









 генетики:")









































 Биология
БиологияПохожие презентации:








")

Генетика развития растений. Меристемы растений
1.
Генетика развития растенийРазвитие =
рост + дифференцировка
Развитие клетки,
ткани, органа
(морфогенез)
2.
Меристемы растенийМеристе́мы (с др.-греч. — «μεριστός» — делимый), или образовательные
ткани, или меристематические ткани, — обобщающее название
для тканей растений, состоящих из интенсивно делящихся и сохраняющих
физиологическую
активность
на
протяжении
всей
жизни
клеток,
обеспечивающих непрерывное нарастание массы растения и предоставляющих
материал для образования различных специализированных тканей.
Сейчас меристемы обычно рассматриваются как специализированные органы.
Общие свойства меристем:
1. Содержат пулы стволовых клеток (СК). По сути представляют собой
ниши СК, способные а). Поддерживать недифференцированное
состояние клеток и б). Продуцировать клетки, способные к
дифференцировке
Наличие центральной (ЦЗ) и периферических зон (ПЗ). ЦЗ состоит из
недифференцированных клеток (СК), ПЗ - из клеток-потомков СК,
вступивших на путь дифференцировки
2. Работа определенных групп ТФ, подавляющих дифференцировку
клеток. Обычно их гены экспрессируются в организующих центрах
(котоорые есть не у всех меристем)
3.
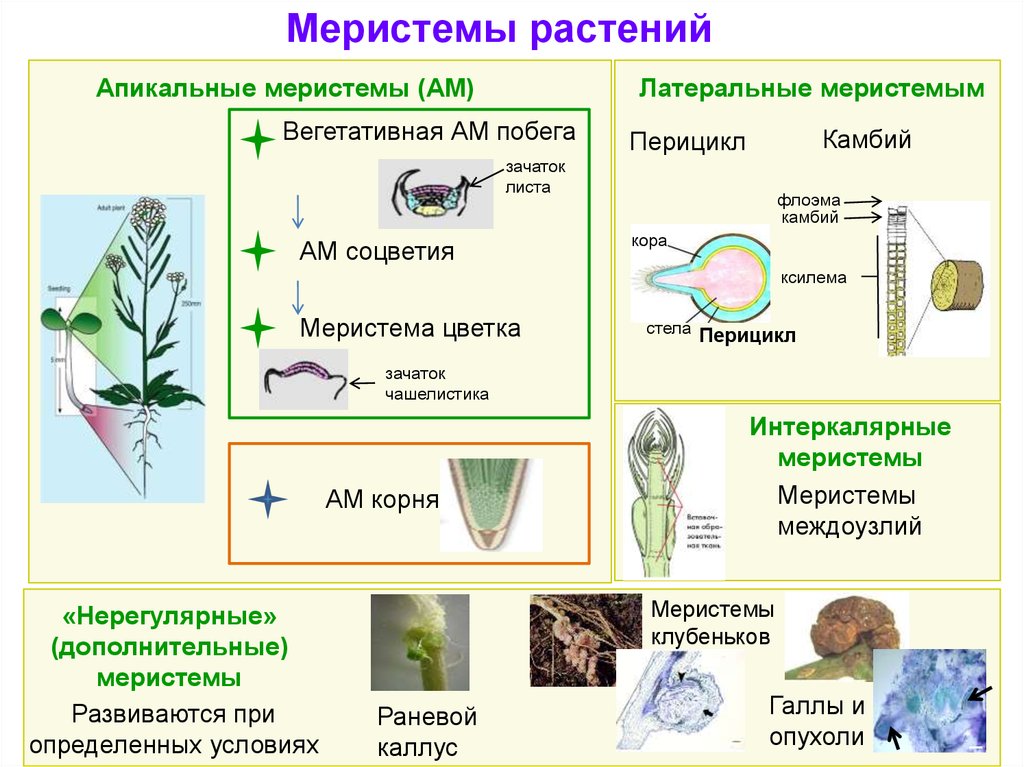
Меристемы растенийАпикальные меристемы (АМ)
Латеральные меристемым
Вегетативная АМ побега
Камбий
Перицикл
зачаток
листа
АМ соцветия
флоэма
камбий
кора
ксилема
Меристема цветка
стела Перицикл
зачаток
чашелистика
АМ корня
«Нерегулярные»
(дополнительные)
меристемы
Развиваются при
определенных условиях
Интеркалярные
меристемы
Меристемы
междоузлий
Меристемы
клубеньков
Раневой
каллус
Галлы и
опухоли
4.
Меристемы растенийПервичные - закладываются в эмбриогенезе
- SAM lineage
- procambium lineage
- RAM lineage
Вторичные – закладываются в постэмбриональном развитии
- xylem pericycle lineage
- phloem pericycle lineage
3
5.
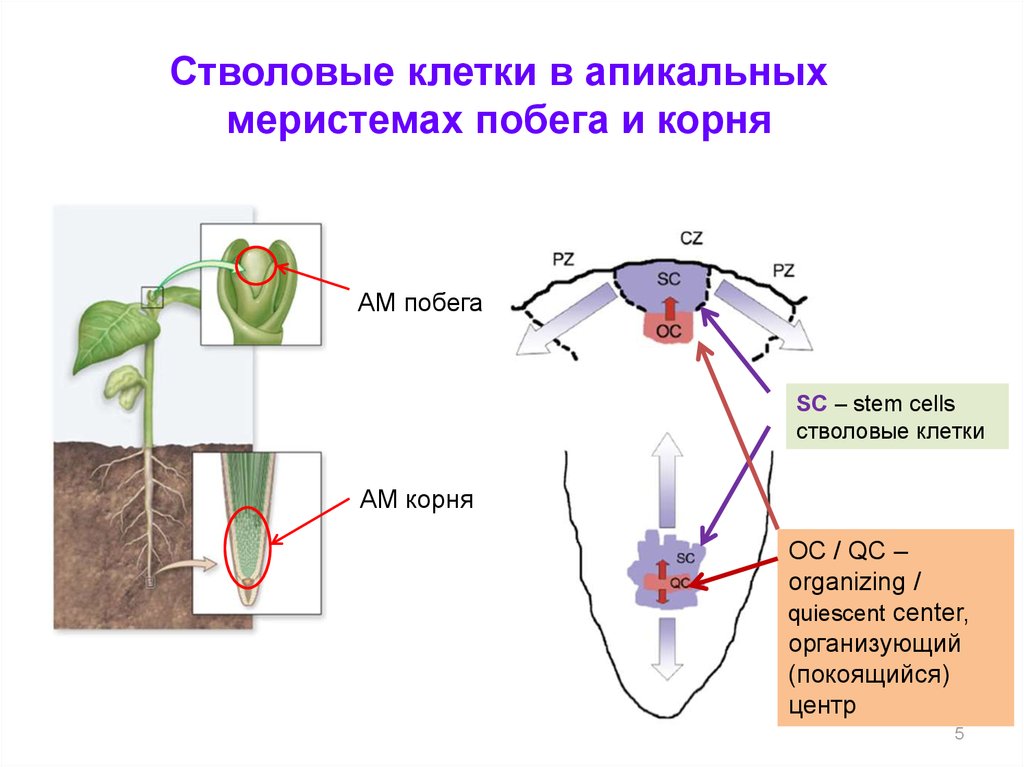
Стволовые клетки в апикальныхмеристемах побега и корня
АМ побега
SC – stem cells
стволовые клетки
АМ корня
OC / QC –
organizing /
quiescent center,
организующий
(покоящийся)
центр
5
6.
Определение стволовых клетокСтволовые клетки (stem cells):
Делящиеся клетки, потомки которых либо остаются в том
же состоянии, что и материнские клетки (обычно
называемое «недифференцированным») и таким образом
пополняют популяцию стволовых клеток, либо
претерпевают дифференцировку и дают начало новым
органам и тканям.
Ниша стволовых клеток (stem cell niche):
локальное микроокружение, среда, поставляющая
факторы, необходимые для поддержания стволовых
клеток.
6
M. R. Tucker and T. Laux, TRENDS in Cell Biology 2007
7.
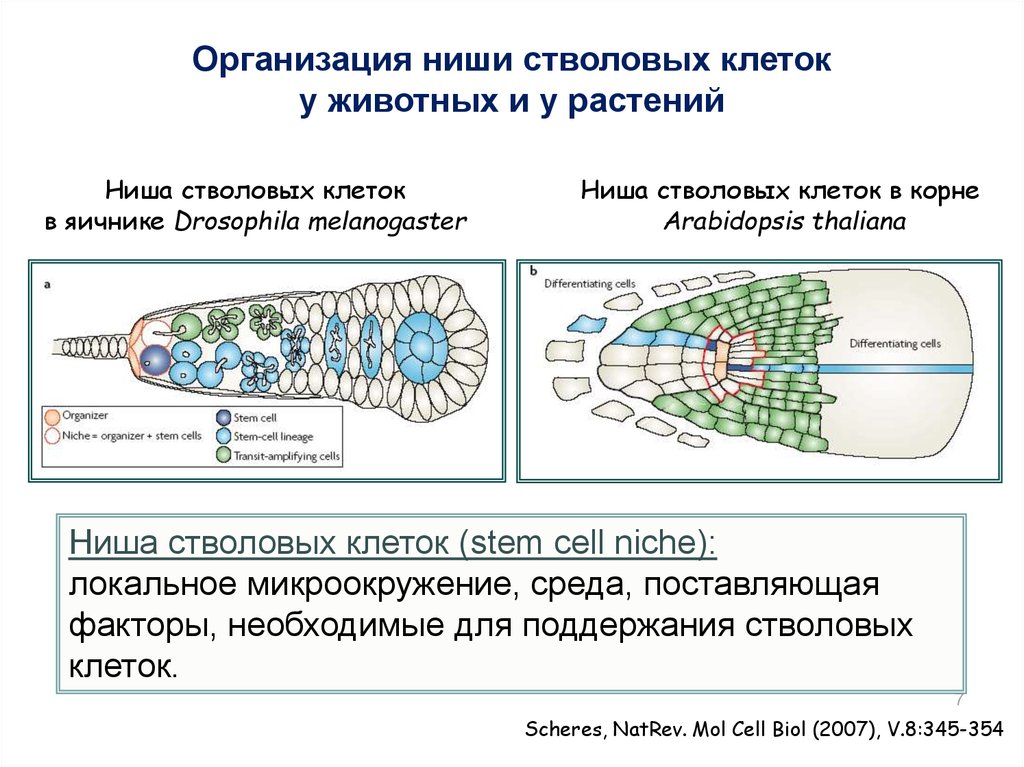
Организация ниши стволовых клетоку животных и у растений
Ниша стволовых клеток
в яичнике Drosophila melanogaster
Ниша стволовых клеток в корне
Arabidopsis thaliana
Ниша стволовых клеток (stem cell niche):
локальное микроокружение, среда, поставляющая
факторы, необходимые для поддержания стволовых
клеток.
7
Scheres, NatRev. Mol Cell Biol (2007), V.8:345-354
8.
Общий принцип организации ниши стволовых клетокдифференцированные клетки
ранние стадии дифференцировки
внутриклеточный
сигнал,
предотвращающий
дифферненцировку
недифференцированные клетки
«организующий центр»
(organizer)
8
Sablowski, Trends in Cell Biology, 2004,14(11):605-611
9.
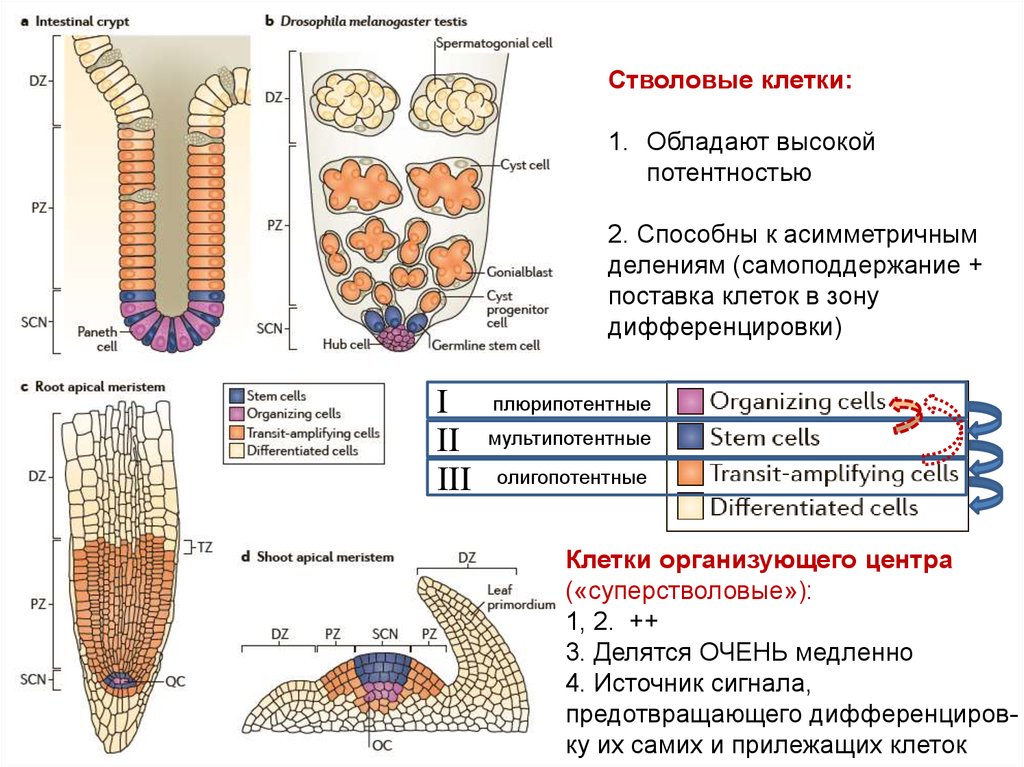
Стволовые клетки:1. Обладают высокой
потентностью
2. Способны к асимметричным
делениям (самоподдержание +
поставка клеток в зону
дифференцировки)
I
II
III
плюрипотентные
мультипотентные
олигопотентные
Клетки организующего центра
(«суперстволовые»):
1, 2. ++
3. Делятся ОЧЕНЬ медленно
4. Источник сигнала,
предотвращающего дифференцировку их самих и прилежащих клеток
10.
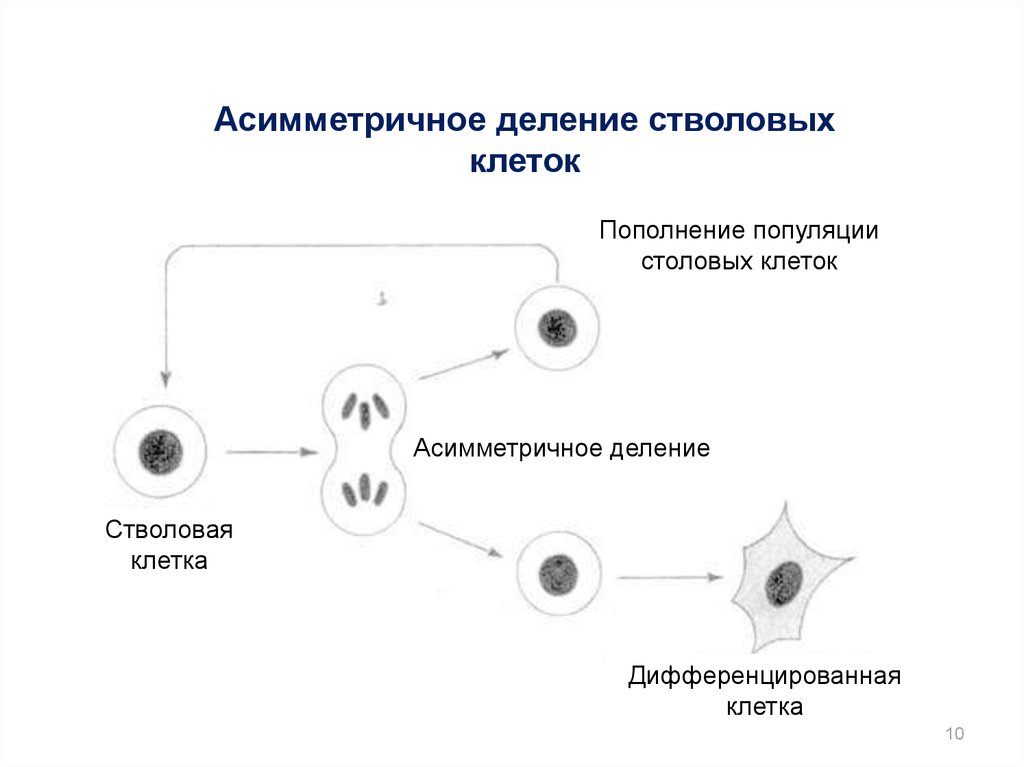
Асимметричное деление стволовыхклеток
Пополнение популяции
столовых клеток
Асимметричное деление
Стволовая
клетка
Дифференцированная
клетка
10
11.
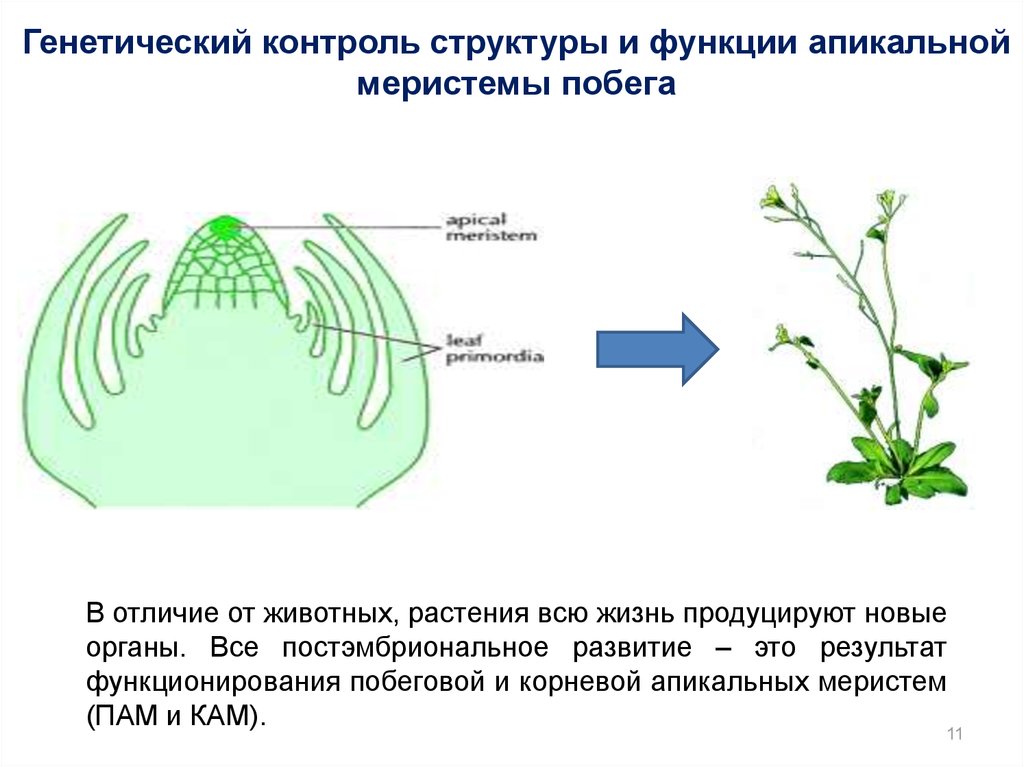
Генетический контроль структуры и функции апикальноймеристемы побега
В отличие от животных, растения всю жизнь продуцируют новые
органы. Все постэмбриональное развитие – это результат
функционирования побеговой и корневой апикальных меристем
(ПАМ и КАМ).
11
12.
Строение апикальной меристемы побегаПАМ
лист
узел
междоузлие
фитомер
12
Clark, Current Opinion in Plant Biology 2001
13.
Строение апикальной меристемы побега1. Клональное разделение: L1 + L2 (туника), L3 (корпус)
2. Гистологическое разделение (зоны): центральная, периферические и риб-зона
CZ
(central
zone,
центральная
зона)
–
стволовые клетки (в т.ч.
организующий центр) и их
ближайшие потомки. Делятся
медленно, нет признаков
дифференцировки
RZ (rib zone, риб-зона) –
зона
закладки
проводящей
системы.
Растущие и делящиеся
клетки на разных стадиях
дифференцировки.
PZ
(peripheral
zone,
периферическая зона) –
дочерние
клетки
CZ.
Скорость и направление
делений
клеток
не
постоянно; клетки способны
13
к дифференцировке.
14.
«Симпластические поля» в апикальной меристеме побега14
15.
Скорость прохождения клеточногоцикла в разных зонах ПАМ
Besnard et al., 2011
16. Распределение гормонов в ПАМ
1617.
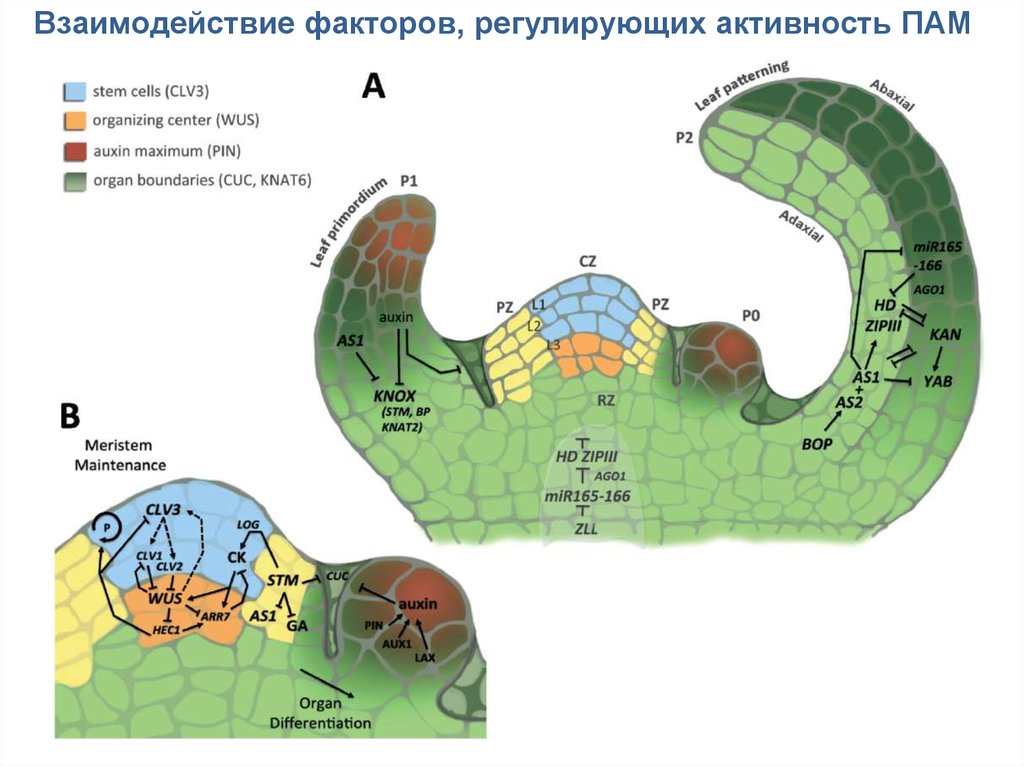
Взаимодействие факторов, регулирующих активность ПАМ17
18. Изучение генетического контроля структуры и функции ПАМ методами классической («прямой») генетики:
Выявлены два классов мутантов, имеющих противоположный фенотип:мутанты с
увеличением размера АМ
(clv1, clv2, clv3, fas1, fas2)
WT
clavata1
(clv1)
мутанты с
недоразвитием АМ
(stm, wus, zll);
wuschel
shootmeristemless
(wus)
(stm)
18
19.
Мутанты с недоразвитием АМWT
wus
stm
19
20.
Изучение взаимодействия генов STM и WUS:1. Анализ фенотипа двойных мутантов stm wus
(метод классической, прямой генетики)
(«сильные» аллели wus и stm)
stm-5/+ x
wus-1/+
По генотипу
9 +/+
: 3 wus-1/+
:
3 stm-5/+ : 1 stm-5/ wus-1
По фенотипу
9 wt
: 3 wus-1
:
4 stm-5
stm-5 wus-1 = stm-5
эпистаз
Вероятно, гены осуществляют последовательные этапы одного
процесса:
продукт STM действует раньше, чем продукт гена WUS
21.
Изучение взаимодействия генов STM и WUS:Анализ трансгенных растений с измененной экспрессией генов:
STM
WUS
Отсутствие изменений
в характере экспрессии
WUS (и CLV3)
ANT::STM
(эктопическая экспрессия
STM
в
листовых
примордиях)
WUS
35S::WUS-GR
(активация
экспрессии
WUS в меристеме)
STM
- Отсутствие изменений в
характере
экспрессии
STM и других
KNOX
генов,
- Активация экспрессии
CLV3
WUS и STM регулируют меристематическую активность клеток
независимо друг от друга
21
22.
Роль генов STM и WUS в апикальной меристеме побегаWUS
WUS
Поддержание пула
стволовых клеток в
меристеме
STM
STM
Пролиферация
стволовых клеток в
меристеме
22
23.
Мутанты по гену WUS (WUSCHEL)дикий
тип
wus
-в эмбриогенезе АМ
образуется, но
уменьшенная, состоящая из
крупных вакуолизированных
клеток;
- АМ побега формирует
несколько листьев и
прекращает свое развитие;
- на придаточных побегах
могут закладываться цветки
с нормальным числом
чашелистиков и лепестков,
но преращающих развитие
с формированием
единственной тычинки
Mayer et al., Cell 1998
Гипотеза о функции гена WUS:
ген необходим для формирования нормальной АМ в эмбриогенезе,
для поддержания клеток ЦЗ апикальной меристемы в стволовом
состоянии в процессе вегетативного и репродуктивного развития. 23
24.
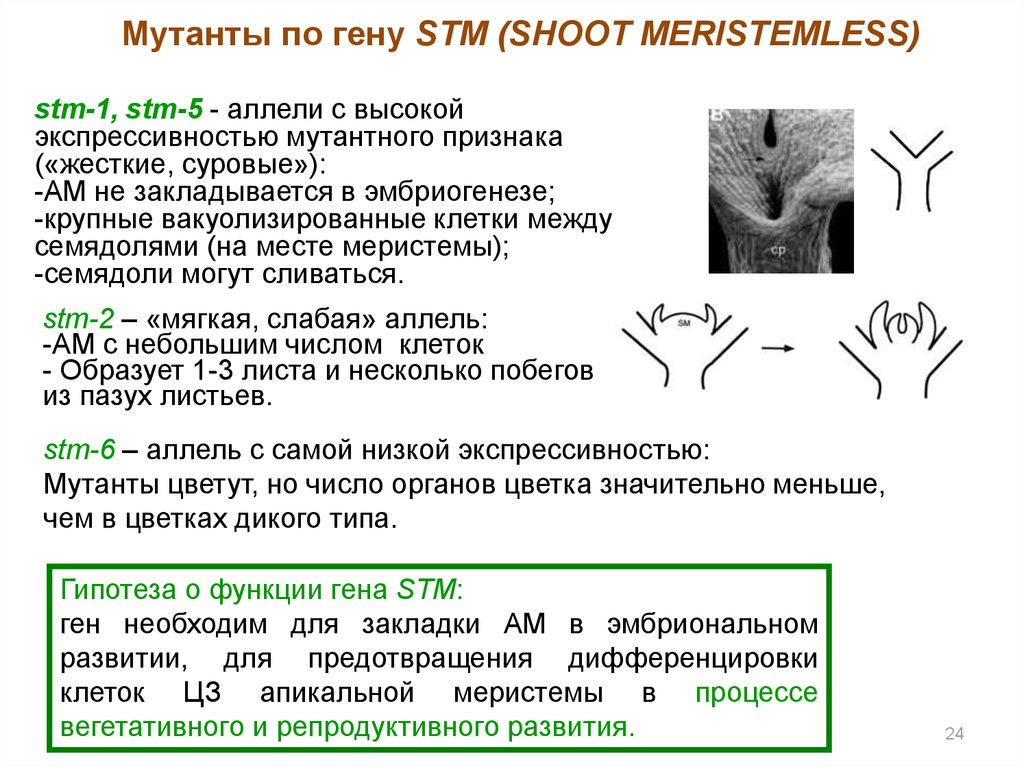
Мутанты по гену STM (SHOOT MERISTEMLESS)stm-1, stm-5 - аллели с высокой
экспрессивностью мутантного признака
(«жесткие, суровые»):
-АМ не закладывается в эмбриогенезе;
-крупные вакуолизированные клетки между
семядолями (на месте меристемы);
-семядоли могут сливаться.
stm-2 – «мягкая, слабая» аллель:
-АМ с небольшим числом клеток
- Образует 1-3 листа и несколько побегов
из пазух листьев.
stm-6 – аллель с самой низкой экспрессивностью:
Мутанты цветут, но число органов цветка значительно меньше,
чем в цветках дикого типа.
Гипотеза о функции гена STM:
ген необходим для закладки АМ в эмбриональном
развитии, для предотвращения дифференцировки
клеток ЦЗ апикальной меристемы в процессе
вегетативного и репродуктивного развития.
24
25.
Мутанты с увеличенной АМWT
clv1
25
26.
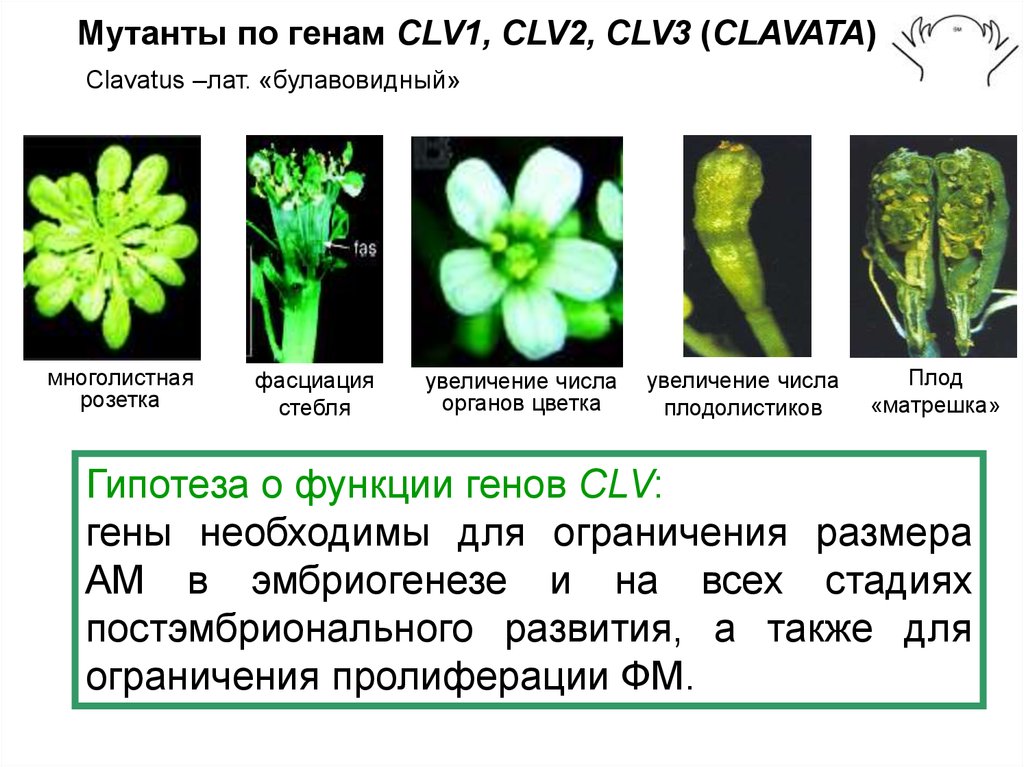
Мутанты по генам СLV1, CLV2, CLV3 (CLAVATA)Clavatus –лат. «булавовидный»
многолистная
розетка
фасциация
стебля
увеличение числа
органов цветка
увеличение числа
плодолистиков
Плод
«матрешка»
Гипотеза о функции генов CLV:
гены необходимы для ограничения размера
АМ в эмбриогенезе и на всех стадиях
постэмбрионального развития, а также для
ограничения пролиферации ФМ.
27.
Экспрессия генов STM, WUS, CLV1 и CLV3 в апикальноймеристеме побега
WUS – в ОЦ
CLV1 – в ОЦ и рядом с ним,
CLV3 – в L1, L2 над ОЦ
27
28.
Экспрессия генов STM и WUS в апикальноймеристеме побега
WUS
STM
Bowman J., Yuval E. Trends in Plant Science, 2000, V. 5., N. 3, pp. 110-115
28
29.
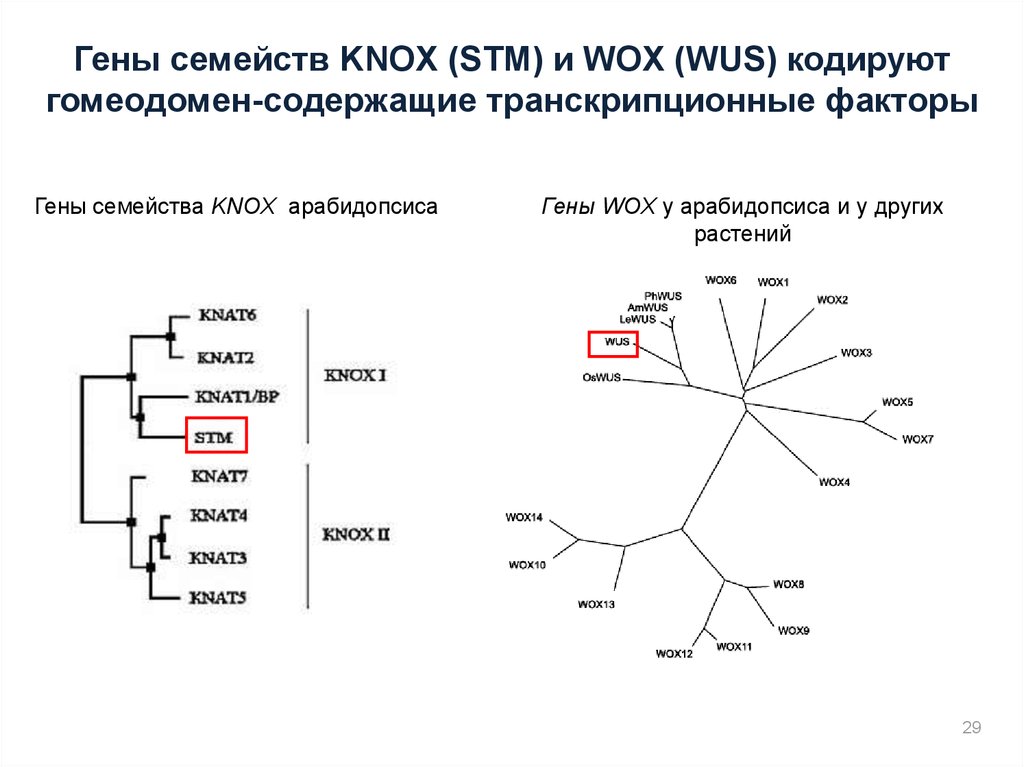
Гены семейств KNOX (STM) и WOX (WUS) кодируютгомеодомен-содержащие транскрипционные факторы
Гены семейства KNOX арабидопсиса
Гены WOX у арабидопсиса и у других
растений
29
30.
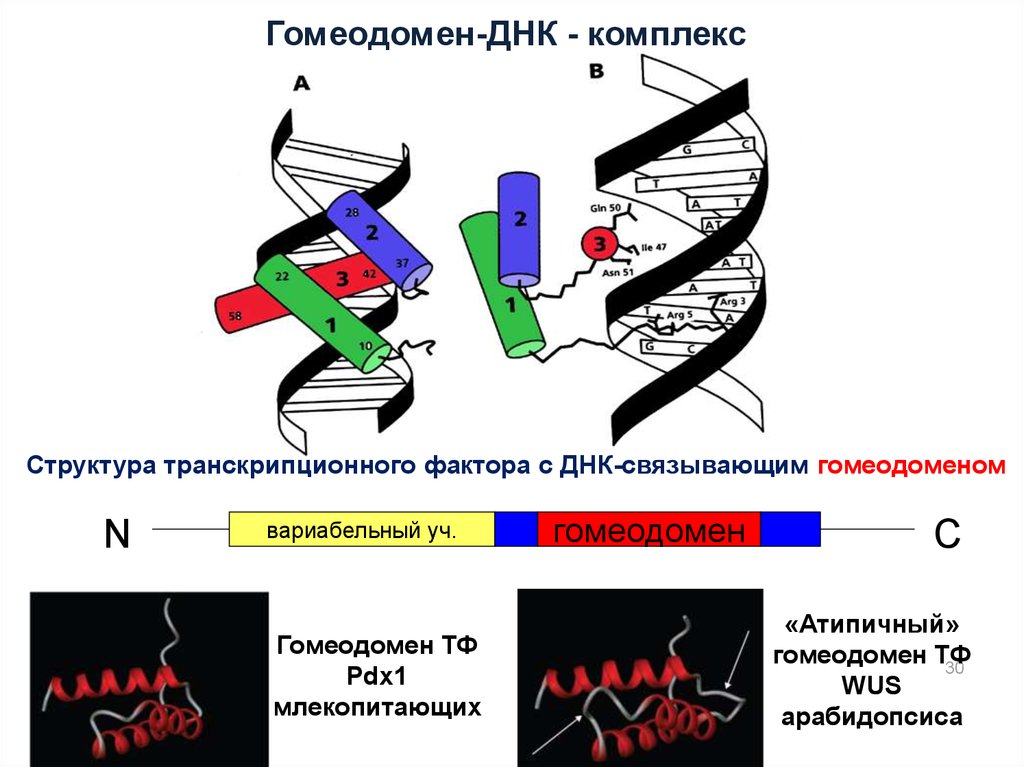
Гомеодомен-ДНК - комплексСтруктура транскрипционного фактора с ДНК-связывающим гомеодоменом
N
вариабельный уч.
Гомеодомен ТФ
Pdx1
млекопитающих
гомеодомен
С
«Атипичный»
гомеодомен ТФ
30
WUS
арабидопсиса
31.
Экспрессия гена STM в АМ побегаСхема доменной организации белка STM
N
KNOX
GSE
ELK
HD
C
Выявление экспрессии гена STM на разных этапах развития АМ
методом гибридизации in situ
31
Moussian et al., The EMBO Journal 199
32.
Транспорт продуктов генов KNOX в апикальноймеристеме побега
32
33.
Нарушениетранспорта белка
STM вызывает
дефекты
развития ПАМ
Plant Journal, 2017
34.
ТФ BELL (тоже из семейства TALE) – партнеры ТФ KNOX(обеспечивают транспорт KNOX в ядро)
35.
Мишени STMЭкспрессия гена AS1 (Asymmetric leaves1), отвечающего за развитие
листового примордия, у мутантов stm-1
дикий тип
stm-1
Взаимодействие STM
и AS1-AS2 в апексе
побега
35
36.
Модель эпигенетической регуляции генов KNOX вдифференцированных клетках листьев
KNOX promoter
wt
as1
hira
Lodha et al., 2013
37.
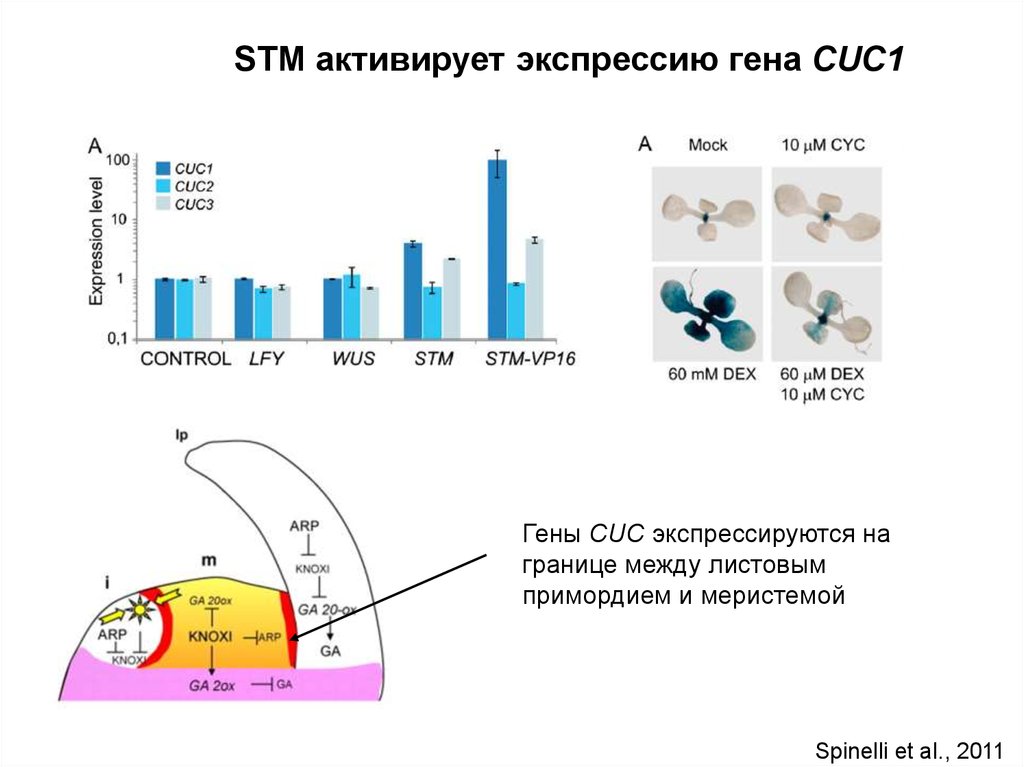
STM активирует экспрессию гена CUC1Гены CUC экспрессируются на
границе между листовым
примордием и меристемой
Spinelli et al., 2011
38.
Мишени STM в ПАМSTM
STM
IPT (биосинтез цитокинина)
GA2ox (дезактивация гибереллинов)
38
Jasinski et al. 2005
39.
Мишени STM в ПАМВзаимодействие STM (генов KNOX) и генов, вовлеченных в метаболизм
цитокинина и гиббереллина в апексе побега
39
Jasinski et al. 2005
40.
Активация экспрессии KNOX важна дляформирования сложных листьев
41.
Транскрипционные факторы семейства WOXклада «WUS»
гомеодомен
WUS-бокс (репрессия транскрипции)
«промежуточная»
клада
домен EAR
(рекрутирование
корепрессоров
трнаскрипции)
«древняя» клада
41
42.
Экспрессия гена WUS в АМ побегаЭкспрессия
гена WUS в
организующем
центре
меристемы
Ядерная локализация
гибридного белка WUSGUS
(в клетках эпидермиса
лука)
42
Mayer et al., Cell 1998
43.
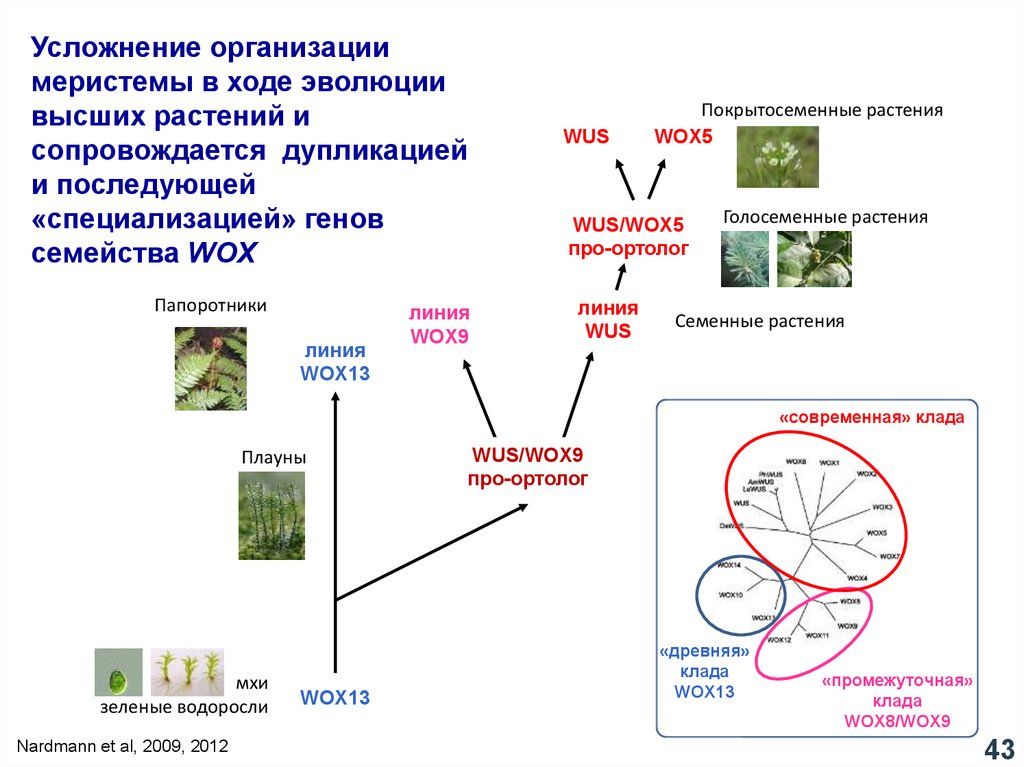
Усложнение организациимеристемы в ходе эволюции
высших растений и
сопровождается дупликацией
и последующей
«специализацией» генов
семейства WOX
Папоротники
линия
WOX13
линия
WOX9
WUS
Покрытосеменные растения
WOX5
WUS/WOX5
про-ортолог
линия
WUS
Голосеменные растения
Семенные растения
«современная» клада
Плауны
мхи
зеленые водоросли
Nardmann et al, 2009, 2012
WOX13
WUS/WOX9
про-ортолог
«древняя»
клада
WOX13
«промежуточная»
клада
WOX8/WOX9
43
44.
Роль про-ортолога WUS/WOX5 у голосеменныхрастений
Геном Gnetum gnemon –
содержит про-ортолог
GgWUS/WOX5 (до дупликации
генов WUS/WOX5 в ходе
эволюции), экспрессирующийся
как в корне, так и в побеге
wt
pWUS-GgWUS/WOX5
Экспрессия pWUS-GgWUS/WOX5
у арабидопсиса обуславливает clvподобный фенотип (фасциация стебля,
формирование многочисленных цветков и
органов цветка)
Nardmann et al, 2009
44
45.
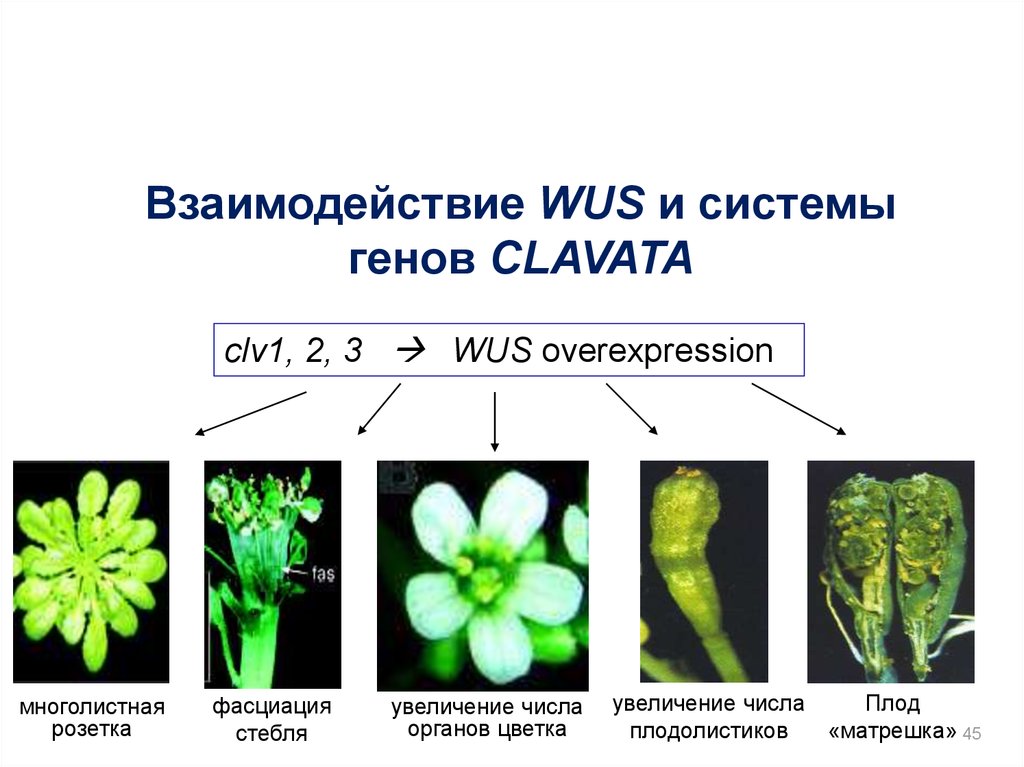
Взаимодействие WUS и системыгенов CLAVATA
clv1, 2, 3 WUS overexpression
многолистная
розетка
фасциация
стебля
увеличение числа
органов цветка
увеличение числа
Плод
плодолистиков
«матрешка» 45
46.
Регуляция экспрессии гена WUS в АМ с участием белковCLAVATA
CLV3
CLV1
CLV2
CLV1 и CLV2 кодируют рецепторные белки (c
лейцин-богатым LRR–доменом), работающие в
виде гетеродимера CLV1/CLV2
CLV3 – лиганд 12 а/к, связывается
рецепторным комплексом CLV1/CLV2
46
с
47.
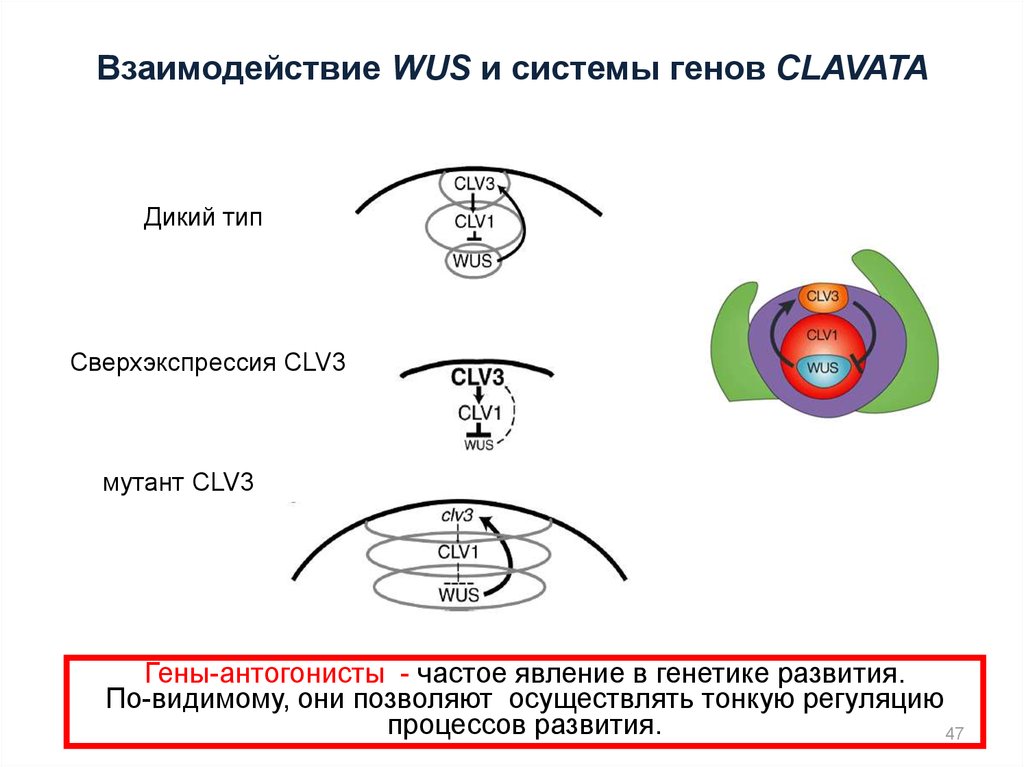
Взаимодействие WUS и системы генов CLAVATAДикий тип
Сверхэкспрессия CLV3
мутант CLV3
Гены-антогонисты - частое явление в генетике развития.
По-видимому, они позволяют осуществлять тонкую регуляцию
процессов развития.
47
48.
Взаимодействие WUS и системы CLAVATAУвеличение домена
экспрессии CLV3 в
растениях ANT::WUS
WUS
дикий тип
мутант clv3
CLV3
CLV3
35S::CLV3
WUS
49.
Экспрессия генов CLV1, CLV3 и WUS в апикальноймеристеме побега
Экспрессия
WUS в ОЦ
CLV1 «защищает» ОЦ от негативного влияния CLV3, связывая его
49
50.
Рецепторные киназы растений c LRR–доменом(Leucine-rich repeat receptor kinases)
Структура гена рецепторной LRR-рецепторной киназы
50
Haffani et al., Can. J. Bot. 2004
51.
Эксперименты по выявлению новых компонентовсигнального пути, опосредованного CLV3
coryne (crn)
Muller et al., 2008
Получение новых мутаций на
фоне сверхэкспрессии CLV3
Известные рецепторы CV3 в ПАМ:
1. CLV1
2. CLV2+CRN
3. RPK2
CORYNE – рецепторная киназа с
коротким внеклеточным доменом
51
52.
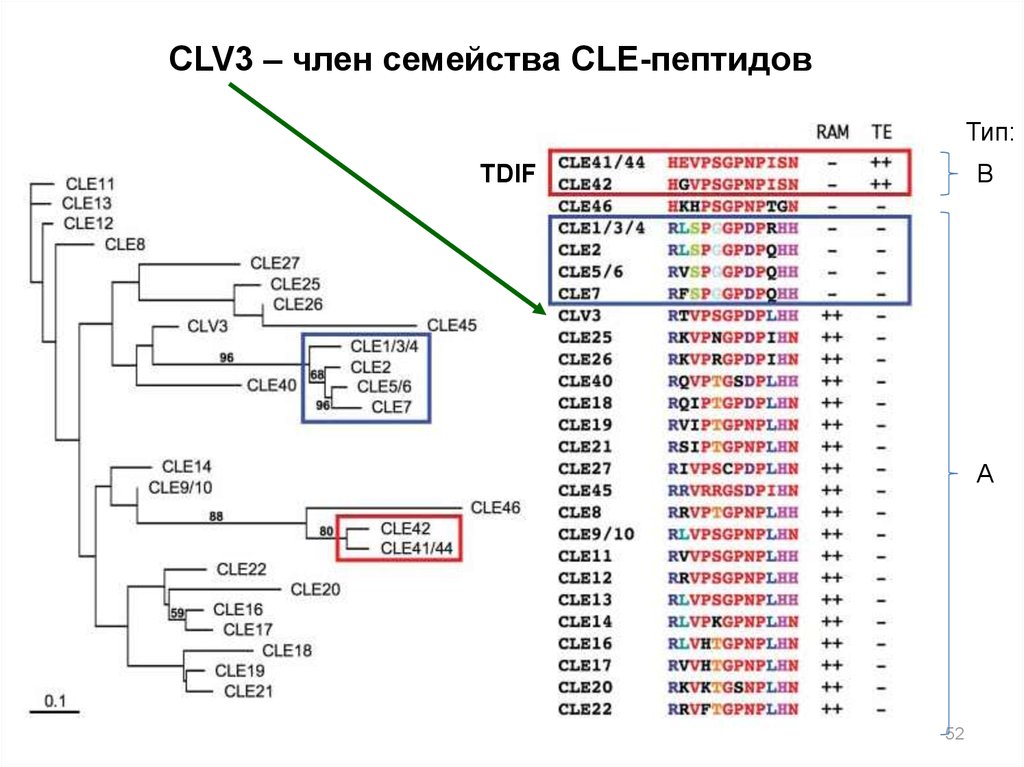
CLV3 – член семейства CLE-пептидовТип:
В
TDIF
А
52
53.
Участие CLE-пептидов в развитии Arabidopsis©2010 by American Society of Plant Biologists
Jun J et al. Plant Physiol. 2010;154:1721-1736
54.
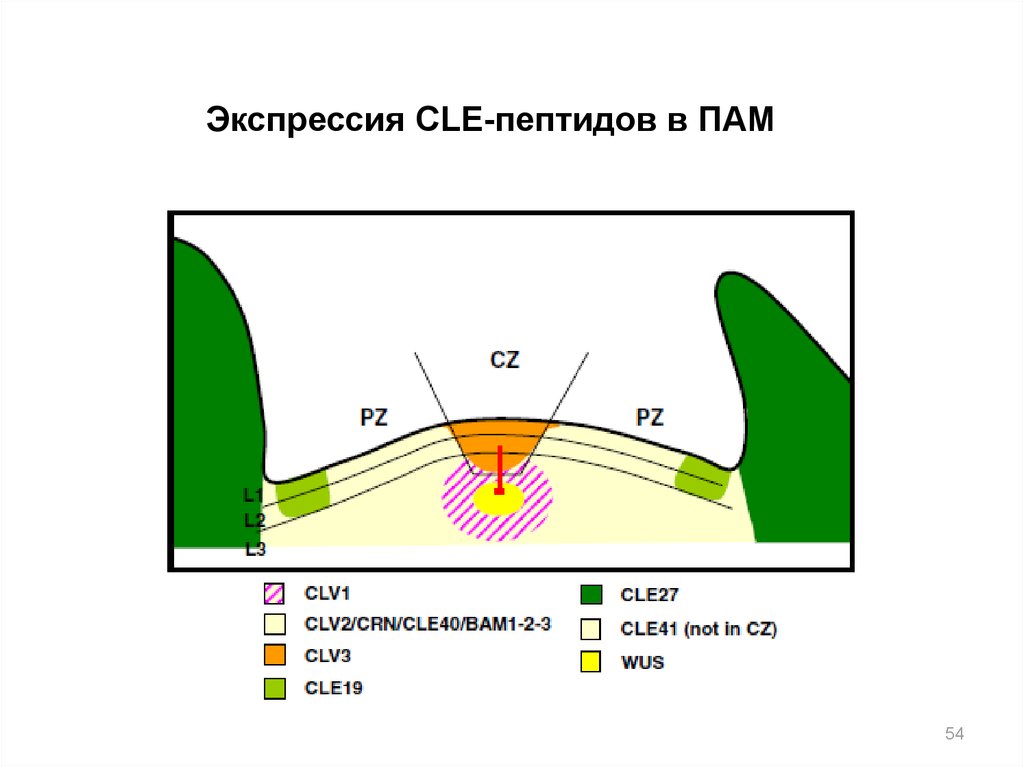
Экспрессия CLE-пептидов в ПАМ54
55.
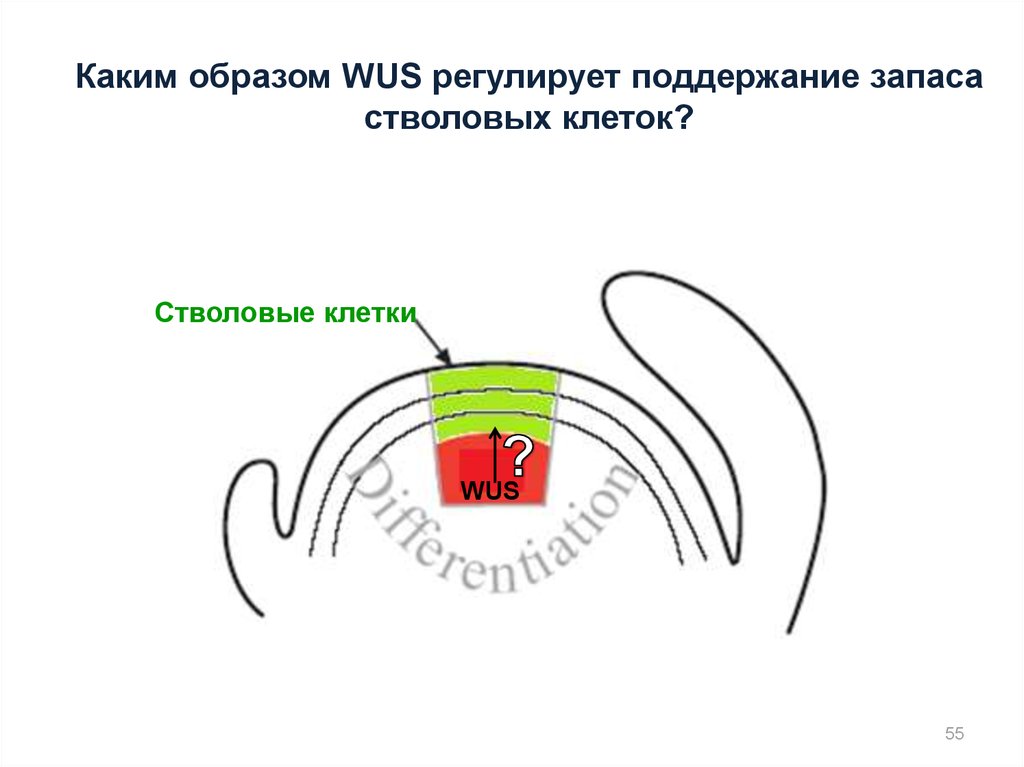
Каким образом WUS регулирует поддержание запасастволовых клеток?
Стволовые клетки
WUS
55
56.
Белок WUS мигрирует в вышележащие слои центральнойзоны меристемы, где расположены стволовые клетки
Yadav et al., 2011
56
57.
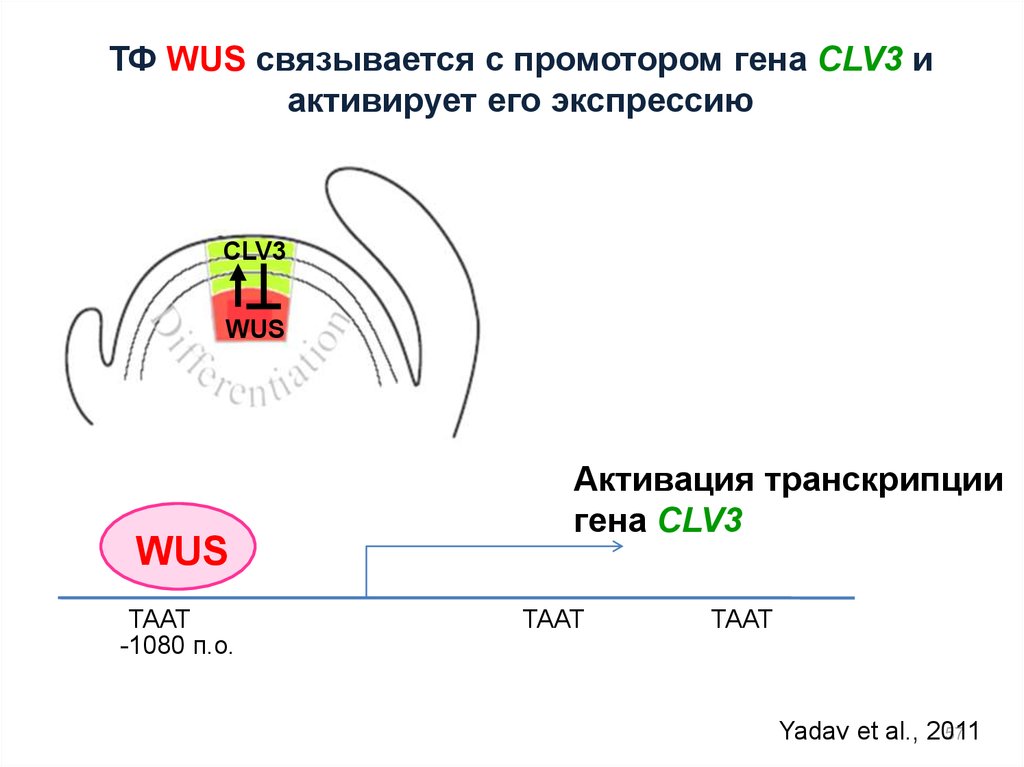
ТФ WUS связывается с промотором гена CLV3 иактивирует его экспрессию
CLV3
WUS
WUS
TAAT
-1080 п.о.
Активация транскрипции
гена CLV3
TAAT
TAAT
Yadav et al., 2011
57
58.
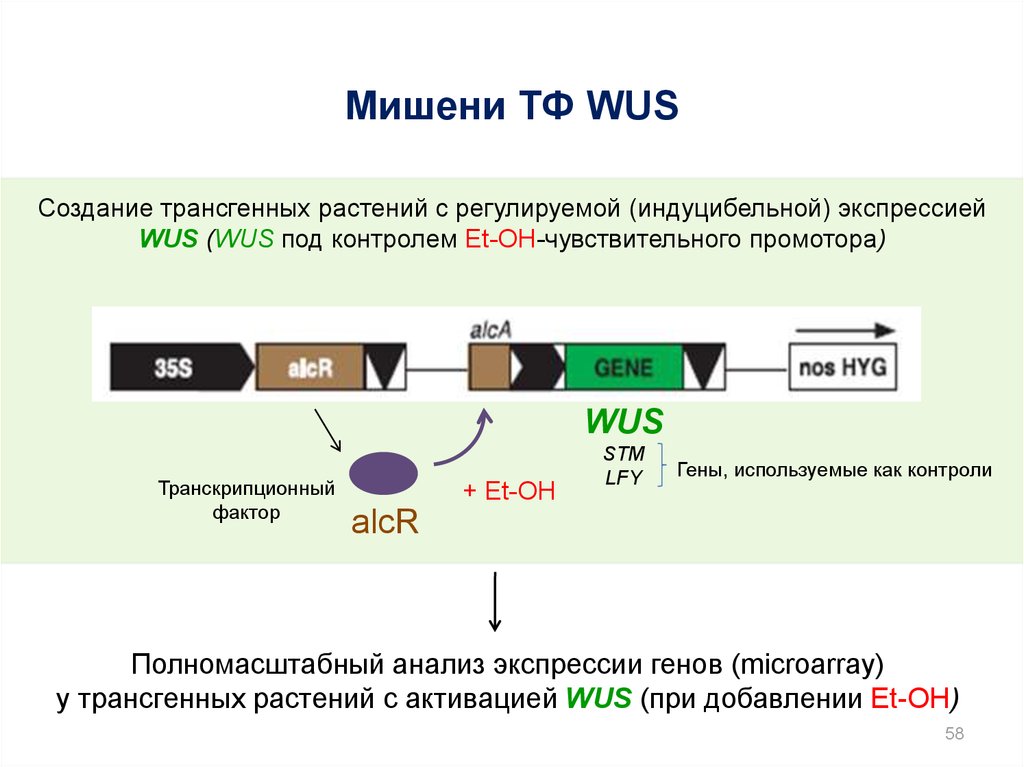
Мишени ТФ WUSСоздание трансгенных растений с регулируемой (индуцибельной) экспрессией
WUS (WUS под контролем Et-OH-чувствительного промотора)
WUS
Транскрипционный
фактор
+ Et-OH
STM
LFY
Гены, используемые как контроли
alcR
Полномасштабный анализ экспрессии генов (microarray)
у трансгенных растений с активацией WUS (при добавлении Et-OH)
58
59.
WUS негативно регулирует экспрессию генов многих ТФ,регулирующих дифференцировку клеток и развитие листа
Yadav et al., 2013
60.
Выявление мишеней ТФ WUS в транскриптомныхисследованиях
активация транскрипции
репрессия транскрипции
WUS
CLV1
ауксиновый
сигнальный
путь
цитокининовый
сигнальный
путь
60
Busch et al., 2010
61.
WUS негативно регулирует экспрессию геновARR А-типа (гены первичного ответа на цитокинин):
WUS
-1000 по
ARR7
61
62.
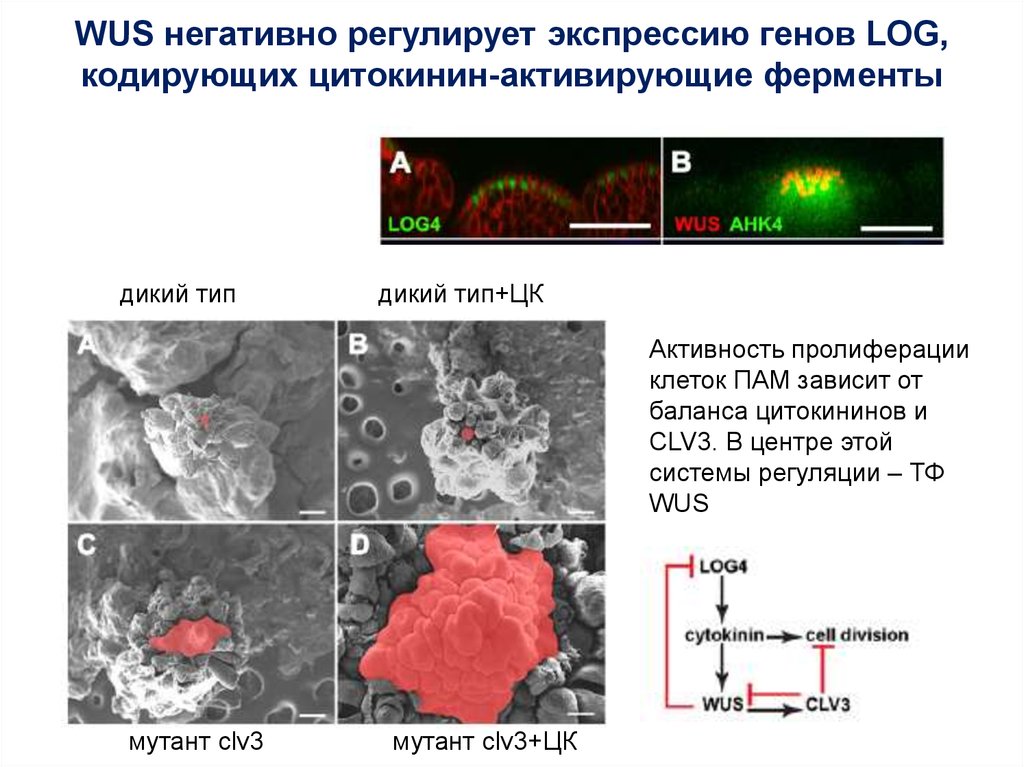
WUS негативно регулирует экспрессию генов LOG,кодирующих цитокинин-активирующие ферменты
дикий тип
дикий тип+ЦК
Активность пролиферации
клеток ПАМ зависит от
баланса цитокининов и
CLV3. В центре этой
системы регуляции – ТФ
WUS
мутант clv3
мутант clv3+ЦК
63.
Биосинтез и сигнальный путь цитокининов:роль WUS в их регуляции
WUS
WUS
63
64.
Взаимодействие транскрипционных факторов STM и WUSи цитокининов
STM
AtIPT
цитокинин
RR B-типа
RR А-типа
Цитокининовый
ответ
WUS
GA20оксидаза
GA2оксидаза
Гиббереллин
Ауксиновый ответ
64
65. Роль гормонов в ПАМ
6566.
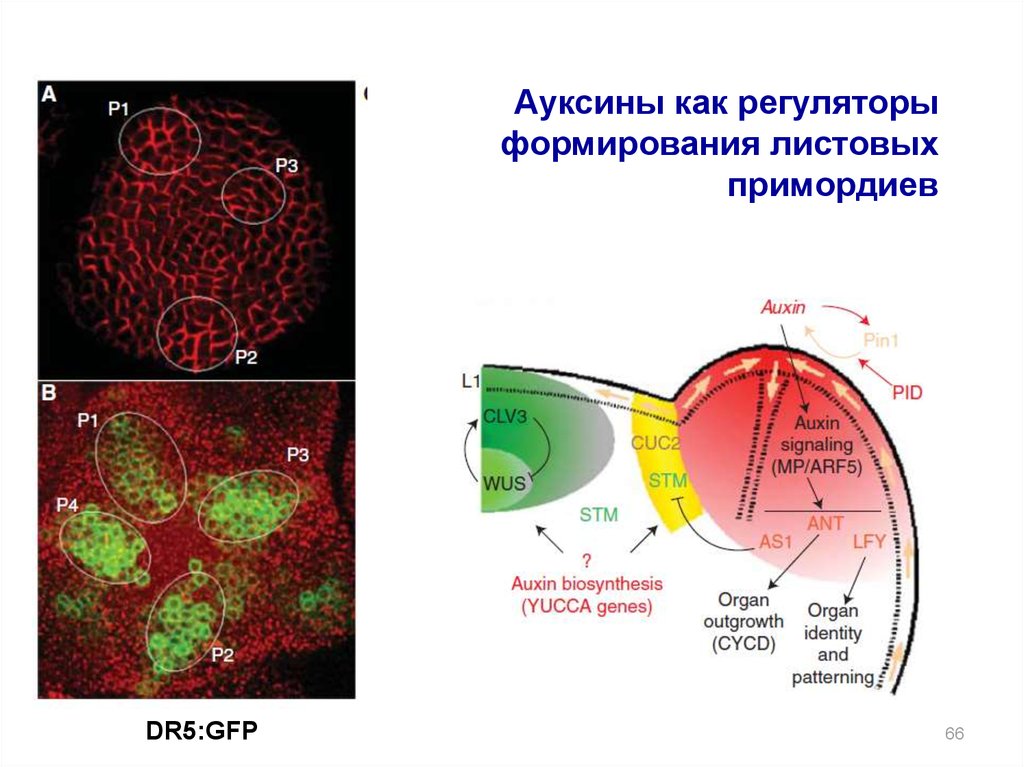
Ауксины как регуляторыформирования листовых
примордиев
DR5:GFP
66
67.
Модель взаимодействия ТФ и гормонов в развитии растенийРаспределение
гормонов в
растительном
организме
Локальная
экспрессия
генов,
кодирующих ТФ
Локальное
изменение
концентрации
гормона (или
чувствительности
клеток к действию
гормона) в области
экспрессии ТФ
Изменение
параметров роста
растения
68.
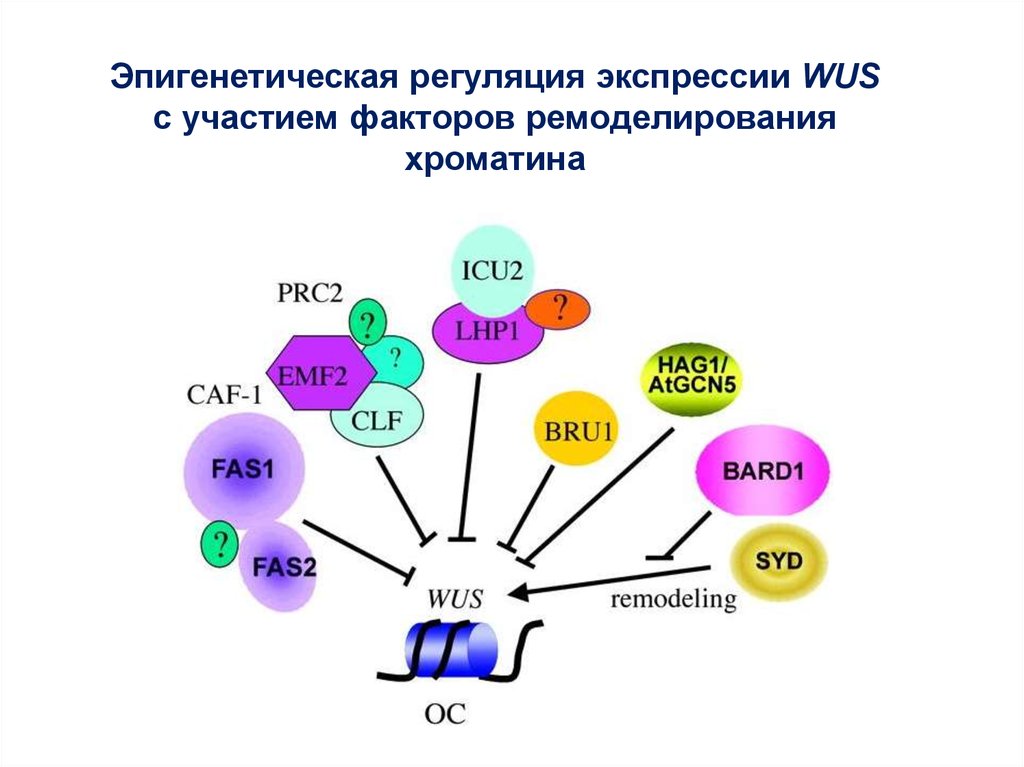
Эпигенетическая регуляция экспрессии WUSс участием факторов ремоделирования
хроматина
69.
Нарушение организации ПАМ у мутантов по генам FASCIATAwt
fas1-1
wt
fas2-1
FAS1 и FAS2 кодируют субъединицы комплекса CAF-1 - (CHROMATIN
ASSEMBLY FACTOR-1) – шапероны гистонов H3/H4
Kaya et al., 2001
70.
При выжигании зон экспрессииWUS или CLV3 они
восстанавливаются в
прилегающих клетках
существуют механизмы, определяющие экспрессию
этих генов в определенной зоне
before
WUS
CLV3
just after
2 days after
71.
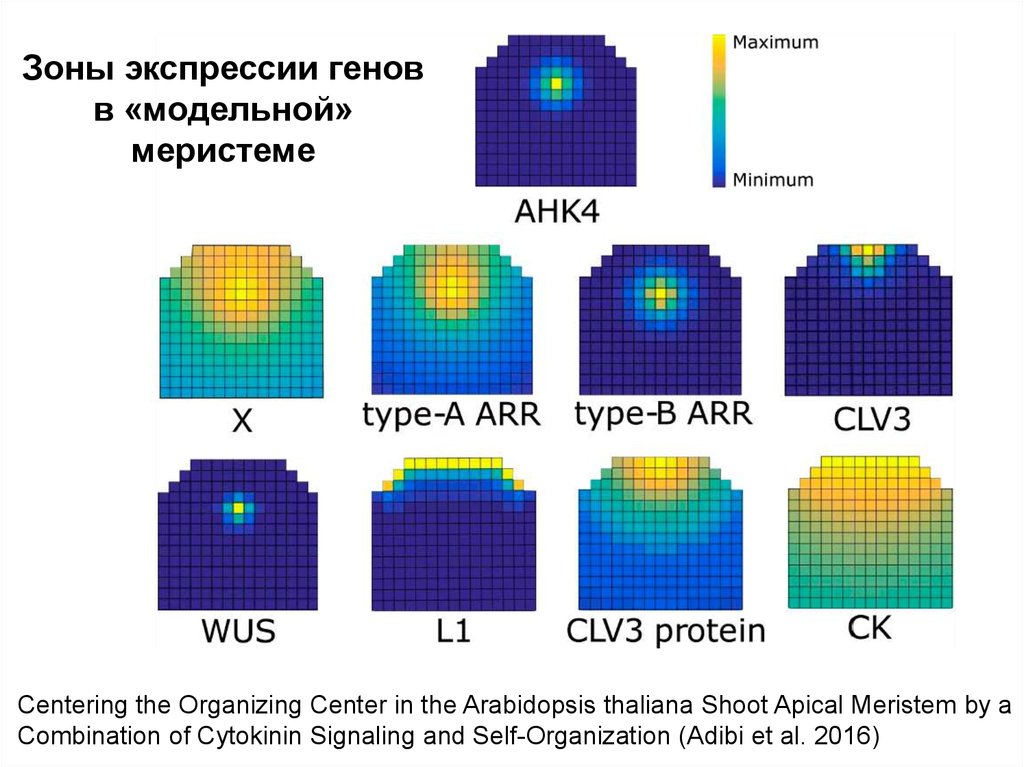
Зоны экспрессии геновв «модельной»
меристеме
Centering the Organizing Center in the Arabidopsis thaliana Shoot Apical Meristem by a
Combination of Cytokinin Signaling and Self-Organization (Adibi et al. 2016)
72.
Регуляции функции меристемы с участиемсигналов от дифференцированных клеток
ham (hairy meristem) – мутанты, характеризующиеся остановкой развития ПАМ и
дифференцировкой меристематических клеток (клетки слоя L1 в клетки
эпидермиса, нижележащие слои клеток – в клетки коры и проводящие ткани).
дикий тип
ham
(hairy meristem)
Дифференцировка
клеток меристемы
HAM
HAM - транскрипционный
фактор семейства GRAS
Stuurman et al., 2002
72
73.
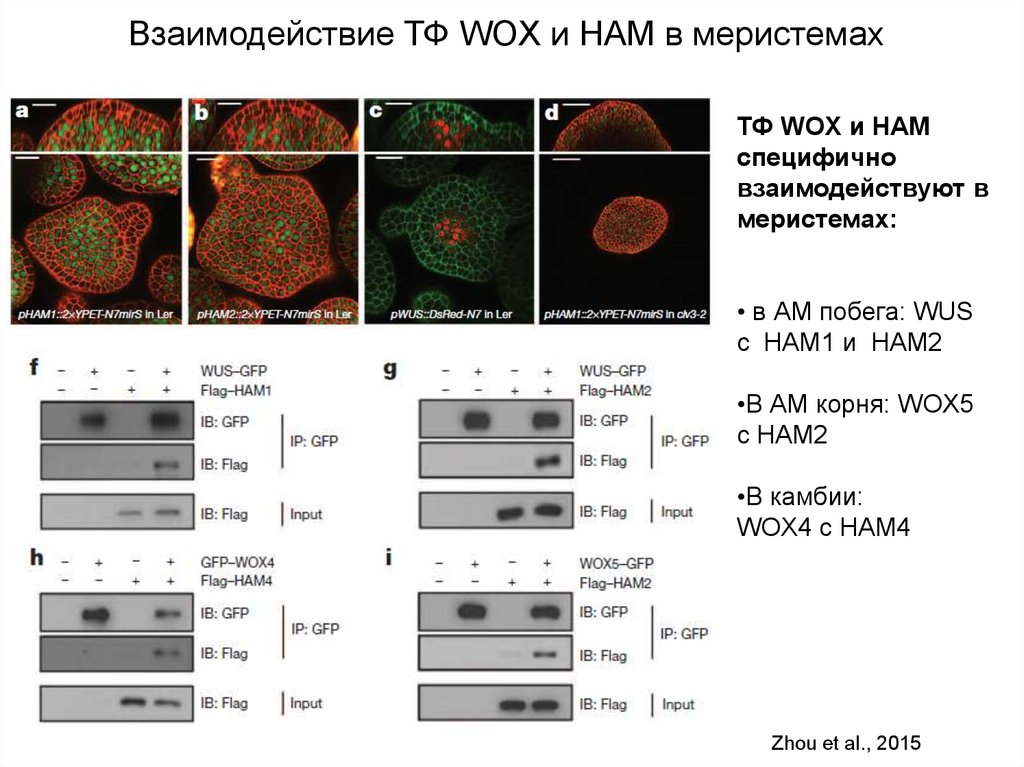
Взаимодействие ТФ WOX и HAM в меристемахТФ WOX и НАМ
специфично
взаимодействуют в
меристемах:
• в АМ побега: WUS
с HAM1 и HAM2
•В АМ корня: WOX5
с HAM2
•В камбии:
WOX4 с HAM4
Zhou et al., 2015
74.
ФиллотаксисЗолотое сечение (1,61803…)
Золотой угол (137,5°…) – «золотое сечение» окружности
75.
Модели филлотаксиса«Ауксиновая» модель
«Механическая» модель
MAP4 (Microtubule Associated Protein4):GFP
76.
Взаимодействие факторов, регулирующих активность ПАМGiacomo et al., 2013
77.
Регуляция клеточного цикла растений77