



















")










 Биология
БиологияПохожие презентации:










Еволюція міжклітинного сигналінгу
1. Еволюція міжклітинного сигналінгу
2. Склад сигнальної системи
СигналЕ
Рецептор (R)
Р
Ефектор (E)
Вторинний месенджер
3. Загальні принципи еволюції сигналінгу
1. Відсутність ампліфікації: 1сигнал – 1 активований
ефектор
2. Ампліфікація сигналу
за рахунок 1) вторинних
месенджерів; 2)
фосфорилування; 3)
транскрипційної
регуляції.
3. Ампліфікація сигналу
додатково за
позитивним зворотнім
зв’язком
5. Ендогенна генерація
сигналів та їх регуляція
за негативним зв’язком
– часові осциляції
4. Регуляції сигналу за
негативним зворотнім
зв’язком
Eukaryota
Metazoa,
Embryophyta
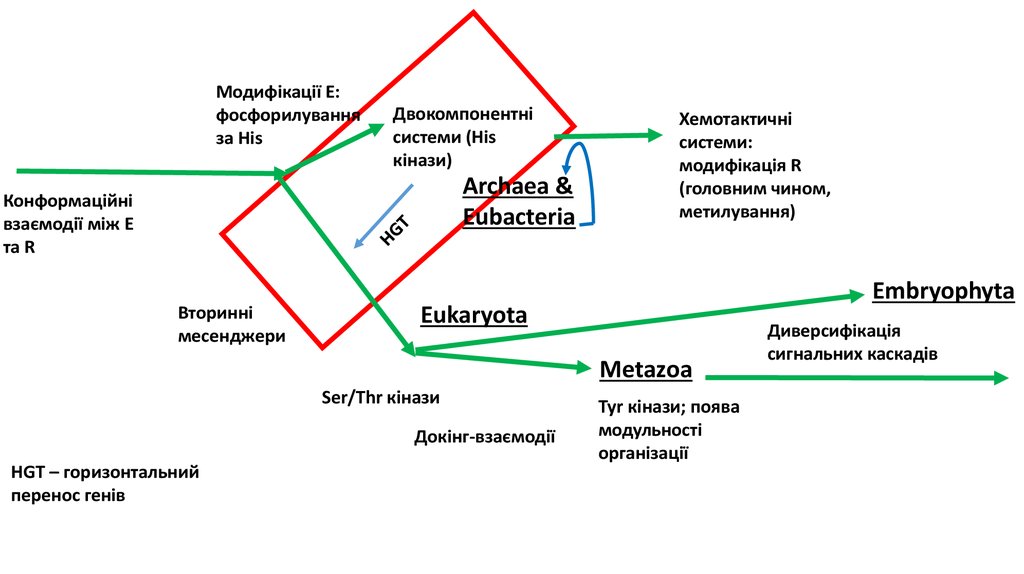
4. Еволюція механізмів сигналінгу
Модифікації E:фосфорилування
за His
Конформаційні
взаємодії між E
та R / R - канал
Вторинні
месенджери
Двокомпонентні
системи (His
кінази)
Archaea &
Eubacteria
HGT
Хемотактичні
системи:
модифікація R
(головним чином,
метилування)
Embryophyta
Eukaryota
Metazoa
Ser/Thr кінази
Докінг-взаємодії
HGT – горизонтальний
перенос генів
Tyr кінази; поява
модульності
організації
Диверсифікація
сигнальних каскадів
5. 1 етап
6. Рання еволюція сигнальної системи
• Одними із перших стимулів, що одночасно являються імесенджерами могли бути йони Ca2+, тому що:
1. Значна концентрація Са2+ токсична, тому має існувати значний
градієнт концентрації цього йону між зовнішньо- та
внутрішньоклітинним середовищем.
2. Кальцій – розповсюджений елемент у земній корі (5%).
3. Можуть існувати небілкові кальцієві транспортери.
7. Іонофори
• Ще один варіант простих переносників йонів (лише заградієнтом).
• Наприклад, A23187 – частково специфічний до Са2+.
8. Небілкові кальцієві канали у сучасних бактерій
• Полі-β-гідроксибутиратні канали відомі у Azotobactervinelandii, Bacillus subtilis, Haemophilus influenzae, і
Escherichia colі. Вони траспортують Са у зв’язаній з
поліфосфатом формі.
• Імовірно, можуть транспортувати в обидва напрямки.
Транспортують йони набагато швидше ніж іонофори.
Мономер
Reusch & Sadoff, 1988
9. Первинні системи сигналінгу без ампліфікації
1. Сигналінг із зовнішнімсередовищем
2. Примітивний
міжклітинний сигналиінг
Зміна зовнішньої
конц. Ca2+
Метаболіт 2
(ліпофільний)
Метаболіт 1
Е
Первинний Са канал
Е
Канал
Р
Р
Зміна внутрішньої
конц. Ca2+
Безпосередня
зміна активності
певного ферменту
Безпосередня
зміна активності
певних ферментів
10. Первинна кальцій-залежна система
• Кальцієва система зв’язана з іншою.• Повноцінний (незалежний від
Е
середовища) кальцієвий сигналінг
Р
може виникнути лише з появою
компартменталізації.
+\-
Індуктор 1
+\-
Синтаза індуктору 2
Індуктор 2
Безпосередня
зміна активності
певного ферменту
11. Можливі функції
1. Груповий гідроліз субстрату.2. Примітивний хемотаксис за принципом роботи аналогічний
прокаріотичному (є індуктор – є рух).
Проблеми такої системи:
1. Слабо підтримується груповим добором через дуже обмежене
поширення сигналу.
2. Залежність інтенсивності відповіді від інтенсивності стимулу –
необхідна ампліфікація сигналу.
12. 2 етап
13. Поява ампліфікації
1.2.
3.
4.
Модифікації білків – > двокомпонентні системи.
Транскрипційна регуляція.
Вторинні месcенджери.
Додатковий шлях у бактерій – ампліфікація позитивним
зв’язком (відчуття кворуму).
14. Еволюція сайтів фосфорилування
• Першим кроком до вирішенняпроблеми необхідності ампліфікації
сигналу була поява
посттрансляційних модифікацій
ефекторів (ефектор І порядка швидше
активує більшу кількість ефекторів ІІ
порядку).
• Поява сайтів фосфорилування
відбулася на місці негативно
заряджених амінокислот у білку (Glu,
Asp).
Pearlman et al., 2011
15. Двокомпонентні системи
Kristin K. Koretke, Andrei N. Lupas,Patrick V. Warren, Martin Rosenberg,
and James R. Brown Evolution of TwoComponent Signal Transduction Mol.
Biol. Evol. 17(12):1956–1970. 2000
Е
Двокомпонентні системи
Рецептор-гістидинова кіназа
Р
Фосфорилування за
залишком His
Ефектор (response regulator)
16.
Модифікації E:фосфорилування
за His
Двокомпонентні
системи (His
кінази)
Archaea &
Eubacteria
Конформаційні
взаємодії між E
та R
Вторинні
месенджери
Хемотактичні
системи:
модифікація R
(головним чином,
метилування)
Embryophyta
Eukaryota
Metazoa
Ser/Thr кінази
Докінг-взаємодії
HGT – горизонтальний
перенос генів
Tyr кінази; поява
модульності
організації
Диверсифікація
сигнальних каскадів
17. Поява модифікації рецепторів – хемотаксис бактерій
ХемоатрактантРегуляція за – зв’язком
Е
Рецептор MCP (Methyl-accepting Chemotaxis Protein)
Метилування рецептору
Р
CheR
Деметилування
рецептору
CheВ
Джгутик
CheW
CheY
CheA
18. Вторинні месенджери
• Циклічні нуклеотиди – найбільш розповсюджені як серед про- такі еукаріот.
cAMP
19. 3 етап
20. Прокаріоти - + зв’язок
21. Регуляція позитивним зворотнім зв’язком
• Повноцінна ампліфікація у сигнальній системі, що активнопідтримується груповим добором.
• Виникає у прокаріот після розходження з еукаріотами.
Ліпофільний автоіндуктор
Е
Р
Синтаза
індуктора
Ефектор
22. Кворум сенсинг (відчуття кворуму)
• Найчастіше ефектор і синтаза автоіндуктора знаходяться в одномуопероні.
Кворум сенсинг у симбіонта
кальмара Euprymna
scolopes Vibrio fischeri, який
забеспечує світіння за
допомогою фермента
люциферази.
Waters & Bassler, 2005
23. Бувають і складніші системи з декількома автоіндукторами
Кворум сенсинг уPseudomonas
aeruginosa
24. Еукаріоти – ускладнення рецепторів
25. Рецептори з ферментативною активністю
Серин-треонін кіназний рецептор– майже всі еукаріоти
Тирозин кіназний рецептор - Metazoa
26. G-білок залежні рецептори
27. Арешт рецепторів – негативна регуляція сигналу
Регуляція за – зв’язком28. Регуляторні мономерні ГТФази – «перемикачі»
29. Ускладнення регуляції
Зв’язування з активнимцентром
Докінг-взаємодії (за
межами активного
центру)
Розвиток модульності
(більший простір для
еволюції, регуляція стає
ще тендітніше)
Bhattacharyya et al., 2006
Скафолди/адаптерні
білки (просторова
колокалізація білків
сигнальних каскадів)
}
Багатоклітинні
30. Диверсифікація сигнальних каскадів
• Збільшення різноманітності сигнальних каскадів – важливапередумова появи багатоклітинності.
• Наприклад, у Metazoa з’являються повністю модульні
трьохкомпонентні системи, основані на фосфорилуванні
тирозину.
• Першими з’являються Eraser (PTP, phosphotyrosine phosphatase) домени
для виправлення випадкового фосфорилювання Tyr Ser/Thr
кіназами. Далі – Reader (SH2); функціональною система змогла
стати лише після появи Writer (TyrK, tyrosine kinase).
1. Eraser
2. Reader
3. Writer
Функціонуюча сигнальна
система
1. Pincus et al., 2008
2. Lim & Pawson, 2010
31. Висновки
• Таким чином, еволюція сигнальних систем йшла в напрямкузбільшення “еволюційної здатності” (evolvability) системи,
кількості шляхів її регуляції і удосконалення систем ампліфікації
сигналу (шлях оптимізації підтримки груповим-добором).