































 Медицина
МедицинаПохожие презентации:

")
")





")
")
Антитела. Клеточный и гуморальный иммунитет (лекция 3)
1. Лекция 3
Антитела. Клеточный игуморальный иммунитет.
2.
АнтителаАнтитела (АТ) - это γ-глобулины (Ig)
(растворимые белки), которые присутствуют в
сыворотке крови и других биологических
жидкостях,
способные
специфически
связываться с антигеном и участвовать во
многих иммунологических реакциях. Они
синтезируются
В-лимфоцитами
и
их
потомками - плазматическими клетками.
Составляют 15-25% белкового содержания
сыворотки крови.
3.
Свойства АТ:Аффинность – сила специфического
взаимодействия АТ с АГ. Зависит от
комплементарности (степени сродства)
антигенсвязывающего центра (паратопа) и
эпитопа.
Авидность – прочность связывания АТ и АГ.
Зависит от аффинности АТ и от количества
паратопов.
4.
Выделяют АТ:1. а)Циркулирующие: сывороточные и секреторные
(sIgA)
б) Рецепторные (мономеры IgM, IgD)
2. а) специфические
б) нормальные - базальный уровень АТ, против АГ
групп крови, против бактерий кишечной группы и
др. Функции: первая линия обороны против
инфекций, удаление из организма отживших клеток
(врожденный иммунитет, В1-лимфоциты).
3. а) полные – имеют два или более паратота
б) неполные – имеют только один
функционирующий паратоп
5.
Моноклональные АТ(АТ, полученные от одного клона лимфоцитов)
В ответ на поступление АГ в организме активируется несколько В-лимфоцитов, продуцирующих АТ
с разной степенью сродства к эпитопу АГ
(поликлональные АТ). Кроме этого один АГ может
иметь
множество
эпитопов.
Поэтому
моноклональные АТ получают искусственно.
Животным вводят АГ, затем выделяют антителопродуцирующую клетку, сливают ее с опухолевой и
получают гибридомы. Они продуцируют АТ строго
определенной специфичности и при этом
«бессмертны».
6.
Получение моноклональных антител7.
Антиглобулиновые антителаIg, как белки, обладают антигенными свойствами и
при попадании в чужеродный организм индуцируют
выработку против себя АТ.
При некоторых условиях в организме могут
образовываться антитела к собственным Ig. Так при
ревматоидном артрите в крови появляются
антитела класса М к собственным Ig G
(ревматоидный фактор).
8.
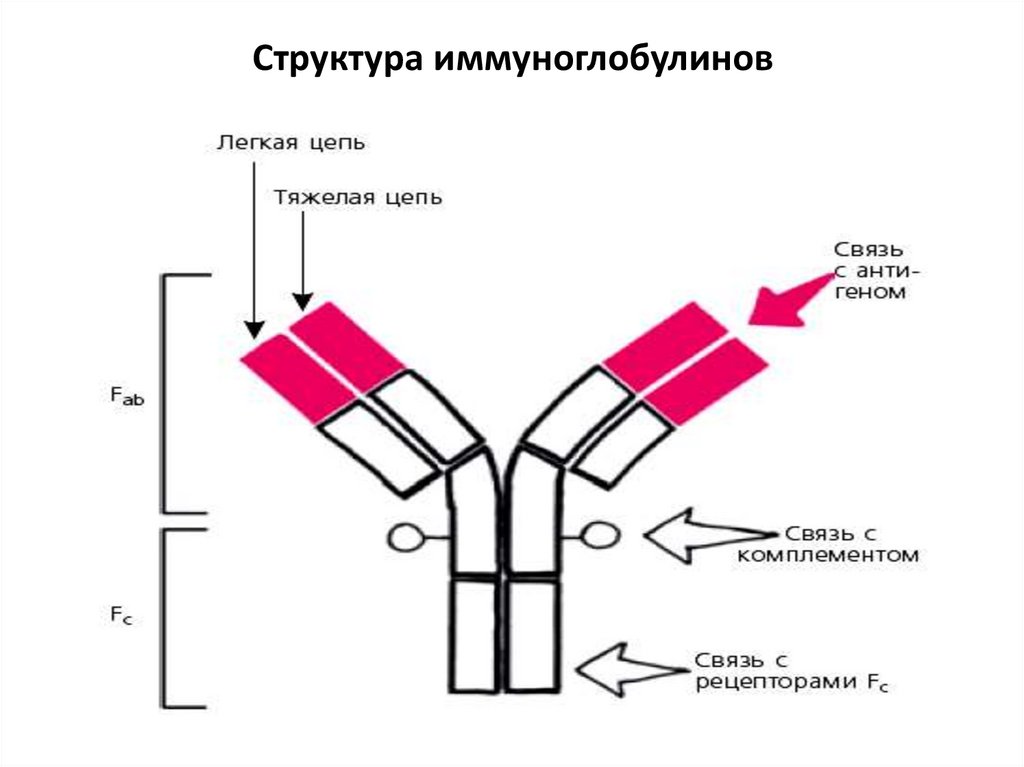
Структура иммуноглобулиновЯвляется белком с определенной биохимической
структурой. Состоит из 2 пар: легких (L) и тяжелых (H)
цепей. Цепи L и H связаны между собой
дисульфидными связями, между тяжелыми цепями
также есть дисульфидная связь (шарнирный участок).
Вторичная структура полипептидных цепей имеет
доменное строение (глобулы). В каждой тяжелой
цепи выделяют 4-5 доменов, в легкой – 2. Домены с
постоянным
аминокислотным
составом
–
константные (С), с переменной структурой –
вариабельные (V) с гипервариабельной областью.
Вариабельные домены легкой и тяжелой цепи
образуют антигенсвязывающий центр – паратоп.
9.
Легкие цепи бывают 2 типов: каппа (κ) и лямбда(λ). Тяжелые 5 типов: альфа (α), гамма (γ), мю (μ),
эпсилон (ε), дельта (δ). Тип тяжелой цепи
определяет класс Ig.
Дополнительные
полипептидные
цепи
в
структуре : J – пептид (соединяет мономеры), S –
пептид
(предохраняет
секреторный
Ig
от
ферментативного расщепления) и М–пептид
(удерживает рецепторный Ig в мембране Влимфоцита).
При обработке молекулы Ig ферментом папаином
она расщепляется на 2 Fab фрагмента ( связываются
с антигеном) и 1 Fc фрагмент (связывается с
рецепторами на мембране клеток макроорганизма).
10.
Структура иммуноглобулинов11.
Классы и типы иммуноглобулиновВыделяют 5 классов Ig:
• Иммуноглобулин класса G.
70-80% всех сывороточных Ig. Период полураспада
21 день. Мономер, имеет 2 антигенсвязывающих
центра, подтипы G1, G2, G3, G4. Связывает
комплемент,
единственный
проходит
через
плацентарный барьер, может выделяться в молоко.
Определяется в сыворотке на пике первичного и при
вторичном
иммунном
ответе.
После
взаимодействия IgG с АГ его Fc фрагмент может
связываться с макрофагами, нейтрофилами и NКклетками.
12.
• Иммуноглобулин класса М.5-10% всех сывороточных белков. Период
полураспада 5 дней. Пентамер, имеет 10
антигенсвязывающих
центров.
Синтезируется
первым при превичном контакте с АГ (первичный
иммунный ответ). Обладает высокой авидностью,
активирует комплемент по классическому пути.
Может выделятся в секрет слизистых, в молоко.
Мономеры имеются на
поверхности В-лимфоцита
в виде рецептора.
13.
• Иммуноглобулин класса А.1) Сывороточный Ig A. 10-15% всех сывороточных
белков. Период полураспада 6 дней. Мономер,
имеет 2 паратопа. Определяется в сыворотке на пике
первичного и при вторичном иммунном ответе. Не
связывает комплемент.
2) Секреторный Ig A (SIgA). Ди- или тример. Его пул
считается самым многочисленным в организме. Имеет S пептид,
который присоединяется к молекуле при прохождении ее через
эпителиальную клетку и защищает ее от ферментов. Обеспечивает
местный иммунитет на слизистых.
14.
• Иммуноглобулин класса Е. 0, 002% всехсывороточных иммуноглобулинов. Мономер. Не
связывает комплемент. Fc фрагмент связывается с
тучными клетками и базофилами. Участвует в
противопаразитарном иммунном ответе и
аллергических реакциях. Называют реагином.
• Иммуноглобулин класса D. Мономер. Может
быть рецептором В-лимфоцитов.
15.
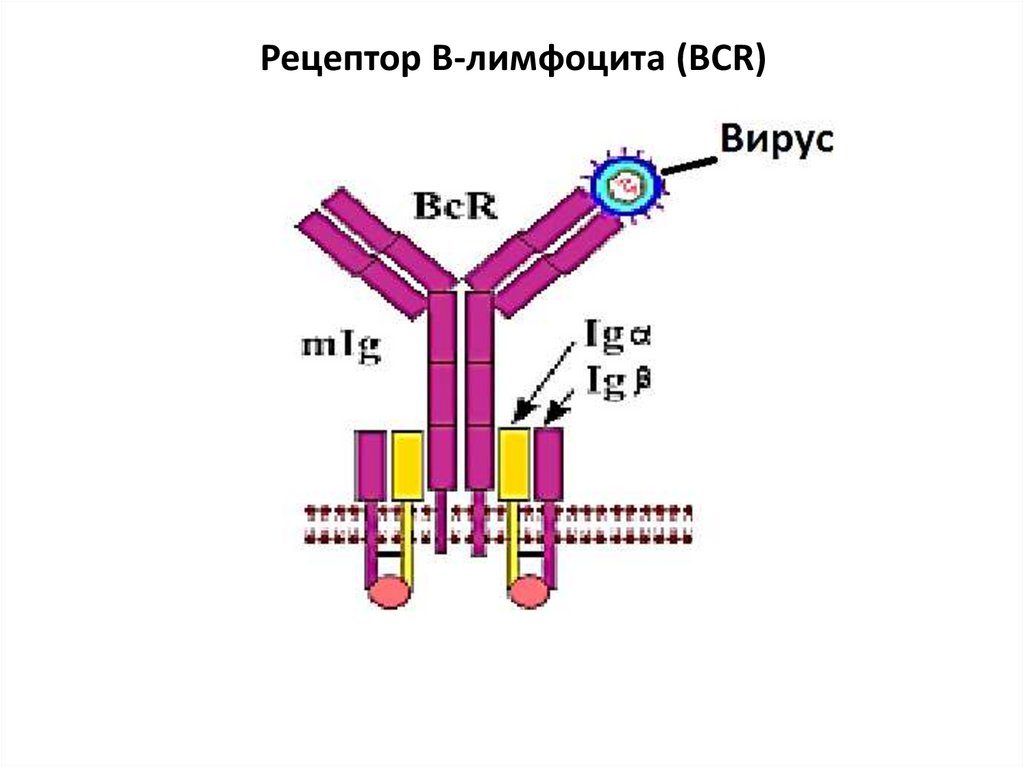
Рецепторы антигенреактивных лейкоцитовВ-клеточный рецептор (BCR) представлен иммуноглобулинами чаще классов М и D (мономеры) (строение
– см. структура Ig). Кроме молекулы иммуноглобулина в
формировании BCR участвуют еще два обязательных
полипептида Ig α и Ig β. Они необходимы для
проведения сигнала внутрь клетки и связаны с
внутриклеточными тирозинкиназами. Для того, чтобы
произошла
эффективная
активация
В-клетки,
необходима перекрестная сшивка антигеном нескольких
BCR. Для этого молекула антигена должна иметь
повторяющиеся эпитопы на своей поверхности. Один
лимфоцит содержит 200-500 тыс. однородных
рецепторов, обладающих специфичностью к одному
антигену.
16.
Рецептор В-лимфоцита (BCR)17.
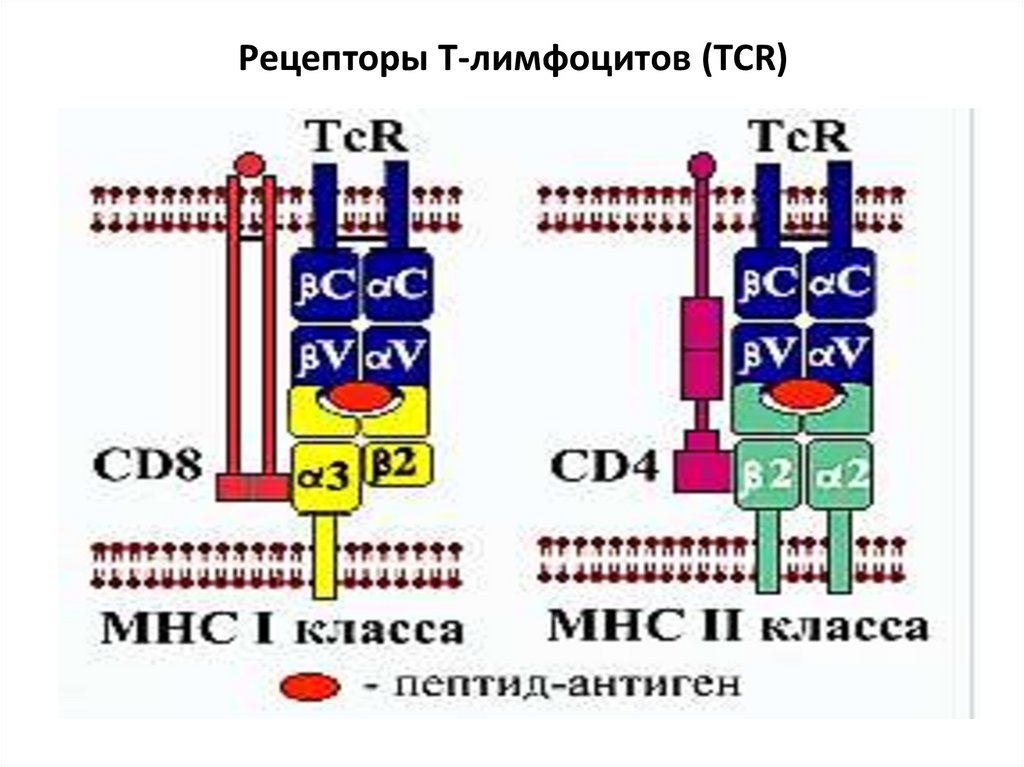
• Т-клеточный рецептор (TCR)Состоит из двух полипептидные цепей (αβ или γδ),
состоящих из вариабельных (V) и константных (С)
доменов, участки которых проходят через мембрану и
погружены в цитоплазму. Они связаны с комплексом
белков – CD3, который передает сигнал от рецептора в
клетку, запуская процесс активации. Вариабельные
домены образуют антигенсвязывающий центр.
TCR совместно с корецепторами CD4 или CD8 распознает
антиген в комплексе МНС I или II класса. Происходит
двойное распознавание: «своего» (МНС) и «чужого» (АГ).
TCR одного Т-лимфоцита распознают только один АГ.
TCR представлены двумя вариантами: αβ (более 90 % Тлимфоцитов крови) и γδ (много в коже, кишечнике,
репродуктивных органах – неспецифический иммунитет).
18.
Рецепторы Т-лимфоцитов (TCR)19.
• Fc-рецепторы (FcR)FcR- трансмембранная белковая молекула,
специализированная к определенному изотипу
тяжелой
цепи
молекулы
АТ.
Бывают
высокоаффинные (на базофилах и тучных клетках) –
связываются
с
интактной
молекулой
АТ,
низкоаффинные – связываются с молекулой АТ в
составе иммунного комплекса.
В свободном виде только IgЕ способны
связываться с тучными клетками и базофилами. Ig
других классов фиксируются на FcR после
связывания антитела с антигеном, то есть
фиксируется не свободное антитело, а комплекс АГАТ через Fc-конец молекулы иммуноглобулина.
20.
Антителозависимые механизмы защитыПрямые эффекты:
1)Нейтрализация.
АТ связывается с активным центром биологически
активной молекулы и тем самым блокирует ее
действие. Например: активный центр токсина,
рецепторы вируса.
2) Энзиматическое действие.
Ig способны расщеплять молекулы некоторых АГ.
Например: деструкция отдельных пептидов или
ДНК.
21.
В большинстве случаев связывание АТ с АГ неприводит к разрушению АГ. Чаще АТ, связываясь с АГ,
маркируют его как мишень для факторов
неспецифического иммунитета.
Непрямые эффекты:
1) Активация комплемента по классическому пути
с образованием МАК и лизисом клетки.
2)
Запуск
антителозависимой
клеточной
цитотоксичности
(АЗКЦТ).
Осуществляют
активированные макрофаги, NК-клетки, эозинофилы. Они имеют FcR, связывающийся с Fcфрагментом антител, которые предварительно
связались с антигеном.
22.
Связывание с иммунным комплексом индуцируету них выброс биологически активных веществ,
разрушающих клетку: перекисные и NО-радикалы,
ферменты у макрофагов; перфорины, гранзимы,
гранулизин у NК-клеток; ферменты, белковые
токсины у эозинофилов при противогельминтном
иммунитете.
3) Иммунный фагоцитоз.
Фагоциты также имеют FcR и способны поглощать
АГ в составе иммунного комплекса.
4) Развитие гиперчувствительности немедленного
типа (ГНТ) (IgЕ).
23.
Генетический контроль иммунного ответа.В организме здорового человека в течение жизни
образуется несколько миллионов вариантов
антител (отличающихся прежде всего паратопами),
которые способны связаться с практически любым
АГ, как природным, так и с искусственно
полученным. Геном человека физически не может
вместить столько отдельных структурных генов.
Количество
генов,
предназначенное
для
программирования
синтеза
АТ,
всего
120
(зародышевые гены). Эта проблема объясняется
теорией С. Тонегавы.
24.
В отличие от генов других белков, гены Ig имеютфрагментарную структуру и разбросаны в
хромосомах во многих экземплярах. Фрагменты
собираются вместе (комбинируются) в единый
функционирующий ген случайным образом, и число
возможных комбинаций исчисляется миллионами.
В каждом отдельном В-лимфоците при созревании
осуществляется своя уникальная рекомбинация
ДНК. Попавший в организм АГ избирательно
активирует определенный специфичный к нему
лимфоцит, который начинает размножаться и
синтезировать АТ.
25.
Гены Ig фрагментированы и образуют три группы,которые располагаются в трех различных
хромосомах и наследуются независимо. 1 группа
генов содержит информацию о первичной
аминокислотной последовательности легкой цепи
каппа (к), вторая – лямбда (λ), третья – всех типов
тяжелых цепей: альфа (α), гамма (γ), мю (μ), эпсилон
(ε), дельта (δ).
Легкая и тяжелая цепи Ig, состоят из константного
домена– С (относительно неизменного во всех АТ) и
вариабельного – V.
26.
Тяжелая цепь (Н) кодируется генами из 4 участков:1) V (вариабельный) имеет около 50 сегментов
2) D (вносящий разнообразие) ≈ 20 сегментов
3) J (соединяющий) ≈ 6 сегментов
4) С (константный) 9 сегментов, определяющих
класс и подкласс.
В результате, для образования вариабельного
домена Ig в структурный ген соединяются по одному
сегменту из V, D, J областей. Вся остальная ДНК этих
областей вырезается и выбрасывается из генома.
Поэтому
дифференцировка
В-лимфоцита
и
приобретение им специфичности необратима на
уровне
ДНК.
Рекомбинацию
катализируют
ферменты рекомбиназы.
27.
Гены легких цепей образованы сочетаниемсегментов из 3 своих областей: V, J (вариабельный
домен) и С.
Так как паратоп образован сочетанием легкой и
тяжелой цепей, это еще больше увеличивает
многообразие вариантов.
Кроме этого есть еще два молекулярных процесса:
1) Запланированная неточность связи сегментов VDJ
2) Гипермутагенез в V-генах – повышенное
количество спонтанных мутаций. Происходит уже
после активации во время пролиферации Влимфоцитов и приводит к селекции в направлении
повышения специфичности рецепторов и АТ.
28.
Гены тяжелой цепи (Н)29.
Переключение классов иммуноглобулиновСтруктурные гены константных частей Ig
расположены в тех же хромосомах, что и VDJ гены и
обозначаются - С. Для каждого изотипа (класса) Ig
есть свой С-ген (Сα, Сγ, Сε, Сδ, Сμ ). Завершившие
лимфопоэз
В-лимфоциты
экспрессируют
иммуноглобулины только М и D. Переключение на
синтез
иммуноглобулинов
других
изотипов
происходит уже в процессе развития иммунного
ответа, то есть после распознавания антигена. Это
переключение
идет
тоже
по
механизму
рекомбинации ДНК. ДНК неиспользуемых С-генов
вырезаются в виде кольцевых структур. С этого
момента В-лимфоцит строго специфичен, он
становится плазматической клеткой и продуцирует
большое количество определенных АТ.
30.
Переключение классов иммуноглобулинов31.
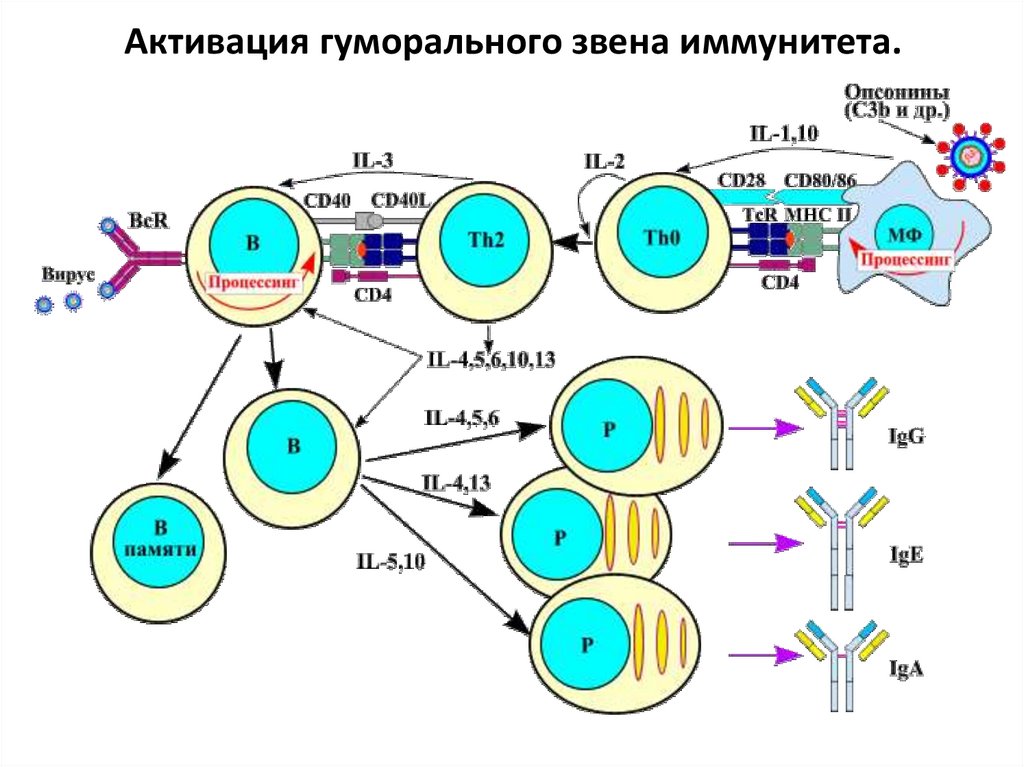
Гуморальный иммунитет.Основа – активация В-лимфоцитов и их
дифференцировка
в
антителопродуцирующие
плазматические клетки.
АГ поступивший в организм захватывается АПК,
происходит его процессинг (неполное переваривание до низкомолекулярных пептидов) и эпитоп АГ
выставляется на поверхности АПК в комплексе с
МНС II класса. Этот АГ в комплексе с МНС II класса
распознает наивный Т-хелпер с помощью своего
рецептора TCR и корецептора CD4. С данным АГ из
всего пула лимфоцитов связывается именно тот Тхелпер, рецептор которого оказался комплементарен данному АГ.
32.
Также между двумя клетками образуются связимолекул CD28 – CD80/86 и др. Получив
необходимую цитокиновую стимуляцию от АПК и
стимуляцию интерлейкином-2, наивный Т-хелпер
активируется,
начинает
пролиферировать
и
дифференцируется в иммунный Т-хелпер 2 типа
(Th2).
В то же время наивный В-лимфоцит, рецептор
(BCR) которого оказался комплементарен данному
виду АГ, также связывается с АГ, осуществляет
эндоцитоз, затем процессинг и выставляет эпитоп
АГ на своей поверхности в комплексе с МНС II
класса. В таком виде его распознает при помощи
своего TCR и CD4 иммунный Т-хелпер 2 типа.
33.
Также между двумя клетками образуются связимолекул CD28 – CD80/86, CD40-CD40L и др. После
этого, получив интерлейкиновый стимул от Тхелпера, наивный В-лимфоцит активируется,
начинает пролиферировать, дифференцируется в
иммунный
и
начинает
продуцировать
АТ
специфичные данному АГ (какой специфичности
рецептор – такие и АТ). Первыми всегда
синтезируются
IgM,
а
затем
происходит
переключение на другие классы и подклассы.
Конечная стадия дифференцировки – плазмоциты.
В то же время часть клонов В-лимфоцита
дифференцируются в долгоживущие клетки памяти.
34.
Активация гуморального звена иммунитета.35.
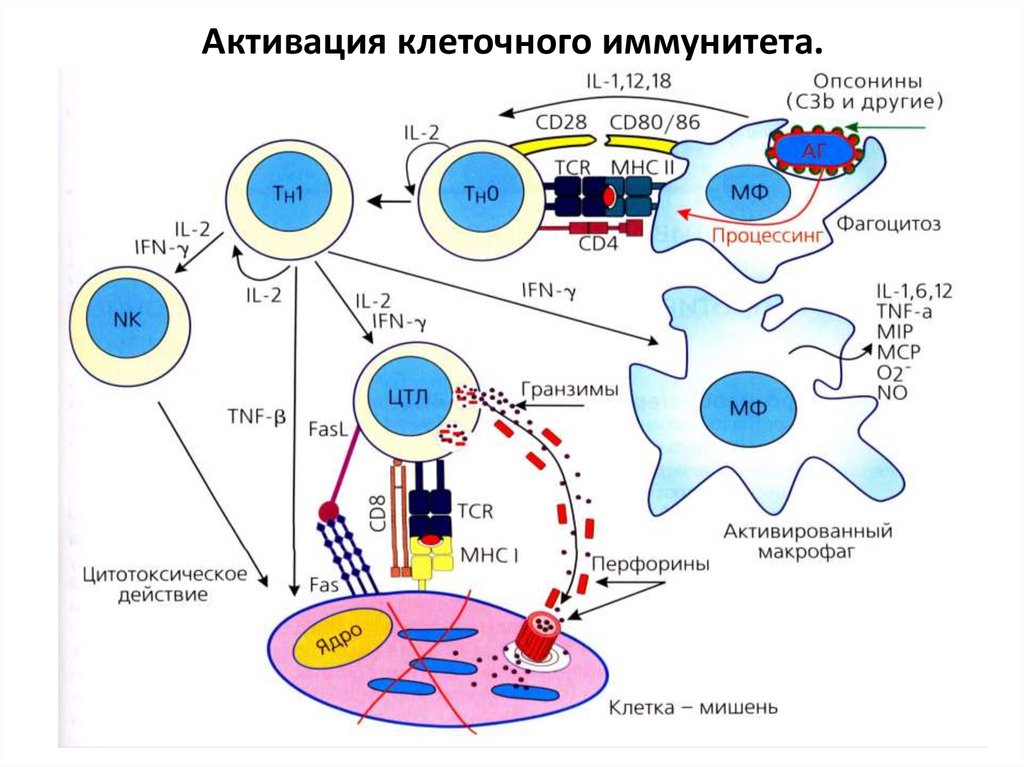
Клеточный иммунитет.Основа – активация ЦТЛ, а также макрофагов и NКклеток.
При поступлении АГ в организм он захватывается
АПК, происходит его процессинг и эпитоп АГ
выставляется на поверхности АПК в комплексе с
МНС II класса. Этот АГ в комплексе с МНС II класса
распознает наивный Т-хелпер с помощью своего
рецептора TCR и корецептора CD4. Также между
двумя клетками образуются связи молекул CD28 –
CD80/86 и др. Получив необходимую цитокиновую
стимуляцию от АПК и аутокринную стимуляцию
интерлейкином-2, наивный Т-хелпер активируется,
начинает пролиферировать и дифференцируется в
иммунный Т-хелпер 1 типа (Th1).
36.
Также эпитоп поглощенного АПК антигенавыставляется на ее поверхности в комплексе с МНС I
класса. ЦТЛ при помощи своего рецептора ТCR и
корецептора CD8 распознает этот АГ в МНС I класса
(эта связь стабилизируется CD28-CD80/86) и, получив
стимуляцию гамма-интерфероном (IFN-γ) от Тхелпера 1 типа и IL-2 , активируется, пролиферирует
и дифференцируется в иммунный ЦТЛ.
Иммунные ЦТЛ начинают при помощи своего
рецептора TCR и CD8 искать клетки-мишени, которые
на своей поверхности в составе МНС I класса имеют
данный чужеродный АГ. Обнаружив такую клетку,
они уничтожают ее, выделяя перфорины, гранзимы,
гранулизин или через образование связи Fasрецептор – Fas-лиганд (Fas- L), запускающей апоптоз.