














































 облегчённой диффузии с участием переносчиков (транслоказ). S1, S2- разные молекулы.")








 Биология
БиологияПохожие презентации:




")





Биологические мембраны. Основные компоненты. Строение. Свойства. Функции. Трансмембранный транспорт. G – белки
1. Биологические мембраны. Основные компоненты. Строение. Свойства. Функции. Трансмембранный транспорт. G – белки.
Иванова И.Б., к.б.н. доцент кафедрыбиологии и генетики ДВГМУ
2. План лекции:
1. Ультраструктура клеточных мембран.2. Молекулярное строение
цитоплазматических мембран.
3. Физическая природа сил взаимодействия
белков и липидов в структурах мембран.
4. Методы изучения мембран.
5. Искусственные модели
цитоплазматических мембран.
3. Важнейшими условиями существования клетки является:
1. Автономность по отношению к окружающейсреде.
2. Связь с окружающей средой.
4.
Клеточная мембра́на (также цитолемма, плазмолемма, илиплазматическая мембрана) — эластическая молекулярная
структура, состоящая из белков и липидов. Отделяет
содержимое любой клетки от внешней среды, обеспечивая её
целостность; регулирует обмен между клеткой и средой;
внутриклеточные мембраны разделяют клетку на
специализированные замкнутые отсеки — компартменты или
органеллы, в которых поддерживаются определённые условия
среды.
5. История исследования
• В 1925 году Гортер и Грендель с помощью осмотического удараполучили так называемые «тени» эритроцитов — их пустые
оболочки. Был открыт липидный бислой.
• В 1935 году Даниэлли и Доусон представили научному
сообществу модель «сендвича», которая говорит о том, что в
основе мембраны лежит липидный бислой, по обеим сторонам
от которого находятся сплошные слои белков, внутри бислоя
ничего нет. Первые электронно-микроскопические исследования
1950-х годов подтвердили эту теорию — на микрофотографиях
были видны 2 электронно-плотных слоя — белковые молекулы и
головки липидов и один электронно-прозрачный слой между
ними — хвосты липидов.
6. Молекулярное строение клеточных мембран
- В 1902 г. - Овертон предположил, что мембранасостоит из тонкого слоя липидов.
- В1925 г. - Гортер и Грендел в опытах с липидами,
экстрагированными из мембран эритроцитов,
обнаружили, что площадь монослоя липидов вдвое
больше суммарной площади поверхности всех
эритроцитов.
Был сделан вывод, что липиды мембраны расположены в
виде бимолекулярного слоя.
7. Молекулярное строение клеточных мембран
В 1935 Даниэлли и Доусон предложилимодель строения мембраны, получившей
название «сэндвич».
Согласно этой модели, имеются 2 слоя
молекул фосфолипидов, которые
окружены слоями белка.
Трехслойная модель СтенаДаниэлли с полярной порой,
образованной молекулами белка.
8. Молекулярное строение клеточных мембран
В 1959 г. Дж. Робертсоном былавыдвинута теория унитарной мембраны,
состоящей из одного слоя
фосфолипидов, покрытого с
цитоплазматической стороны слоем
фибриллярного белка, а с наружной
поверхности – мукополисахаридами или
мукопротеидами.
Модель Шестранда – липидная фаза
мембраны существует в виде
глобулярных мицелл,
представляющих комплекс
липидных молекул, залитых в
белковый матрикс.
9.
Все это привело к созданию в 1972году Сингером (S. Jonathan Singer) и
Николсоном (Garth L. Nicolson)
жидкостно-мозаичной модели
строения мембраны.
Согласно этой модели белки в
мембране не образуют сплошной
слой на поверхности, а делятся на
интегральные, полуинтегральные
и периферические.
10.
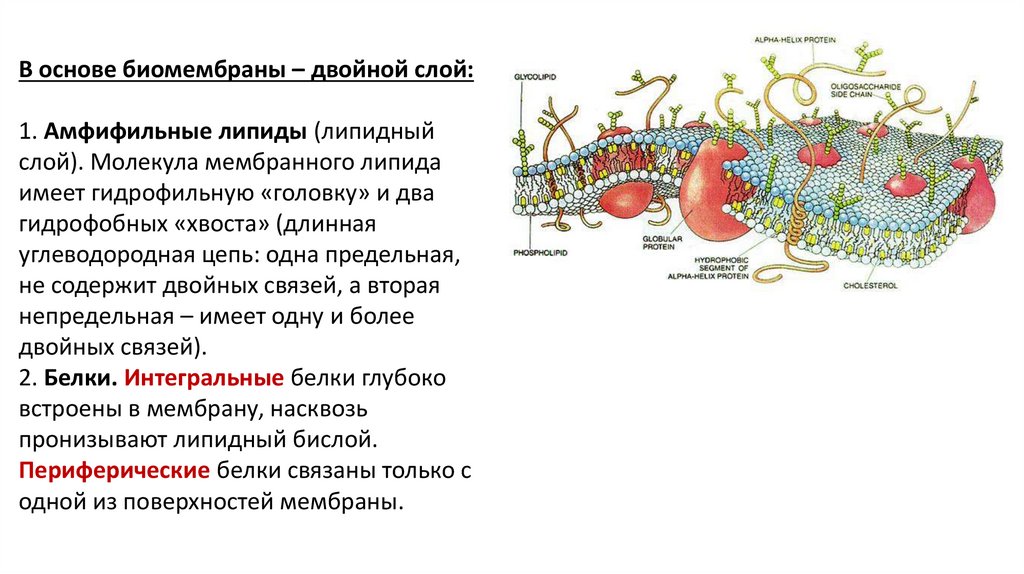
В основе биомембраны – двойной слой:1. Амфифильные липиды (липидный
слой). Молекула мембранного липида
имеет гидрофильную «головку» и два
гидрофобных «хвоста» (длинная
углеводородная цепь: одна предельная,
не содержит двойных связей, а вторая
непредельная – имеет одну и более
двойных связей).
2. Белки. Интегральные белки глубоко
встроены в мембрану, насквозь
пронизывают липидный бислой.
Периферические белки связаны только с
одной из поверхностей мембраны.
11. Структура и состав биомембран
• Мембраны состоят из липидов трёх классов: фосфолипиды,гликолипиды и холестерол.
• Фосфолипиды и гликолипиды (липиды с присоединёнными к ним
углеводами) состоят из двух длинных гидрофобных
углеводородных «хвостов», которые связаны с заряженной
гидрофильной «головой».
• Холестерол придаёт мембране жёсткость, занимая свободное
пространство между гидрофобными хвостами липидов и не
позволяя им изгибаться. Поэтому мембраны с малым
содержанием холестерола более гибкие, а с большим — более
жёсткие и хрупкие. Также холестерол служит «стопором»,
препятствующим перемещению полярных молекул из клетки и в
клетку.
12.
Липидный слой – это основная структура мембраны,которая создает относительно непроницаемый барьер
для большинства водорастворимых молекул.
Классы:
• Фосфолипиды (ФЛ)
• Сфинголипиды (СЛ)
• Гликолипиды (ГЛ)
• Стероиды, холестерин (ХС)
13.
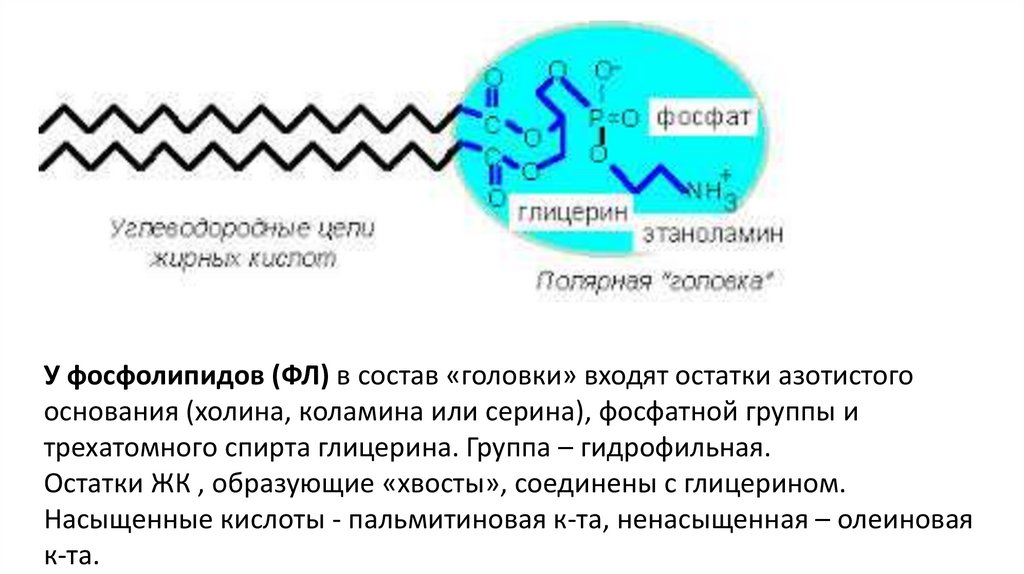
У фосфолипидов (ФЛ) в состав «головки» входят остатки азотистогооснования (холина, коламина или серина), фосфатной группы и
трехатомного спирта глицерина. Группа – гидрофильная.
Остатки ЖК , образующие «хвосты», соединены с глицерином.
Насыщенные кислоты - пальмитиновая к-та, ненасыщенная – олеиновая
к-та.
14. Строение фосфолипида
15.
Сфинголипиды (СЛ) – вместо глицерина и одной изжирной кислот, включают сфингозин (сфингенин) –
2-х атомный аминоспирт.
Существует несколько заболеваний, связанных с
нарушенным метаболизмом сфинголипидов
(сфинголипидозы). Наиболее распространённая из
них — это болезнь Гоше.
16.
Гликолипиды - сложные липиды, образующиеся в результатесоединения липидов с углеводами. В молекулах гликолипидов
есть полярные «головы» (углевод) и неполярные «хвосты»
(остатки жирных кислот). Благодаря этому гликолипиды
(вместе с фосфолипидами) входят в состав клеточных
мембран.
Гликолипиды широко представлены в тканях, особенно в
нервной ткани, в частности в ткани мозга. Они локализованы
преимущественно на наружной поверхности плазматической
мембраны, где их углеводные компоненты входят в число
других углеводов клеточной поверхности.
17.
Холестери́н (др.-греч. χολή — желчь и στερεός — твёрдый) —органическое соединение, природный жирный (липофильный)
спирт, содержащийся в клеточных мембранах всех живых
организмов, за исключением безъядерных (прокариоты).
Нерастворим в воде, растворим в жирах и органических
растворителях.
Холестерин обеспечивает стабильность клеточных мембран в
широком интервале температур. Он необходим для выработки
витамина D, выработки надпочечниками различных стероидных
гормонов, включая кортизол, альдостерон, женских половых
гормонов эстрогенов и прогестерона, мужского полового гормона
тестостерона, играет важную роль в деятельности нервной и
иммунной системы.
18.
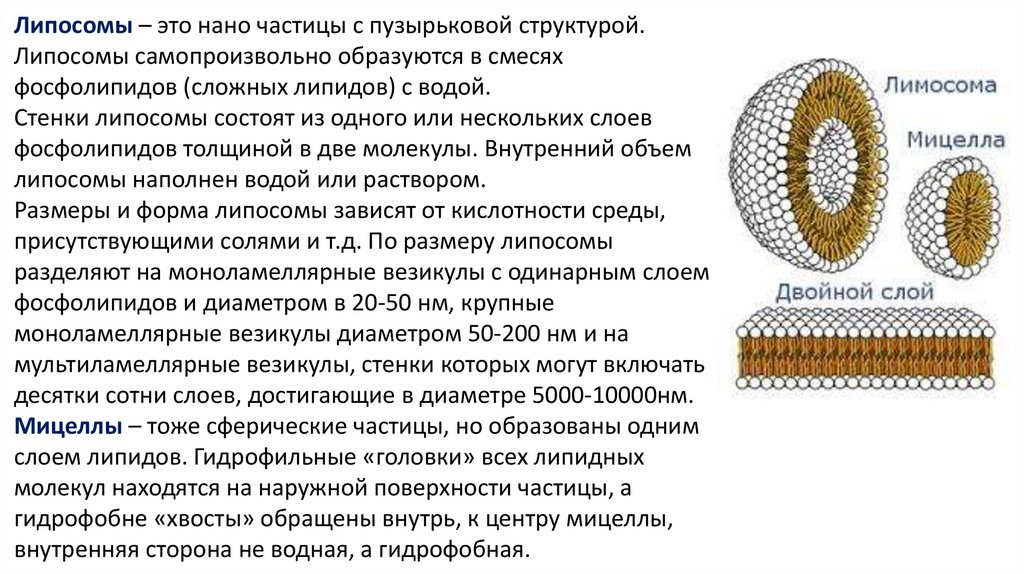
Липосомы – это нано частицы с пузырьковой структурой.Липосомы самопроизвольно образуются в смесях
фосфолипидов (сложных липидов) с водой.
Стенки липосомы состоят из одного или нескольких слоев
фосфолипидов толщиной в две молекулы. Внутренний объем
липосомы наполнен водой или раствором.
Размеры и форма липосомы зависят от кислотности среды,
присутствующими солями и т.д. По размеру липосомы
разделяют на моноламеллярные везикулы с одинарным слоем
фосфолипидов и диаметром в 20-50 нм, крупные
моноламеллярные везикулы диаметром 50-200 нм и на
мультиламеллярные везикулы, стенки которых могут включать
десятки сотни слоев, достигающие в диаметре 5000-10000нм.
Мицеллы – тоже сферические частицы, но образованы одним
слоем липидов. Гидрофильные «головки» всех липидных
молекул находятся на наружной поверхности частицы, а
гидрофобне «хвосты» обращены внутрь, к центру мицеллы,
внутренняя сторона не водная, а гидрофобная.
19.
Мембранные липиды – амфипатические молекулы,и в водной среде большинство из них
самопроизвольно образует бислои.
Именно это свойство обусловливает самосборку
липидных бислоев в клетке и их способность
самопроизвольно восстанавливаться при
повреждениях.
Различный липидный состав характерен и для
разных мембран одной и той же эукариотической
клетки.
20.
Важную часть мембраны составляют белки,пронизывающие её и отвечающие за
разнообразные свойства мембран. Их состав и
ориентация в разных мембранах различаются.
Рядом с белками находятся аннулярные липиды —
они более упорядочены, менее подвижны, имеют в
составе более насыщенные жирные кислоты и
выделяются из мембраны вместе с белком. Без
аннулярных липидов белки мембраны не работают.
21.
Белки мембраны клеток представлены, восновном, гликопротеинами.
Различают:
•интегральные белки, проникающие через всю
толщу мембраны и
•периферические белки, прикрепленные
только к поверхности мембраны, в основном, к
внутренней ее части.
22.
Периферические белки почти все функционируют какэнзимы (ацетилхолинестераза, кислая и шелочная фосфатазы
и др.). Но некоторые энзимы также представлены
интегральными белками — АТФ-аза.
Интегральные белки обеспечивают селективный обмен
ионов через каналы мембран между экстрацеллюлярной и
интрацеллюлярной жидкостью, а также действуют как белки
— переносчики крупных молекул.
Рецепторы и антигены мембраны могут быть представлены
как интегральными, так и периферическими белками.
Белки, примыкающие к мембране с цитоплазматической
стороны, относятся к цитоскелету клетки. Они могут
прикрепляться к мембранным белкам.
23.
Схема расположения белков в примембранном цитоскелетеэритроцитов.
1 — спектрин; 2 — анкирин; 3 — белок полосы 3; 4 — белок полосы 4,1; 5
— белок полосы 4,9; 6 — олигомер актина; 7 — белок 6; 8 — гпикофорин А;
9 — мембрана.
24.
Спектрин является основным белком цитоскелета,составляющим двумерную сеть, к которой
прикрепляется актин.
Актин образует микрофиламенты, представляющие
собой сократительный аппарат цитоскелета.
Цитоскелет позволяет клетке проявлять
гибкоэластические свойства, обеспечивает
дополнительную прочность мембраны.
25.
Гликофорин – интегральный белок. Онпронизывает всю мембрану и с обей сторон
выступает над ее поверхностью.
В состав белка входит сиаловая кислота,
благодаря ей, эритроцит несет отрицательный
заряд, который препятствует слипанию
эритроцитов друг с другом.
26. Виды мембранных белков
• Структурные• Транспортные
• Белки, обеспечивающие межклеточное
взаимодействие
• Белки, участвующие в передачи сигналов
27. Структурные белки
• Придают клетке и органеллам определеннуюформу
• Придают мембране механические свойства
(эластичность и т.д.)
• Обеспечивают связь мембраны с цитоскелетом
или (в случае с ядерной мембраной) с
хромосомами.
28. Транспортные белки
• Создают устойчивые транспортные потокиопределенных веществ через мембраны
• Транспорт ионов приводит к возникновению
трансмембранного потенциала во всех клетках, а
также к его изменениям в нервных и мышечных
клетках и волокнах (возбудимость и
проводимость).
29. Белки, обеспечивающие межклеточное взаимодействие
• Адгезивные белки – для связывания клетокдруг с другом или неклеточными структурами
(базальной мембраной, волокнами).
• Белки, участвующие в образовании
специализированных межклеточных
контактов (демосом и др.).
30. Белки, участвующие в передачи сигналов
•Рецепторные белки•Белки эффекторного устройства
•Фермент инактивации медиатора.
31.
Клеточные мембраны часто асимметричны, то есть слоиотличаются по составу липидов, в наружном содержатся
преимущественно фосфатидилинозитол, фосфатидилхолин,
сфингомиелины и гликолипиды, во внутреннем —
фосфатидилсерин, осфатидилэтаноламин и
фосфатидилинозитол. Переход отдельной молекулы из
одного слоя в другой (так называемый флип-флоп)
затруднён, но может происходить спонтанно, примерно раз
в 6 месяцев или с помощью белков-флиппаз и скрамблазы
плазматической мембраны. Если в наружном слое
появляется фосфатидилсерин, это является сигналом для
макрофагов о необходимости уничтожения клетки.
32. Гидрофобные и гидрофильные силы взаимодействия
В подержание структуры мембраны большуюроль играют гидрофобные и гидрофильные силы
взаимодействия. За счет гидрофобных сил
взаимодействия формируется липидный бислой,
а также прикрепление к липидному слою
неполярных областей в белке.
33. Гидрофильные силы взаимодействия способствуют удержанию белков на поверхности бислоя фосфолипидов. Таким образом, гидрофильные
игидрофобные силы взаимодействия способствуют
поддержанию стабильной целостности структуры
биологической мембраны, а также обеспечивают
высокую подвижность, приспособляемость к
выполнению многочисленных функций.
34. Основные свойства мембран
•Замкнутость•Латеральная подвижность
•Асимметрия
35. Замкнутость
Липидные бислои (и мембраны) всегдасамостоятельно замыкаются на себе с
образованием полностью отграниченных
отсеков.
При нарушении целостности мембраны
происходит ее «самосшивание».
36. Латеральная подвижность
Компоненты мембраны могут перемещаться впределах своего слоя.
Мембраны обладают свойствами двумерных
жидкостей. По этой причине модель строения
биомембран называется жидкостно-мозаичной.
Некоторые мембранные белки способны совершать
вращательные движения, меняя свою ориентацию
относительно поверхностей мембраны.
37. Асимметрия
Наружная и внутренняя поверхности мембраны различаются по своемусоставу:
1. Углеводные компоненты находятся с внешней поверхности
плазмолеммы;
2. Многие белки расположены всегда только с наружной, а другие только с внутренней стороны;
3. Нередко различается липидный состав слоев бислоя.
Полярность (асимметрия) мембраны возникает на ранних стадиях ее
формирования и затем все время сохраняется.
38. Свойства мембраны
• 1. Проницаемость.• 2. Полупроницаемость.
• 3. Избирательная проницаемость.
• 4. Активная проницаемость.
• 5. Управляемая проницаемость.
• Как видим, основное свойство мембраны - это её проницаемость
по отношению к различным веществам.
• 6. Фагоцитоц и пиноцитоз.
• 7. Экзоцитоз.
• 8. Наличие электрических и химических потенциалов.
• 9. Изменения электрического потенциала.
39.
• 10. Раздражимость.• 11. Каталитическая ферментативная активность. Ферменты
могут быть встроены в мембрану или связаны с её
поверхностью (как внутри, так и снаружи клетки), и там они
осуществляют свою ферментативную деятельность.
40. Основные функции биологических мембран:
1. Барьерная – обеспечивает селективныйрегулируемый, пассивный и активный обмен веществ
2. Матричная – обеспечивает определенное взаимное
расположение и ориентацию мембранных белков,
обеспечивает их оптимальное взаимодействие
41. Основные функции биологических мембран:
3. Механическая – обеспечивает прочность иавтономность клетки, внутриклеточных структур
4. Энергетическая – синтез АТФ на внутренних
мембранах митохондрий и фотосинтез в
мембранах хлоропластов
42. Основные функции биологических мембран:
5. Рецепторная (механическая, акустическая,обонятельная, зрительная, химическая,
терморецепция и тд.)
6. Генерация и проведение биопотенциалов
43. Существуют 4 способа ассоциации белков с липидным слоем:
1. интегральные белки, пронизывающиемембрану насквозь
2. поверхностные белки, погруженные в
липидном бислое частично
3. белки, удерживающиеся
нековалентными взаимодействиями с
другими мембранными белками
4. белки, ковалентно соединенные с одной
или двумя цепями жирных кислот.
44. Методы изучения и исследования биомембран
Метод рентгеноструктурного анализа – основанна явлении дифракции. Дифракция наблюдается в
тех случаях, когда на пути лучей имеются
препятствия, сравнимые по размерам с длиной
волны лучей. Метод рентгеноструктурного анализа
заключается в том, что на исследуемый объект
направ-ляют параллельный пучок рентгеновских
лучей. За объектом помещают фотопленку, на
которой регистрируется получающаяся
дифракционная картинка.
45. Методы изучения и исследования биомембран
1. Метод рентгеновского рассеивания нейтронов.Метод рассеивания нейтронов под малыми
углами позволяет определить толщину бислоя
фосфолипидов и расстояние между слоями.
46. Метод моделирования мембран
1. В 1917 г. Лэнгмюр разработал техникуполучения модели мономолекулярного слоя
фосфолипидов – модель «частокол
Лэнгмюра». На поверхность воды наносили
каплю фосфолипидов, растворенных в
каком-либо растворителе. Молекулы
липидов располагаются перпендикулярно к
поверхности воды: гидрофильная полярная
группа «головка» погружена в воду, а
неполярный «хвостик» направлен вверх.
воздух
вода
47. Метод моделирования мембран
2.В 1962 Мюллер и Рудин разработали способ получения больших двуслойных
мембран (d до 2 мм), используя тефлоновую пластину, разделяющую две водные
фазы. На отверстие пластины наносили смесь фосфолипидов в растворе летучих
растворителей. Происходило самопроизвольное формирование бислоя
фосфолипидов толщиной 6 нм.
48. Метод моделирования мембран
Получение липосом - замкнутыемембранные пузырьки, содержащие
водную фазу внутри и находящиеся в
водной среде. Липосомы формируют из
липидов в водной среде путем
обработки ультразвуком или быстрого
смешивания раствора фосфолипида в
этаноле с водой.
49. Перенос веществ через мембраны
1. Низкомолекулярные соединения:А) простая диффузия
Б) облегченная диффузия
В) активный транспорт
50. Облегченная диффузия
Молекула глюкозы связываетсяпереносчиком на наружной поверхности
плазматической мембраны. Происходит
конформационное изменение, и центр
переносчика, занятый глюкозой,
оказывается открытым внутрь клетки.
Вследствие конформационных изменений
переносчик теряет сродство к глюкозе, и
молекула высвобождается в цитозоль
клетки. Отделение глюкозы от переносчика
вызывает конформационные изменения
белка, и он возвращается к исходной
"информации.
51. Типы (виды) облегчённой диффузии с участием переносчиков (транслоказ). S1, S2- разные молекулы.
Некоторые транслоказымогут переносить два
разных вещества по
градиенту концентраций в
одном направлении пассивный симпорт, или в
противоположных
направлениях - пассивный
антипорт.
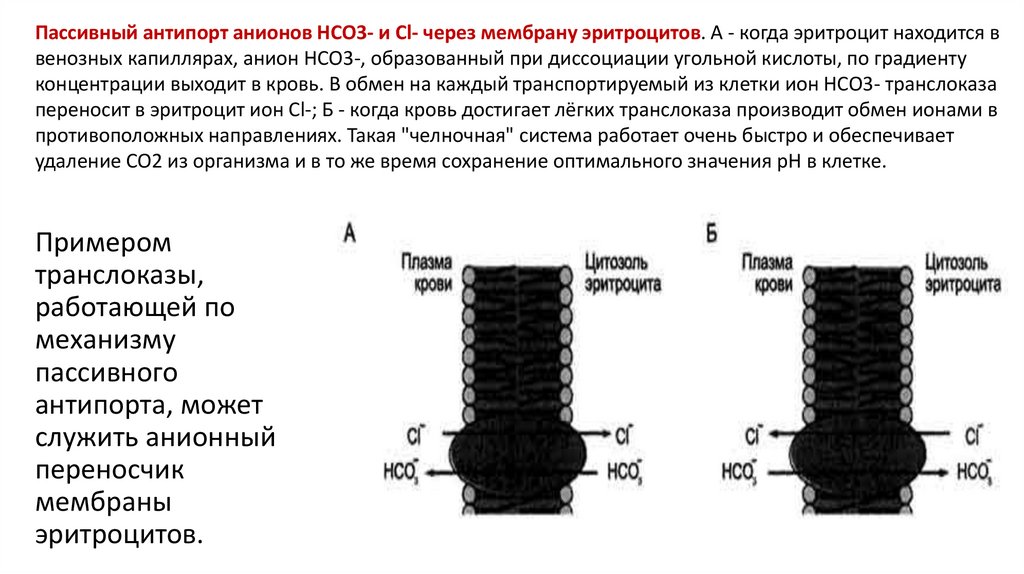
52.
Пассивный антипорт анионов НСО3- и Сl- через мембрану эритроцитов. А - когда эритроцит находится ввенозных капиллярах, анион НСО3-, образованный при диссоциации угольной кислоты, по градиенту
концентрации выходит в кровь. В обмен на каждый транспортируемый из клетки ион НСО3- транслоказа
переносит в эритроцит ион Cl-; Б - когда кровь достигает лёгких транслоказа производит обмен ионами в
противоположных направлениях. Такая "челночная" система работает очень быстро и обеспечивает
удаление СО2 из организма и в то же время сохранение оптимального значения рН в клетке.
Примером
транслоказы,
работающей по
механизму
пассивного
антипорта, может
служить анионный
переносчик
мембраны
эритроцитов.
53. Строение и функционирование Nа+,К+-АТФ-азы плазматической мембраны.
1 - три иона натрия связываются специфическим центромтранслоказы; 2 - изменение конформации транслоказы,
вызванное присоединением 3Na+, приводит к активации
каталитической субъединицы и увеличению сродства
активного центра к субстрату (АТФ). Протекает реакция
аутофосфорилирования по карбоксильной группе
аспарагиновой кислоты; 3 - аутофосфорилирование
изменяет заряд и конформацию транслоказы, она
закрывается с внутренней стороны мембраны и
открывается с наружной, уменьшается сродство к ионам
натрия и они диссоциируют от переносчика; 4 - Na+, К+АТФ-аза открытая с наружной стороны мембраны имеет
специфический центр связывания для 2К+; Присоединение
двух ионов калия к фосфорилированной транслоказе
вызывает изменение конформации и появление
аутофосфатазной активности. Протекает реакция
аутодефосфорилирования; 5 - дефосфорилирование
изменяет заряд и конформацию транслоказы, она
закрывается с наружной стороны мембраны и открывается
с внутренней, уменьшается сродство к ионам калия и они
диссоциируют от Na+, К+-АТФ-азы; 6 - АТФ-аза
возвращается в первоначальное состояние.
54. Вторичный активный транспорт
Перенос некоторыхрастворимых веществ против
градиента концентрации
зависит от одновременного
или последовательного
переноса другого вещества по
градиенту концентрации в том
же направлении (активный
симпорт) или в
противоположном (активный
антипорт).
55. Перенос через мембрану макромолекул и частиц: эндоцитоз и экзоцитоз
• Перенос вещества из среды в клетку вместе с частьюплазматической мембраны называют "эндоцитоз". Путем
эндоцитоза (фагоцитоза) клетки могут поглощать большие
частицы, такие как вирусы, бактерии или обломки клеток. Захват
больших частиц осуществляется в основном
специализированными клетками - фагоцитами.
• Поглощение жидкости и растворённых в ней веществ с помощью
небольших пузырьков называют "пиноцитоз". Усвоение веществ
механизмом эндоцитоза (пиноцитоза) характерно для всех
клеток.
56.
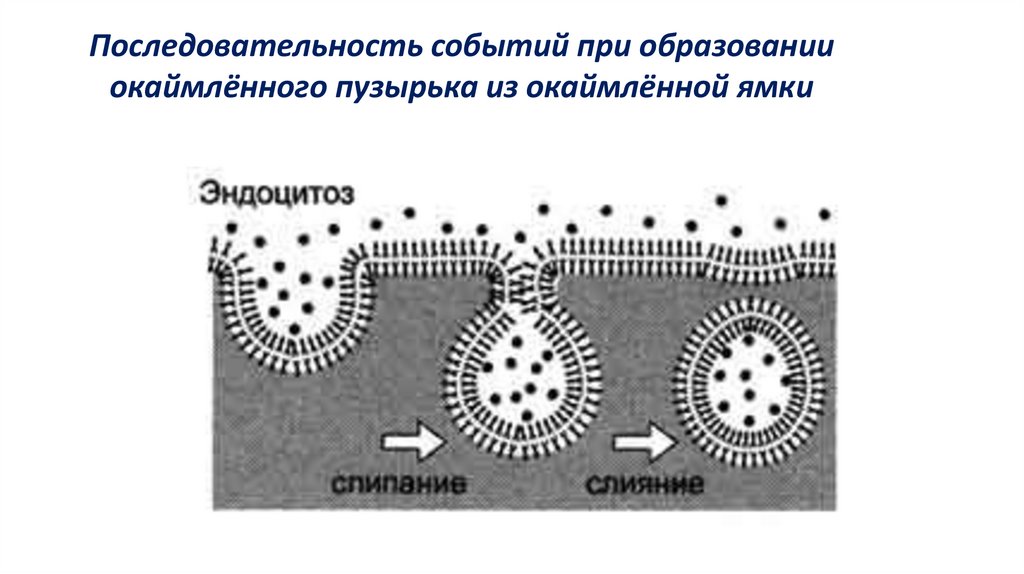
Последовательность событий при образованииокаймлённого пузырька из окаймлённой ямки
57. Положение рецепторов ЛПНП в цитоплазматической мембране
А - положение рецепторовЛПНП в окаймлённой ямке;
Б - положение дефектных
рецепторов ЛПНП вне
окаймлённой ямки.
58. G-белки
•Сигнальные G-белки являютсяуниверсальными посредниками при
передаче гормональных сигналов от
рецепторов клеточной мембраны к
эффекторным белкам , вызывающим
конечный клеточный ответ.
59.
60. Патологии клеточных мембран
• нарушения мембранного транспорта,• изменения проницаемости мембран,
• изменения коммуникации клеток и их "узнавания",
• изменения подвижности мембран и формы
клеток,
• нарушения биогенеза мембран.
61. вывод
• мембрана клетки - это активная изменчивая структура, котораяактивно работает в интересах всей клетки и связывает её с
окружающим миром, а не просто является "защитной
оболочкой". Это - самое важное, что надо знать про клеточную
мембрану.
• В медицине мембранные белки зачастую используются как
“мишени” для лекарственных средств. В качестве таких мишеней
выступают рецепторы, ионные каналы, ферменты, транспортные
системы. В последнее время кроме мембраны мишенью для
лекарственных веществ становятся также гены, спрятанные в
клеточном ядре.