

 Биология
БиологияПохожие презентации:


")







Нейруляция и дифференцировка нейроэктодермы
1.
Нейруляцияи
дифференцировка
нейроэктодермы
2.
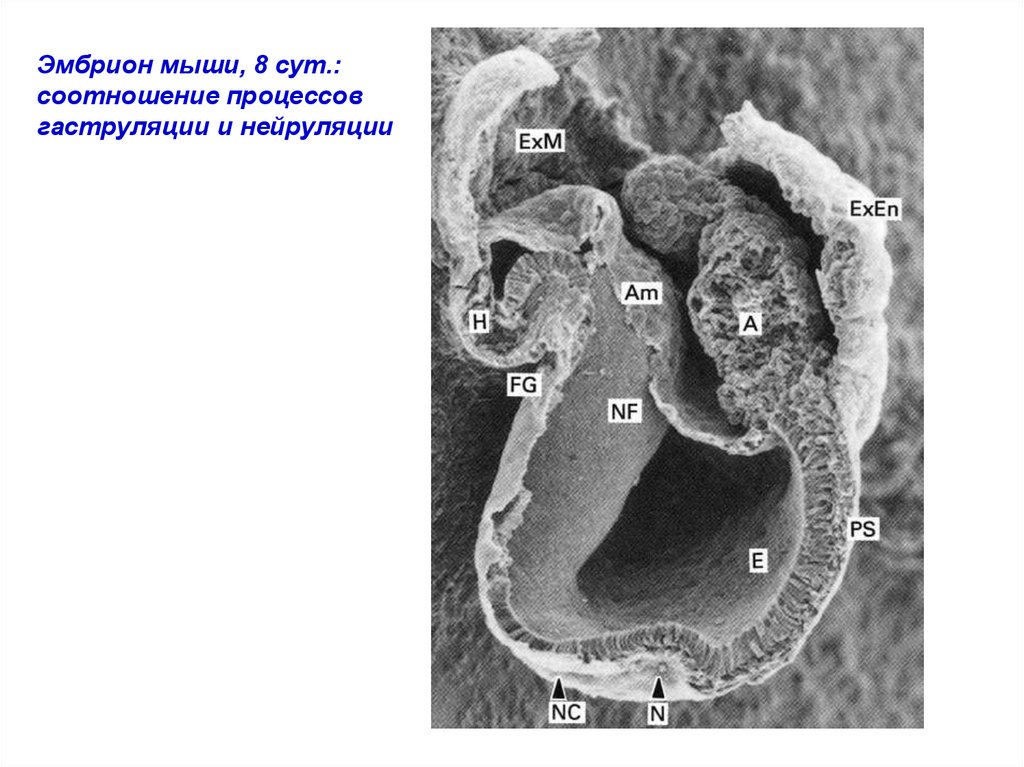
Эмбрион мыши, 8 сут.:соотношение процессов
гаструляции и нейруляции
3.
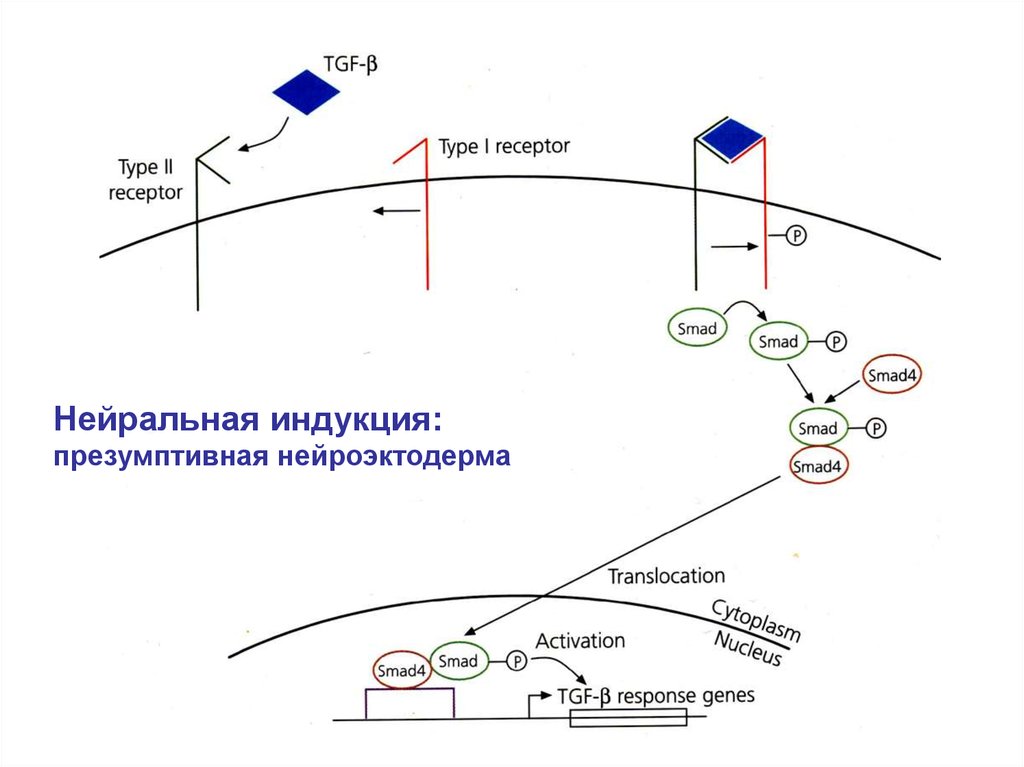
Нейральная индукция:презумптивная нейроэктодерма
4.
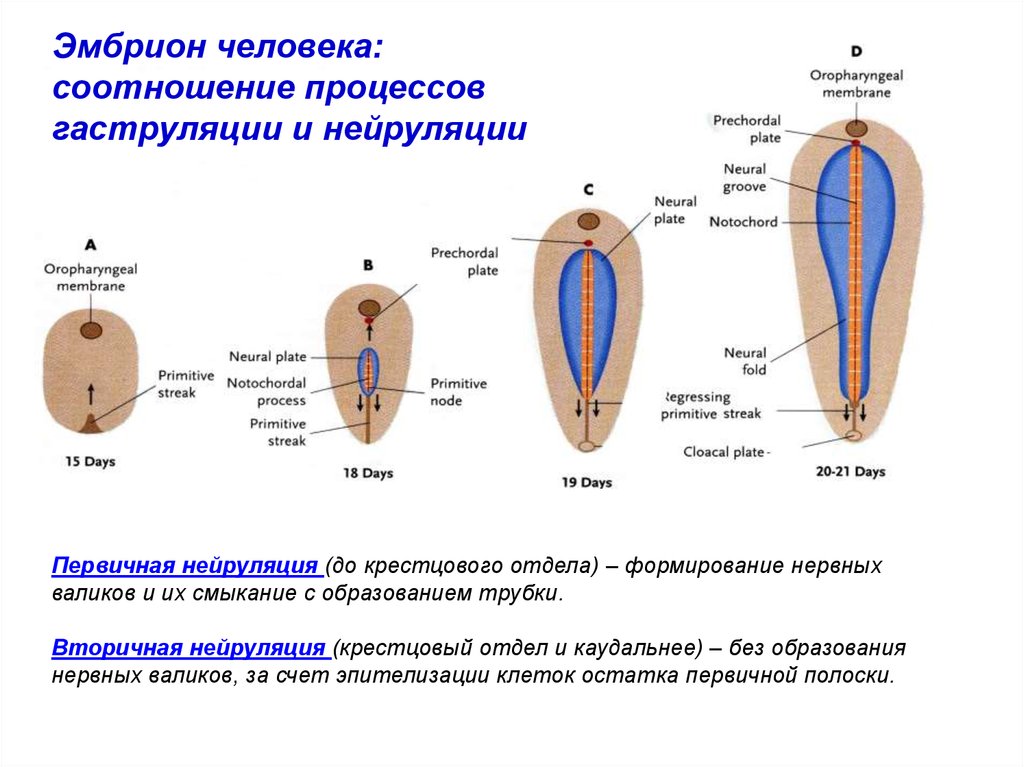
Эмбрион человека:соотношение процессов
гаструляции и нейруляции
Первичная нейруляция (до крестцового отдела) – формирование нервных
валиков и их смыкание с образованием трубки.
Вторичная нейруляция (крестцовый отдел и каудальнее) – без образования
нервных валиков, за счет эпителизации клеток остатка первичной полоски.
5.
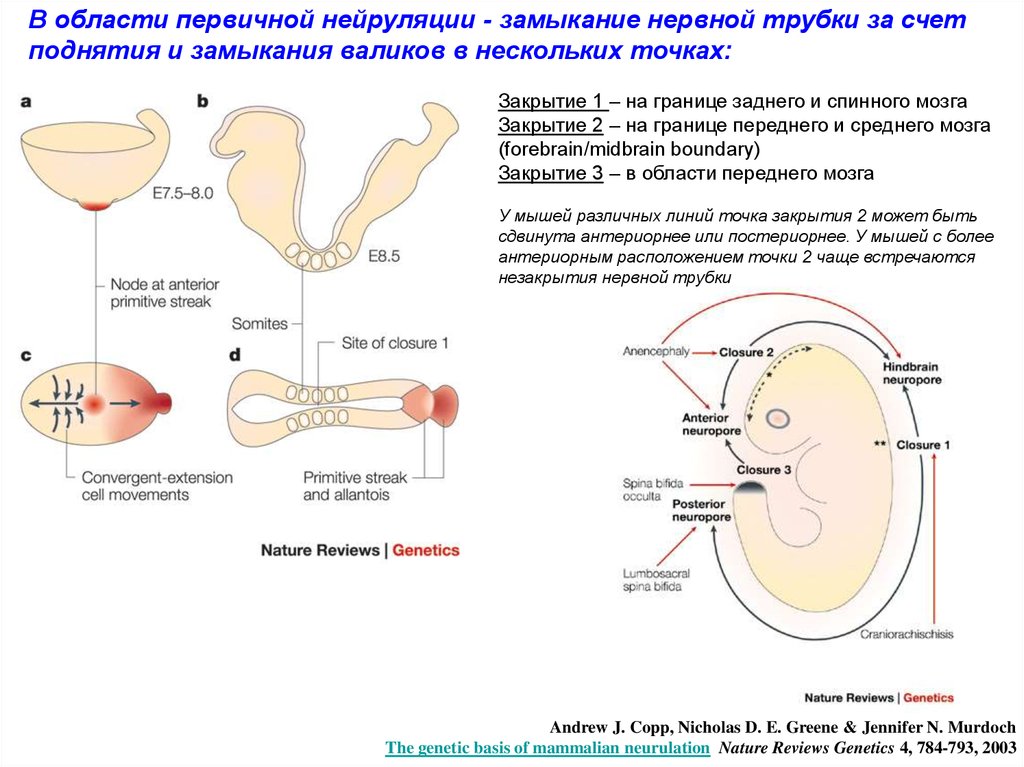
В области первичной нейруляции - замыкание нервной трубки за счетподнятия и замыкания валиков в нескольких точках:
Закрытие 1 – на границе заднего и спинного мозга
Закрытие 2 – на границе переднего и среднего мозга
(forebrain/midbrain boundary)
Закрытие 3 – в области переднего мозга
У мышей различных линий точка закрытия 2 может быть
сдвинута антериорнее или постериорнее. У мышей с более
антериорным расположением точки 2 чаще встречаются
незакрытия нервной трубки
Andrew J. Copp, Nicholas D. E. Greene & Jennifer N. Murdoch
The genetic basis of mammalian neurulation Nature Reviews Genetics 4, 784-793, 2003
6.
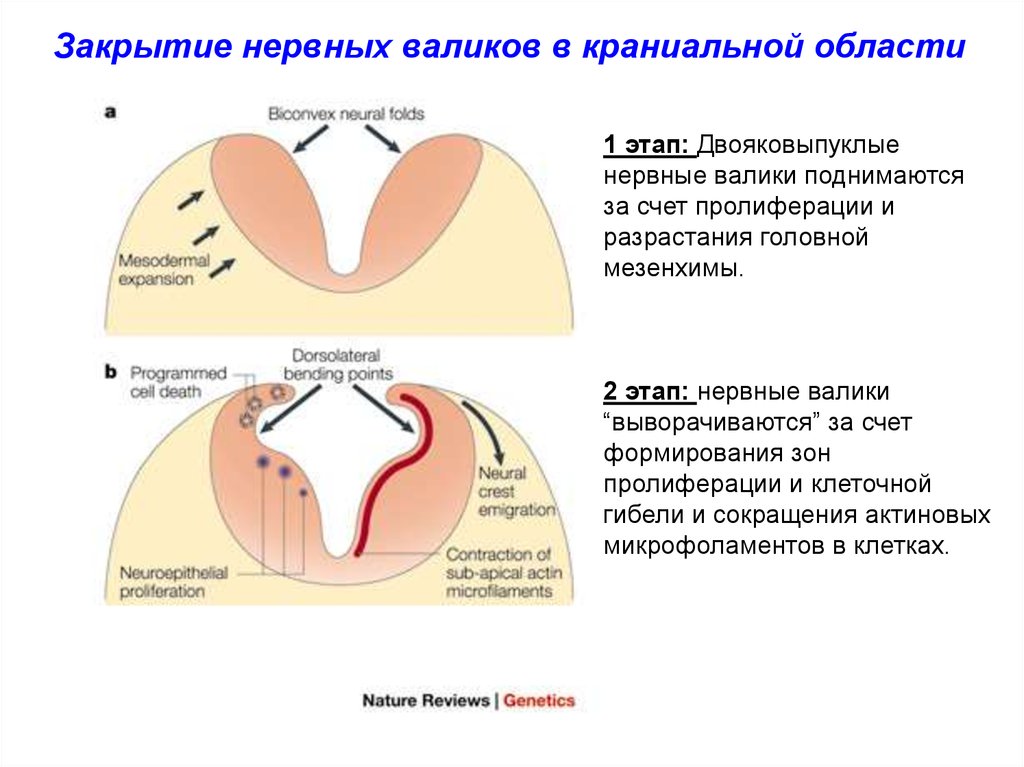
Закрытие нервных валиков в краниальной области1 этап: Двояковыпуклые
нервные валики поднимаются
за счет пролиферации и
разрастания головной
мезенхимы.
2 этап: нервные валики
“выворачиваются” за счет
формирования зон
пролиферации и клеточной
гибели и сокращения актиновых
микрофоламентов в клетках.
7.
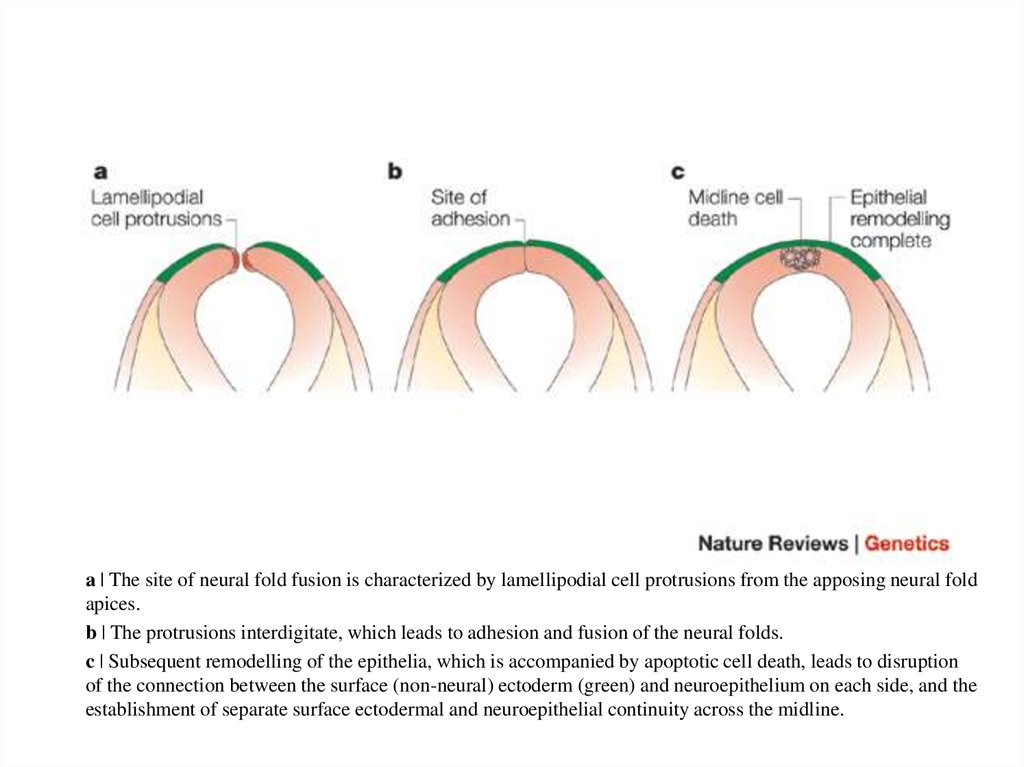
a | The site of neural fold fusion is characterized by lamellipodial cell protrusions from the apposing neural foldapices.
b | The protrusions interdigitate, which leads to adhesion and fusion of the neural folds.
c | Subsequent remodelling of the epithelia, which is accompanied by apoptotic cell death, leads to disruption
of the connection between the surface (non-neural) ectoderm (green) and neuroepithelium on each side, and the
establishment of separate surface ectodermal and neuroepithelial continuity across the midline.
8.
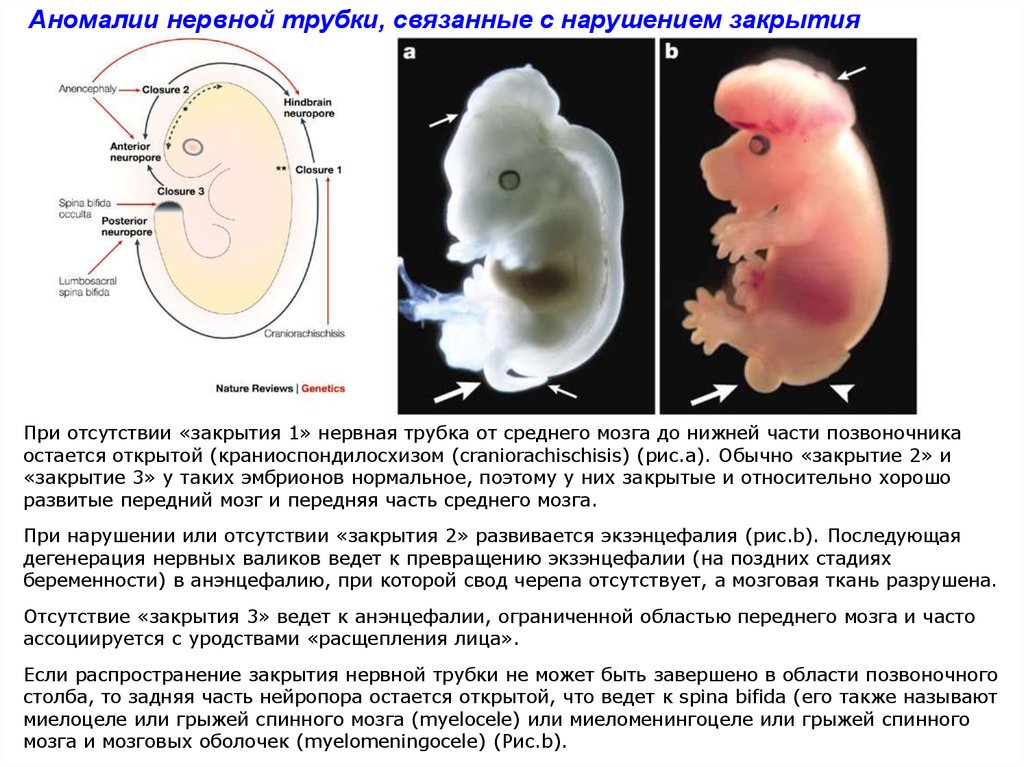
Аномалии нервной трубки, связанные с нарушением закрытияПри отсутствии «закрытия 1» нервная трубка от среднего мозга до нижней части позвоночника
остается открытой (краниоспондилосхизом (craniorachischisis) (рис.а). Обычно «закрытие 2» и
«закрытие 3» у таких эмбрионов нормальное, поэтому у них закрытые и относительно хорошо
развитые передний мозг и передняя часть среднего мозга.
При нарушении или отсутствии «закрытия 2» развивается экзэнцефалия (рис.b). Последующая
дегенерация нервных валиков ведет к превращению экзэнцефалии (на поздних стадиях
беременности) в анэнцефалию, при которой свод черепа отсутствует, а мозговая ткань разрушена.
Отсутствие «закрытия 3» ведет к анэнцефалии, ограниченной областью переднего мозга и часто
ассоциируется с уродствами «расщепления лица».
Если распространение закрытия нервной трубки не может быть завершено в области позвоночного
столба, то задняя часть нейропора остается открытой, что ведет к spina bifida (его также называют
миелоцеле или грыжей спинного мозга (myelocele) или миеломенингоцеле или грыжей спинного
мозга и мозговых оболочек (myelomeningocele) (Рис.b).
9.
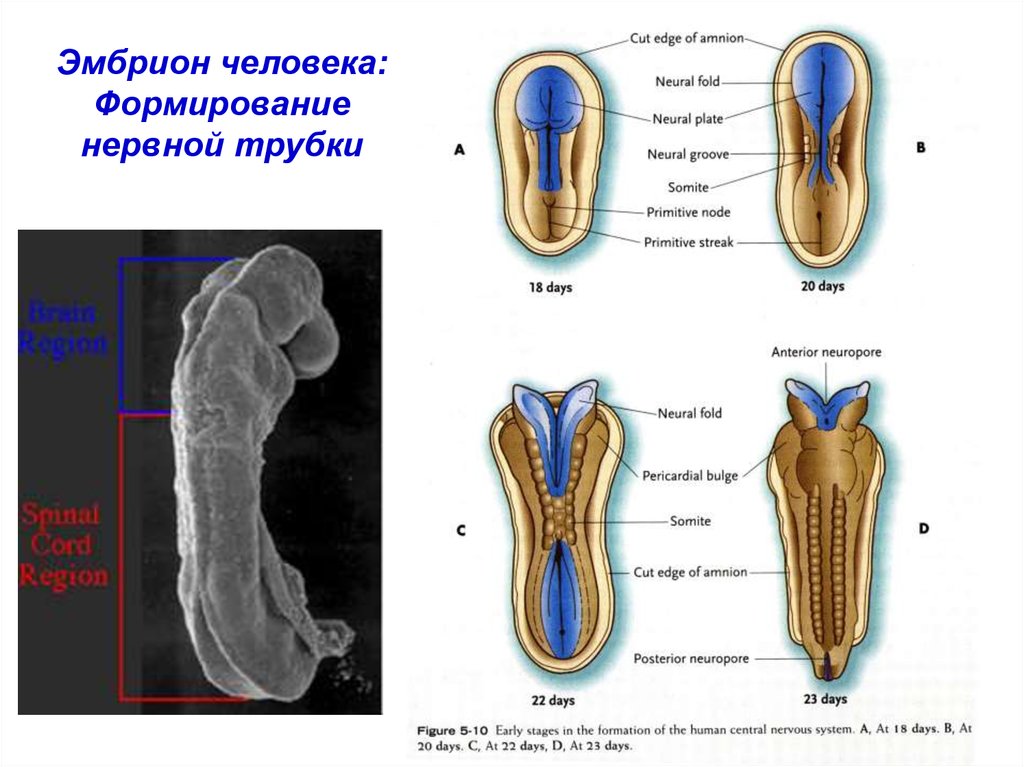
Эмбрион человека:Формирование
нервной трубки
10.
11.
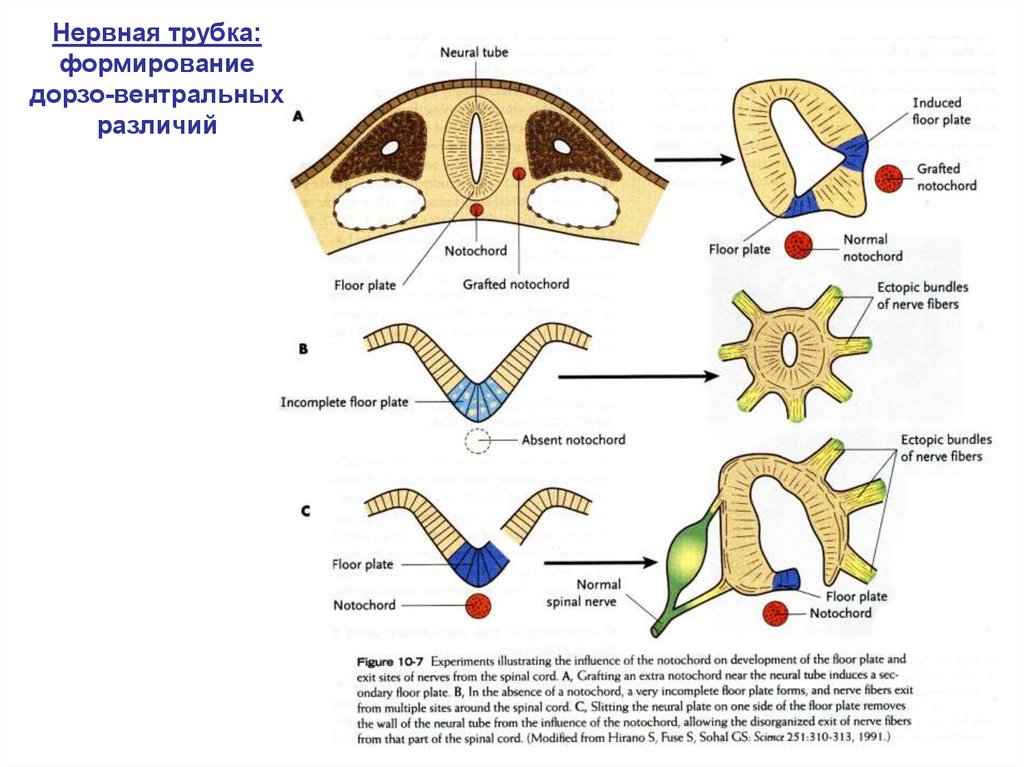
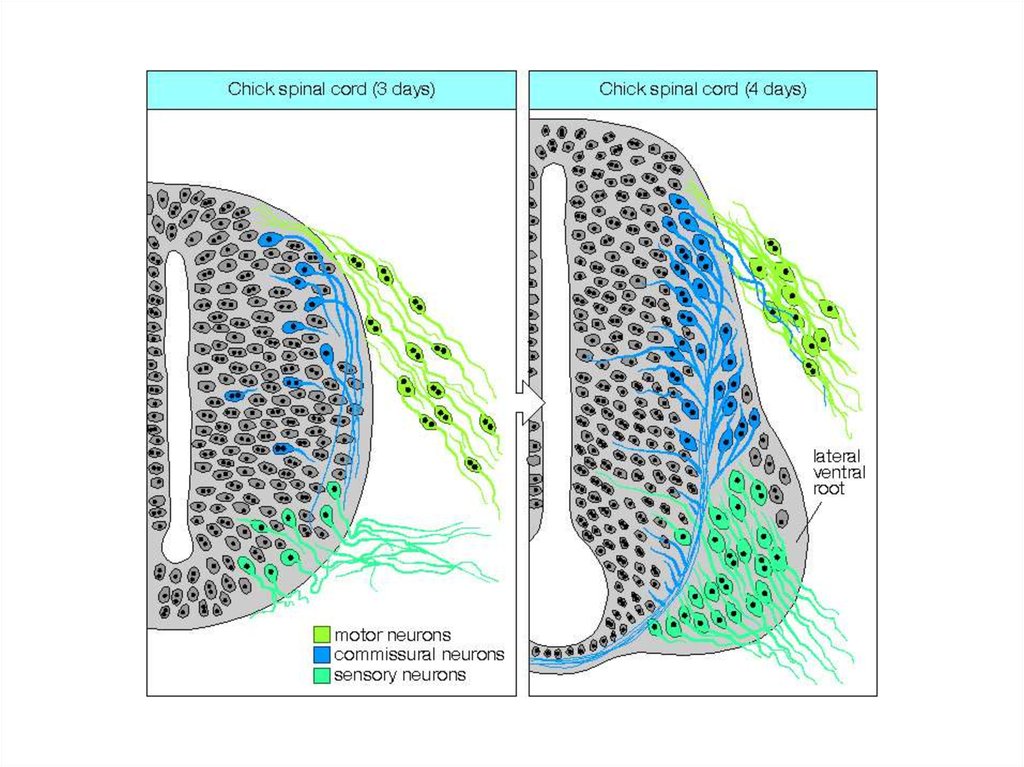
Нервная трубка:формирование
дорзо-вентральных
различий
12.
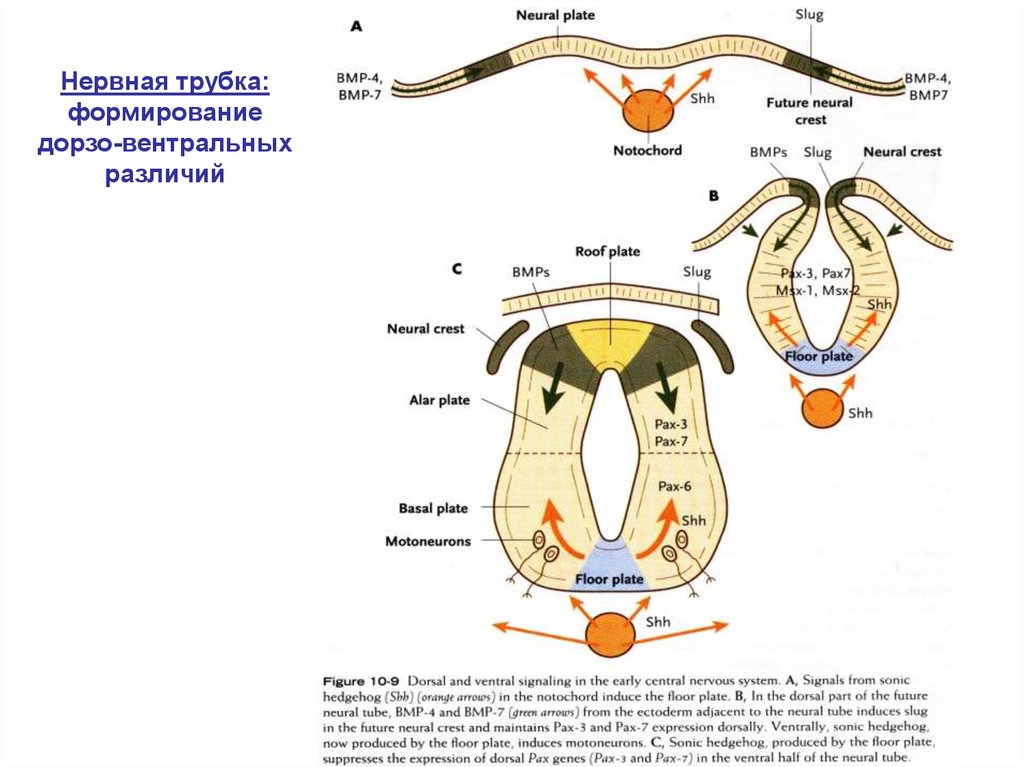
Нервная трубка:формирование
дорзо-вентральных
различий
13.
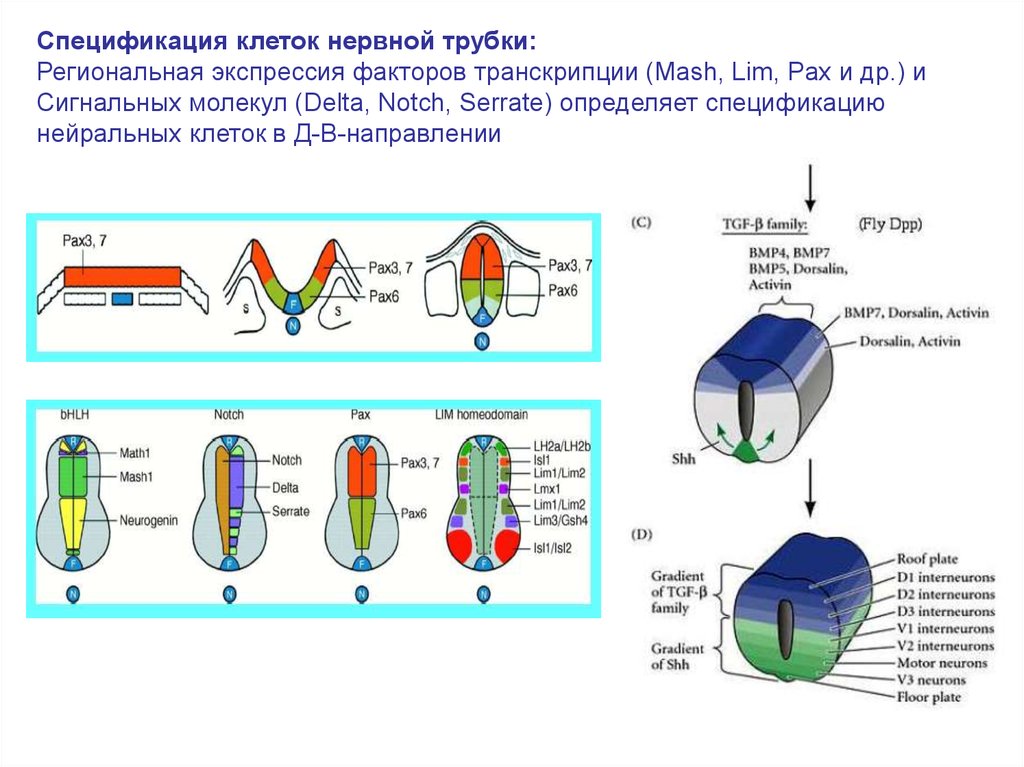
Спецификация клеток нервной трубки:Региональная экспрессия факторов транскрипции (Mash, Lim, Pax и др.) и
Сигнальных молекул (Delta, Notch, Serrate) определяет спецификацию
нейральных клеток в Д-В-направлении
14.
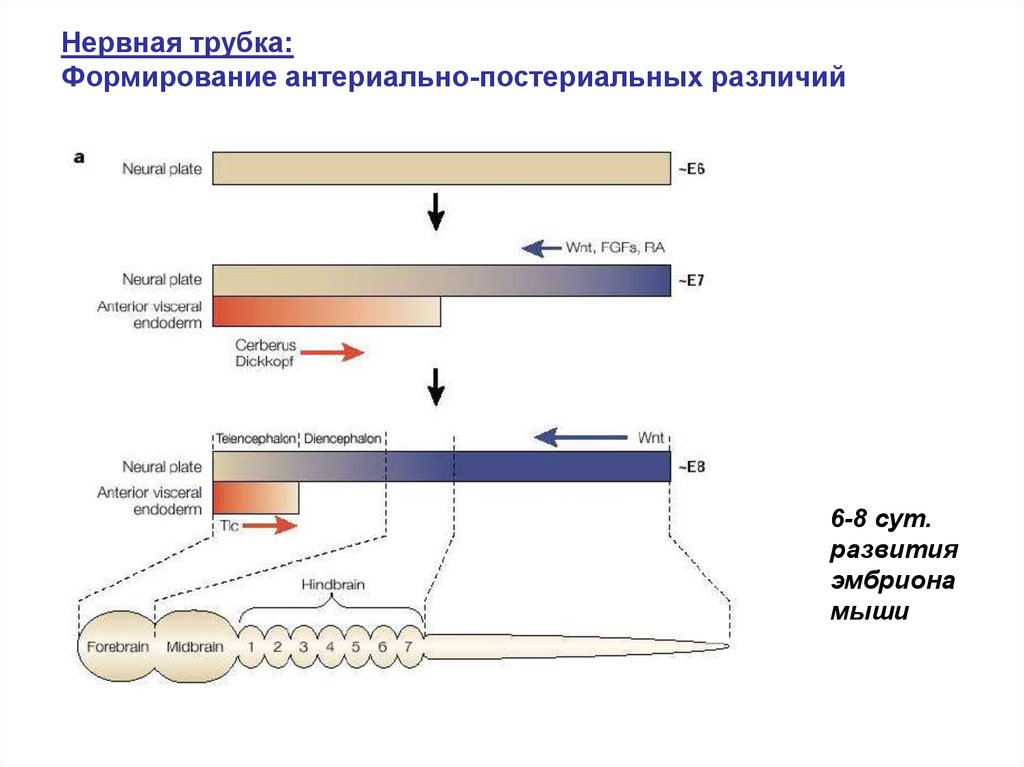
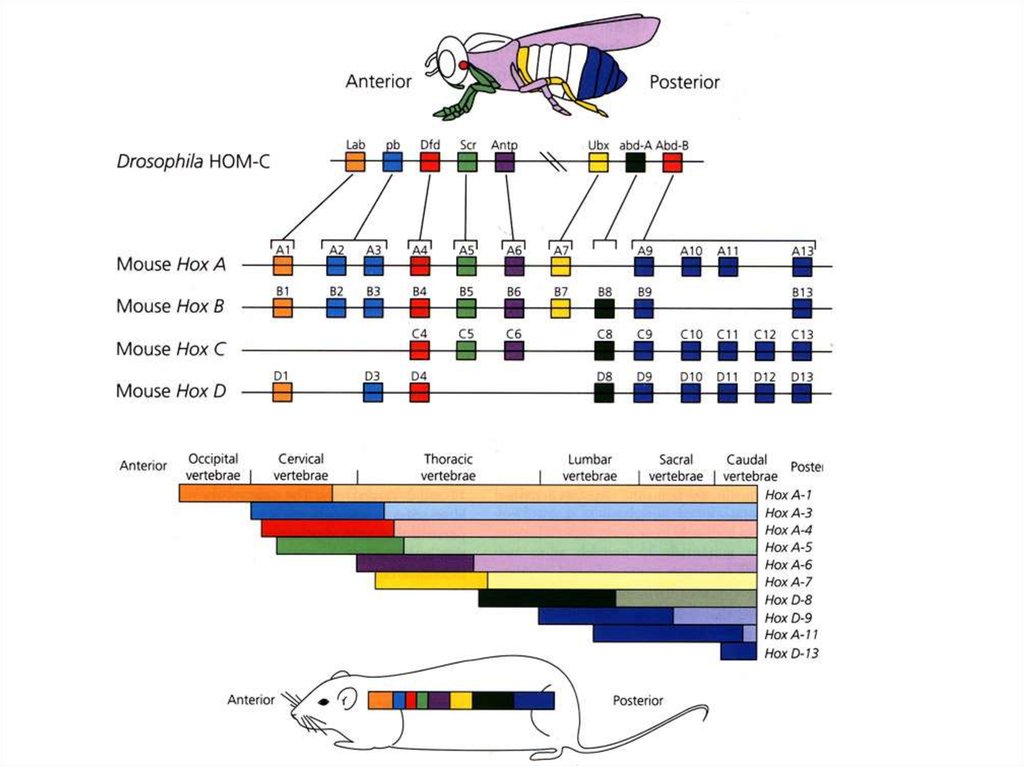
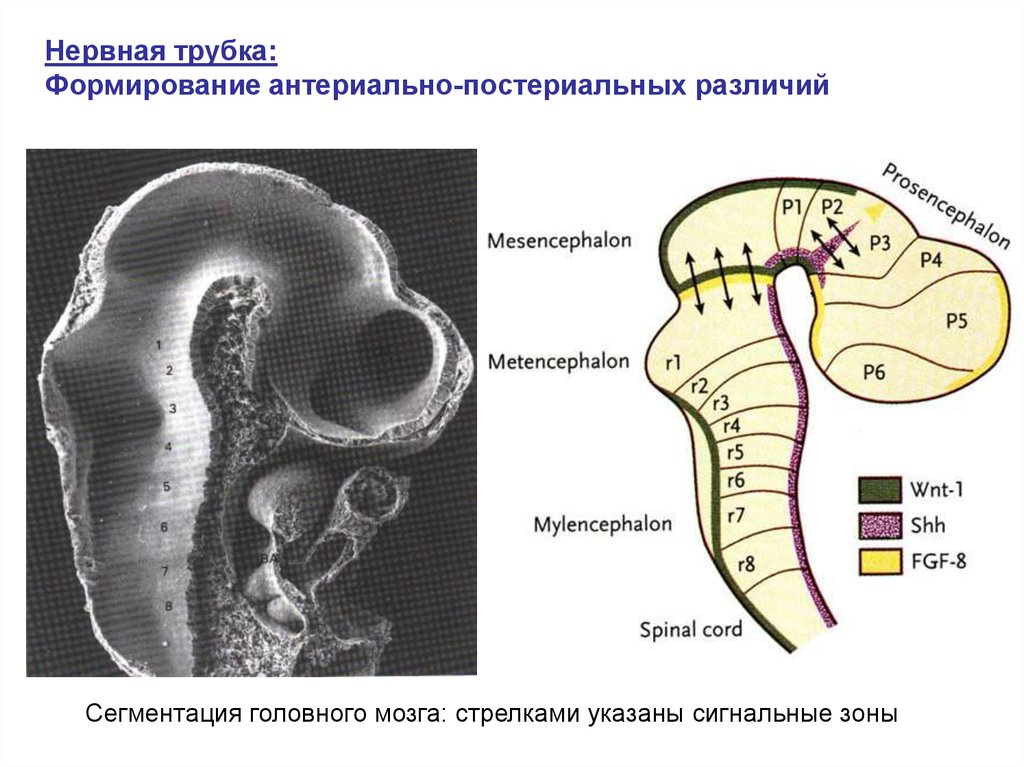
Нервная трубка:Формирование антериально-постериальных различий
6-8 сут.
развития
эмбриона
мыши
15.
16.
17.
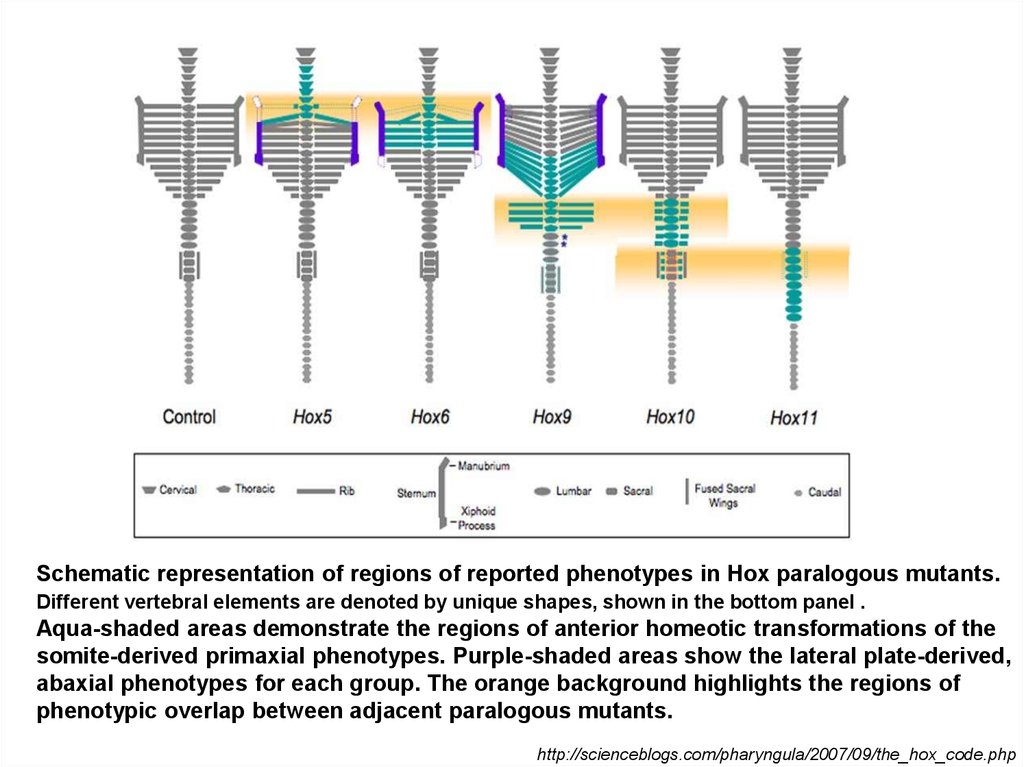
Schematic representation of regions of reported phenotypes in Hox paralogous mutants.Different vertebral elements are denoted by unique shapes, shown in the bottom panel .
Aqua-shaded areas demonstrate the regions of anterior homeotic transformations of the
somite-derived primaxial phenotypes. Purple-shaded areas show the lateral plate-derived,
abaxial phenotypes for each group. The orange background highlights the regions of
phenotypic overlap between adjacent paralogous mutants.
http://scienceblogs.com/pharyngula/2007/09/the_hox_code.php
18.
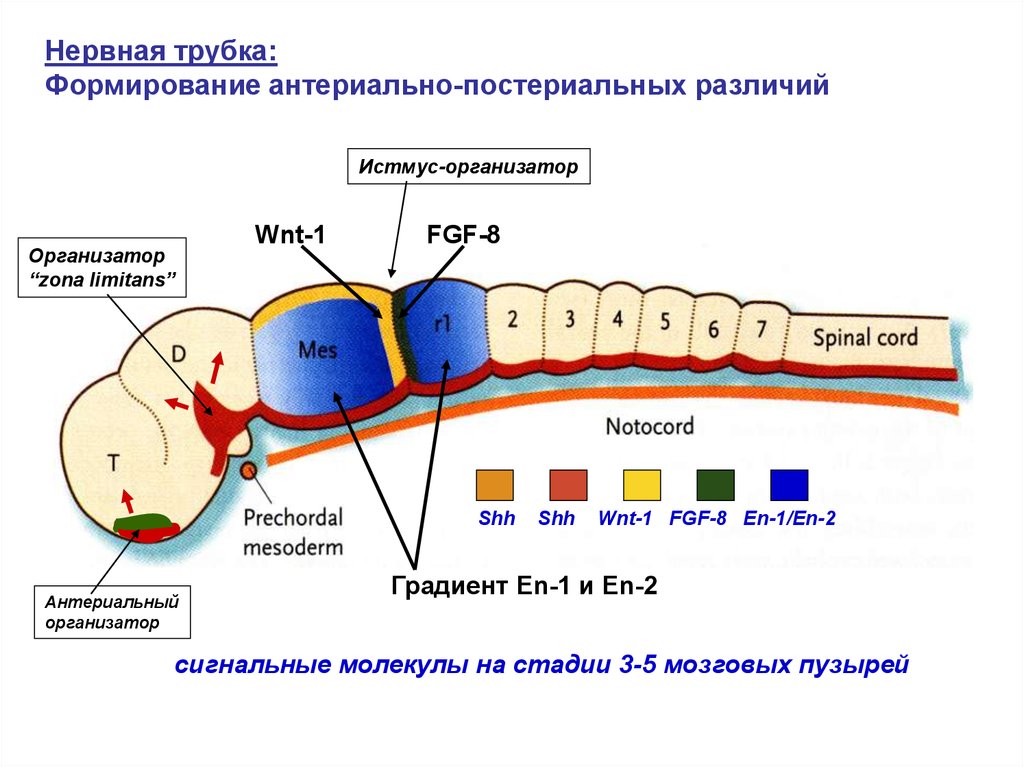
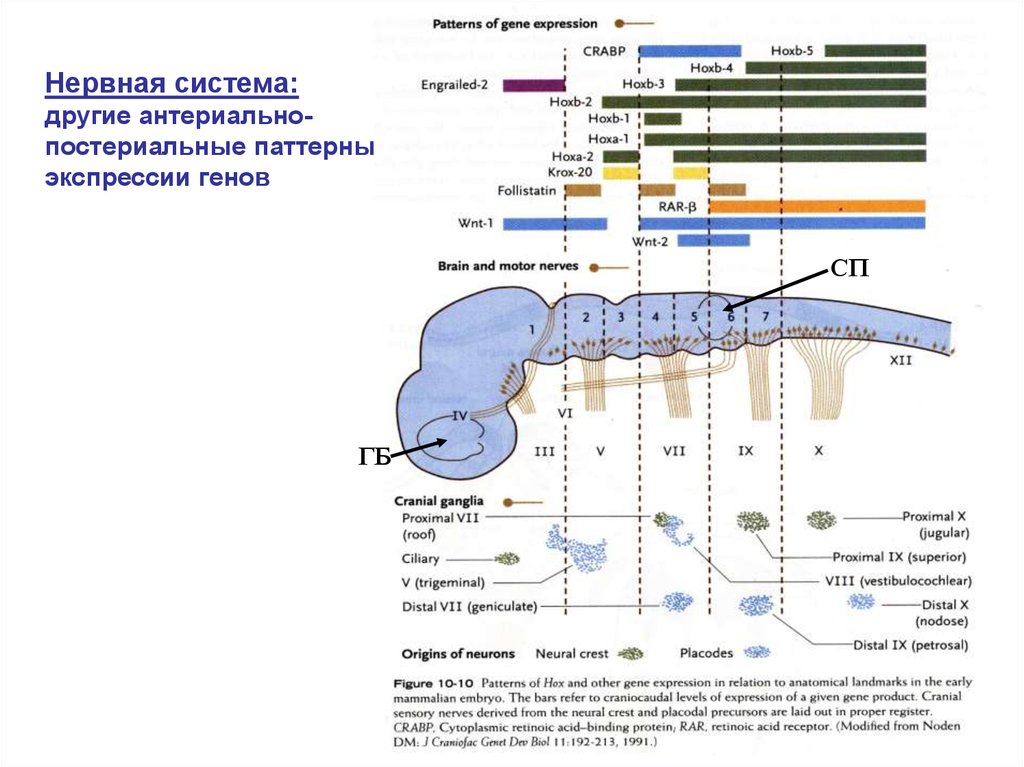
Нервная трубка:Формирование антериально-постериальных различий
Истмус-организатор
Организатор
“zona limitans”
Wnt-1
FGF-8
Shh
Антериальный
организатор
Shh
Wnt-1 FGF-8 En-1/En-2
Градиент En-1 и Еn-2
сигнальные молекулы на стадии 3-5 мозговых пузырей
19.
Нервная трубка:Формирование антериально-постериальных различий
Сегментация головного мозга: стрелками указаны сигнальные зоны
20.
Нервная система:другие антериальнопостериальные паттерны
экспрессии генов
СП
ГБ
21.
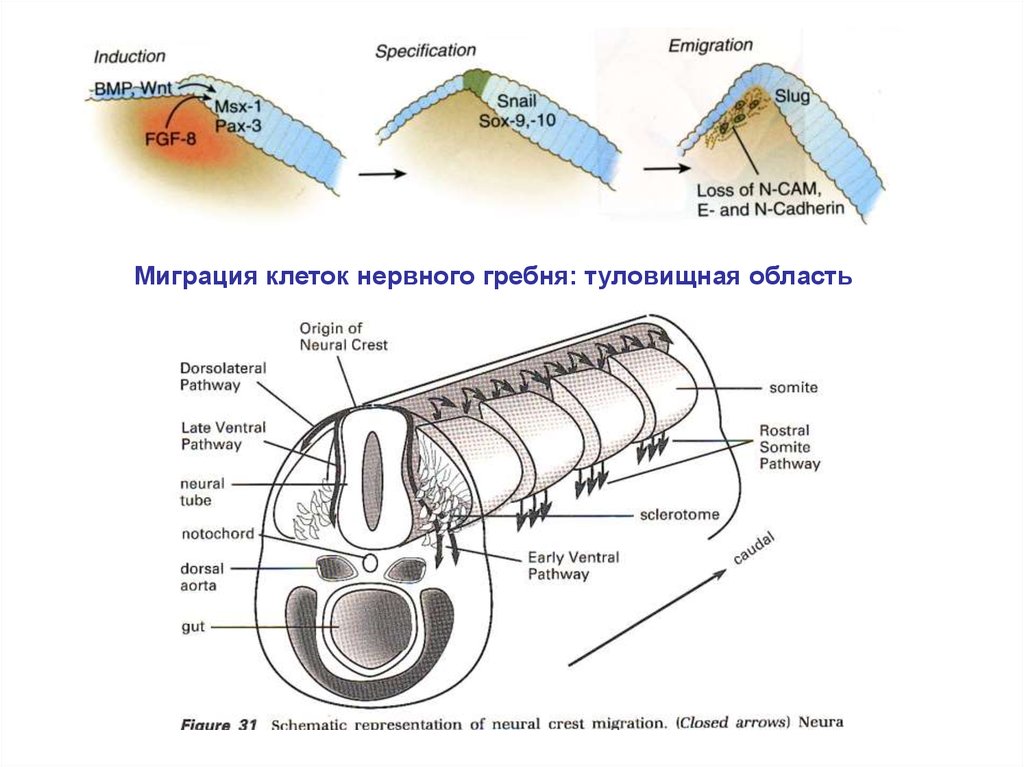
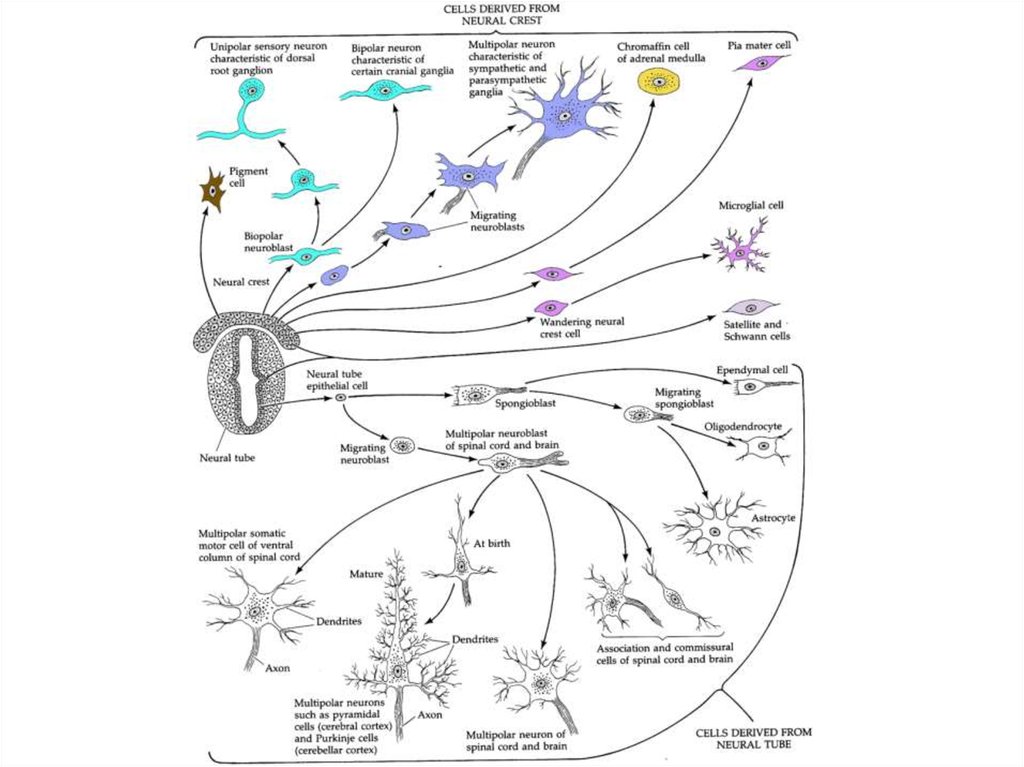
Миграция клеток нервного гребня: туловищная область22.
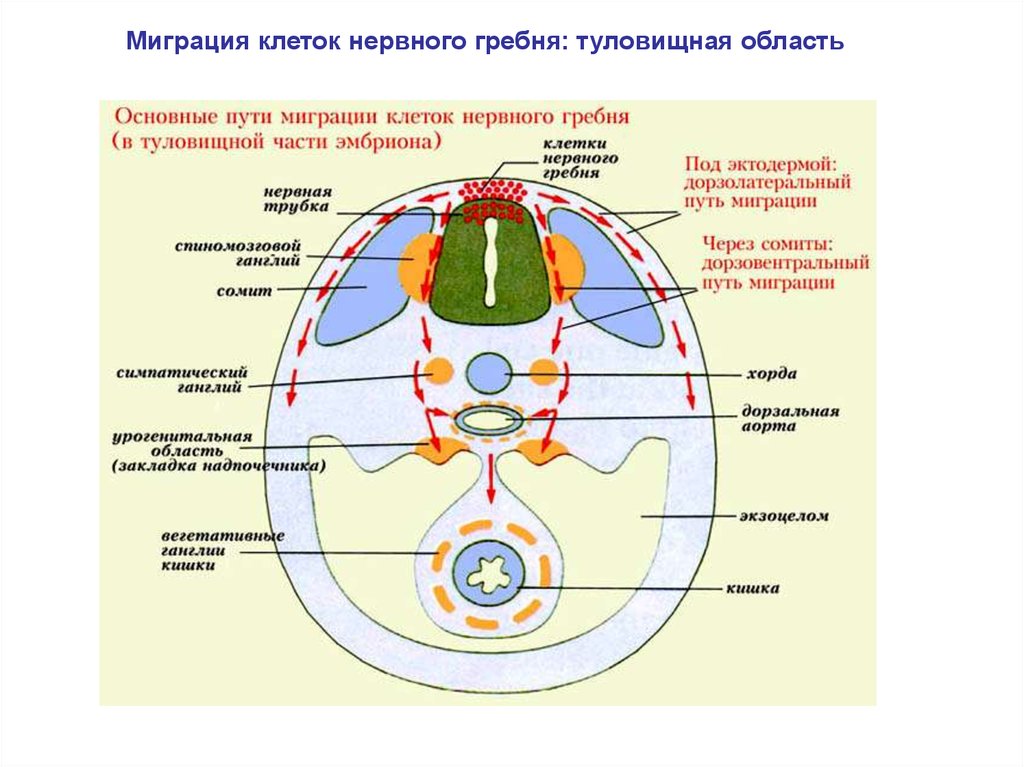
Миграция клеток нервного гребня: туловищная область23.
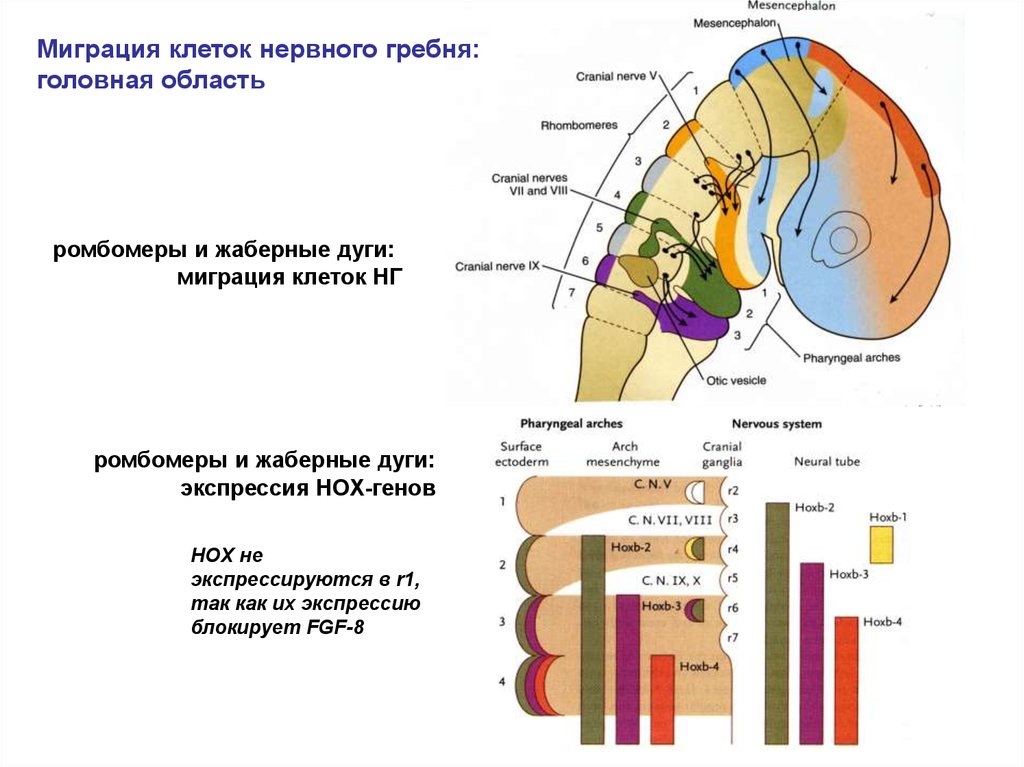
Миграция клеток нервного гребня:головная область
ромбомеры и жаберные дуги:
миграция клеток НГ
ромбомеры и жаберные дуги:
экспрессия НОХ-генов
HOX не
экспрессируются в r1,
так как их экспрессию
блокирует FGF-8
24.
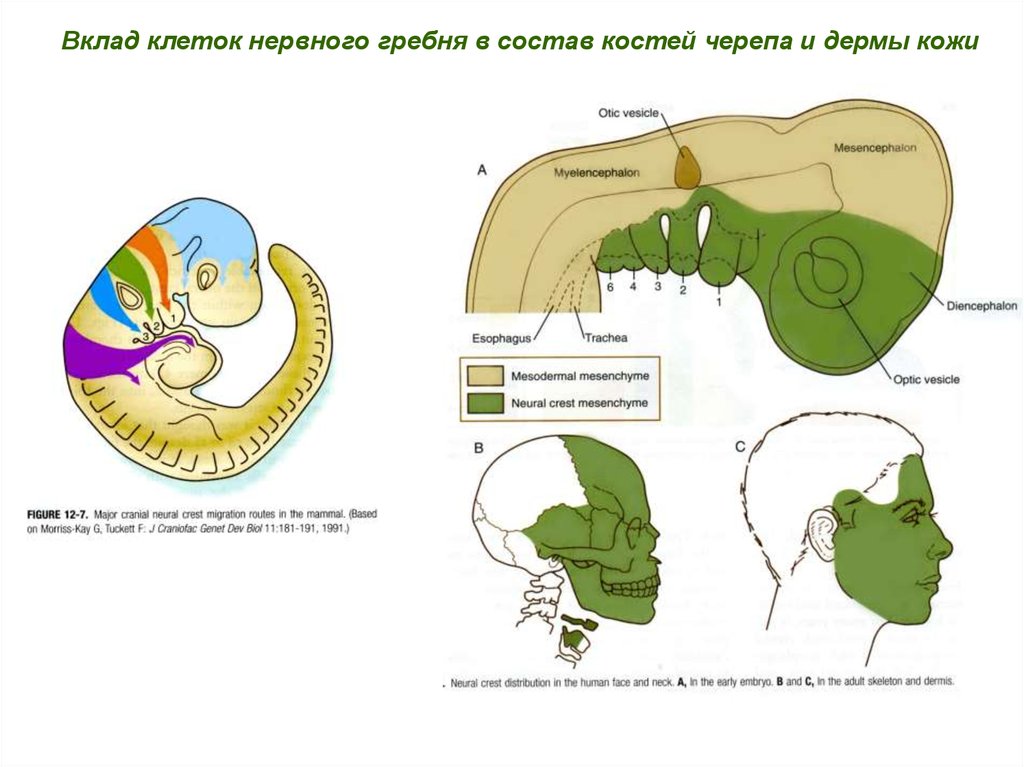
Вклад клеток нервного гребня в состав костей черепа и дермы кожи25.
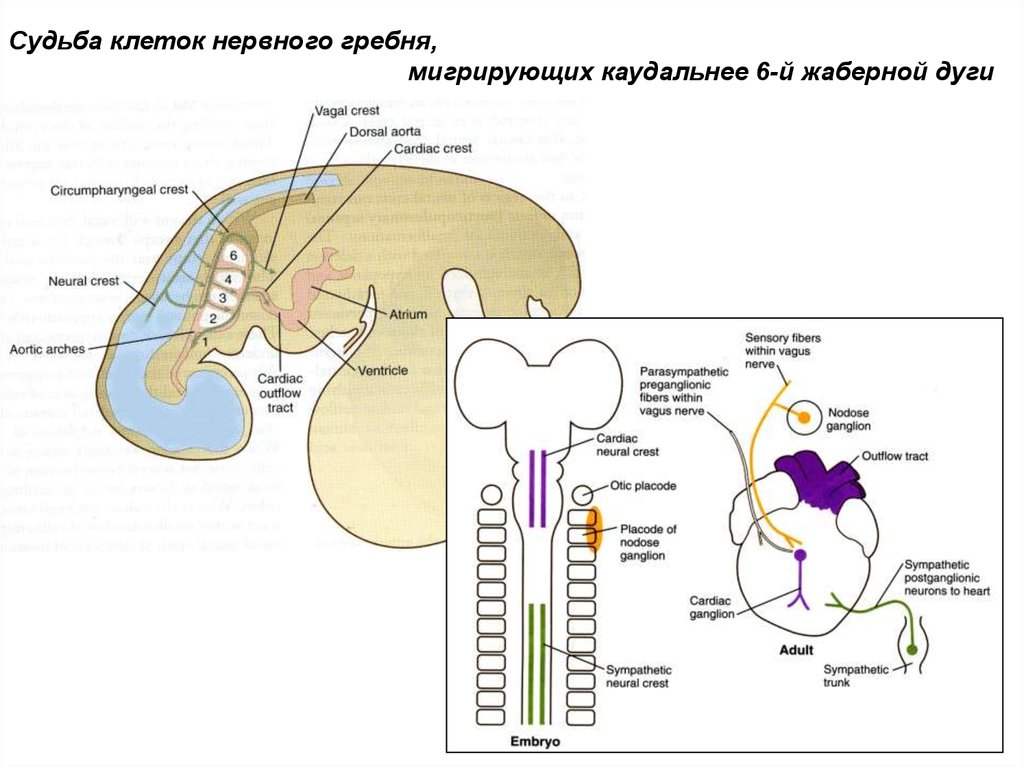
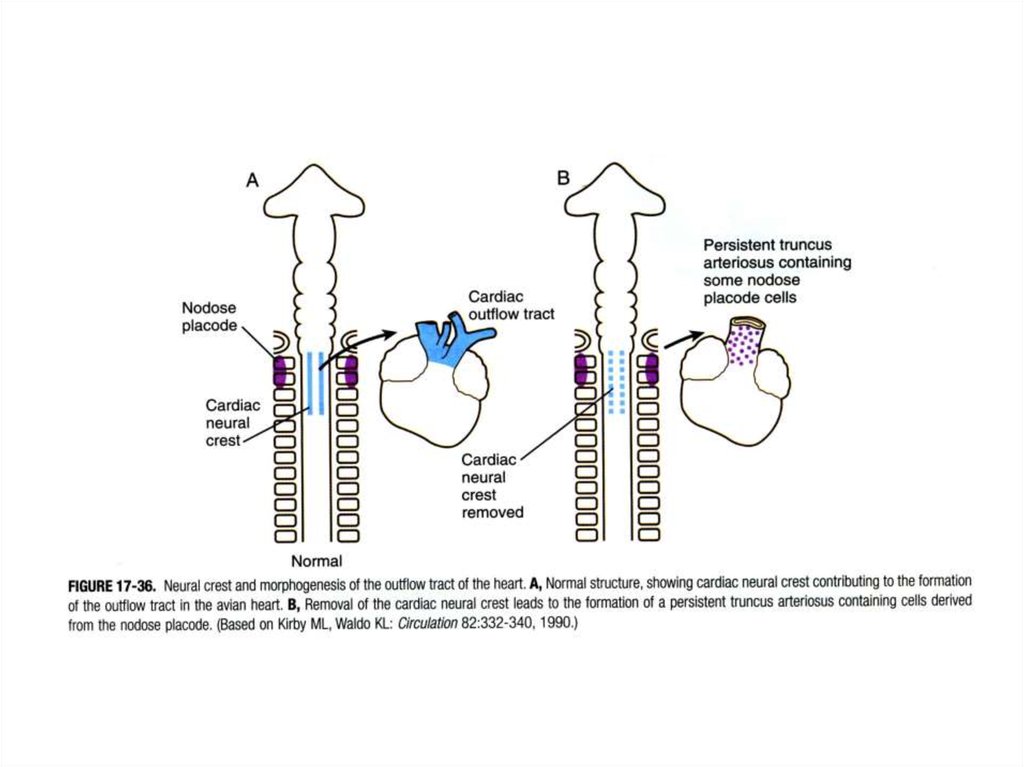
Судьба клеток нервного гребня,мигрирующих каудальнее 6-й жаберной дуги
26.
27.
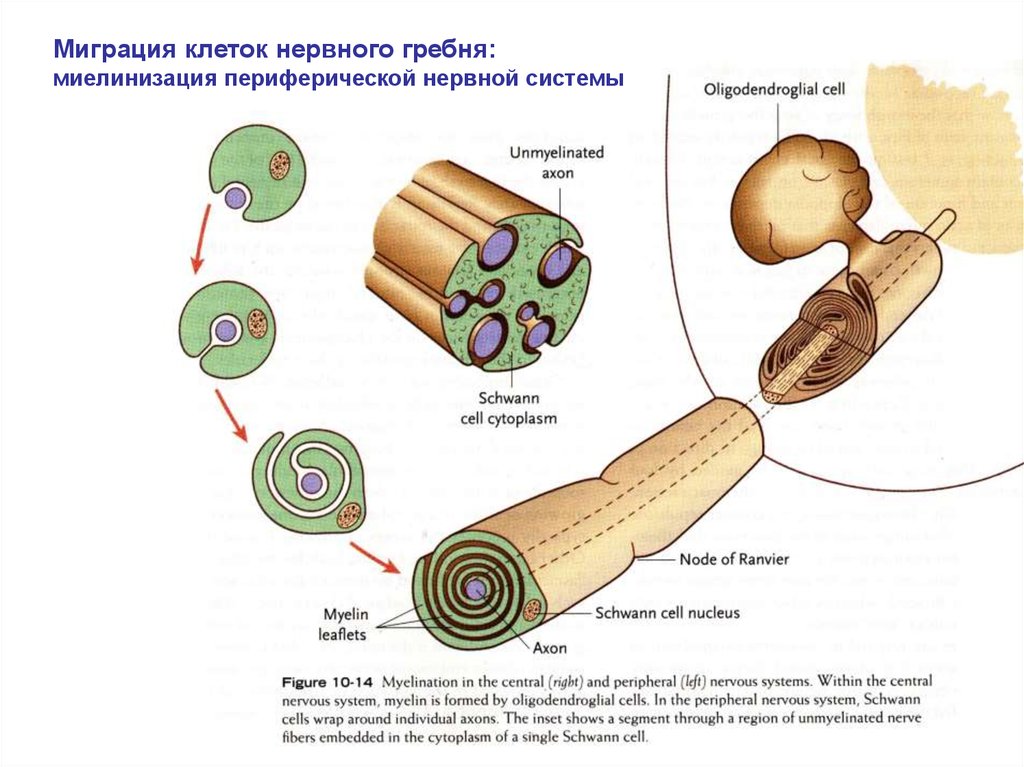
Миграция клеток нервного гребня:миелинизация периферической нервной системы
28.
29.
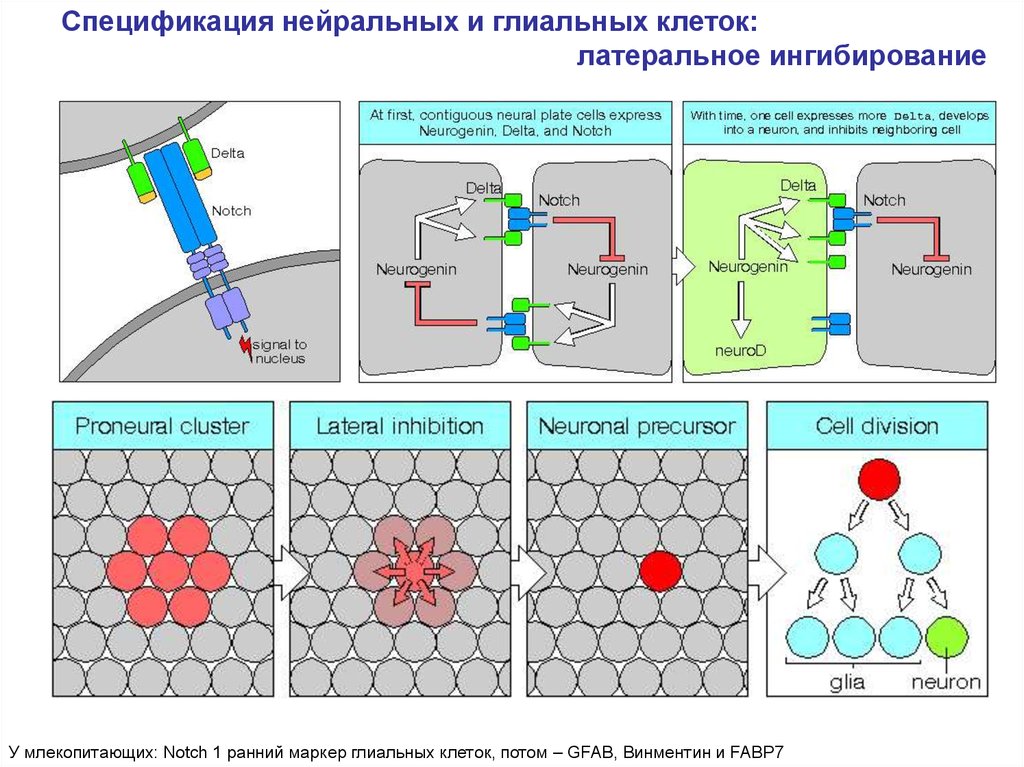
Спецификация нейральных и глиальных клеток:латеральное ингибирование
У млекопитающих: Notch 1 ранний маркер глиальных клеток, потом – GFAB, Винментин и FABP7
30.
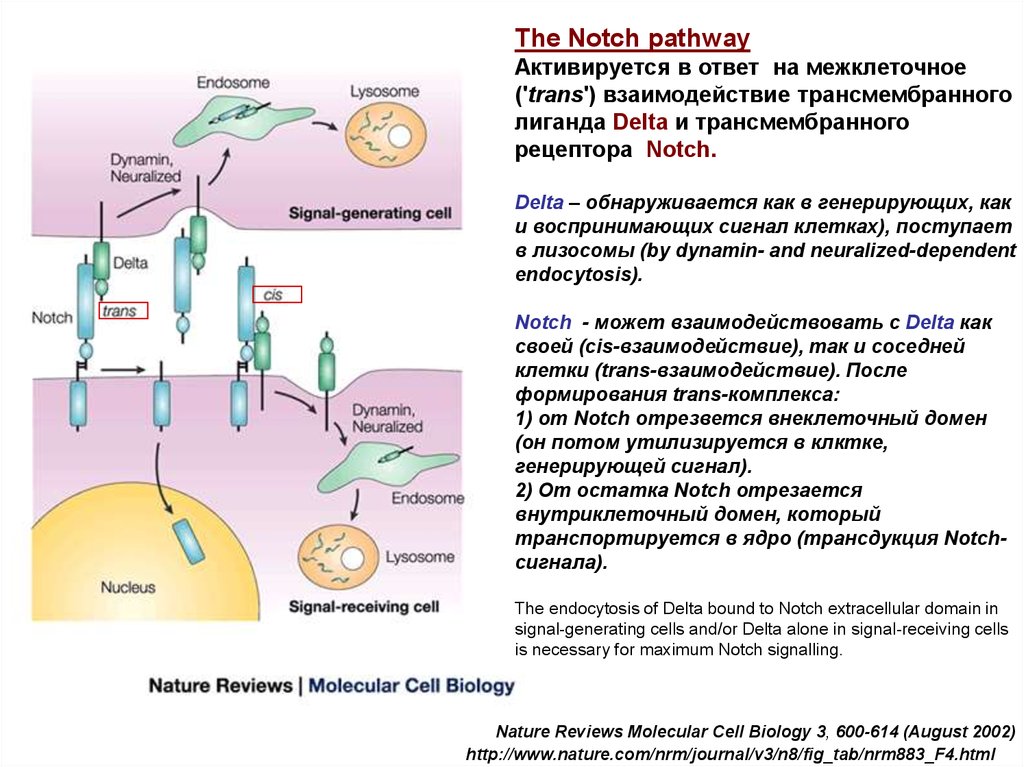
The Notch pathwayАктивируется в ответ на межклеточное
('trans') взаимодействие трансмембранного
лиганда Delta и трансмембранного
рецептора Notch.
Delta – обнаруживается как в генерирующих, как
и воспринимающих сигнал клетках), поступает
в лизосомы (by dynamin- and neuralized-dependent
endocytosis).
Notch - может взаимодействовать с Delta как
своей (cis-взаимодействие), так и соседней
клетки (trans-взаимодействие). После
формирования trans-комплекса:
1) от Notch отрезвется внеклеточный домен
(он потом утилизируется в клктке,
генерирующей сигнал).
2) От остатка Notch отрезается
внутриклеточный домен, который
транспортируется в ядро (трансдукция Notchсигнала).
The endocytosis of Delta bound to Notch extracellular domain in
signal-generating cells and/or Delta alone in signal-receiving cells
is necessary for maximum Notch signalling.
Nature Reviews Molecular Cell Biology 3, 600-614 (August 2002)
http://www.nature.com/nrm/journal/v3/n8/fig_tab/nrm883_F4.html
31.
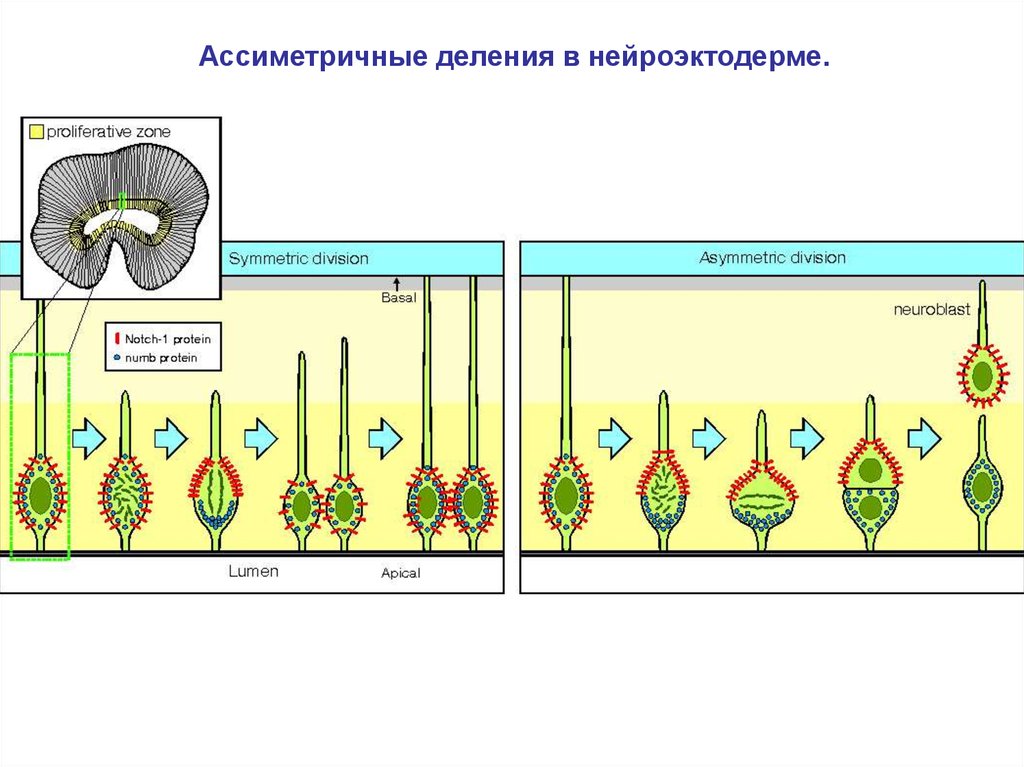
Ассиметричные деления в нейроэктодерме.32.
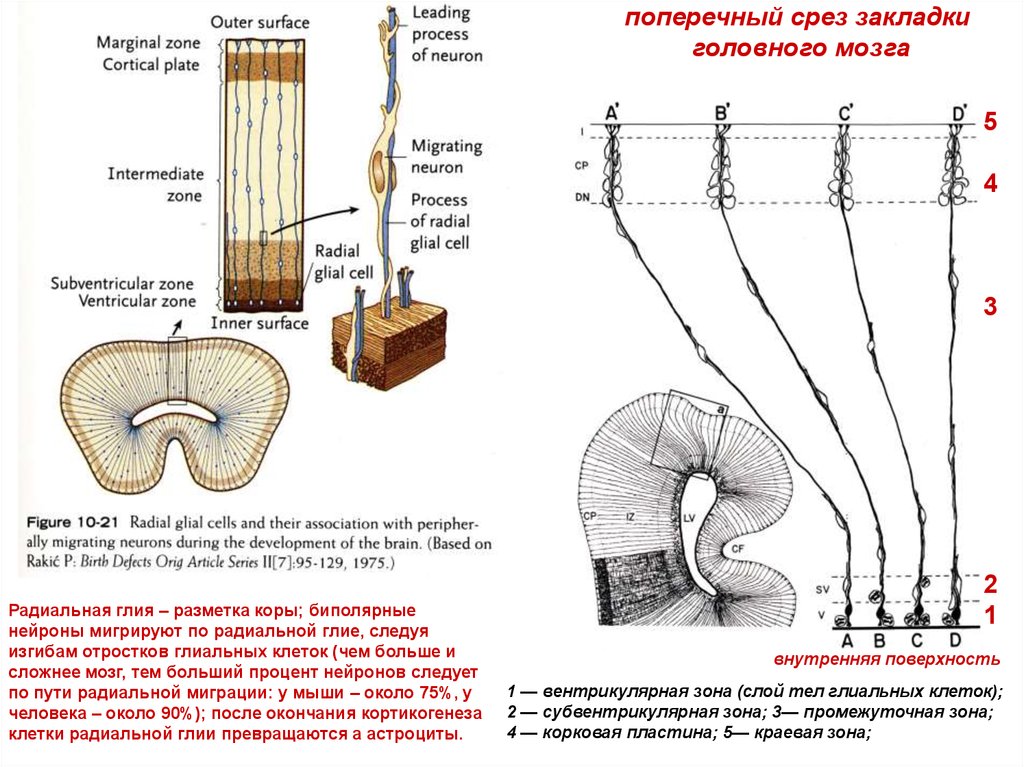
поперечный срез закладкиголовного мозга
5
4
3
Радиальная глия – разметка коры; биполярные
нейроны мигрируют по радиальной глие, следуя
изгибам отростков глиальных клеток (чем больше и
сложнее мозг, тем больший процент нейронов следует
по пути радиальной миграции: у мыши – около 75%, у
человека – около 90%); после окончания кортикогенеза
клетки радиальной глии превращаются а астроциты.
2
1
внутренняя поверхность
1 — вентрикулярная зона (слой тел глиальных клеток);
2 — субвентрикулярная зона; 3— промежуточная зона;
4 — корковая пластина; 5— краевая зона;
33.
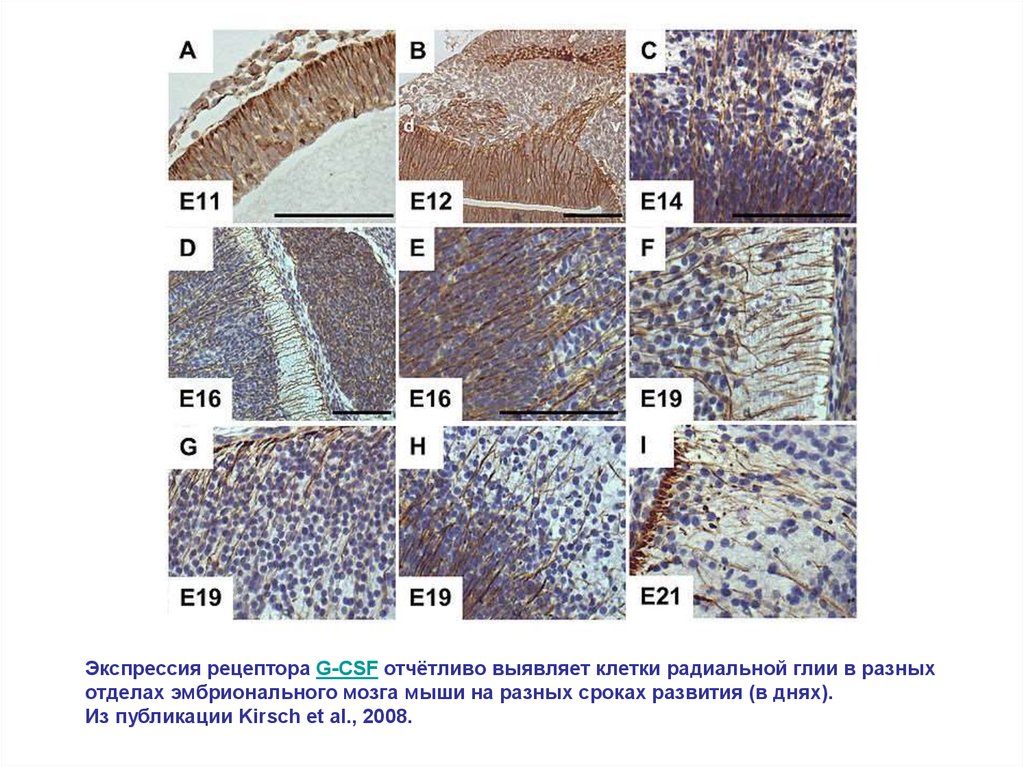
Экспрессия рецептора G-CSF отчётливо выявляет клетки радиальной глии в разныхотделах эмбрионального мозга мыши на разных сроках развития (в днях).
Из публикации Kirsch et al., 2008.
34.
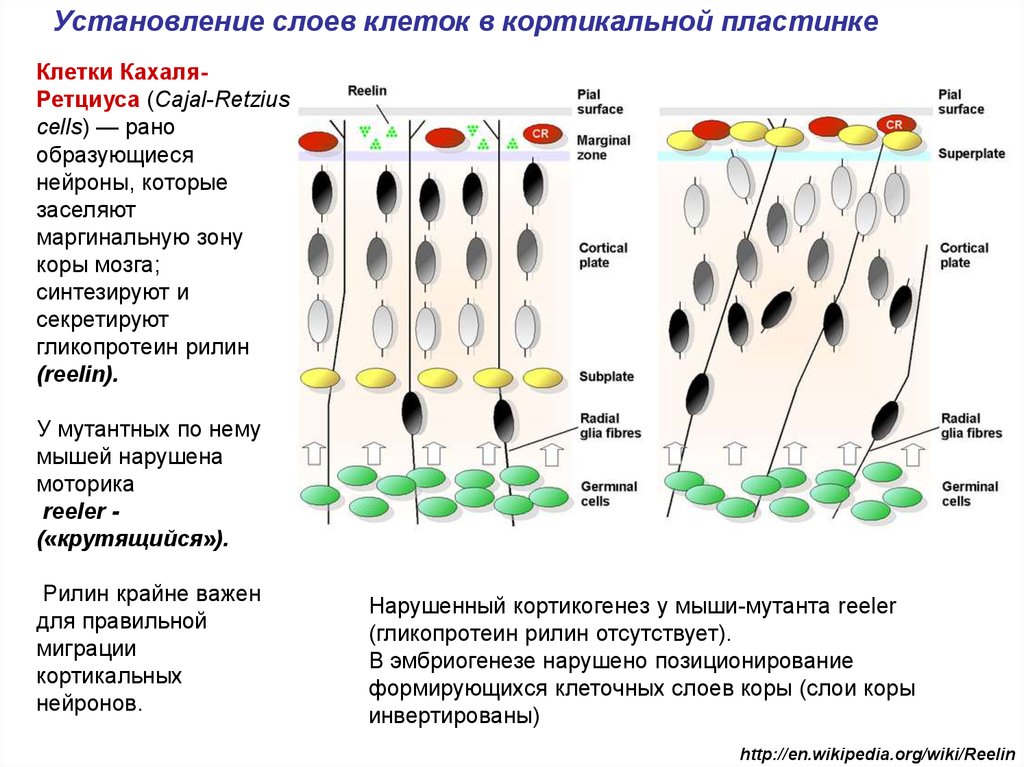
Установление слоев клеток в кортикальной пластинкеКлетки КахаляРетциуса (Cajal-Retzius
cells) — рано
образующиеся
нейроны, которые
заселяют
маргинальную зону
коры мозга;
синтезируют и
секретируют
гликопротеин рилин
(reelin).
У мутантных по нему
мышей нарушена
моторика
reeler («крутящийся»).
Рилин крайне важен
для правильной
миграции
кортикальных
нейронов.
Нарушенный кортикогенез у мыши-мутанта reeler
(гликопротеин рилин отсутствует).
В эмбриогенезе нарушено позиционирование
формирующихся клеточных слоев коры (слои коры
инвертированы)
http://en.wikipedia.org/wiki/Reelin
35.
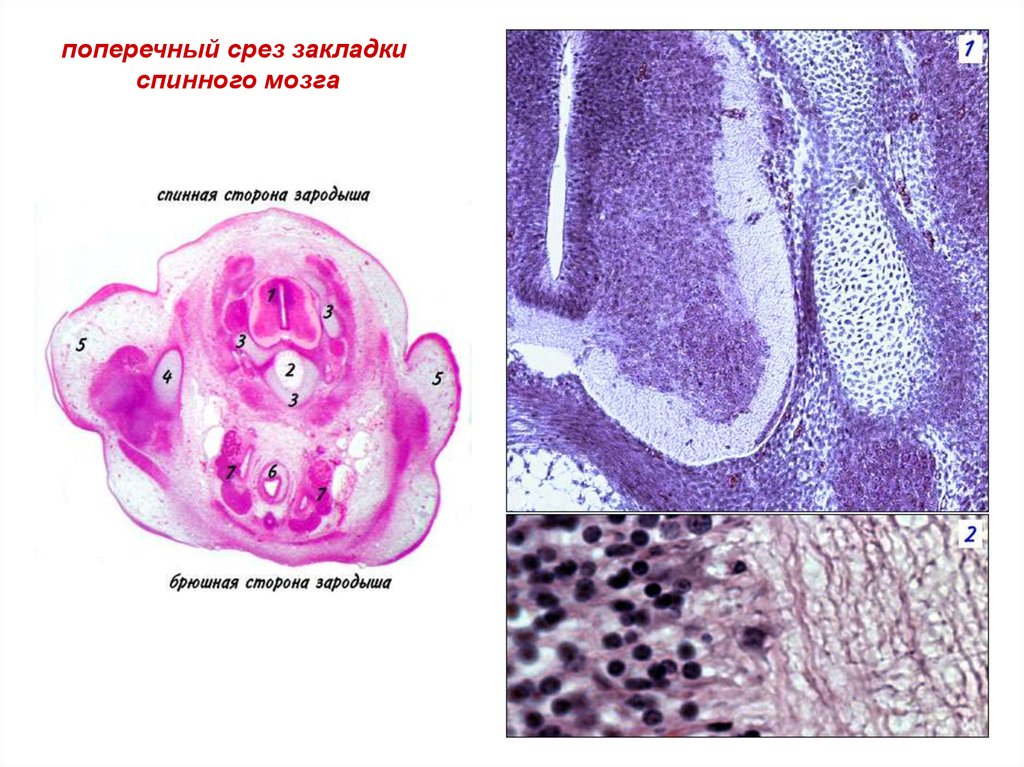
поперечный срез закладкиспинного мозга
36.
37.
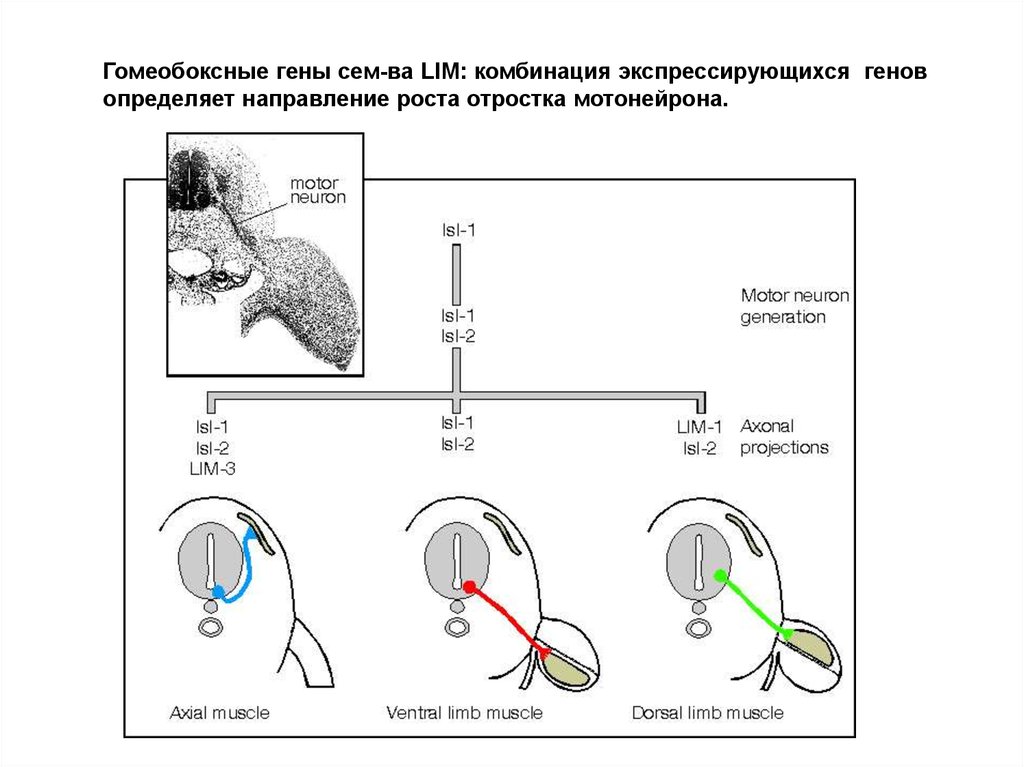
Гомеобоксные гены сем-ва LIM: комбинация экспрессирующихся геновопределяет направление роста отростка мотонейрона.