



















 География
ГеографияПохожие презентации:










Bunkový cyklus
1.
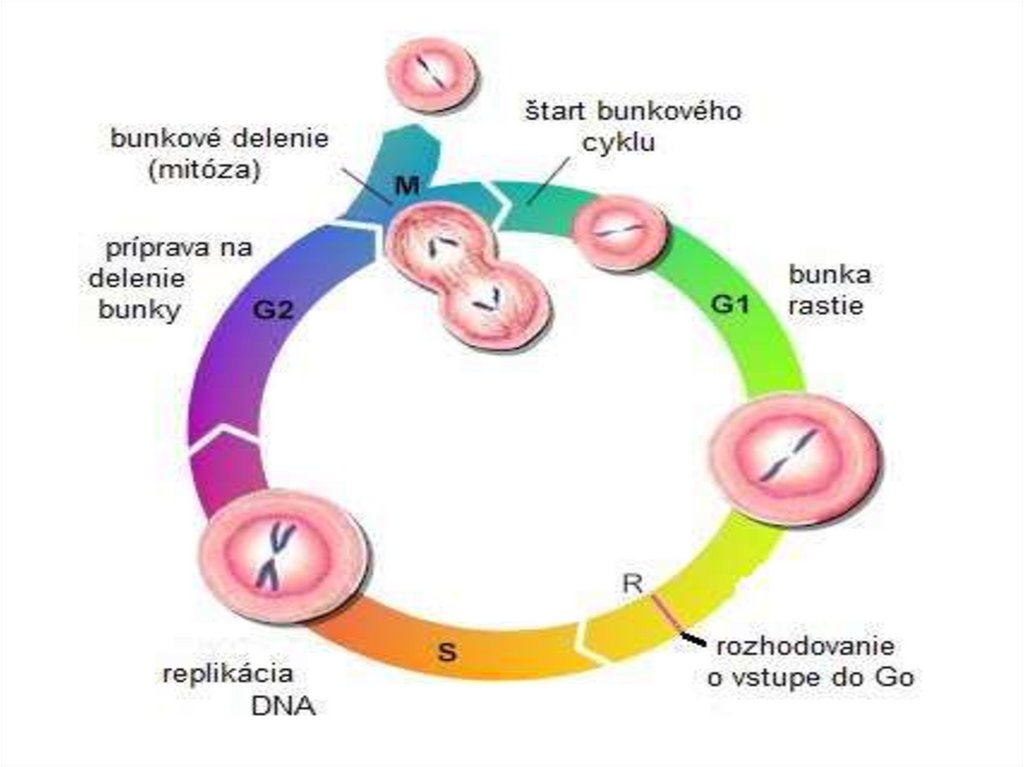
Bunkový cyklus2.
Bunkový cyklusbunka sa rozmnožuje vykonávaním usporiadaného
sledu úkonov = pritom zväčšuje svoj obsah a delí sa
na dve bunky = bunkový cyklus
je
charakterizovaný
sériou
udalostí
v
eukaryotickej bunke medzi dvoma bunkovými
deleniami
jednobunkové organizmy – bunkovým delením
vzniká celý nový organizmus
mnohobunkové organizmy – potrebné viaceré
bunkové delenia – postupná diferenciácia – tkanivá a
orgány
3.
Eukarytický bunkový cyklus – dve základné časti1) Interfáza – najdlhšie obdobie, bunka sa nedelí, dochádza
ku duplikácii celého obsahu bunky
Výsledok: dvojnásobná veľkosť bunky
- delí sa na tri časti: G1, S a G2-fázu
2) M-fáza – dochádza ku rozdeleniu jadra (karyokinéza) a
rozdeleniu na dve dcérske bunky (cytokinéza)
časové trvanie = dĺžka bunkového cyklu = generačná
doba bunky
dĺžku ovplyvňuje predovšetkým G1 fáza (epitelové
bunky črevnej
výstelky, prekurzorové krvné bunky sa delia raz za
niekoľko dní,
pečeňové bunky raz za rok, svalové a nervové vôbec
4.
5.
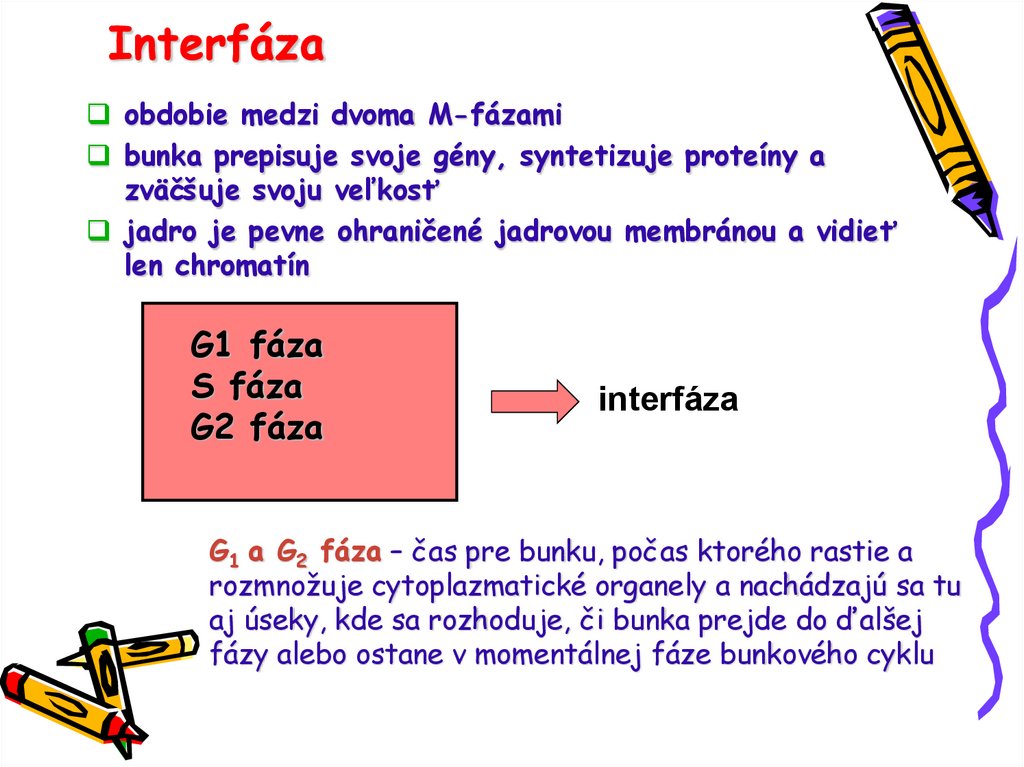
Interfázaobdobie medzi dvoma M-fázami
bunka prepisuje svoje gény, syntetizuje proteíny a
zväčšuje svoju veľkosť
jadro je pevne ohraničené jadrovou membránou a vidieť
len chromatín
G1 fáza
S fáza
G2 fáza
interfáza
G1 a G2 fáza – čas pre bunku, počas ktorého rastie a
rozmnožuje cytoplazmatické organely a nachádzajú sa tu
aj úseky, kde sa rozhoduje, či bunka prejde do ďalšej
fázy alebo ostane v momentálnej fáze bunkového cyklu
6.
G1 fázaZačína hneď po skončení mitózy (predchádzajúceho
delenia)
Najdlhšia a časovo najvariabilnejšia fáza bun. cyklu
(epiteliálne bunky G1 7-170 hodín)
V G1 fáze bunka vykonáva obvyklé metabolické aktivity a
pripravuje sa na replikáciu DNA.
intenzívne syntetické procesy - syntéza RNA a
proteosyntéza
Zvyšovanie počtu bunkových organel (mitochondrie,
ribozómy, DER atď.) bunka rastie
Vytváranie zásob nukleotidov a enzýmov potrebných pre
replikáciu DNA v nasledujúcej S - fáze
kontrola genómu bunky – reparačné mechanizmy
odstraňujú chyby genetickej informácie – poškodenia, ktoré
vznikli v DNA v priebehu mitózy
G1- checkpoint (bod bez návratu)
30 – 40% z trvania bunkového cyklu
7.
S – fáza (S=syntéza)V S fáze dochádza k replikácii
jadrovej DNA
rýchlosť syntézy DNA v
eukarytických chromozómoch je
50 nukleotidov za sekundu
replikujúca sa DNA tvorí stále
súčasť chromozómov
súčasne sa teda syntetizujú aj
históny
30 – 50 % trvania bunkového
cyklu
Extranukleárna DNA
(mitochondrie a chloroplasty) sa
replikuje počas celého bunkového
cyklu, nezávisle na jadrovej DNA a
S fáze.
8.
G2 - fázav G2 fáze bunka ďalej rastie, prebieha v nej
metabolizmus a pripravuje sa na následnú mitózu
v tejto fáze bunka obsahuje dvojnásobné množstvo
DNA,
každý chromozóm sa skladá z dvoch identických
molekúl DNA teda z dvoch sesterských chromatíd
intenzívna proteosyntéza a syntéza RNA
zvyšovanie počtu bunkových organel
syntetické procesy pripravujú bunku na mitotické
delenie
G2 – checkpoint (regulácia bunkového cyklu)
10 – 20 % trvania bunkového cyklu
9.
M – fáza = MitózaMitóza delia sa výhradne eukaryotické
bunky
najčastejší typ delenia somatických eukar. buniek
dej, pri ktorom bunka rozdelí svoj zreplikovaný genóm na
dve identické polovice
cieľ: rozdeliť zreplikované chromozómy, aby obe
dcérske bunky získali kompletné sady chromozómov
zvyčajne je nasledovaná cytokinézou, pri ktorej sa
rozdelí cytoplazma a bunková membrána. Tak vniknú dve
dcérske bunky s približne rovnakou distribúciou
bunkových organel a iných bunkových súčastí.
mitóza a cytokinéza mitotická (M) fáza bunkového
cyklu
výsledkom mitotického delenia je vznik dvoch geneticky
rovnakých dcérskych buniek z jednej bunky materskej
10.
Regulácia bunkového cyklujednotlivé udalosti bunkového cyklu (BC) sa objavujú v
presne určenom poradí a časovej následnosti (S, G2,
M, G1 fáza)
systém regulácie BC musí v správnom čase aktivovať
enzýmy a proteíny pre uskutočnenie každého procesu a
po jeho dokončení ich inaktivovať
každý krok musí byť dokončený pred začatí ďalšieho
kroku
na komlexnej činnosti riadiaceho systému sa podieľajú
mechanizmy s protichodným účinkom – stimulujú aj
inhibujú bunkovú proliferáciu vplyvom vonkajších a
vnútorných faktorov
rovnováha stimulujúcich a inhibujúcich mechanizmov –
inak napr. nekontrolovateľné delenie buniek – vznik
nádorov
11.
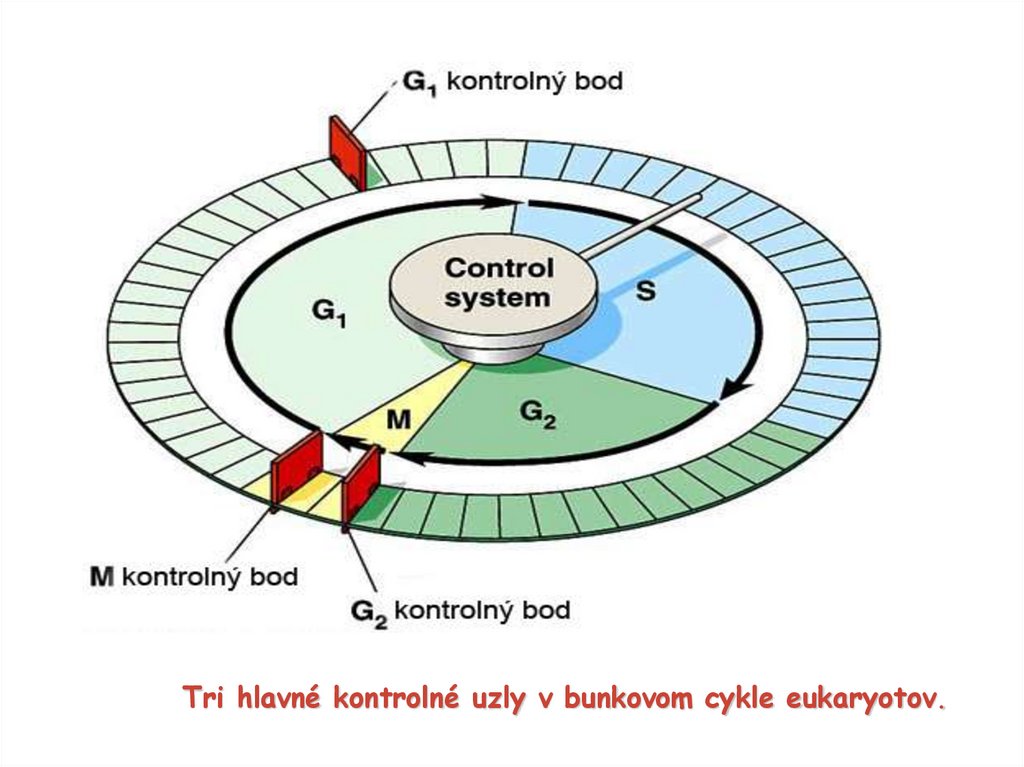
Tri hlavné kontrolné uzly v bunkovom cykle eukaryotov.12.
G1 kontrolný uzol – (hlavný kontrolný uzol – bod, z ktoréhoniet návratu) rozhodnutie, či bunka prejde do ďalšej fázy
BC a bude sa deliť a dôjde k replikácii DNA – kontrolný bod
pre replikáciu DNA alebo nastúpi do kľudového štádia,
označovaného ako G0 fáza
kontrola veľkosti bunky, dostatku živín, prítomnosti rast.
faktorov a poškodenia DNA
kontrolný uzol pre replikáciu
G0 fáza
proteosyntéza redukovaná na 30 % oproti iným fázam
zastavená expresia génov potrebných pre reguláciu a ďalšie
pokračovanie BC
dĺžku zastavenia BC v tomto bode ovplyvňujú pozitívne a
negatívne signály
pre cicavčie bunky sú pre následné pokračovanie BC
potrebné signály ostatných buniek – rastové faktory
(mitogény)
ak tieto signály nemajú, BC sa zastavuje v G1 fáze a bunky
vstupujú do stavu G0
ak už raz bunka prejde G1 kontrolným bodom, je
predurčená k dokončeniu celého BC
13.
G2 kontrolný uzol (kontrolný uzol pre karyokinézu) –replikácia DNA úspešná ? a spustenie prechodu do M
fázy mitóza
- kontrola veľkosti buniek a úspešnosti replikácie, rozhoduje sa, či
dôjde k deleniu jadra
M kontrolný uzol (kontrolný uzol pre cytokinézu) – nachádza
sa v metafáze, kontrola úspešnosti priebehu mitózy
opustenie mitózy a začiatok cytokinézy prechod do
G1 fázy
- kontrola pripojenia všetkých chromozómov na deliace
vretienko, rozhoduje sa o tom, či dôjde k deleniu bunky
až po prekonaní týchto bodov môže dôjsť k prechodu do
ďalšej fázy BC
na kontrole BC v týchto bodoch sa podieľajú faktory, ktoré
na základe spätnej väzby pôsobia tak, že regulačný systém
nedovolí spustenie nasledujúcej fázy, ak neboli dokončené
reakcie predchádzajúcej fázy (napr. zastavenie v G2, pokiaľ
nebola ukončená replikácia DNA)
detekčný systém na odhalenie abnormálnych alebo neúplných
procesov BC (napr. poškodenie DNA)
14.
Mitóza je kontinuálny proces. Delenie na jednotlivé fázyslúži len na lepší popis a orientáciu.
Mitóza sa obyčajne delí na päť fáz:
Profáza
Prometafáza
Metafáza
Anafáza
Telofáza
5 – 10% trvania bunkového cyklu
(najkratšia fáza)
15.
ProfázaPočas profázy možno pozorovať
2 základné procesy:
kondenzácia všetkých chromozómov, stávajú sa viditeľné
(každý z chromozómov sa skladá z dvoch sesterských
chromatíd spojených spolu v mieste centroméry)
vznik mitotického aparátu bunky (cytoplazmatické
mikrotubuly depolymerizujú na molekuly tubulínu)
postupné vymiznutie jadierka
centrozóm – zdvojenie na konci interfázy, v profáze sa
vzďaľujú na opačné póly bunky (ich súčasťou sú centrioly)
presúvanie centriolov k opačným pólom bunky
agregáciu rôznych proteínov okolo oboch centriolov
(centrosféra, astrálne mikrotubuly, astrosféra)
zhromažďovanie siete mikrotubulov pri centrozómoch,
počiatok vzniku deliaceho vretienka (MTOC) – polárne
mikrotubuly
Rastliny neobsahujú centrioly
16.
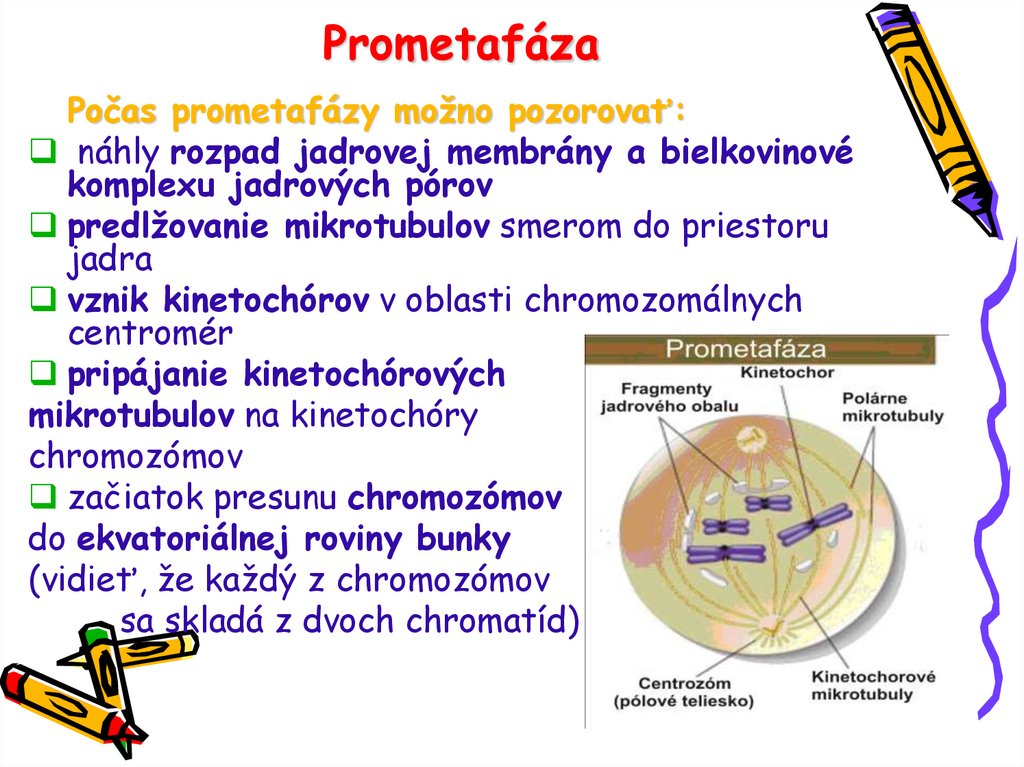
PrometafázaPočas prometafázy možno pozorovať:
náhly rozpad jadrovej membrány a bielkovinové
komplexu jadrových pórov
predlžovanie mikrotubulov smerom do priestoru
jadra
vznik kinetochórov v oblasti chromozomálnych
centromér
pripájanie kinetochórových
mikrotubulov na kinetochóry
chromozómov
začiatok presunu chromozómov
do ekvatoriálnej roviny bunky
(vidieť, že každý z chromozómov
sa skladá z dvoch chromatíd)
17.
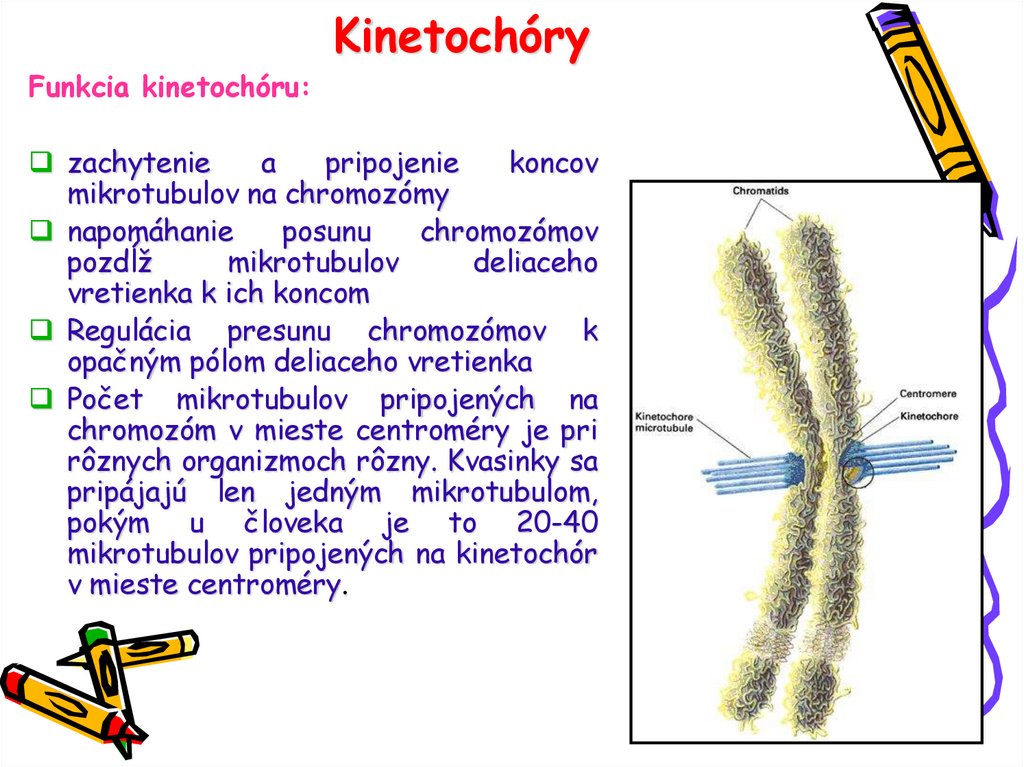
KinetochóryFunkcia kinetochóru:
zachytenie
a
pripojenie
koncov
mikrotubulov na chromozómy
napomáhanie
posunu
chromozómov
pozdĺž
mikrotubulov
deliaceho
vretienka k ich koncom
Regulácia presunu chromozómov k
opačným pólom deliaceho vretienka
Počet mikrotubulov pripojených na
chromozóm v mieste centroméry je pri
rôznych organizmoch rôzny. Kvasinky sa
pripájajú len jedným mikrotubulom,
pokým u človeka je to 20-40
mikrotubulov pripojených na kinetochór
v mieste centroméry.
18.
Metafázamaximálnu kondenzáciu chromozómov (najvhodnejšie na
pozorovania svetelným mikroskopom)
usporiadanie všetkých chromozómov do ekvatoriálnej roviny bunky
(stred bunky)
deliace vretienko tvorené polárnymi mikrotubulami
sesterské chromatídy sa od seba oddeľujú, ale ostávajú spojené
v oblasti centroméry
Každá zo sesterských chromatíd sa
kinetochórovými mikrotubulmi pripája
k opačnému
pólu deliaceho vretienka
19.
Anafázarozdelenie chromozómov v mieste centroméry
priťahovanie každej z dvoch chromatíd chromozómu k opačným
pólom bunky pomocou skracovania kinetochórových
mikrotubulov (rýchlosť pohybu chromozómov 1 m za minútu)
predlžovanie bunky, oba póly bunky
sa pohybujú od seba predlžovanie
polárnych mikrotubulov
(tiež to napomáha separácii
chromozómov)
sesterské chromatídy sa
po oddelení v anafáze stávajú
nezávislými dcérskymi chromozómami
lokalizovaných pri každom póle
deliaceho vretienka
20.
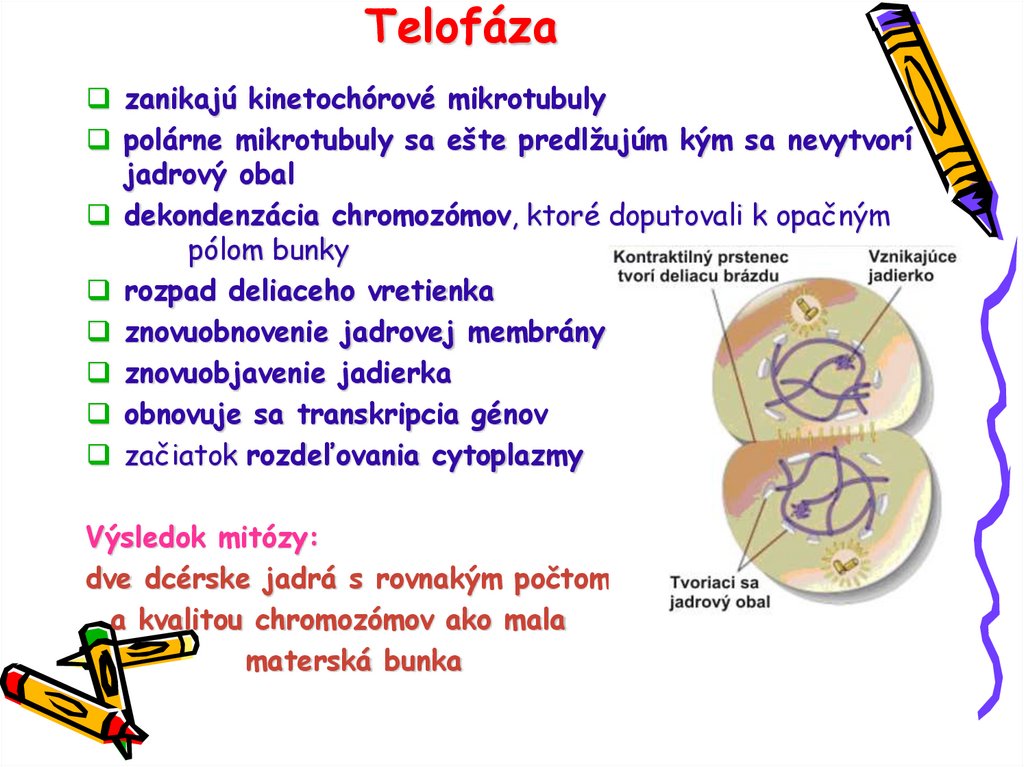
Telofázazanikajú kinetochórové mikrotubuly
polárne mikrotubuly sa ešte predlžujúm kým sa nevytvorí
jadrový obal
dekondenzácia chromozómov, ktoré doputovali k opačným
pólom bunky
rozpad deliaceho vretienka
znovuobnovenie jadrovej membrány
znovuobjavenie jadierka
obnovuje sa transkripcia génov
začiatok rozdeľovania cytoplazmy
Výsledok mitózy:
dve dcérske jadrá s rovnakým počtom
a kvalitou chromozómov ako mala
materská bunka
21.
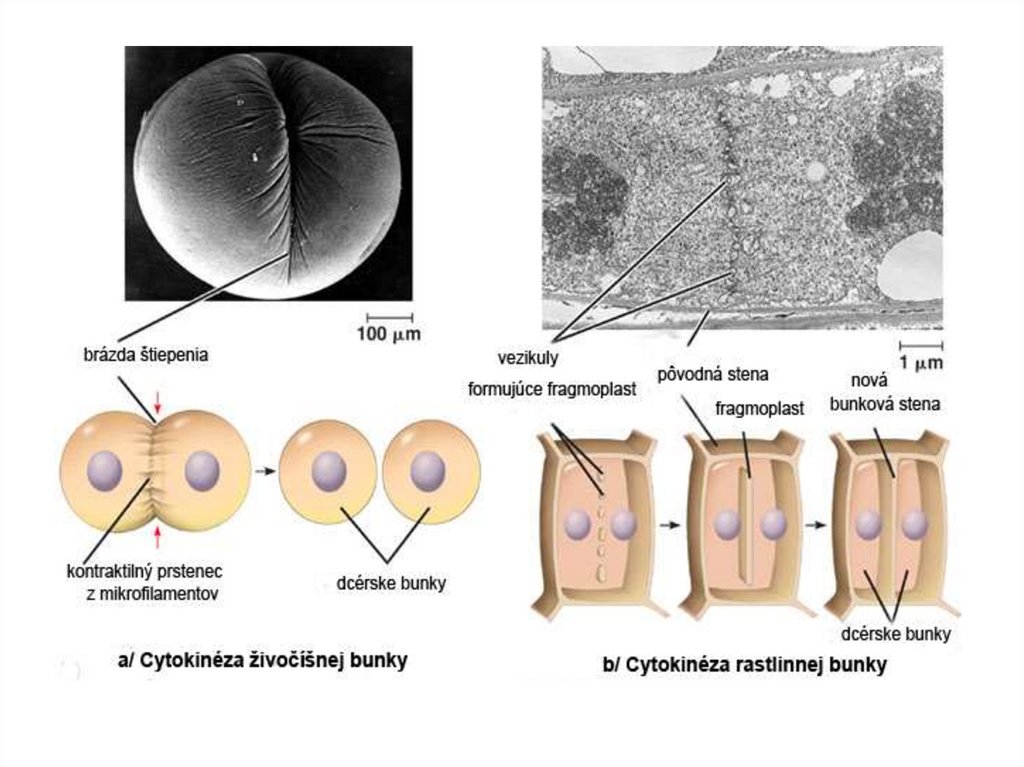
Cytokinézafyzické rozdelenie cytoplazmy a vznik dvoch dcérskych buniek
materská bunka sa rozdelí na dve dcérske bunky schopné
samostatnej existencie
odlišný priebeh u živočíšnych a rastlinných buniek
Živočíšna bunka:
začiatok krátko po rozdelení chromatíd v anafáze mitózy
kontraktilný prstenec – vlákna nesvalového myozínu II a
aktínové filamenty (ekvatoriálne zhlukovanie v blízkosti
plazmatickej membrány bunky
kontrakcia prstenca a plazmatickej membrány
zaškrcovanie cytoplazmy (centripetálne)
22.
Rastlinná bunka:vznik bunkovej steny je riadený útvarom fragmoplast
(formuje sa zo zbytkov polárnych mikrotubulov, ktoré
zostávajú v mieste ekvatoriálnej roviny)
presun vezikúl vzniknutých z Golgiho aparátu podĺž
mikrotubulov do centra bunky (obsahujú polysacharidy a
glykoproteíny potrebné pre matrix bunkovej steny)
vezikuly sa zlievajú a fragmoplast zväčšuje centrifugálne
(smerom k povrchu bunky) až kým nedôjde k spojeniu s
cytoplazmatickou membránou a pôvodnou bunkovou stenou
rozdelenie na dve dcérske bunky
23.
24.
MEIÓZAšpecifické delenie –– vznik haploidných buniek (gamét)
Dve po sebe nasledujúce delenia:
prvé meiotické delenie
– heterotypické - redukčné
druhé meiotické delenie – homeotypické - ekvačné
Prvé meiotické delenie:
profáza
metafáza
anafáza
telofáza
25.
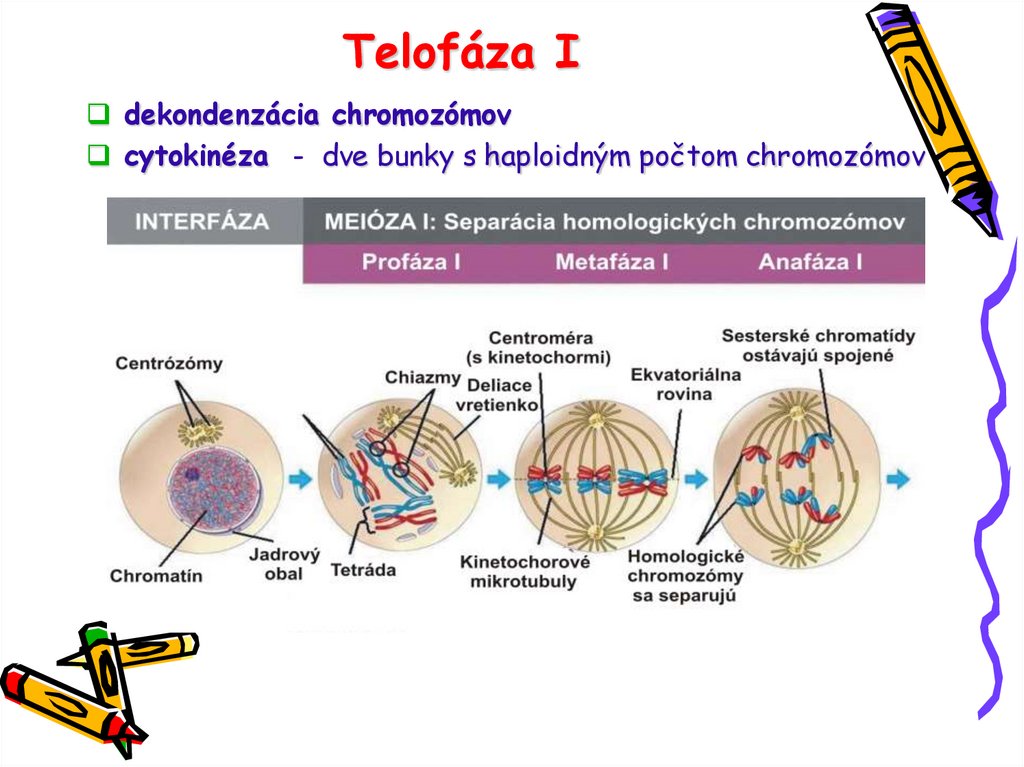
PROFÁZA Iprebieha v jadre so zachovaným jadrovým obalom a
jadierkom a jednotlivé páry homologických chromozómov
podliehajú zložitým zmenám
5 štádií
1)
Leptotén
o
o
kondenzácia chromozómov, stávajú sa viditeľné,
oboma koncami prichytené k vnútornej membráne
obalu
jadrového
2)
Zygotén
o
začína sa vytvárať synaptonemálny komplex, ktorý spája
homológne chromozómy
dochádza k tzv. synapsii a vytvárajú sa bivalenty (homologické
chromozómy sa párujú, pozdĺžne sa k sebe prikladajú)
o
26.
3) Pachyténo
o
o
pokračuje kondenzácia chromozómov
vznik tetrád (na každom chromozóme si rozlíšiteľné 4
chromatídy),
nesesterské chromatídy sa okolo seba obtáčajú, vznik
prekríženia = chiazmy --- vzájomná výmena určitých oblastí
chromozómov – crossing – over
4) Diplotén
o
o
zánik synaptonemálneho komplexu
postupné oddeľovanie homologických chromozómov
5) Diakinéza
o
o
o
o
o
o
kondenzácia chromozómov vrcholí
ukončuje sa terminalizácia chiaziem (posun na konce chromatíd)
jadrový obal sa rozpadáva
jadierko mizne
vytvára sa deliace vretienko
centrozómy dosiahli opačné póly bunky
27.
Metafáza Itetrády sa posúvajú do ekvatoriálnej roviny
centroméry náhodne orientované k opačným pólom bunky
Anafáza I
homologické chromozómy sa od seba oddeľujú, bivalenty
zanikajú
oddelené chromozómy sa na rozdiel od mitózy skladajú z
dvoch chromatíd spojených centromérou (dyády) a sú
priťahované mikrotubulmi deliaceho vretienka k opačným
pólom bunky
rozchod párových chromozómov je náhodný --- zodpovedá
Mendelovmu zákonu o voľnej kombinovateľnosti
týmto okamihom sa zakladá haploidný počet chromozómov v
každej bunke
chyby pri rozchode chromozómov --- vznik numerických
aberácií – spontánne potraty alebo vznik syndrómov
28.
Telofáza Idekondenzácia chromozómov
cytokinéza - dve bunky s haploidným počtom chromozómov
29.
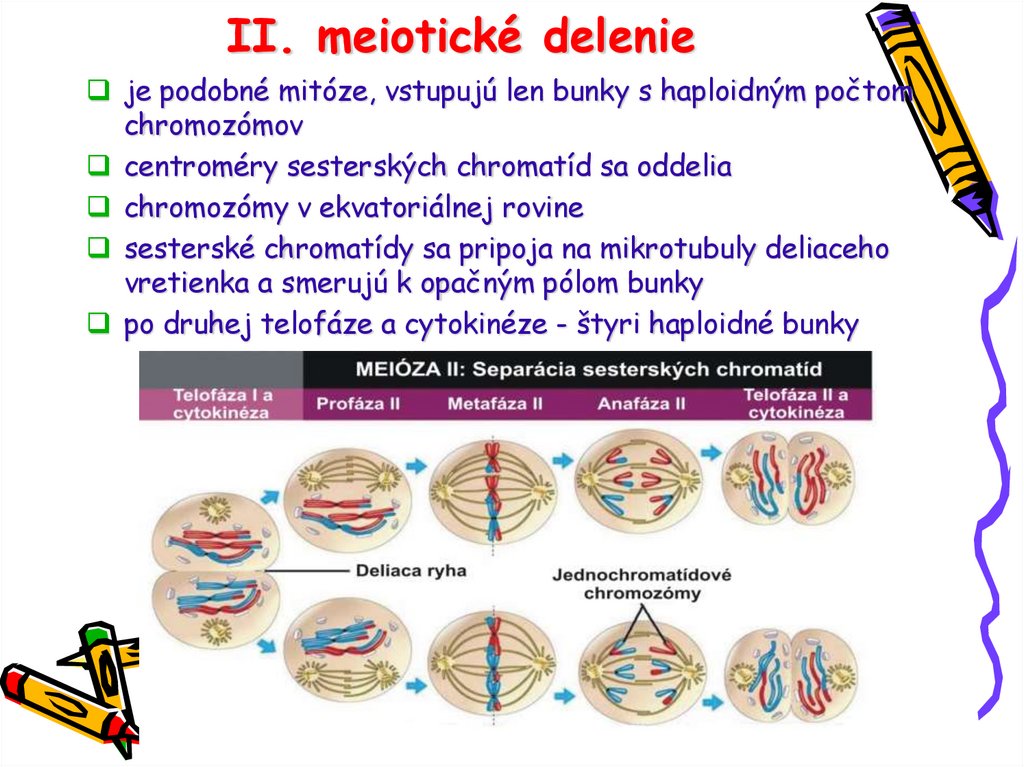
II. meiotické delenieje podobné mitóze, vstupujú len bunky s haploidným počtom
chromozómov
centroméry sesterských chromatíd sa oddelia
chromozómy v ekvatoriálnej rovine
sesterské chromatídy sa pripoja na mikrotubuly deliaceho
vretienka a smerujú k opačným pólom bunky
po druhej telofáze a cytokinéze - štyri haploidné bunky
30.
Genetický význam meiózysegregácia
a
náhodná
distribúcia
chromozómov
redukcia počtu chromozómov na polovicu
po meióze má každé jadro dcérskej bunky
náhodnú kombináciu chromozómov - t. j.
génov od matky a od otca
crossing–over
rozširuje
kombinačnú
premenlivosť v pohlavných bunkách
prostredníctvom meiózy a oplodnenia sa
udržuje rovnaký počet chromozómov
v generáciach pohlavne sa rozmnožujúcich
organizmov zachováva sa druhová stálosť