
















")

 Химия
ХимияПохожие презентации:










Нуклеиновые кислоты. Схема образования пуринового нуклеозида
1.
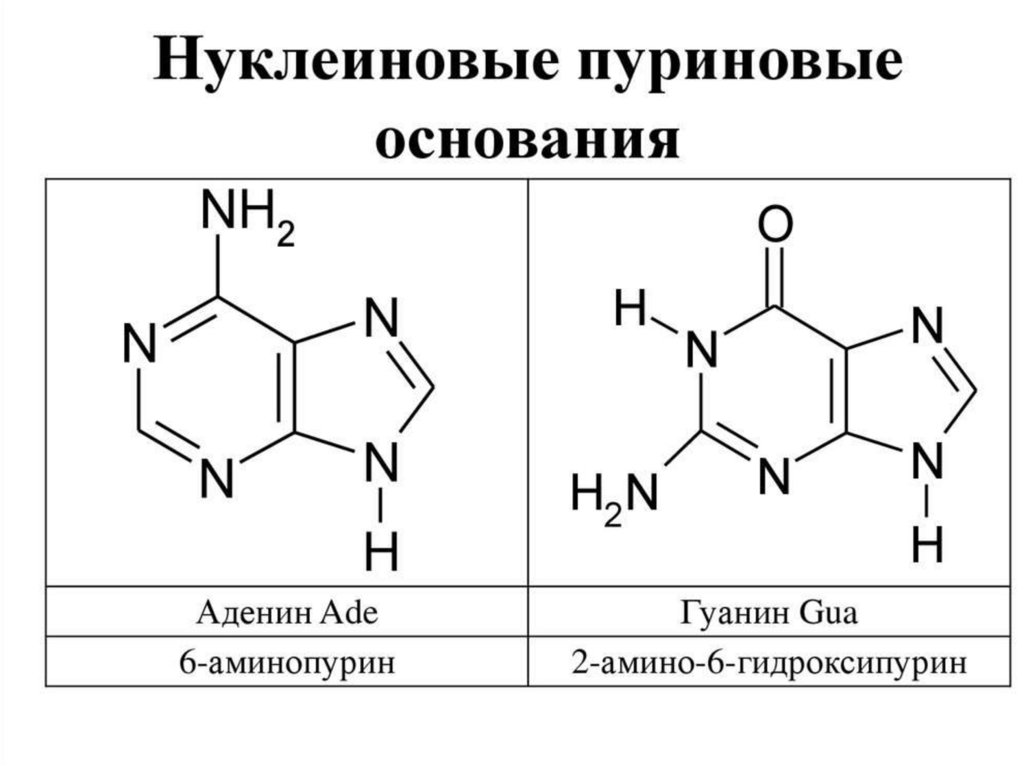
НУКЛЕИНОВЫЕ КИСЛОТЫ2.
3.
4. Углеводы нк
5.
6.
7.
8.
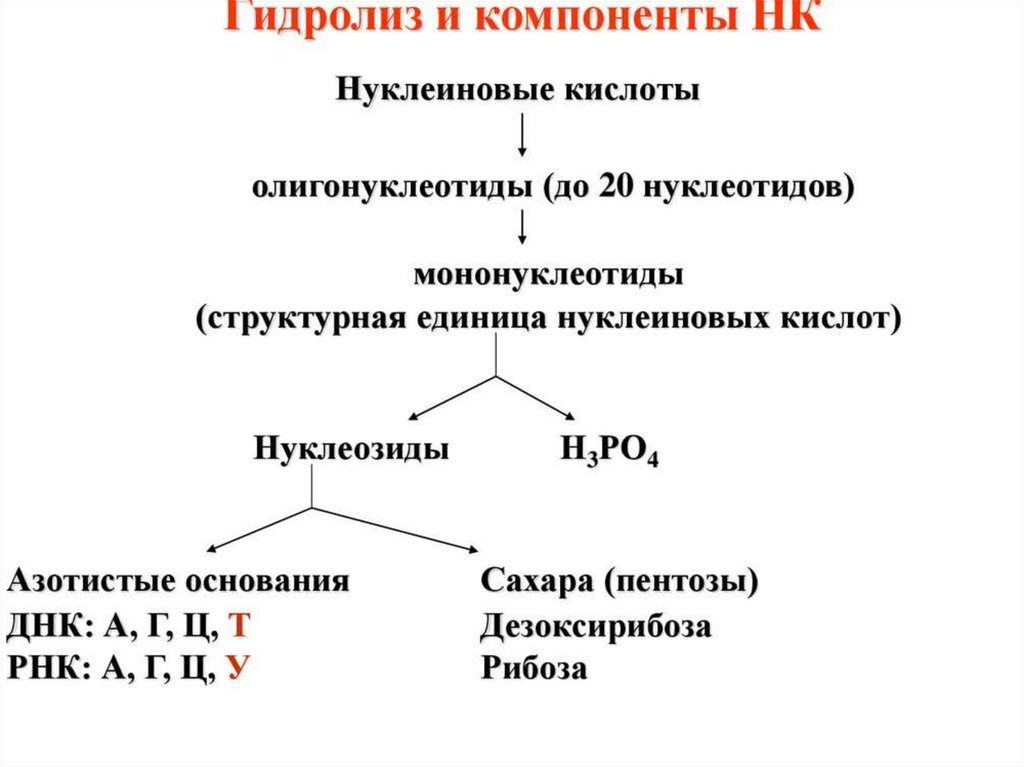
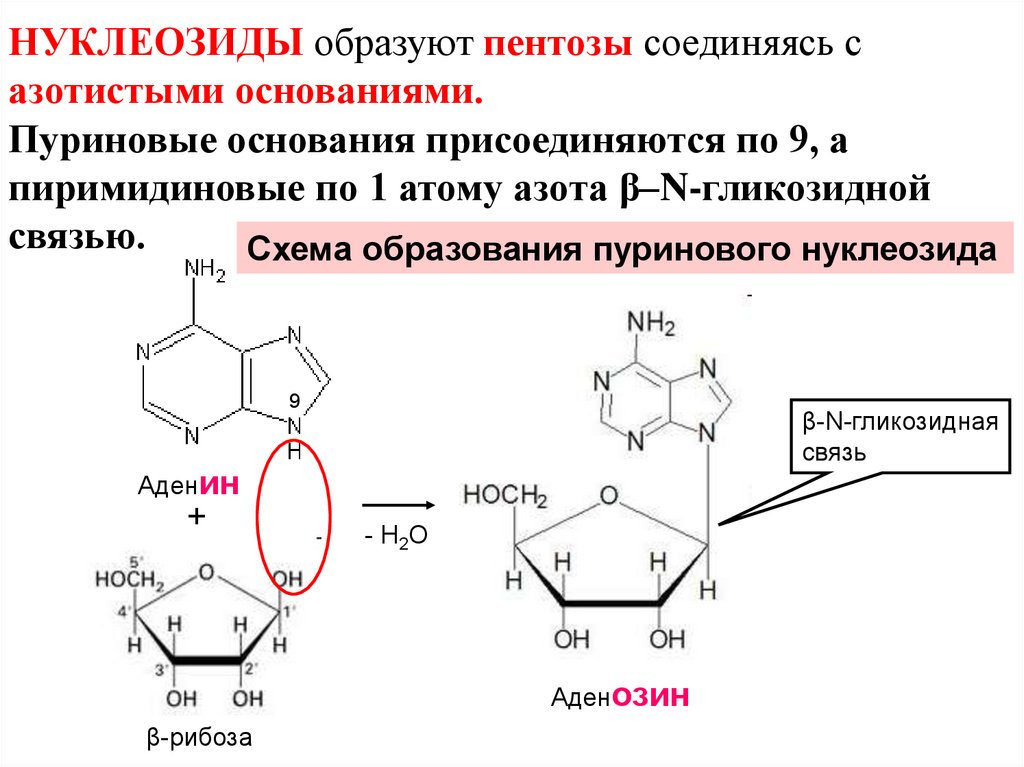
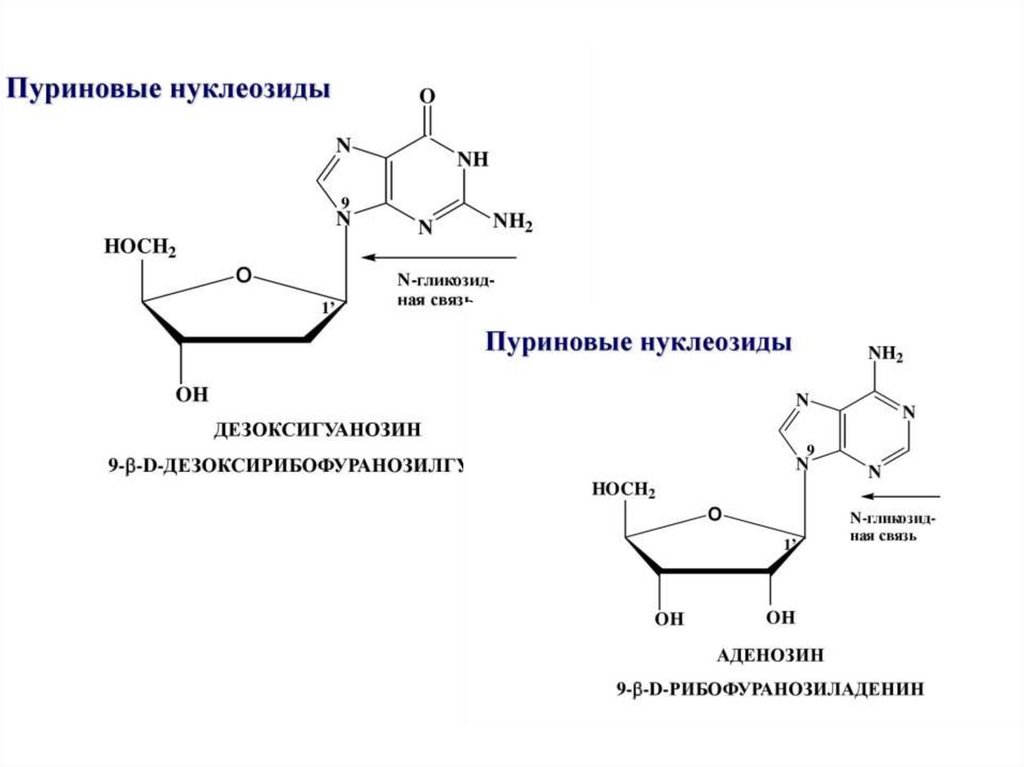
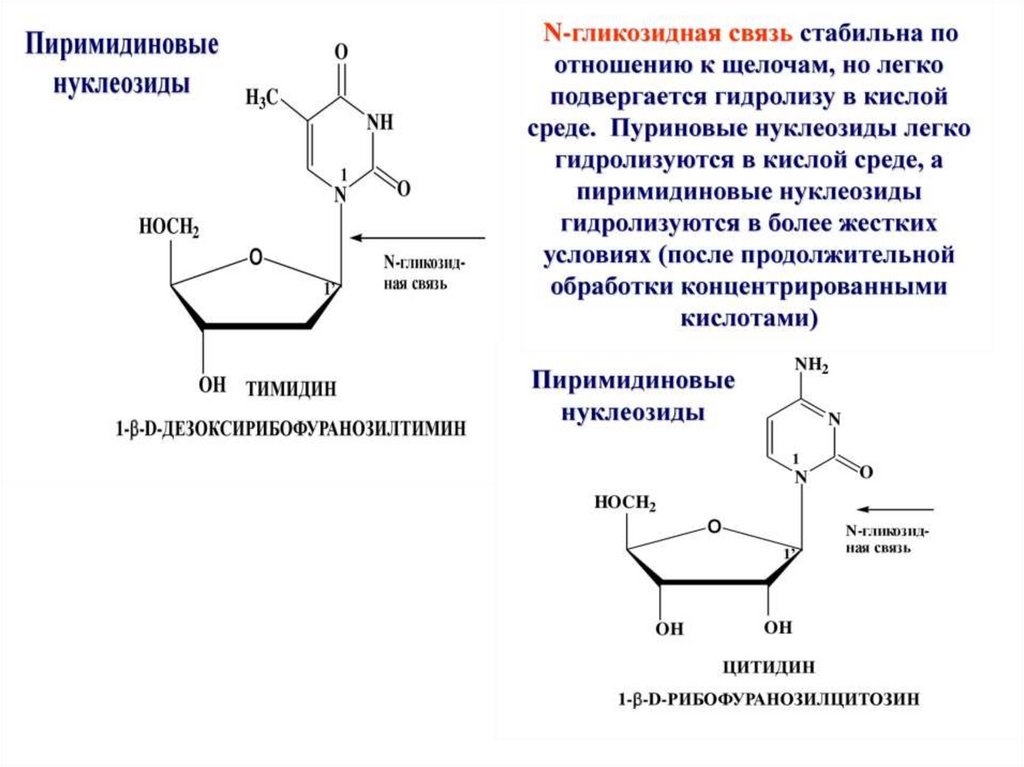
НУКЛЕОЗИДЫ образуют пентозы соединяясь сазотистыми основаниями.
Пуриновые основания присоединяются по 9, а
пиримидиновые по 1 атому азота β–N-гликозидной
связью.
Схема образования пуринового нуклеозида
9
β-N-гликозидная
связь
Аденин
+
- Н2О
Аденозин
β-рибоза
9.
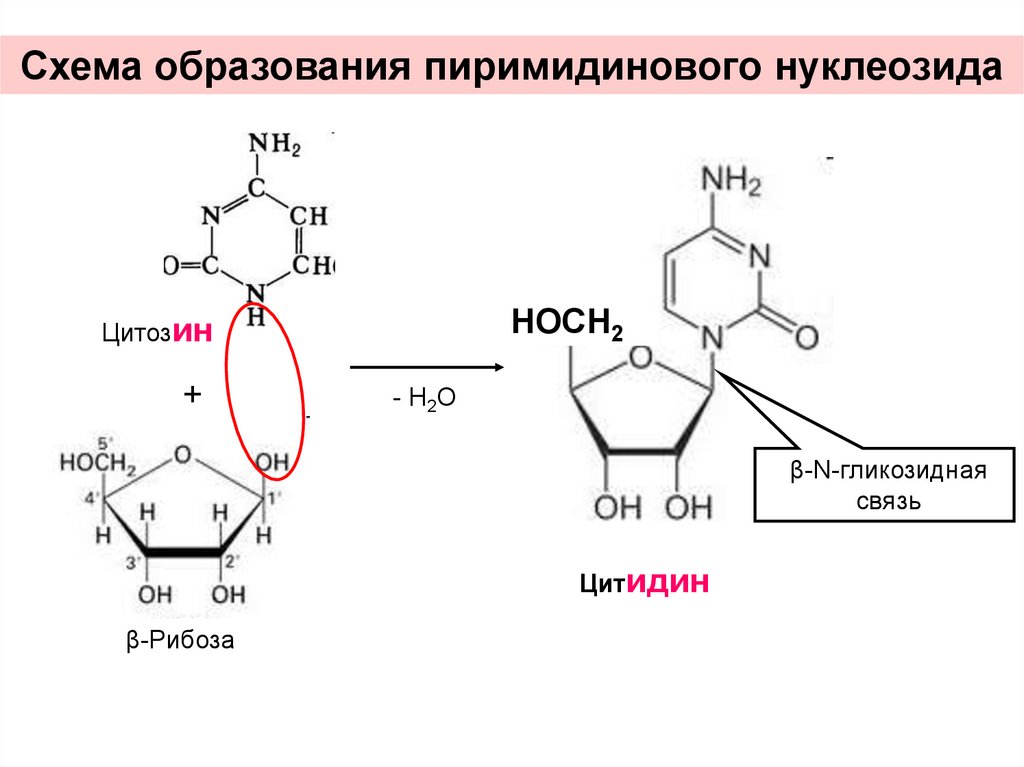
Схема образования пиримидинового нуклеозидаНОСН2
Цитозин
+
- Н2О
β-N-гликозидная
связь
Цитидин
β-Рибоза
10.
11.
12.
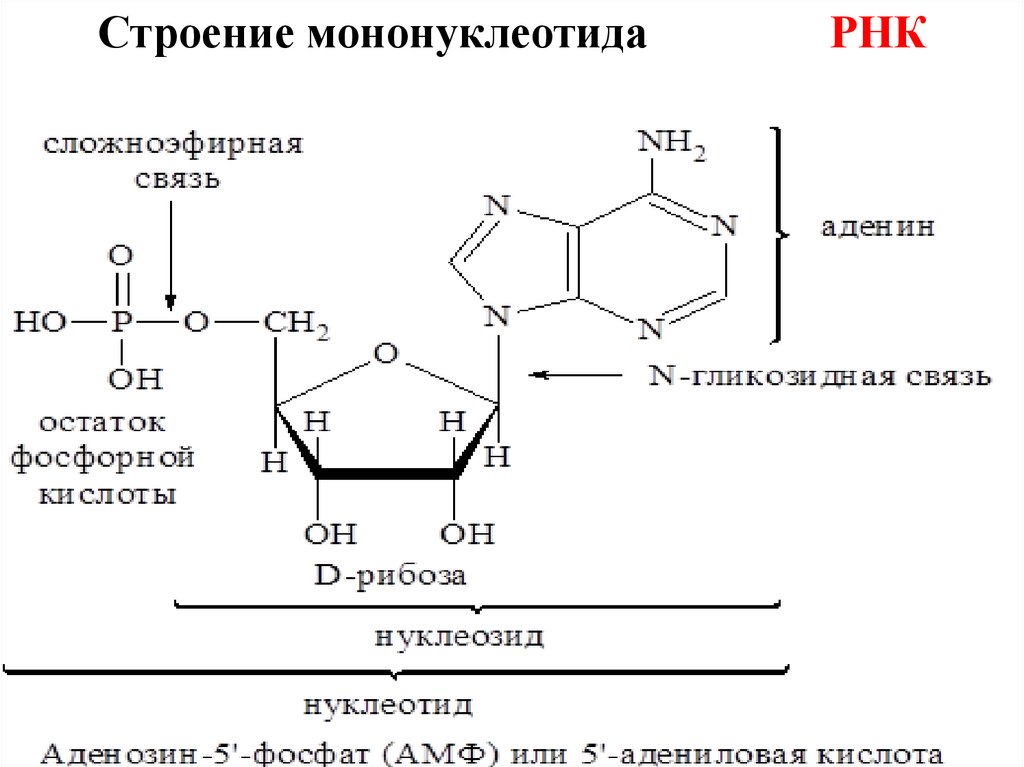
Строение мононуклеотидаРНК
(Н3РО4 присоединяется по 5-ому атому пентозы)
13.
РНК14.
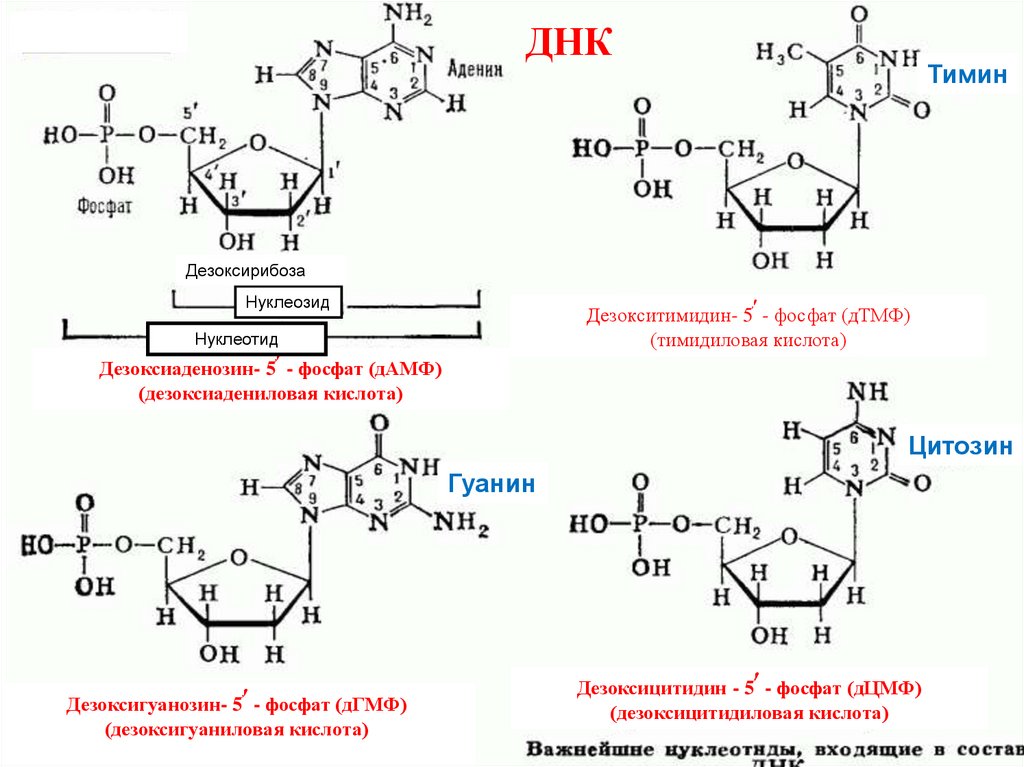
ДНКАденин
Тимин
Дезоксирибоза
׳
Нуклеозид
Дезокситимидин- 5҆ - фосфат (дТМФ)
(тимидиловая кислота)
Нуклеотид
׳
Дезоксиаденозин- 5҆ - фосфат (дАМФ)
(дезоксиадениловая кислота)
Цитозин
Гуанин
׳
Дезоксигуанозин- 5҆ - фосфат (дГМФ)
(дезоксигуаниловая кислота)
׳
Дезоксицитидин - 5҆ - фосфат (дЦМФ)
(дезоксицитидиловая кислота)
15.
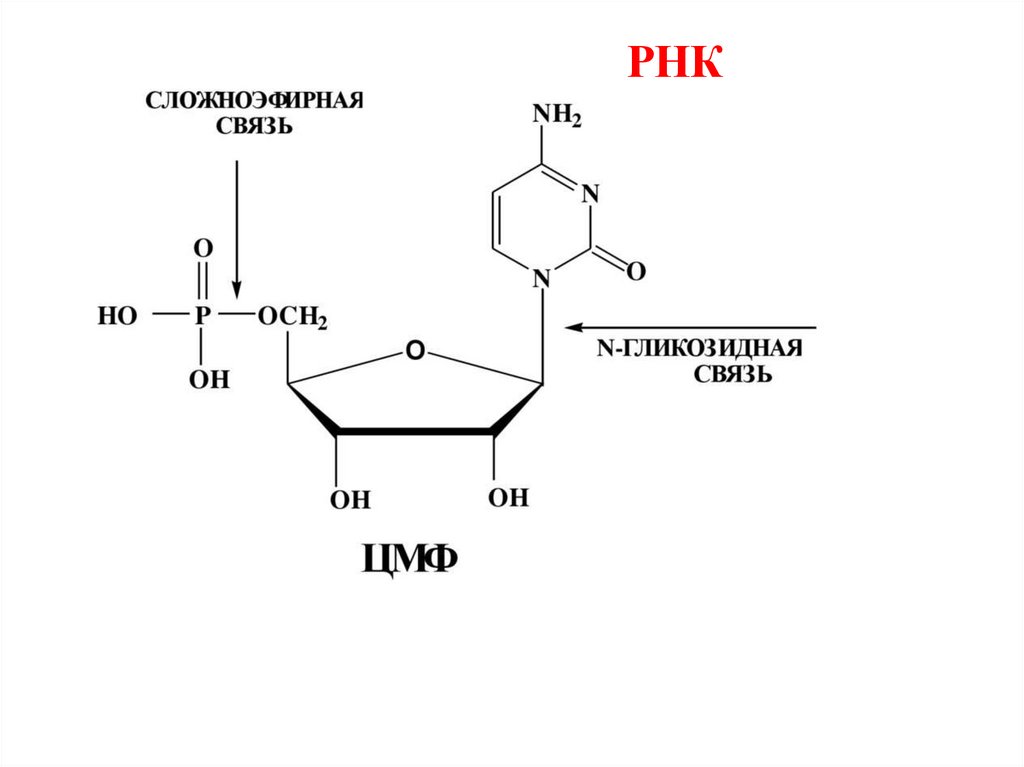
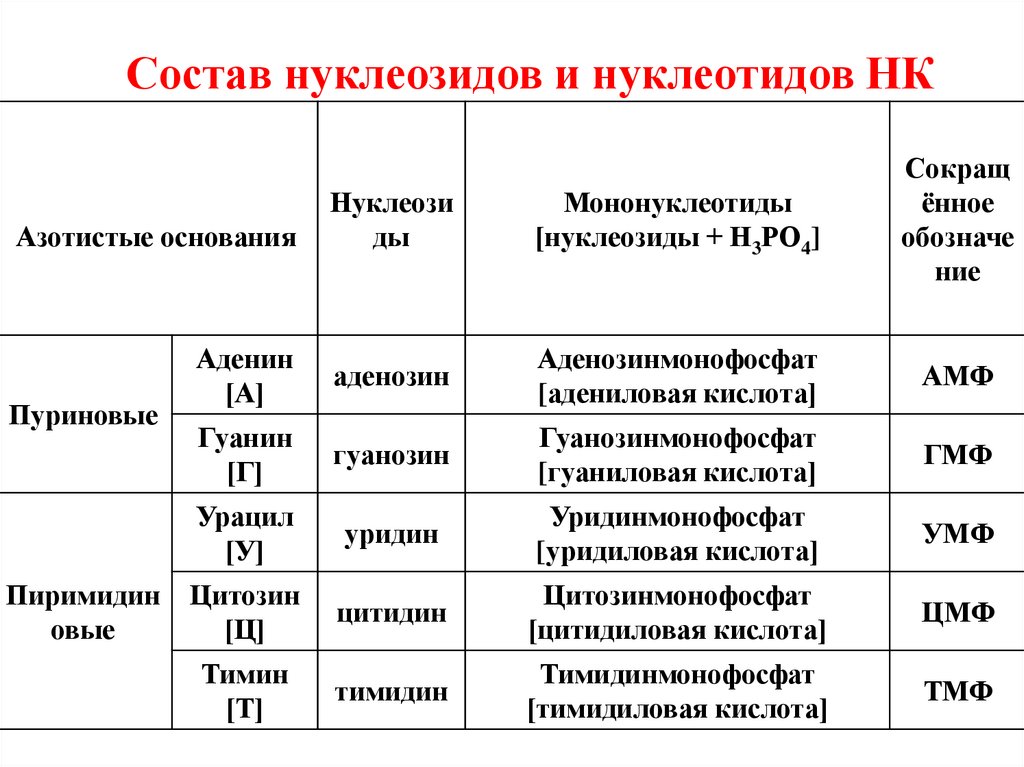
Состав нуклеозидов и нуклеотидов НКНуклеози
ды
Мононуклеотиды
[нуклеозиды + Н3РО4]
Сокращ
ённое
обозначе
ние
Аденин
[А]
аденозин
Аденозинмонофосфат
[адениловая кислота]
АМФ
Гуанин
[Г]
гуанозин
Гуанозинмонофосфат
[гуаниловая кислота]
ГМФ
Урацил
[У]
уридин
Уридинмонофосфат
[уридиловая кислота]
УМФ
Цитозин
[Ц]
цитидин
Цитозинмонофосфат
[цитидиловая кислота]
ЦМФ
Тимин
[Т]
тимидин
Тимидинмонофосфат
[тимидиловая кислота]
ТМФ
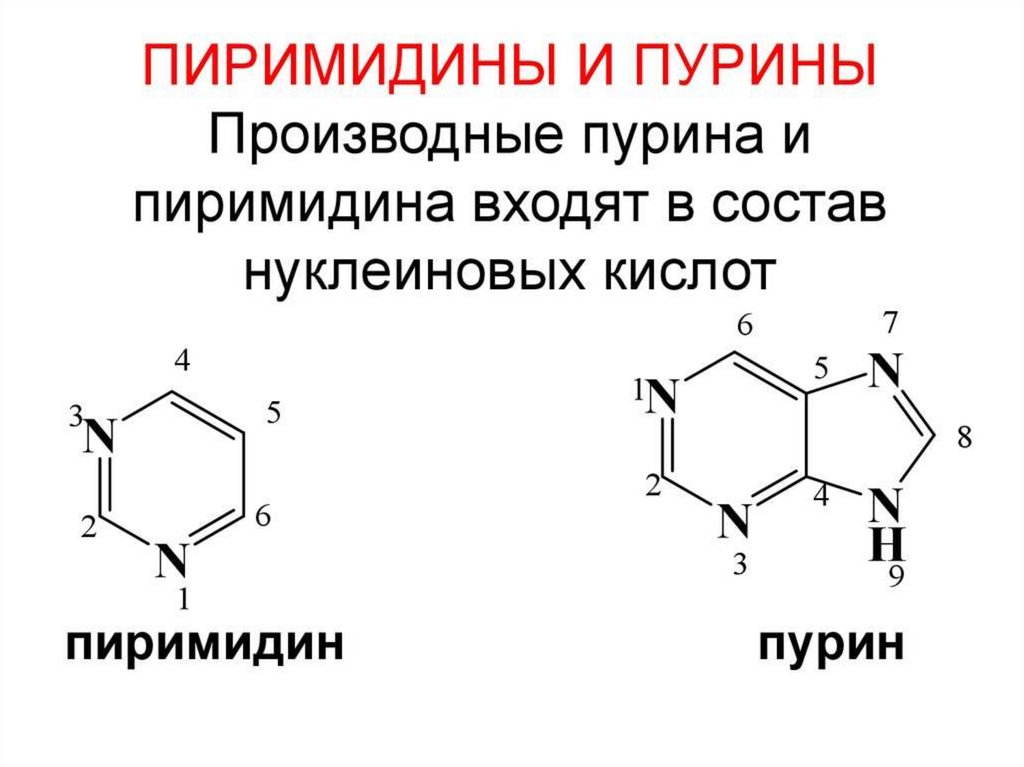
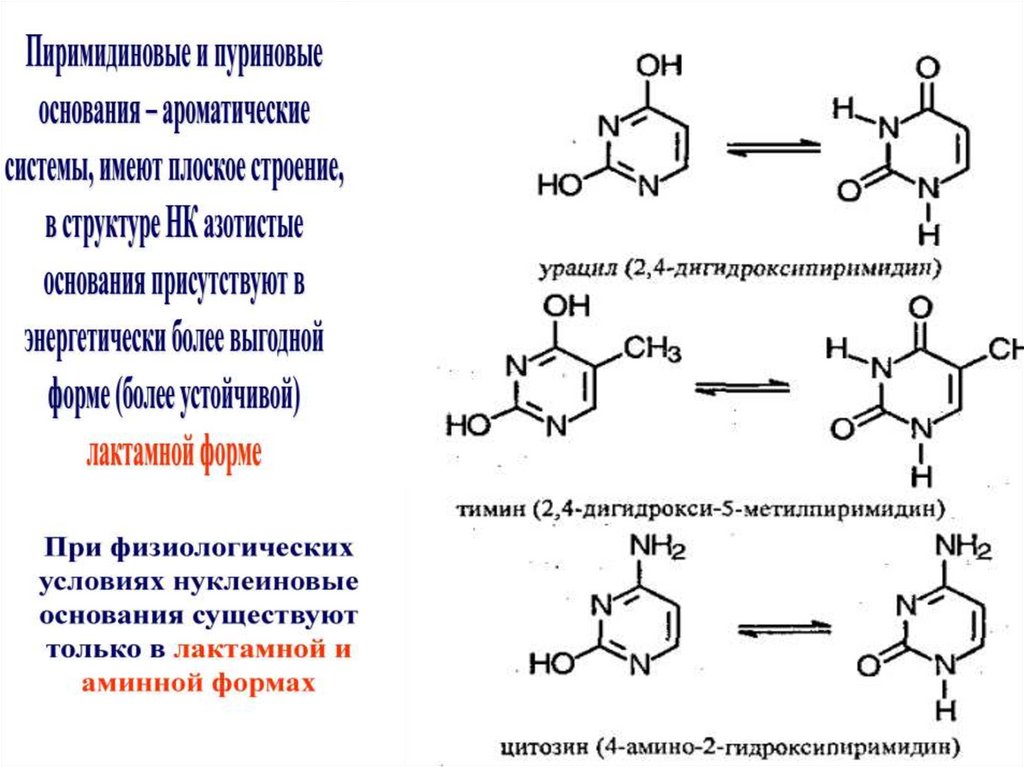
Азотистые основания
Пуриновые
Пиримидин
овые
16.
17.
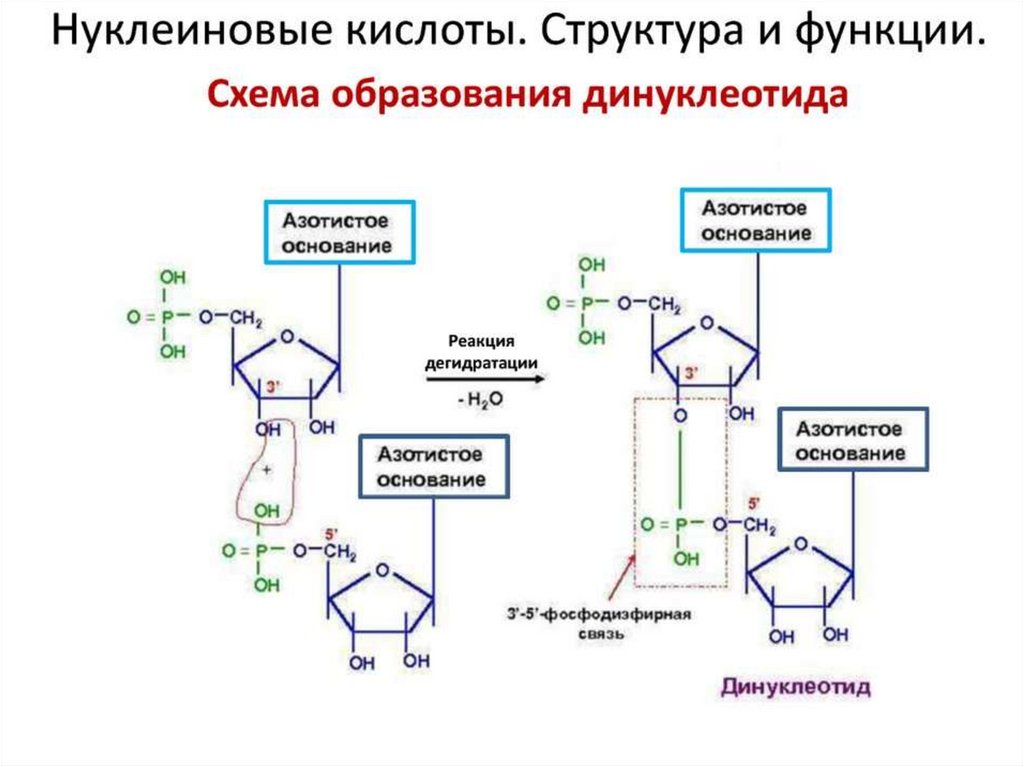
Первичнаяструктура
нуклеиновых
кислот - порядок
расположения
мононуклеотидов в
полинуклеотидной
цепи ДНК и РНК:
связи 3’,5’-фосфодиэфирные
.
Известна первичная
структура почти всех
т-РНК, ряда молекул
р-РНК.
18.
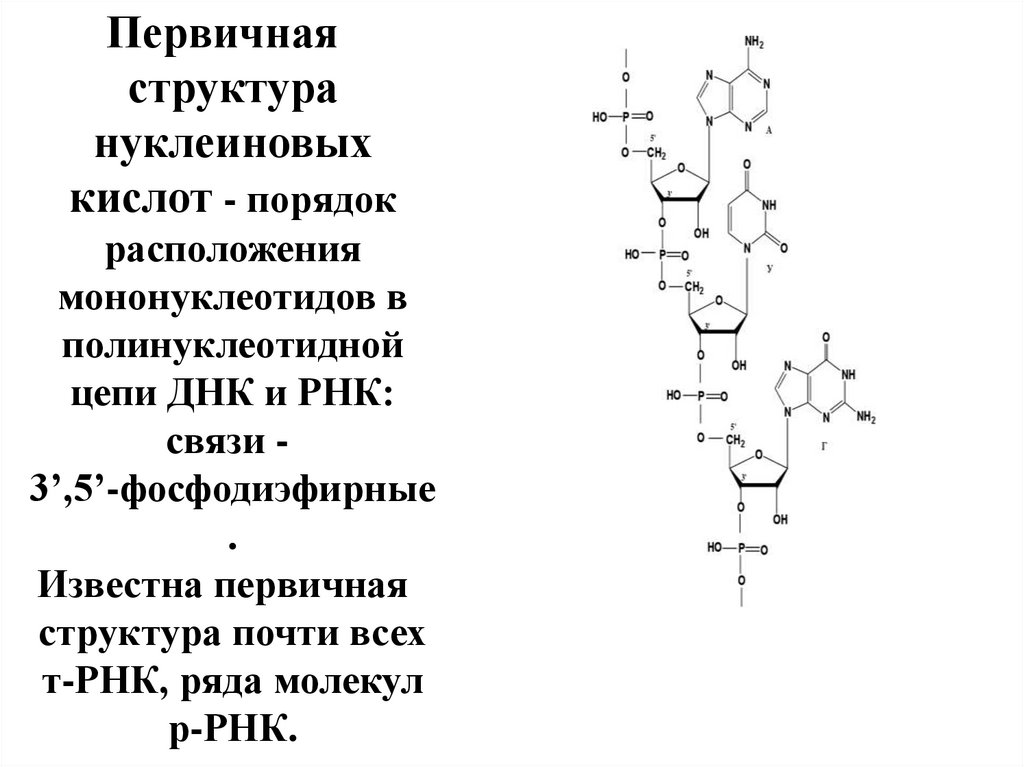
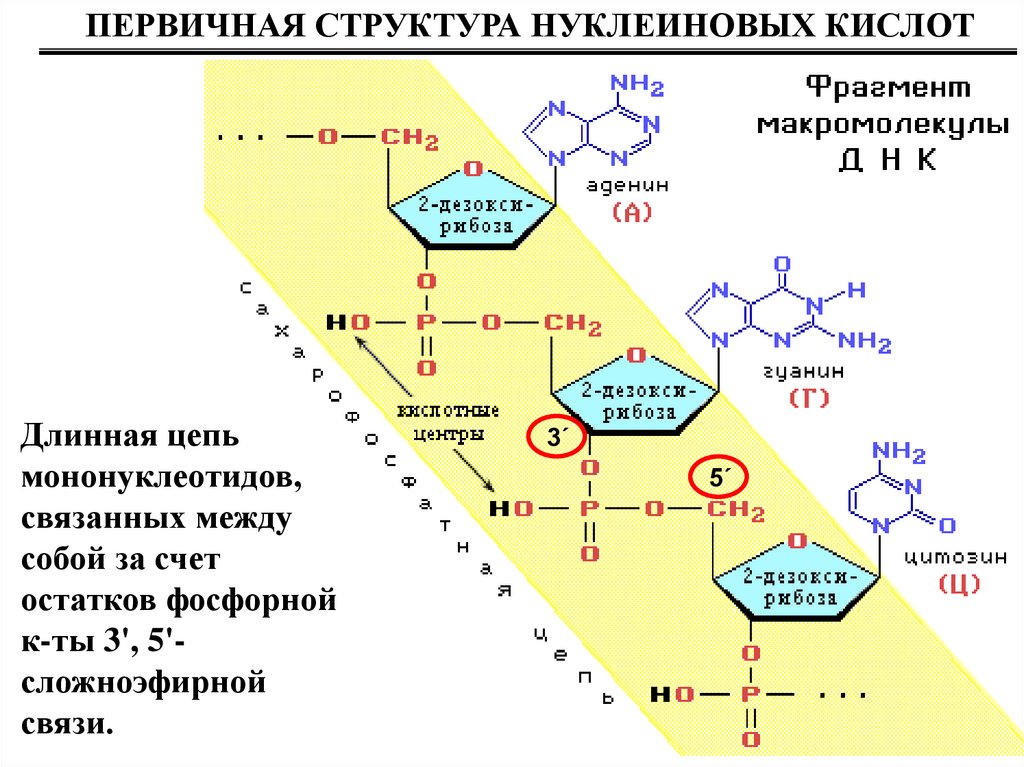
ПЕРВИЧНАЯ СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТДлинная цепь
мононуклеотидов,
связанных между
собой за счет
остатков фосфорной
к-ты 3', 5'сложноэфирной
связи.
3´
5´
19.
ДНК20. Структура ДНК
Геном человека - нуклеотиднаяпоследовательность всей геномной
ДНК человека, а также
идентификация генов и их
локализация в геноме.
Три миллиарда пар оснований могут быть
записаны на одном компакт-диске.
21.
22.
23.
Вторичная структура ДНКВторичная структура ДНК – это двухцепочечная
правозакрученная спираль из комплементарных друг другу
антипараллельных полинуклеотидных нитей. Молекула ДНК
состоит из 2-х цепей, закрученных в правовращающуюся
спираль вокруг одной и той же оси. Азотистые основания
находятся внутри, а фосфорные и углеводные компоненты –
снаружи.
Диаметр спирали 1,8 нм.
Шаг спирали 3,4 нм и содержит 10 пар оснований.
Полинуклеотидные цепи ориентированы в противоположном
направлении (антипараллельны). На одном конце спирали одна
цепь имеет 5/ - конец, другая 3/ -конец.
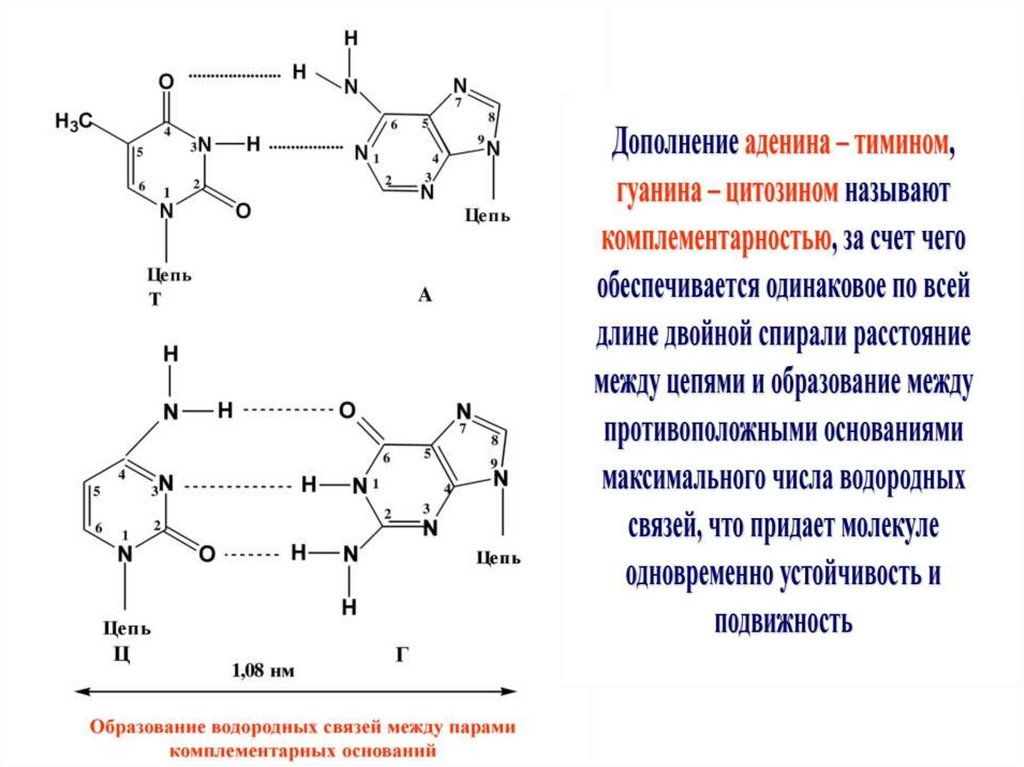
Азотистые основания в молекуле ДНК расположены
по принципу комплементарности: А взаимодействует
только с Т, Г с Ц,
24.
25.
идрофобныезаимодействия между
роматическими
снованиями
табилизируют структуру.
статки дезоксирибозы
вух антипараллельных
епей образуют
равозакрученную
пираль, где на каждый
иток приходится 10 пар
снований. Внешняя
торона спирали заряжена
трицательно за счет
осфатных групп.
26.
27.
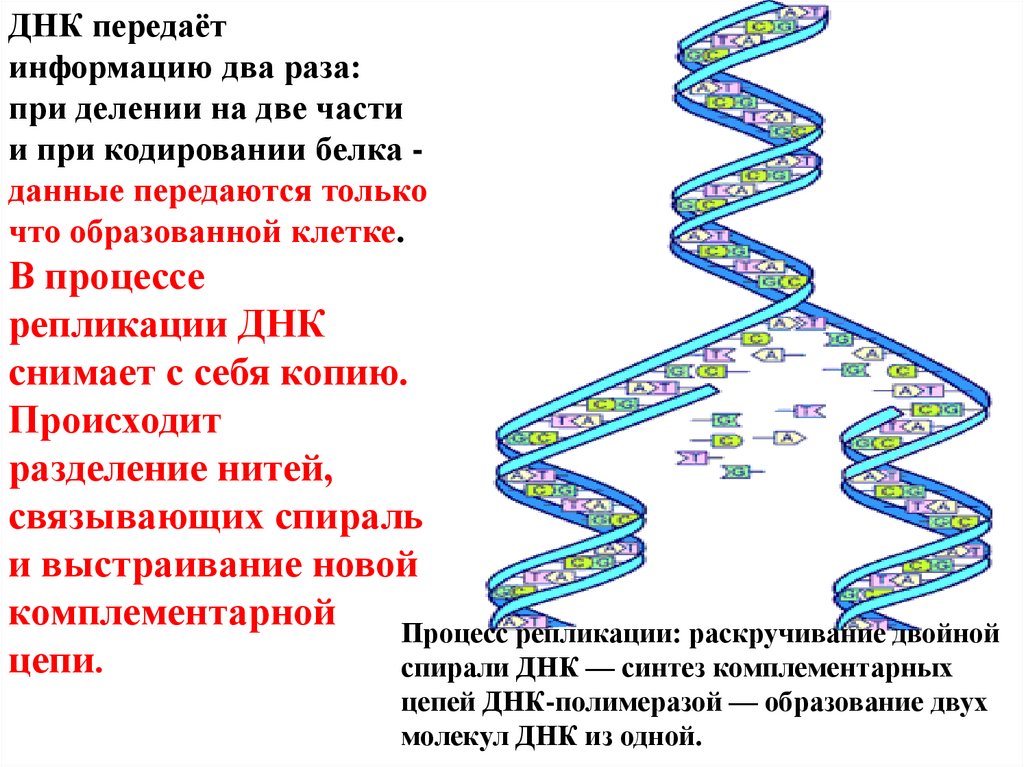
ДНК передаётинформацию два раза:
при делении на две части
и при кодировании белка данные передаются только
что образованной клетке.
В процессе
репликации ДНК
снимает с себя копию.
Происходит
разделение нитей,
связывающих спираль
и выстраивание новой
комплементарной
Процесс репликации: раскручивание двойной
цепи.
спирали ДНК — синтез комплементарных
цепей ДНК-полимеразой — образование двух
молекул ДНК из одной.
28.
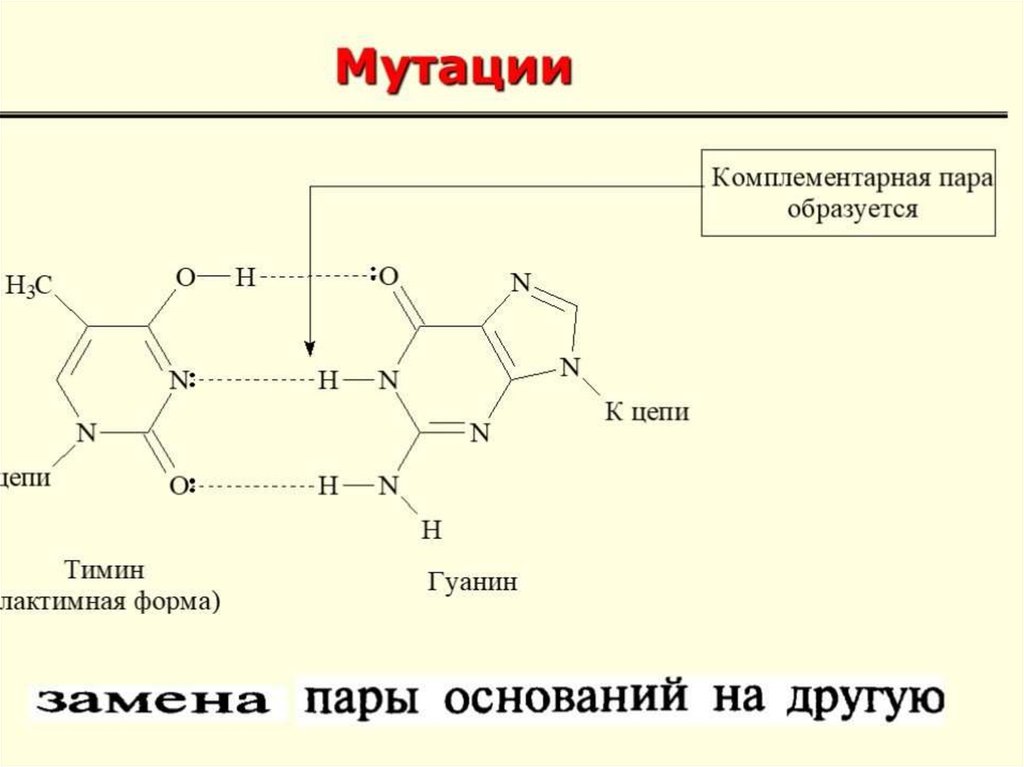
Мута́ция— стойкое изменение геномаВ основе серповидно-клеточной анемии лежит генная
мутация, обусловливающая синтез аномального гемоглобина
S (HbS). Причина: замена глутаминовой кислоты валином в
ß-полипептидной цепи. Гемоглобин S после потери
присоединенного кислорода приобретает консистенцию
высокополимерного геля и становится в 100 раз менее
растворимым, чем нормальный гемоглобин А.
29.
30.
Третичная структура ДНК - двойная спираль ДНК нанекоторых участках может подвергаться дальнейшей
спирализации с образованием суперспирали или
открытой кольцевой формы.
Суперспирализация обеспечивает экономную упаковку —
вместо 8 см длины огромная молекула ДНК занимает 5 нм.
Суперспирали соединяются с белками – гистонами
(положительно заряженными), обеспечивающими стабильность
ДНК.
Хромосомы образуют компактные структуры только в фазу
деления. В период покоя комплексы ДНК с белками
распределены равномерно по объему ядра, образуя
хроматин. Белки хроматина включают две группы: гистоны и
негистоновые белки. В состав хроматина входят ДНК и
гистоны — белки с высоким содержанием лизина и аргинина.
Предполагают, что аминогруппы радикалов этих аминокислот
взаимодействуют с кислотными группами ДНК.
31.
32.
33.
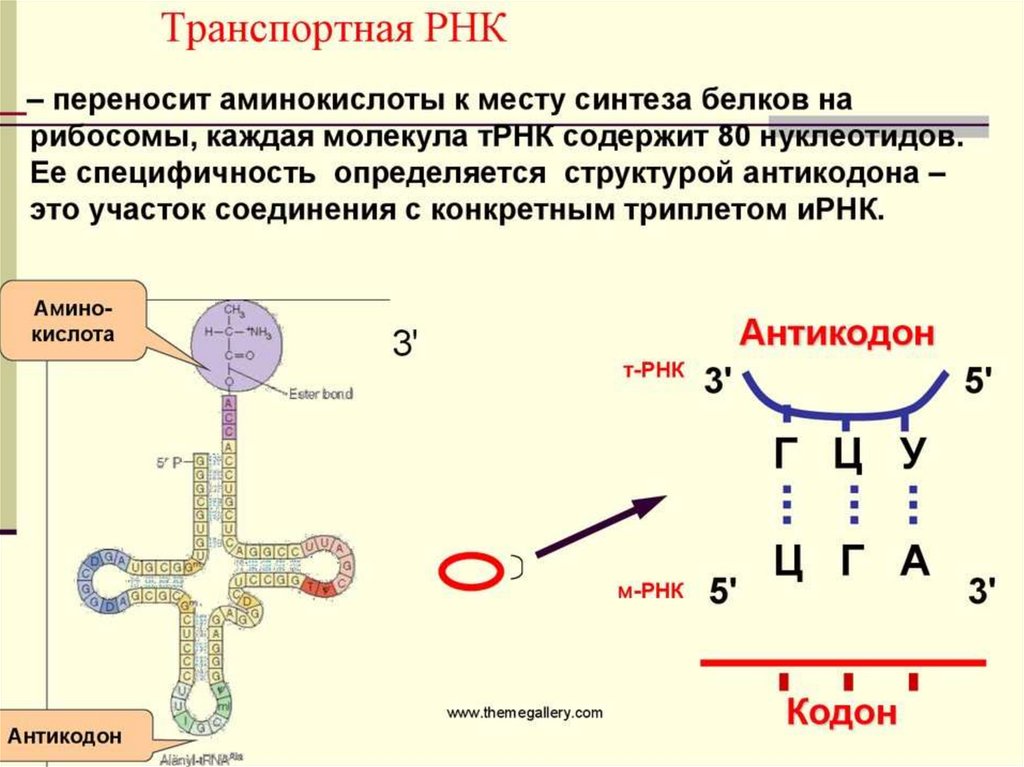
Рибосомные РНК (рРНК) Составляют около80% всей РНК клетки и входят в состав
рибосом. рРНК выполняет структурную и
ферментативную функции, обеспечивают
правильное взаимодействие рибосом с
мРНК и тРНК. Рибосомы состоят из двух
субъединиц: большой и малой. Функциясинтез белка в клетке
34.
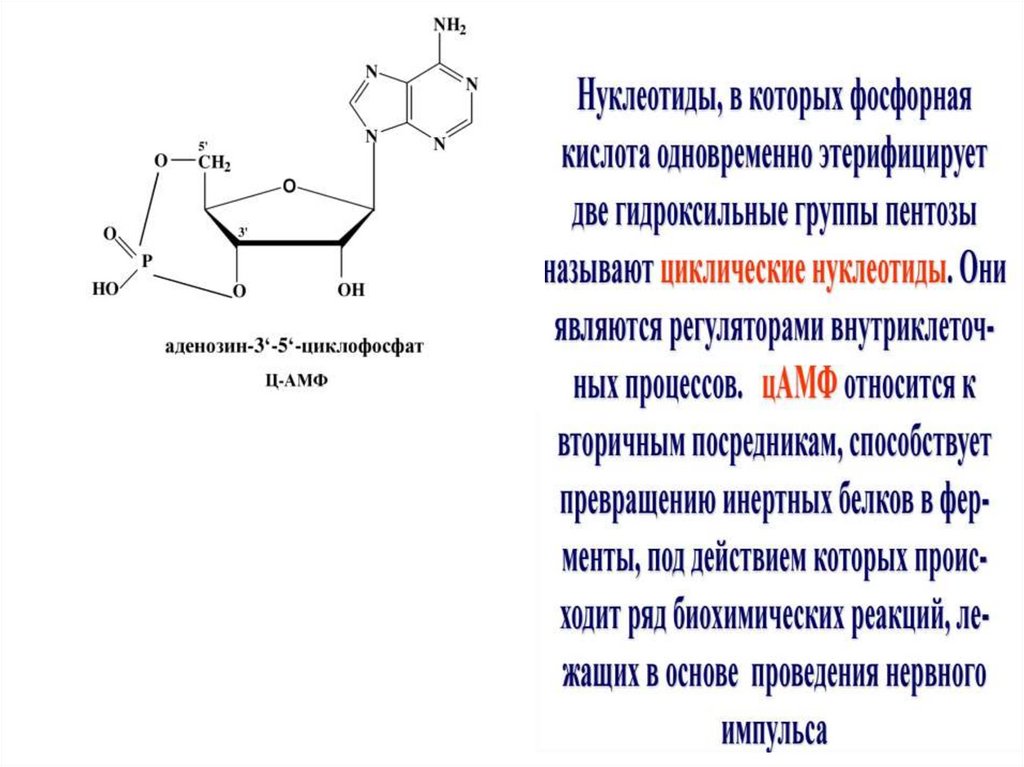
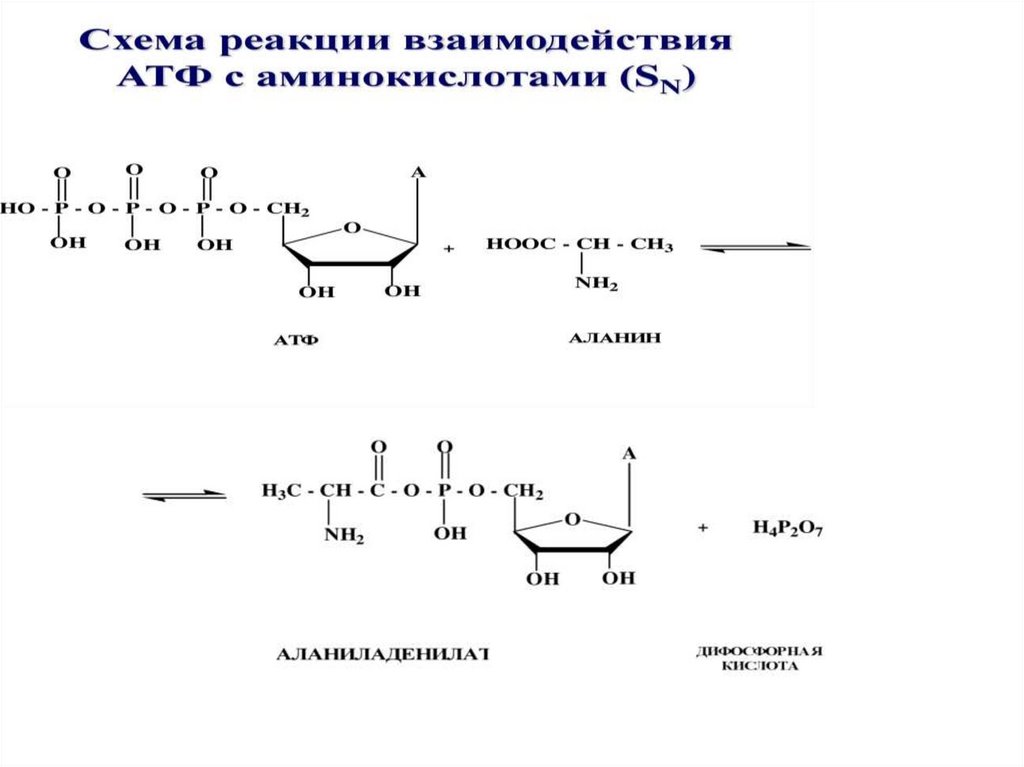
35. Другие биологически активные формы нуклеотидов
1. Цикломонофосфаты (цАМФ, цГМФ)передают сигнал внутрь клетки2. Полифосфаты (аденозинтрифосфат АТФ,
ГТФ, УТФ, ЦТФ ) – макроэргические
соединения, которые обладают высокой
энергией (активностью)
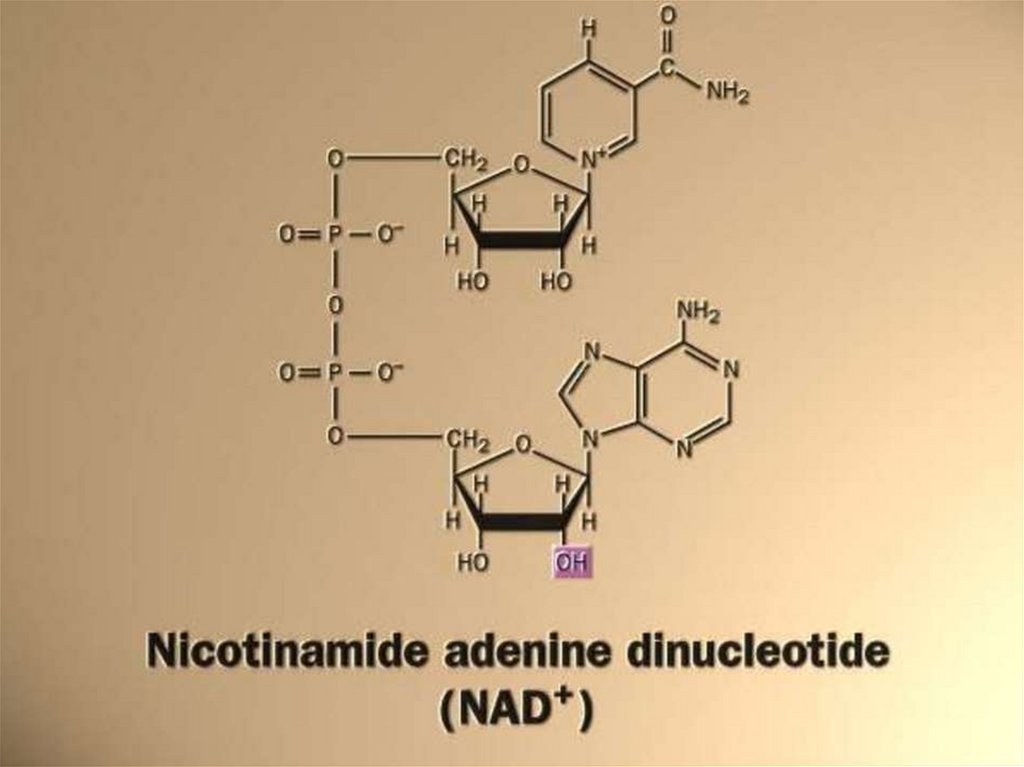
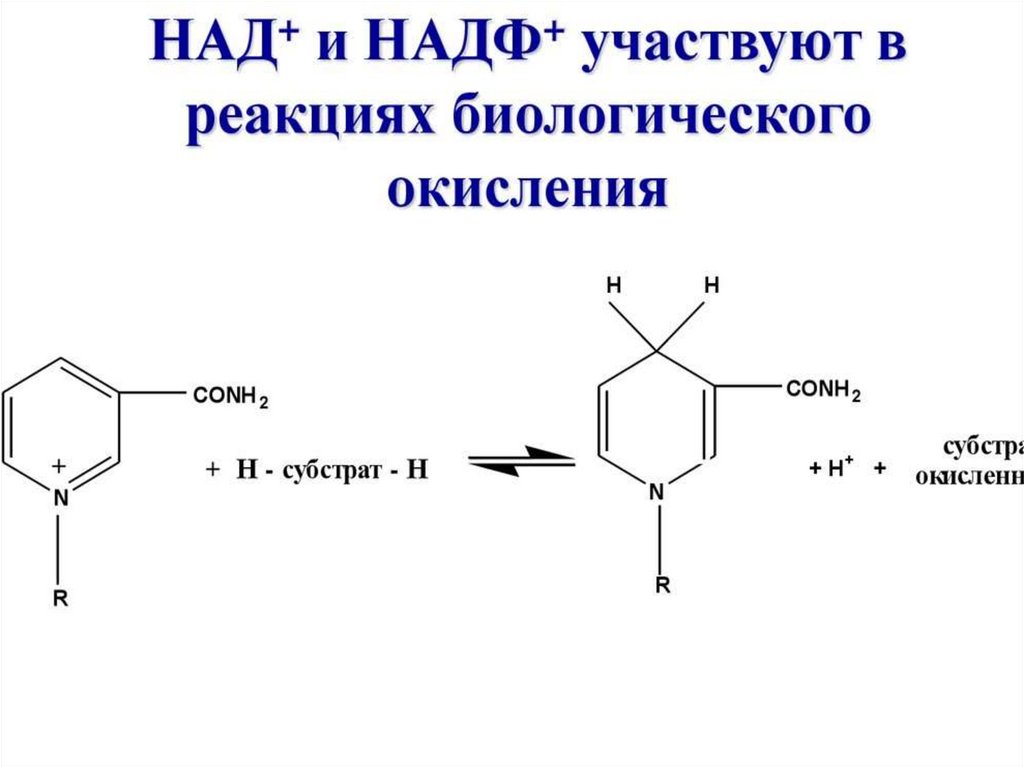
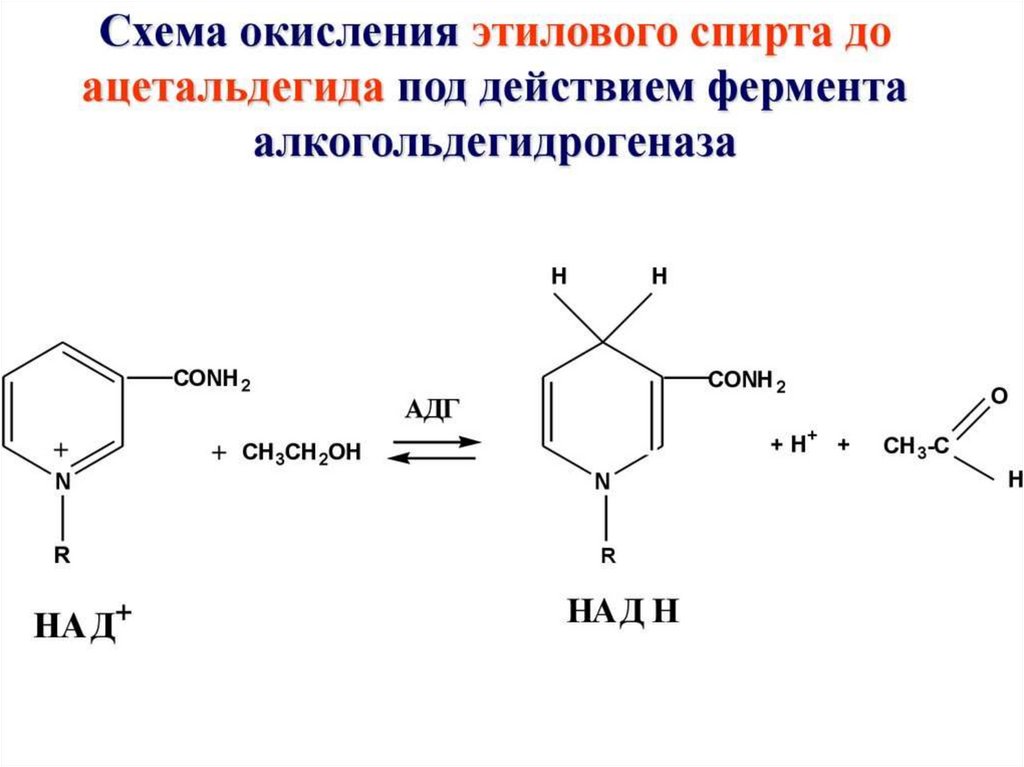
3. Динуклеотиды (НАД, ФАД)- коферменты
ферментов дегидрогеназ
36.
37.
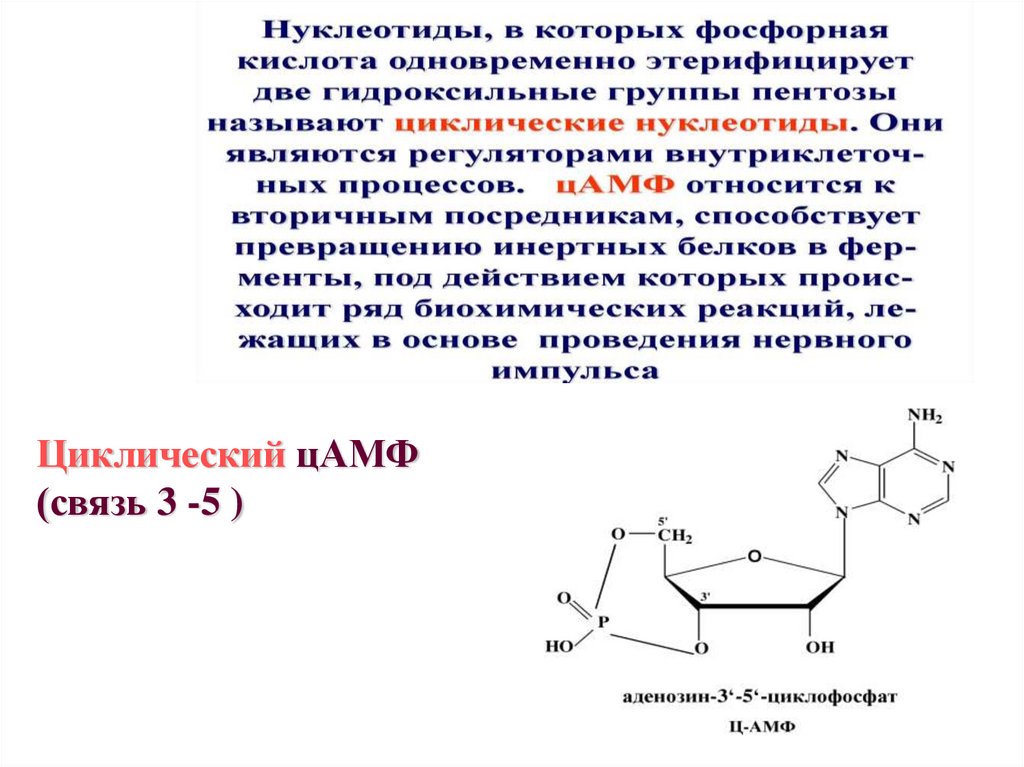
•цЦиклический цАМФ
(связь 3 -5 )
38.
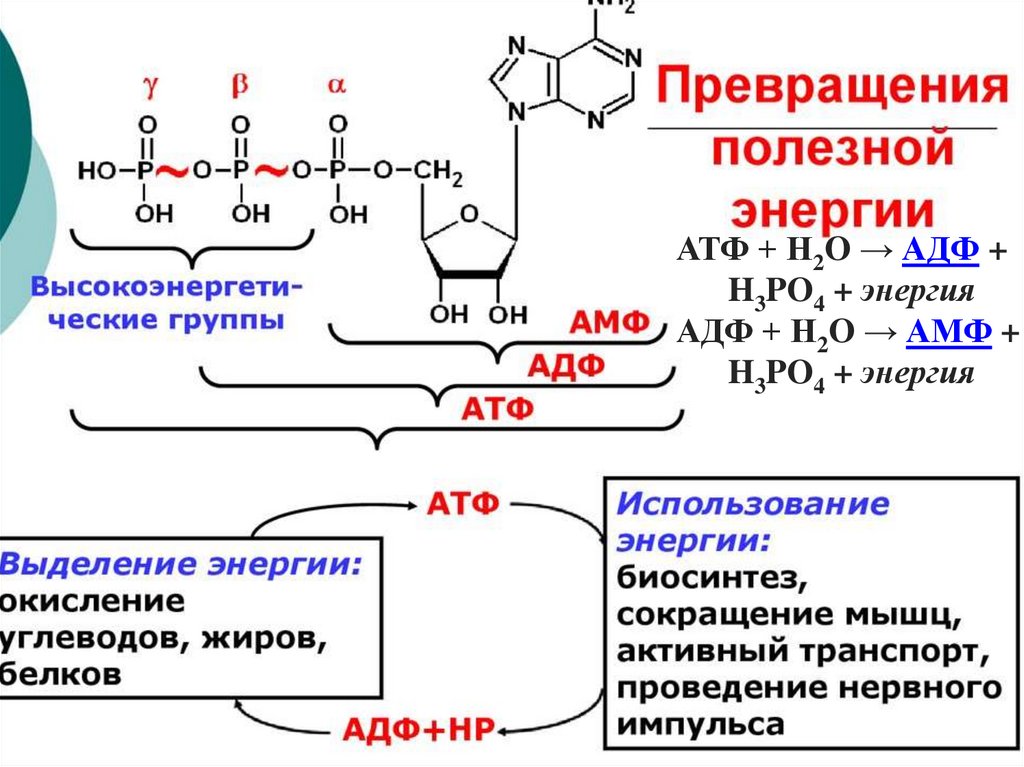
АТФ + H2O → АДФ +H3PO4 + энергия
АДФ + H2O → АМФ +
H3PO4 + энергия
39.
40.
41.
42.
43.
44.
45.
46.
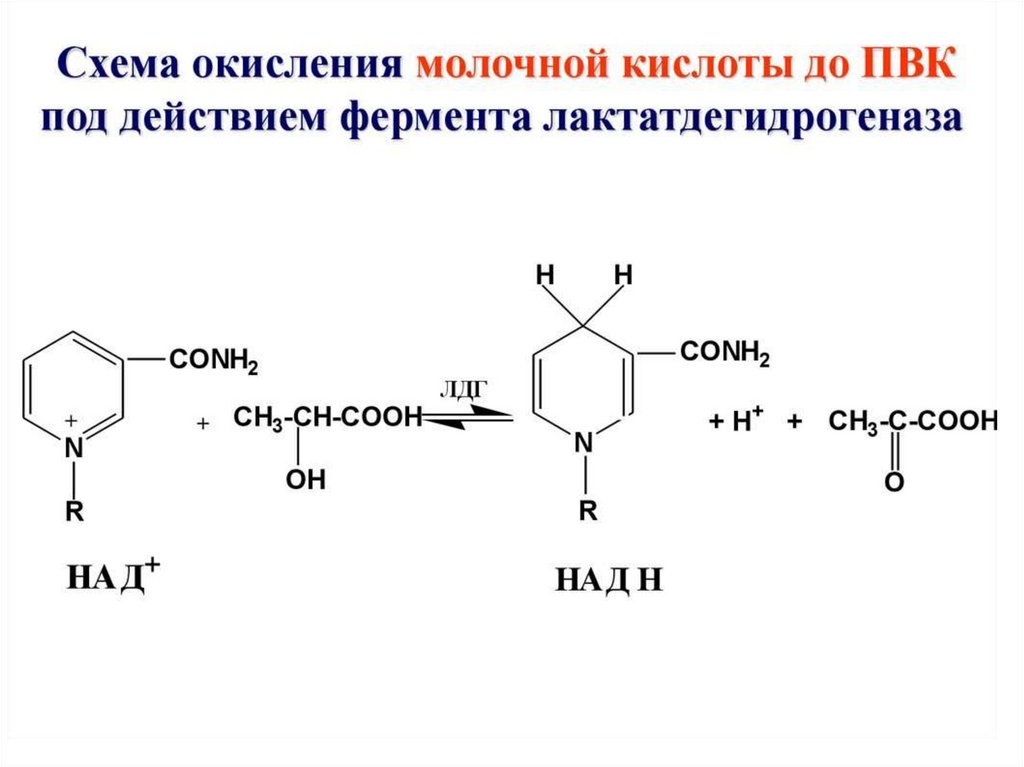
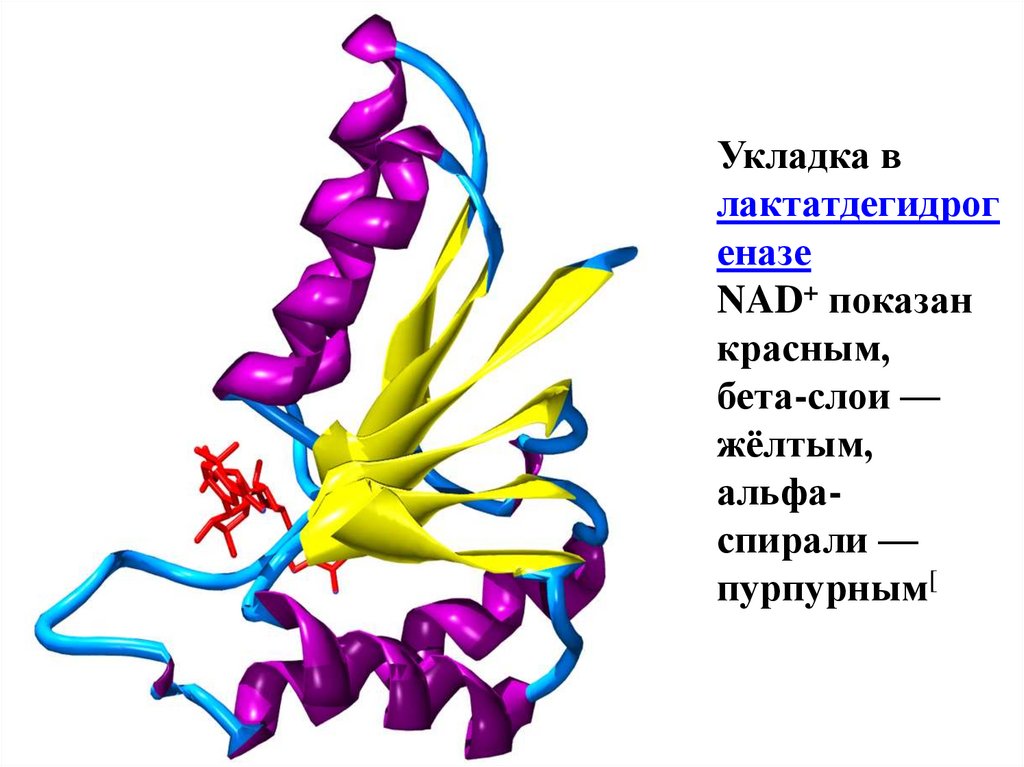
Укладка влактатдегидрог
еназе
NAD+ показан
красным,
бета-слои —
жёлтым,
альфаспирали —
пурпурным[
47.
48. ФАД(флавинадениндинуклеотид)
49.
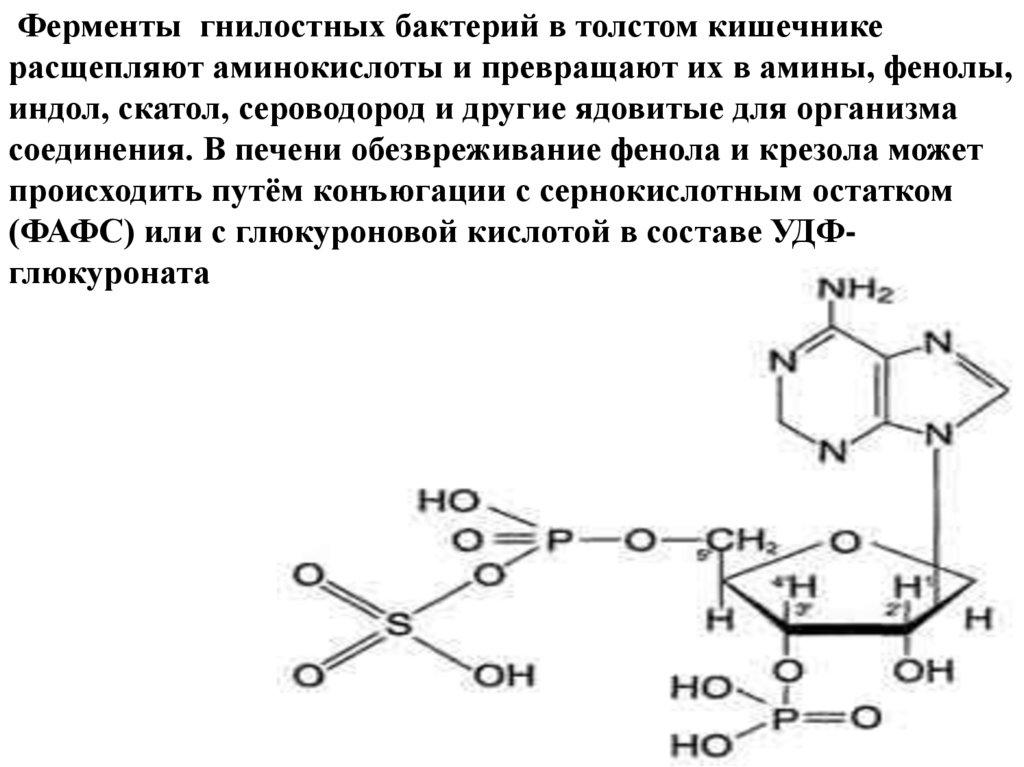
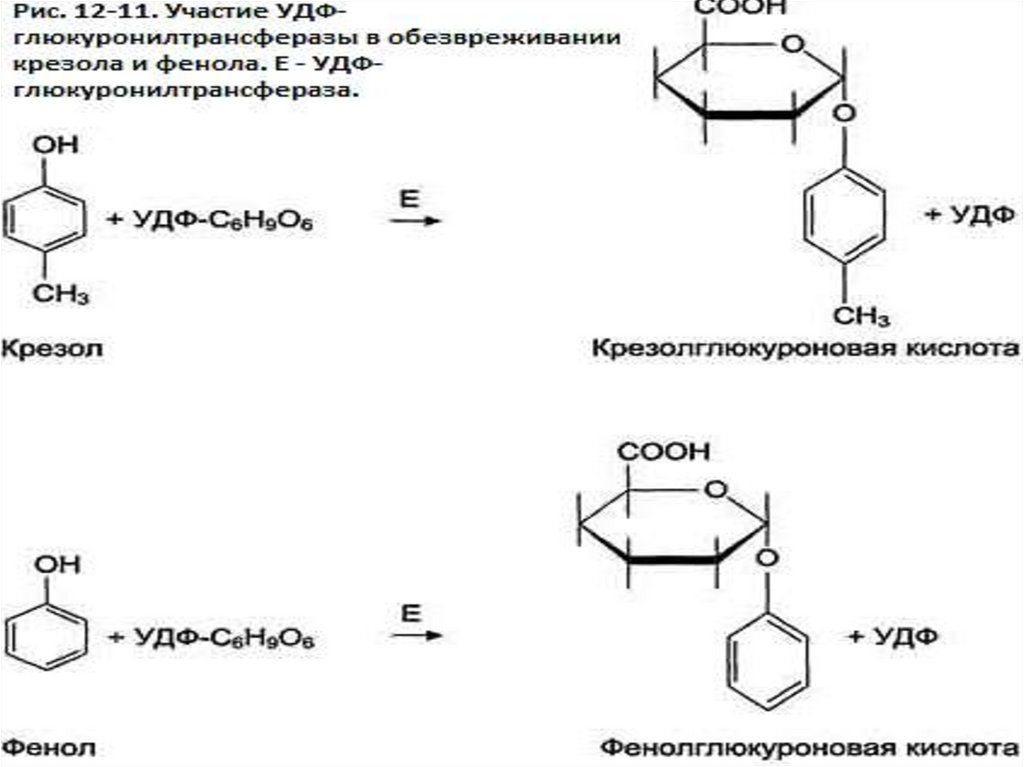
Ферменты гнилостных бактерий в толстом кишечникерасщепляют аминокислоты и превращают их в амины, фенолы,
индол, скатол, сероводород и другие ядовитые для организма
соединения. В печени обезвреживание фенола и крезола может
происходить путём конъюгации с сернокислотным остатком
(ФАФС) или с глюкуроновой кислотой в составе УДФглюкуроната
50.
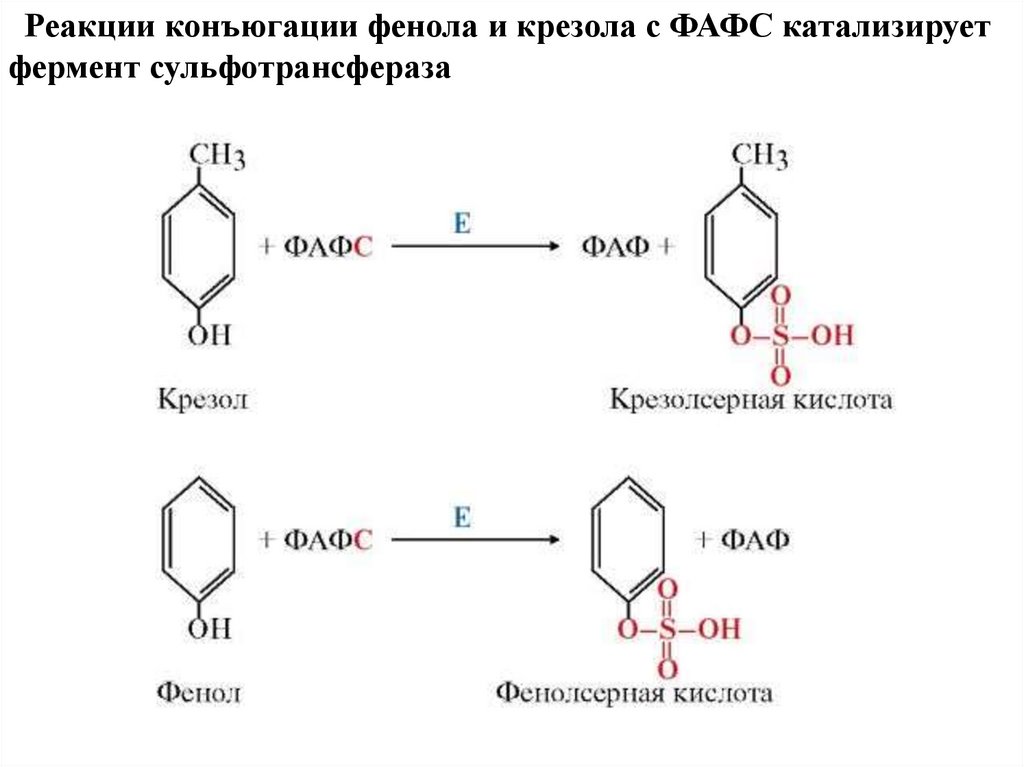
Реакции конъюгации фенола и крезола с ФАФС катализируетфермент сульфотрансфераза