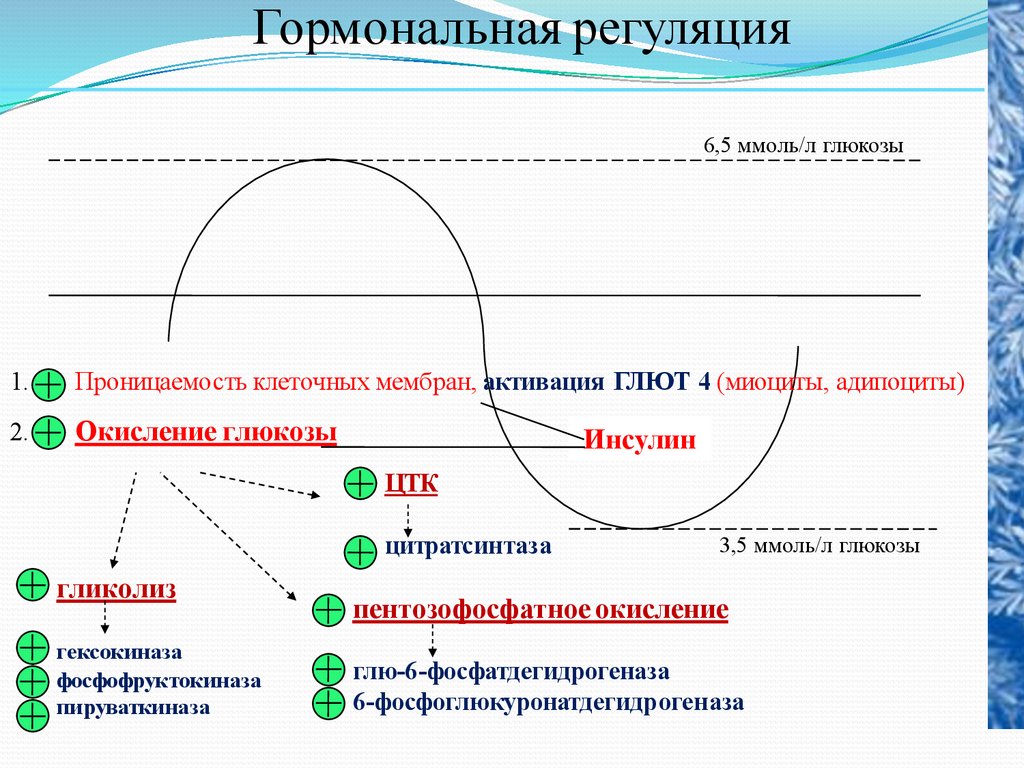
Гормональная регуляция6,5 ммоль/л глюкозы
1.
Проницаемость клеточных мембран, активация ГЛЮТ 4 (миоциты, адипоциты)
2.
Окисление глюкозы
Инсулин
ЦТК
цитратсинтаза
гликолиз
гексокиназа
фосфофруктокиназа
пируваткиназа
3,5 ммоль/л глюкозы
пентозофосфатное окисление
глю-6-фосфатдегидрогеназа
6-фосфоглюкуронатдегидрогеназа
67.
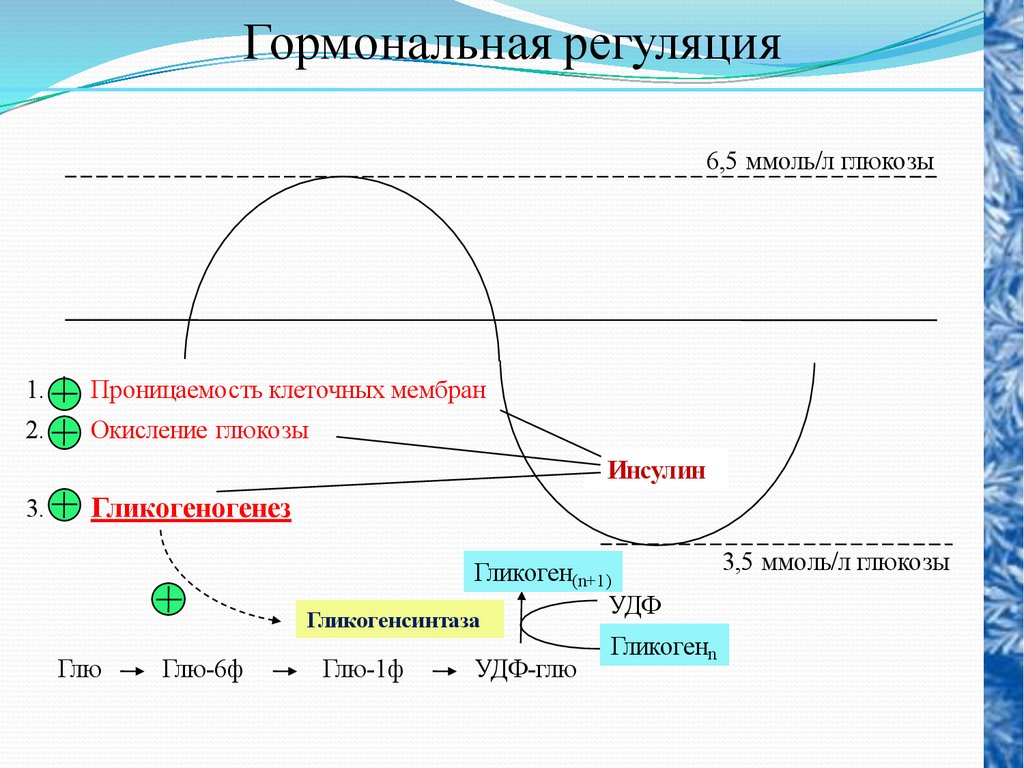
Гормональная регуляция6,5 ммоль/л глюкозы
1.
Проницаемость клеточных мембран
2.
Окисление глюкозы
Инсулин
3.
Гликогеногенез
Гликоген(n+1)
УДФ
Гликогенсинтаза
Глю
Глю-6ф
Глю-1ф
УДФ-глю
Гликогенn
3,5 ммоль/л глюкозы
68.
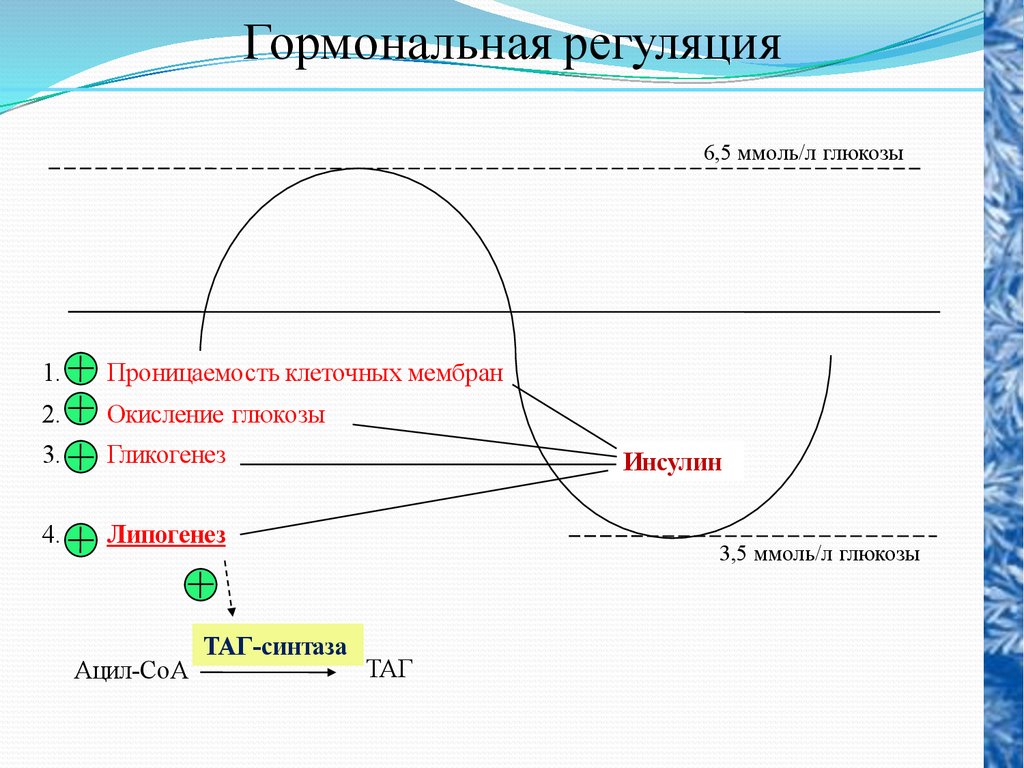
Гормональная регуляция6,5 ммоль/л глюкозы
1.
Проницаемость клеточных мембран
2.
Окисление глюкозы
3.
Гликогенез
4.
Липогенез
Ацил-СоА
ТАГ-синтаза
Инсулин
3,5 ммоль/л глюкозы
ТАГ
69.
Гормональная регуляция6,5 ммоль/л глюкозы
Липолиз
Инсулин
ТАГ
Липаза
Ацил-СоА
Гликогенn
Гликогенолиз
Фосфорилаза
3,5 ммоль/л глюкозы
Гликоген(n-1) + Глю-1ф
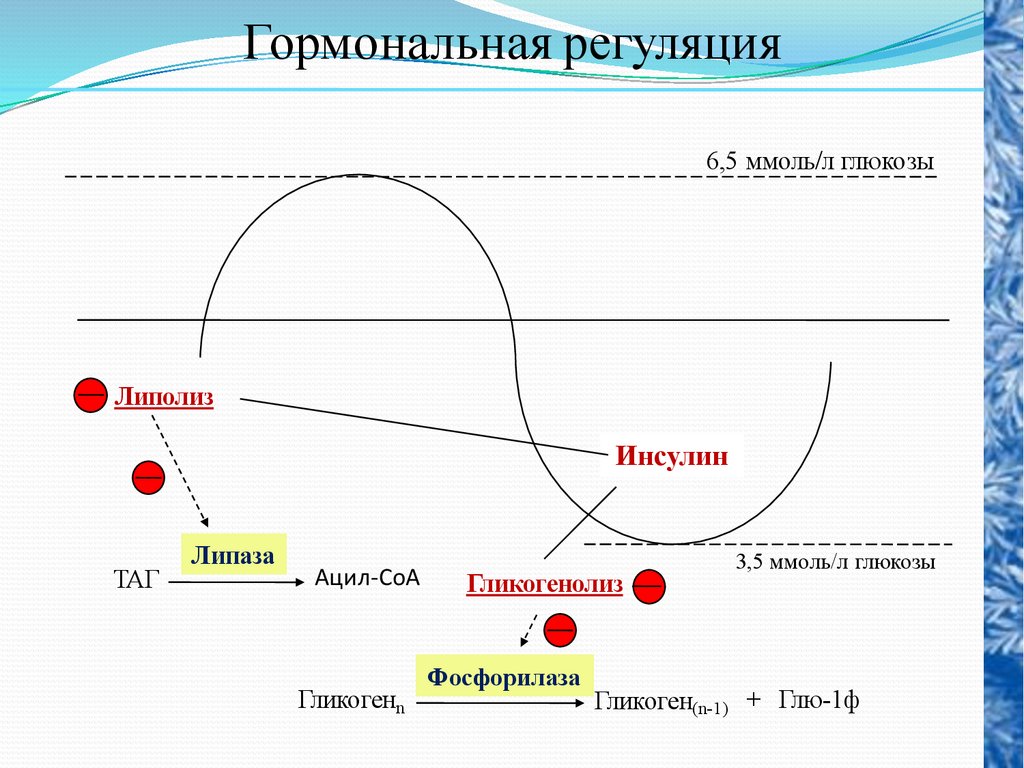
70.
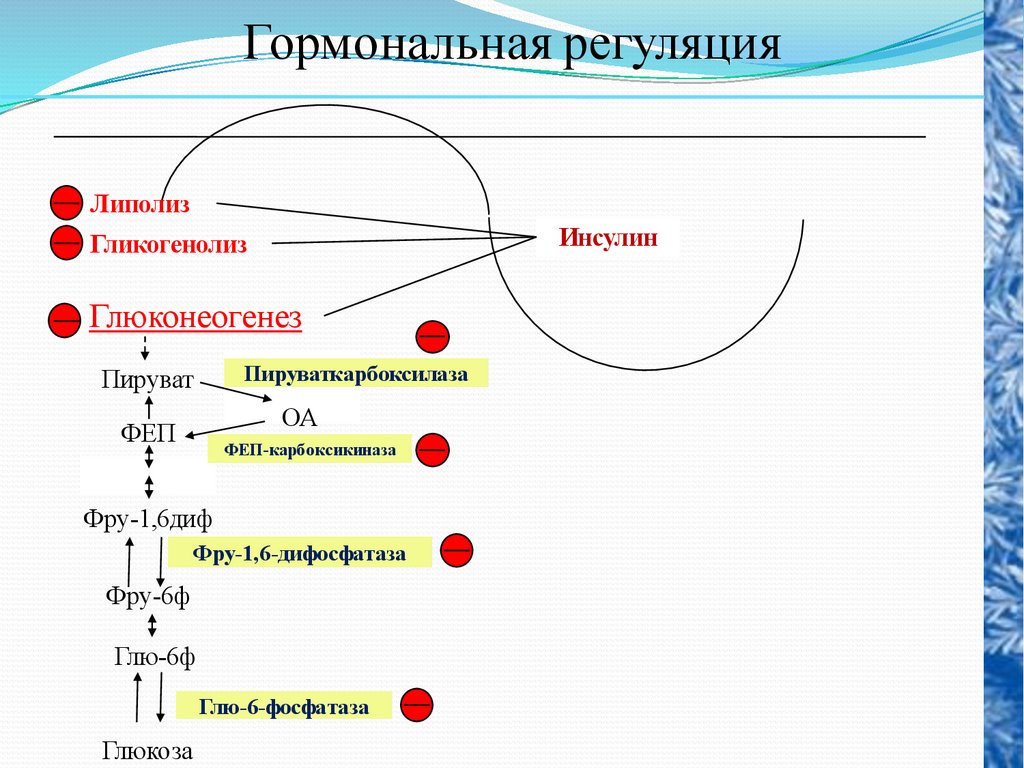
Гормональная регуляцияЛиполиз
Инсулин
Гликогенолиз
Глюконеогенез
Пируваткарбоксилаза
Пируват
ОА
ФЕП
ФЕП-карбоксикиназа
Фру-1,6диф
Фру-1,6-дифосфатаза
Фру-6ф
Глю-6ф
Глю-6-фосфатаза
Глюкоза
71.
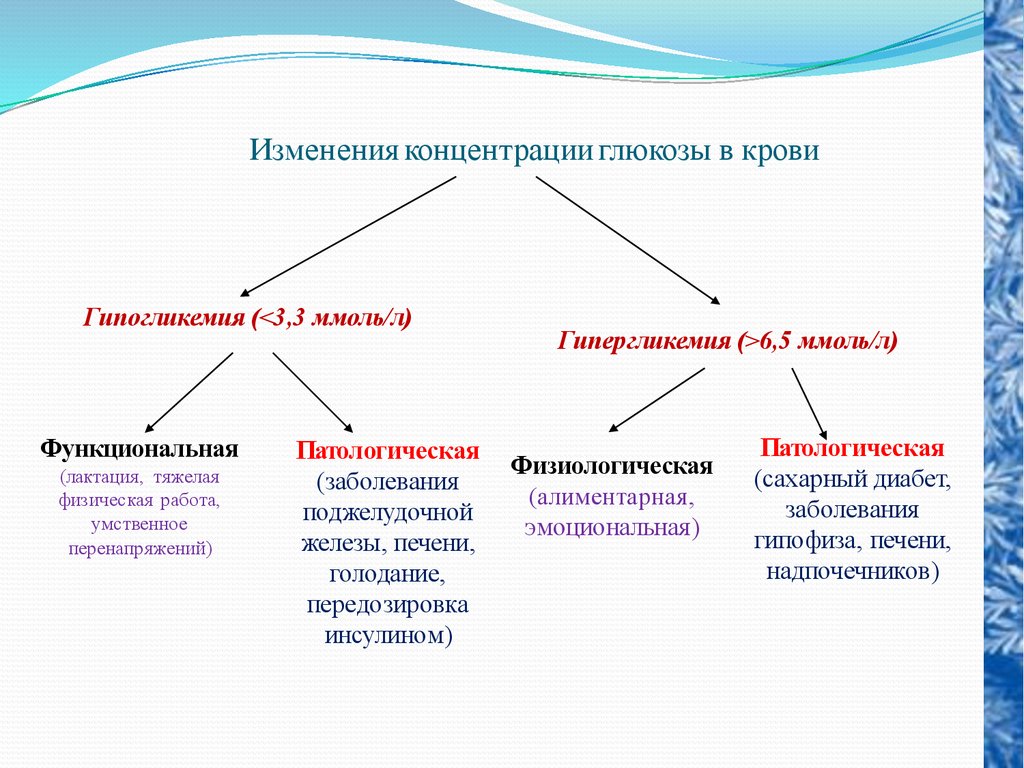
Изменения концентрации глюкозы в кровиГипогликемия (<3,3 ммоль/л)
Функциональная
(лактация, тяжелая
физическая работа,
умственное
перенапряжений)
Патологическая
(заболевания
поджелудочной
железы, печени,
голодание,
передозировка
инсулином)
Гипергликемия (>6,5 ммоль/л)
Физиологическая
(алиментарная,
эмоциональная)
Патологическая
(сахарный диабет,
заболевания
гипофиза, печени,
надпочечников)
72.
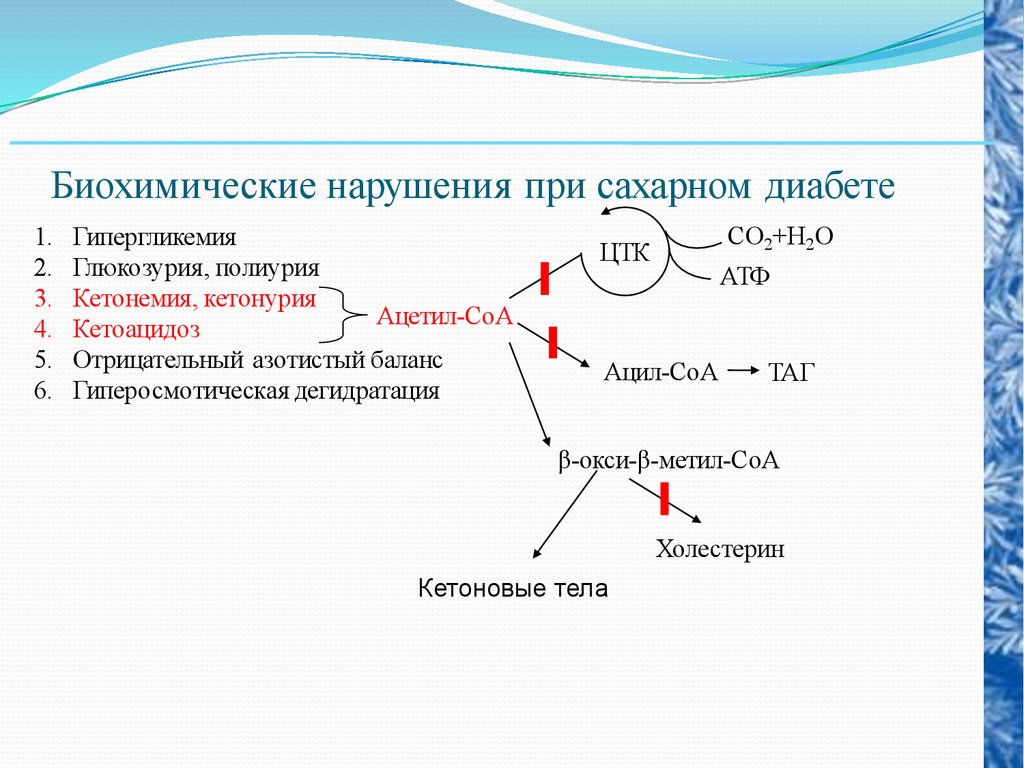
Биохимические нарушения при сахарном диабете1.
2.
3.
4.
5.
6.
Гипергликемия
Глюкозурия, полиурия
Кетонемия, кетонурия
Ацетил-СоА
Кетоацидоз
Отрицательный азотистый баланс
Гиперосмотическая дегидратация
СО2+Н2О
ЦТК
АТФ
Ацил-СоА
ТАГ
β-окси-β-метил-СоА
Холестерин
Кетоновые тела
73.
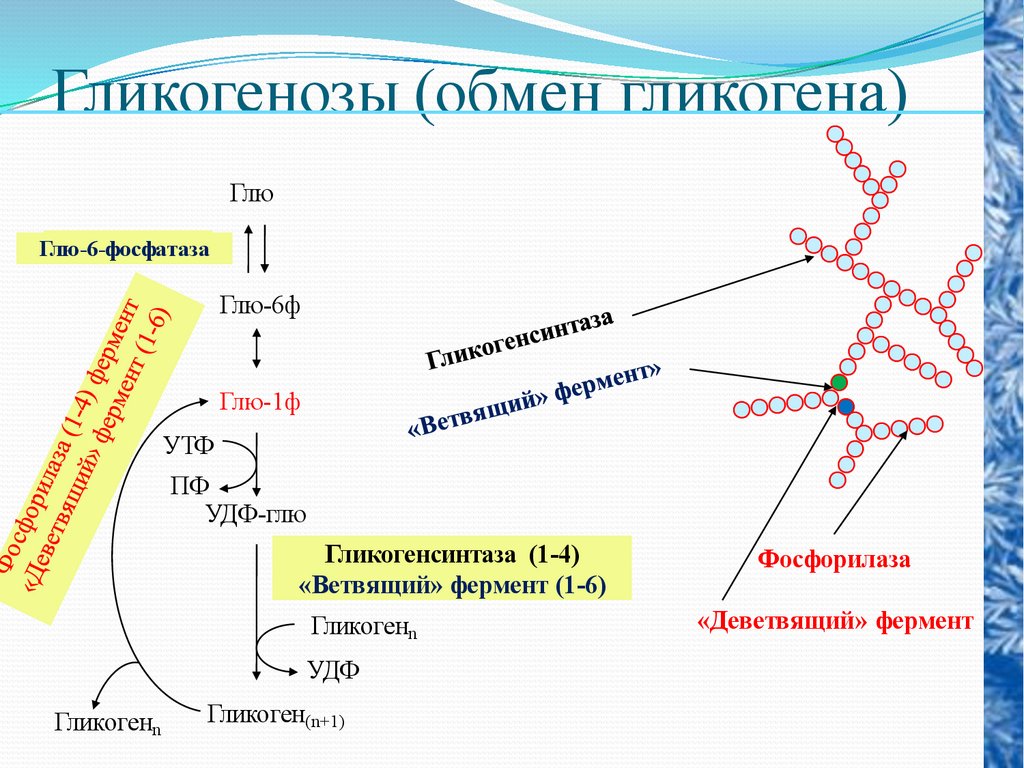
Гликогенозы (обмен гликогена)Глю
Глю-6-фосфатаза
Глю-6ф
Глю-1ф
УТФ
ПФ
УДФ-глю
Гликогенсинтаза (1-4)
«Ветвящий» фермент (1-6)
Гликогенn
УДФ
Гликогенn
Гликоген(n+1)
Фосфорилаза
«Деветвящий» фермент
74.
Гликогенозы — группа наследственныхзаболеваний, вызванных недостаточностью
одного
или
нескольких
ферментов,
вовлечённых в синтез и распад гликогена, и
характеризующихся
накоплением
патологических количеств или типов
гликогена
в
тканях.
Симптоматика
возникает
вследствие
накопления
гликогена,
или
его
промежуточных
метаболитов,
или
из-за
недостатка
конечных продуктов распада гликогена,
особенно глюкозы.
75.
Всего известно 12 типов гликогенозов.По
патогенетическому
признаку
гликогенозы делят:
печеночные – 0, I, III, IV, VI, VIII, IX, Х,
ХI типов,
мышечные – V и VII типов,
смешанные – II типа.
76.
Печеночные гликогенозыСамый частый гликогеноз I типа или болезнь фон
Гирке
обусловлен
аутосомно-рецессивным
дефектом глюкозо-6-фосфатазы. Из-за того, что этот
фермент есть только в печени и почках, преимущественно
страдают эти органы, и болезнь носит еще одно название
– гепаторенальный гликогеноз. Даже у новорожденных
детей наблюдаются гепатомегалия и нефромегалия,
обусловленные накоплением гликогена не только в
цитоплазме, но и в ядрах клеток. Кроме этого,
активируется синтез липидов с возникновением стеатоза
печени.
Так
как
фермент
необходим
для
дефосфорилирования глюкозо-6-фосфата с последующим
выходом глюкозы в кровь, у больных отмечается
гипогликемия
и,
как
следствие,
ацетонемия,
метаболический ацидоз, ацетонурия.
77.
Гликогеноз III типа или болезнь ФорбсаКори – это аутосомно-рецессивныйдефект
амило-α1,6-глюкозидазы,
"деветвящего" фермента, гидролизующего
α1,6-гликозидную связь. Болезнь имеет
более доброкачественное течение, и
частота ее составляет примерно 25% от всех
гликогенозов. Для больных характерна
гепатомегалия,
умеренная
задержка
физического развития, в подростковом
возрасте возможна небольшая миопатия.
78.
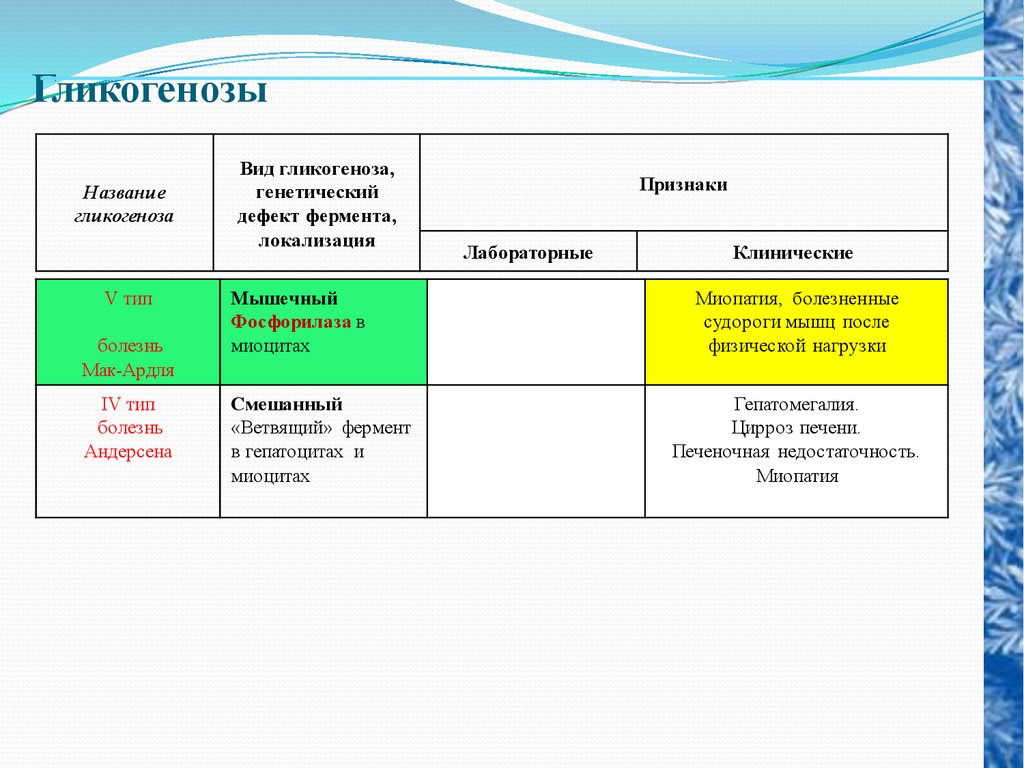
При гликогенозе IV типа (болезнь Андерсена,1%
всех
гликогенозов),
связанного
с
дефектом ветвящего фермента, образуется
гликоген с малым количеством ветвлений, что
резко уменьшает скорость гликогенолиза.
Гликогеноз VI типа (болезнь Херса, 25% всех
гликогенозов), связан с дефицитом печеночной
фосфорилазы гликогена. При этом отсутствует
мобилизация
гликогена,
развивается
гепатомегалия и гипогликемия.
79.
Мышечные гликогенозыДля этой группы гликогенозов характерны
изменения ферментов мышечной ткани. Это
приводит к нарушению энергообеспечения
мышц при физической нагрузке, к болям в
мышцах, судорогам.
Гликогеноз V типа (болезнь Мак-Ардля) –
отсутствие мышечной фосфорилазы. При
тяжелой мышечной нагрузке возникают
судороги, миоглобинурия, хотя легкая
работа не вызывает каких-либо проблем.
80.
Смешанные гликогенозыЭти заболевания касаются и печени, и мышц, и
других органов.
Гликогеноз II типа (болезнь Помпе, 10% всех
гликогенозов) – поражаются все гликогенсодержащие
клетки из-за отсутствия лизосомальной (кислой) α1,4-глюкозидазы, поэтому данная болезнь относится
к лизосомным болезням накопления. Происходит
накопление гликогена в лизосомах и в цитоплазме.
Заболевание составляет почти 10% всех гликогенозов
и является наиболее злокачественным. Больные при
отсутствии лечения умирают в раннем возрасте из-за
кардиомегалии
и
тяжелой
сердечной
недостаточности.
81.
АгликогенозыАгликогенозы – состояния, связанные с
отсутствием гликогена. В качестве примера
агликогеноза можно привести наследственный
аутосомно-рецессивный дефицит гликогенсинтазы. Симптомами является резкая
гипогликемия натощак, особенно утром,
появляется рвота, судороги, потеря сознания.
В результате гипогликемии наблюдается
задержка
психомоторного
развития,
умственная отсталость. Болезнь несмертельна
при адекватном лечении (частое кормление),
хотя и опасна.
82.
ГликогенозыНазвание
гликогеноза
I тип
Вид гликогеноза,
генетический
дефект фермента,
локализация
Печеночный.
болезнь Гирке
Глю-6-фосфатаза в
гепатоцитах
VI тип,
Печеночный.
Гликогенфосфорилаза в гепатоцитах
болезнь Херса
Признаки
Лабораторные
Клинические
Увеличение содержания:
в гепатоцитах гликогена, в крови
лактата,
кетоновых
тел
триглицеридов,
мочевой
к-ты.
Ацидоз. Гипогликемия натощак.
Гепатомегалия,
гипотрофия,
замедление роста,
кровоточивость,
судороги
Гипогликемия натощак, кетонемия,
ацидоз, увеличение содержания в
крови лактата, в печени - гликогена
Гепатомегалия,
слабовыраженное
замедление роста
83.
ГликогенозыНазвание
гликогеноза
V тип
болезнь
Мак-Ардля
IV тип
болезнь
Андерсена
Вид гликогеноза,
генетический
дефект фермента,
локализация
Мышечный
Фосфорилаза в
миоцитах
Смешанный
«Ветвящий» фермент
в гепатоцитах и
миоцитах
Признаки
Лабораторные
Клинические
Миопатия, болезненные
судороги мышц после
физической нагрузки
Гепатомегалия.
Цирроз печени.
Печеночная недостаточность.
Миопатия
84.
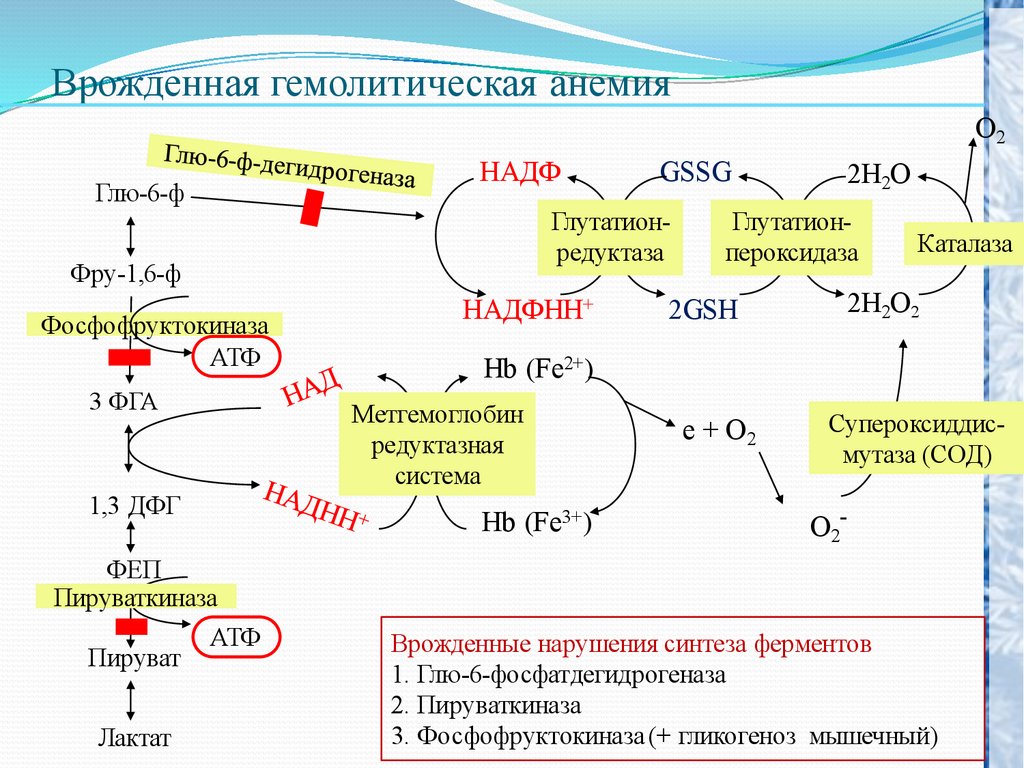
Врожденная гемолитическая анемияO2
НАДФ
Глю-6-ф
GSSG
Глутатионредуктаза
Фру-1,6-ф
Фосфофруктокиназа
АТФ
3 ФГА
НАДФНН+
Глутатионпероксидаза
2GSH
Каталаза
2Н2О2
Hb (Fe2+)
Метгемоглобин
редуктазная
система
1,3 ДФГ
2Н2О
Hb (Fe3+)
e + O2
Супероксиддисмутаза (СОД)
O2-
ФЕП
Пируваткиназа
Пируват
Лактат
АТФ
Врожденные нарушения синтеза ферментов
1. Глю-6-фосфатдегидрогеназа
2. Пируваткиназа
3. Фосфофруктокиназа (+ гликогеноз мышечный)
85.
Гликолиз в эритроцитах и транспорт кислорода связаны участиемв обоих процессах 2,3 –дифосфоглицерата.
2,3- дифосфоглицерат снижает сродство гемоглобина к кислороду и
облегчает освобождение кислорода в тканях.
1. Пи
дефекте
гексокиназы
снижается
концентрация
промежуточн6ых продуктов гликолиза, в том числе снижается
концентрация 2,3 –дифосфоглицерата. В таких эритроцитах
гемоглобин обладает осень высоким сродством с кислородом.
Наступает гемолиз эритроцитов, гемоглобин плохо отдает
кислород.
2. При дефекту пируваткиназы нарушается энергетический
обмен и мембрана не получает энергию необходимую для
ионного обмена наряду с этим образуется избыток 2,3 –
дифосфоглицерата и связь гемоглобина с кислородом
становится слабой, сродство гемоглобина к кислороду
становится низким.