







































 Медицина
МедицинаПохожие презентации:



")





")
Защитные функции антител
1.
Защитные функции антителЛекция 9
2.
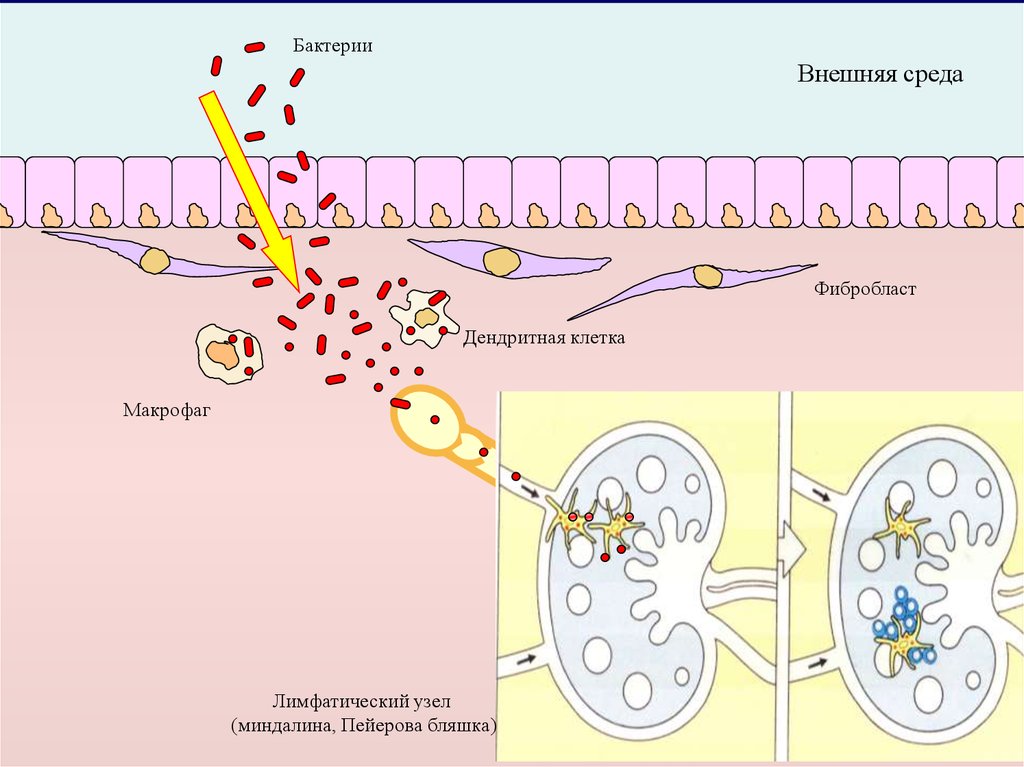
БактерииВнешняя среда
Фибробласт
Дендритная клетка
Макрофаг
Лимфатический узел
(миндалина, Пейерова бляшка)
3. В-клетка, у которой сработали рецепторы, секретирует огромное количество молекул, подобных рецепторам
Black box4.
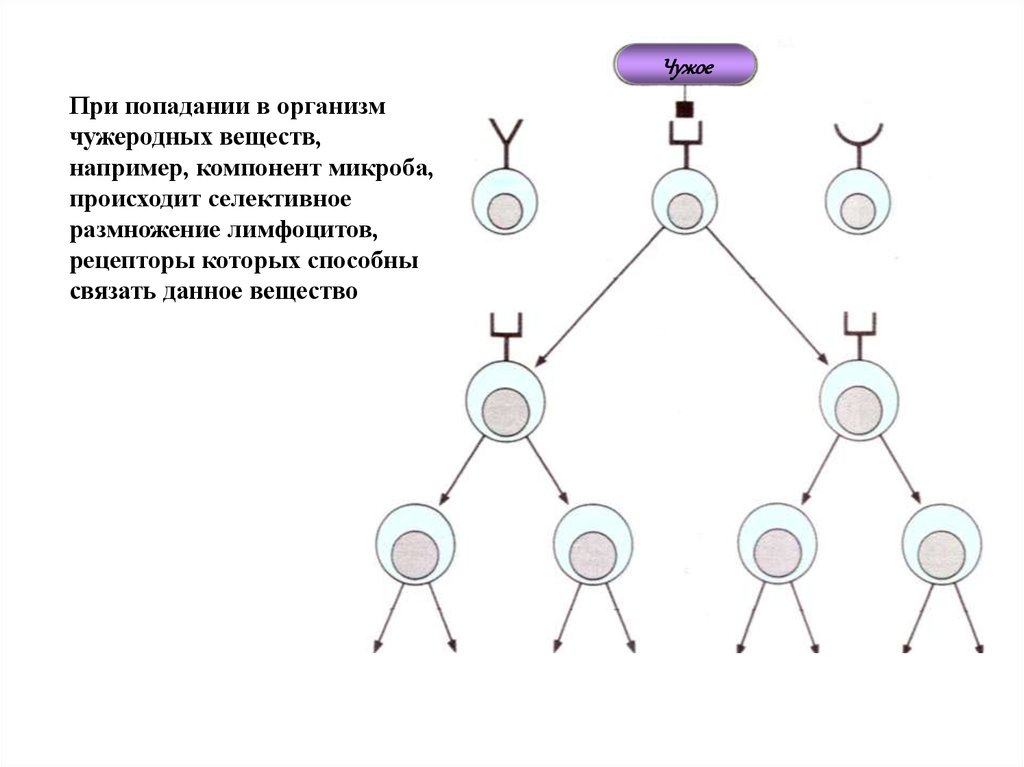
ЧужоеПри попадании в организм
чужеродных веществ,
например, компонент микроба,
происходит селективное
размножение лимфоцитов,
рецепторы которых способны
связать данное вещество
5.
В-лимфоциты, «схватившие» антиген ,не только размножаются (~ 1 деление в сутки),
но и сильно изменяются – дифференцируются.
Значительные изменения клетки, сопровождающиеся изменением активности
многих генов, изменением структуры клетки и ее функциональных свойств,
называется клеточной дифференцировкой.
В-лимфоциты, «схватившие» антиген, дифференцируются в плазматические
клетки, секретирующие триллионы молекул антител, идентичных их Igрецепторам. Эта дифференцировка – терминальная, то есть плазматическая
клетка больше ни во что не превращается, живет несколько дней и гибнет.
6.
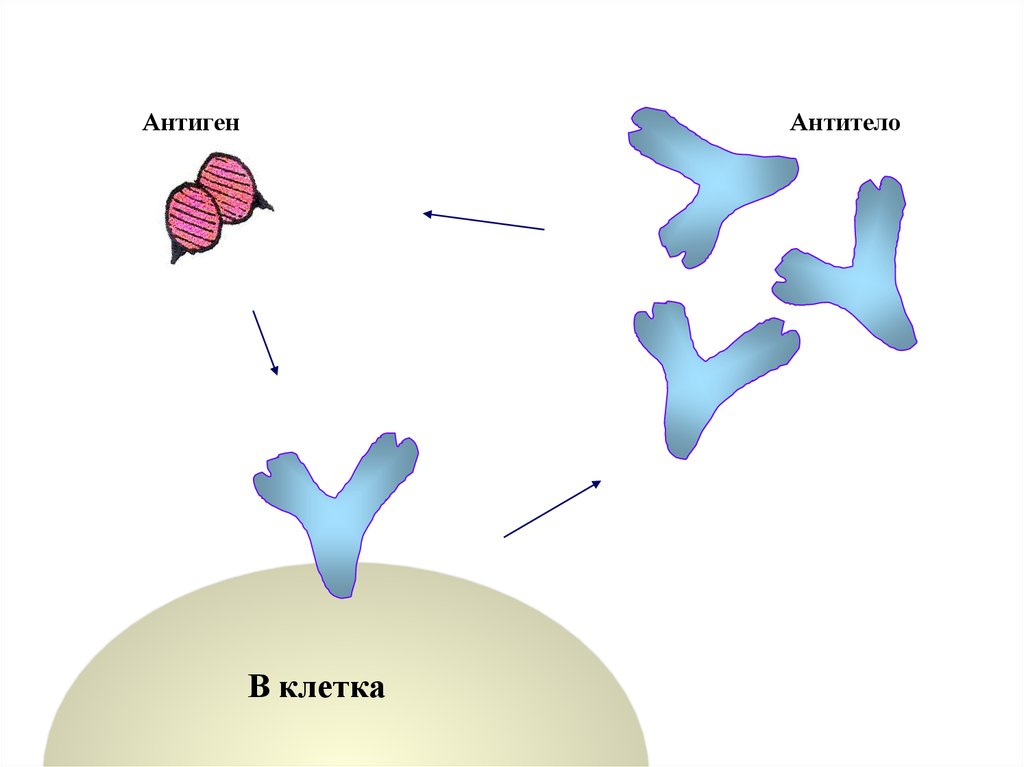
В клеткаПлазматическая
клетка
7.
АнтигенАнтитело
В клетка
8.
Антитела находят соответствующие антигеныи связываются с ними.
Растворимые антигены переводятся в нерастворимые
комплексы [антиген + антитело].
9.
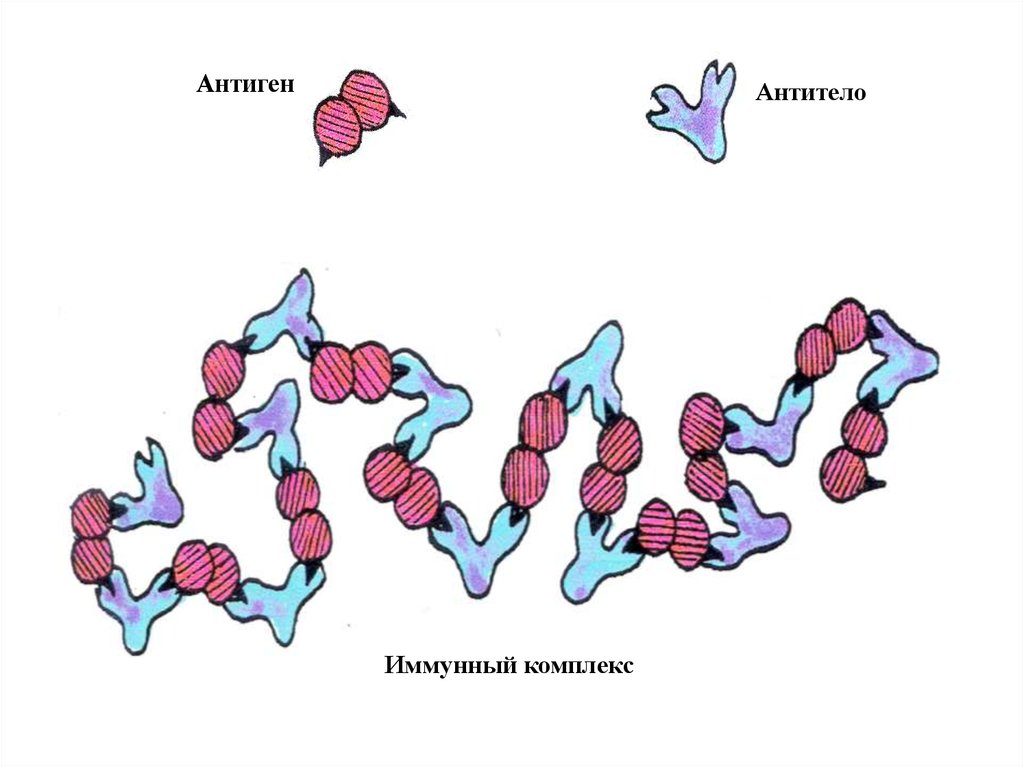
АнтигенАнтитело
Иммунный комплекс
10.
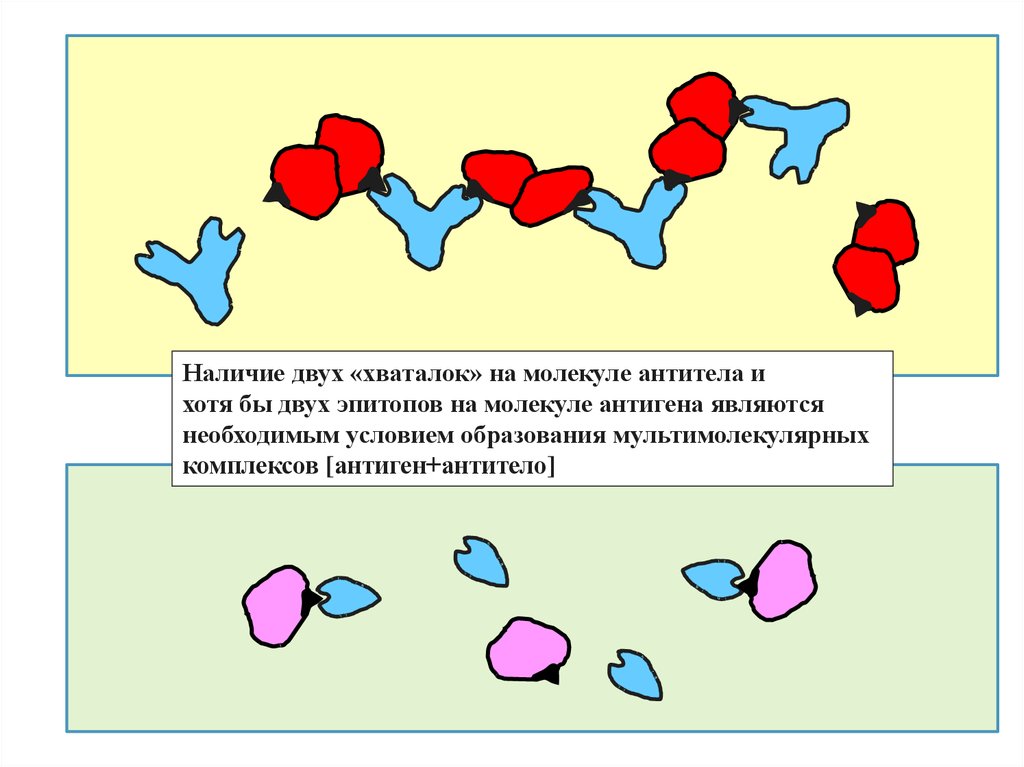
Наличие двух «хваталок» на молекуле антитела ихотя бы двух эпитопов на молекуле антигена являются
необходимым условием образования мультимолекулярных
комплексов [антиген+антитело]
11.
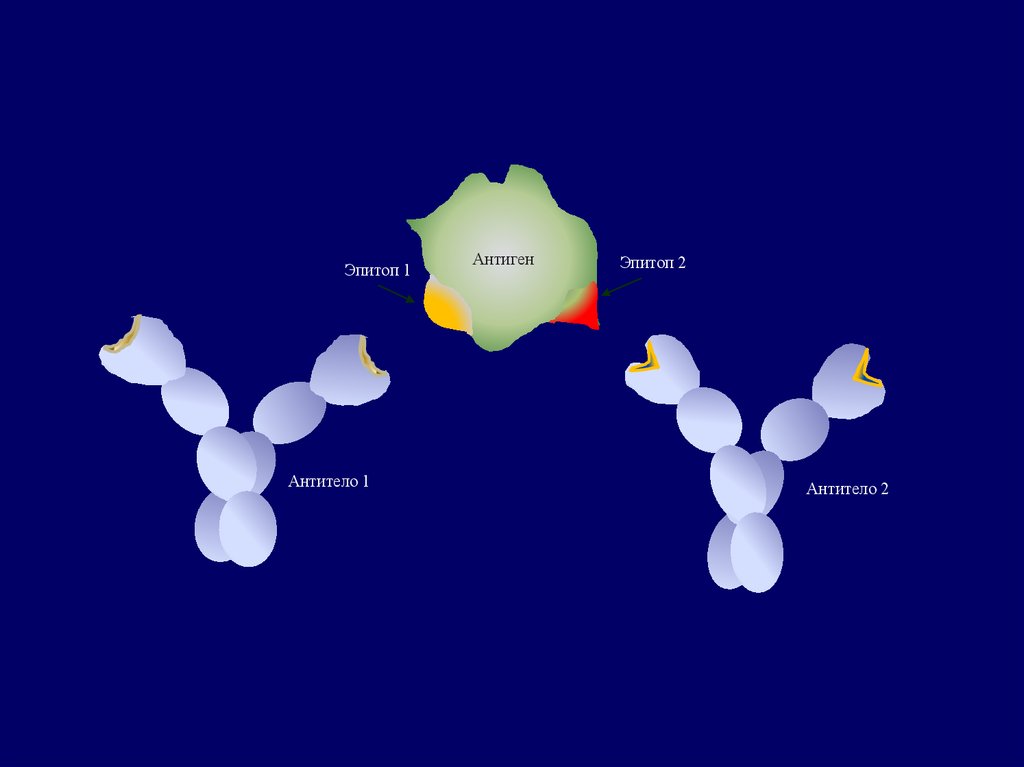
Эпитоп 1Антитело 1
Антиген
Эпитоп 2
Антитело 2
12.
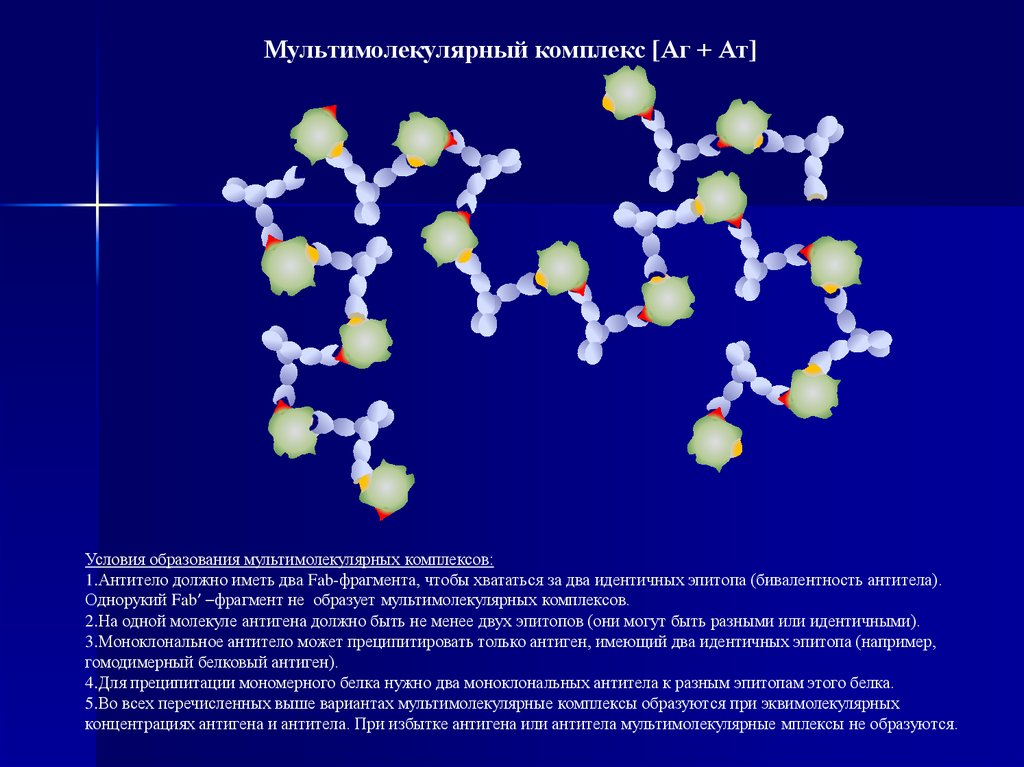
Мультимолекулярный комплекс [Аг + Ат]Условия образования мультимолекулярных комплексов:
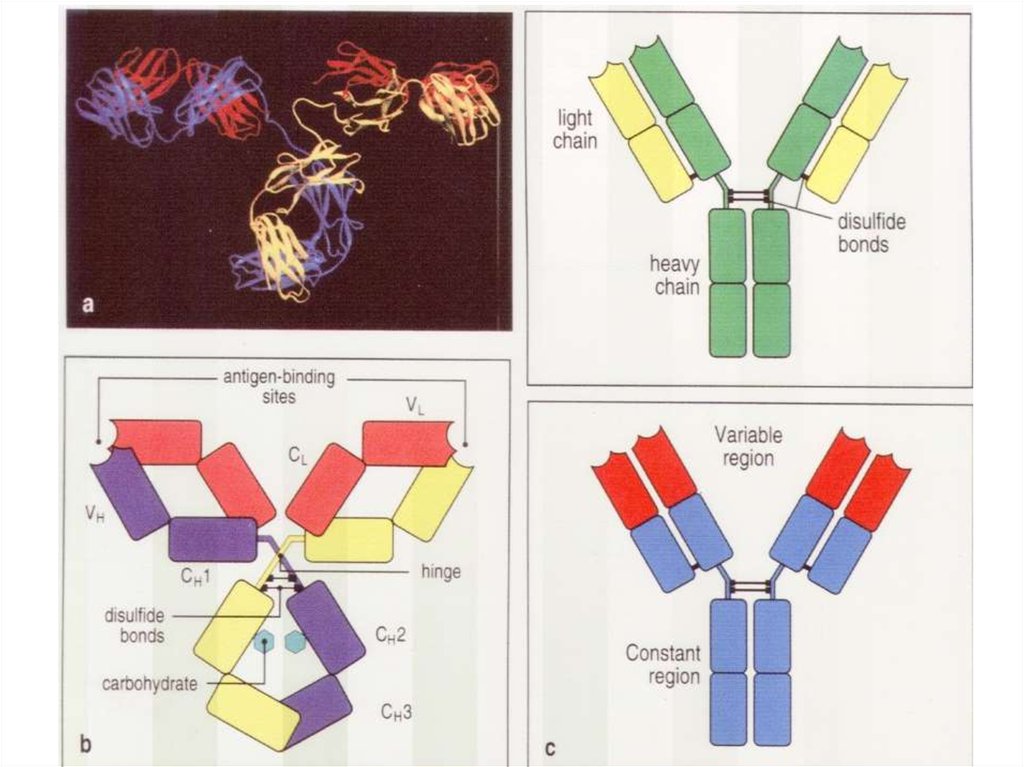
1.Антитело должно иметь два Fab-фрагмента, чтобы хвататься за два идентичных эпитопа (бивалентность антитела).
Однорукий Fab’ –фрагмент не образует мультимолекулярных комплексов.
2.На одной молекуле антигена должно быть не менее двух эпитопов (они могут быть разными или идентичными).
3.Моноклональное антитело может преципитировать только антиген, имеющий два идентичных эпитопа (например,
гомодимерный белковый антиген).
4.Для преципитации мономерного белка нужно два моноклональных антитела к разным эпитопам этого белка.
5.Во всех перечисленных выше вариантах мультимолекулярные комплексы образуются при эквимолекулярных
концентрациях антигена и антитела. При избытке антигена или антитела мультимолекулярные мплексы не образуются.
13.
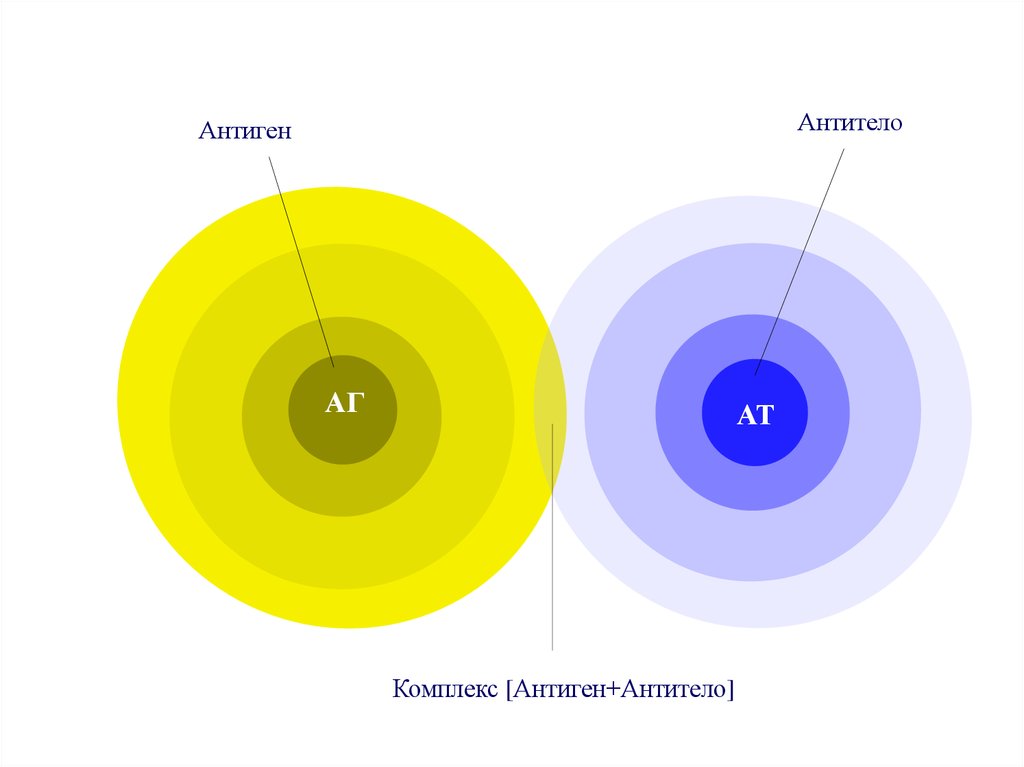
АнтителоАнтиген
АГ
АТ
Комплекс [Антиген+Антитело]
14.
15.
16.
17.
18.
Как и от чего можно защититься с помощью антител?Римский додекаэдр – не известно, зачем он был нужен
19. ЗАЩИТНЫЕ ФУНКЦИИ АНТИТЕЛ
Нейтрализация токсинаУдаление токсина в виде иммунных комплексов
Нейтрализация вируса
Препятствие адгезии бактерий
Активация комплемента на поверхности вирусов,
бактерий и других существ
Опсонизация бактерий, вирусов и других патогенов
(за счет антител и комплемента)
Нацеливание киллеров (через FcR)
Защита границ за пределами организма (IgA)
20.
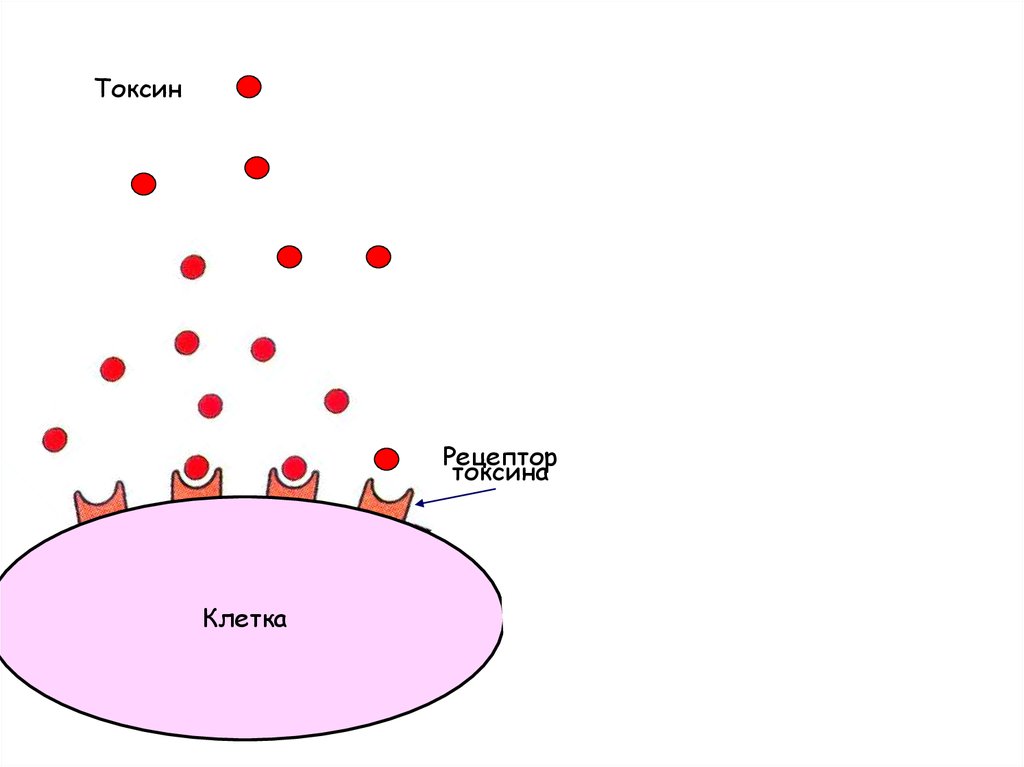
АнтителоТоксин
Рецептор
токсина
Клетка
Клетка
21.
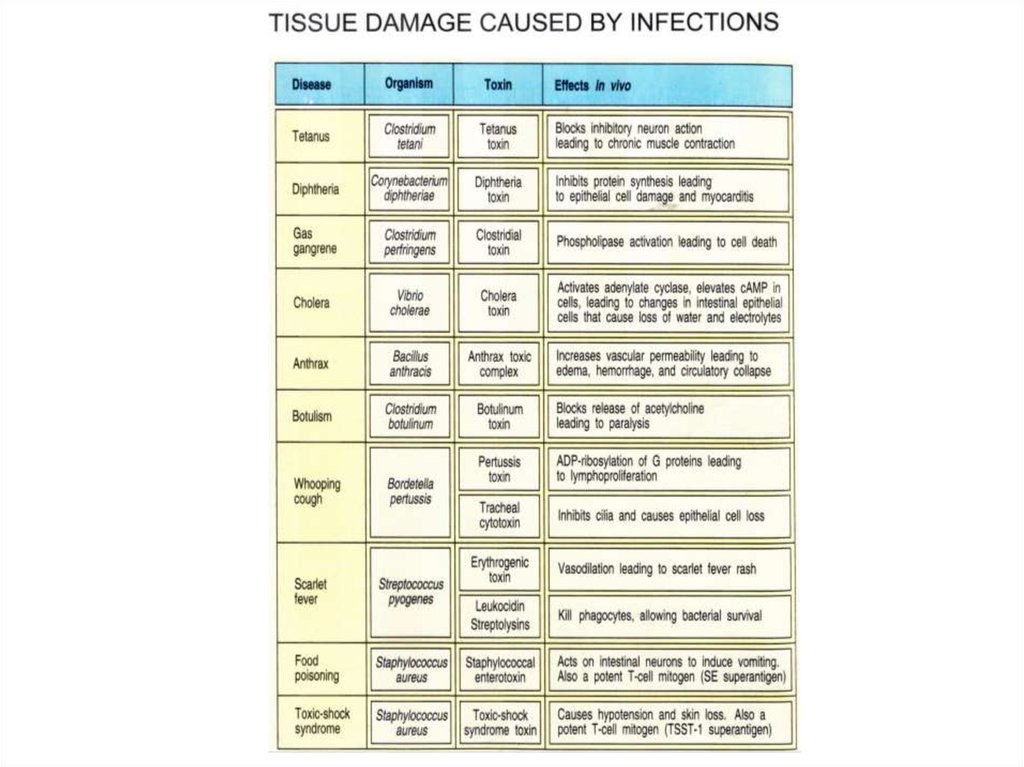
Зачастую вред наносит не бактерия, а еетоксины.
В таких случаях, если обезвредить
токсины, то присутствие бактерий не
вызовет болезни (примеры – холера,
дифтерия, столбняк и др.)
Старая истина –
мал клоп, да вонюч
22.
23.
Токсичность ядов бактерийТоксин
Дифтерийный
токсин
Ботулотоксин
100 нг/кг
1 грамм =
смертельных доз
100 000 человек
1 нг/кг
10 000 000 человек
LD50
Перфрингенс бета- 400 нг/кг
токсин
Перфрингенс
100 нг/кг
эпсилон-токсин
Тетанус-токсин
1 нг/кг
25 000 человек
100 000 человек
10 000 000 человек
Сравните:
•Яд змеи тайпан (самая ядовитая змея) LD50 = 10 мкг/кг, 1 грамма достаточно убить 1000 чел
•Яд кобры LD50 ~ 1мг/кг, 1 грамма достаточно убить 10 чел
24.
Нейтрализация опасного зверя25.
Антитела препятствуютадгезии бактерий
26.
Антитела нейтрализуют вирусы27.
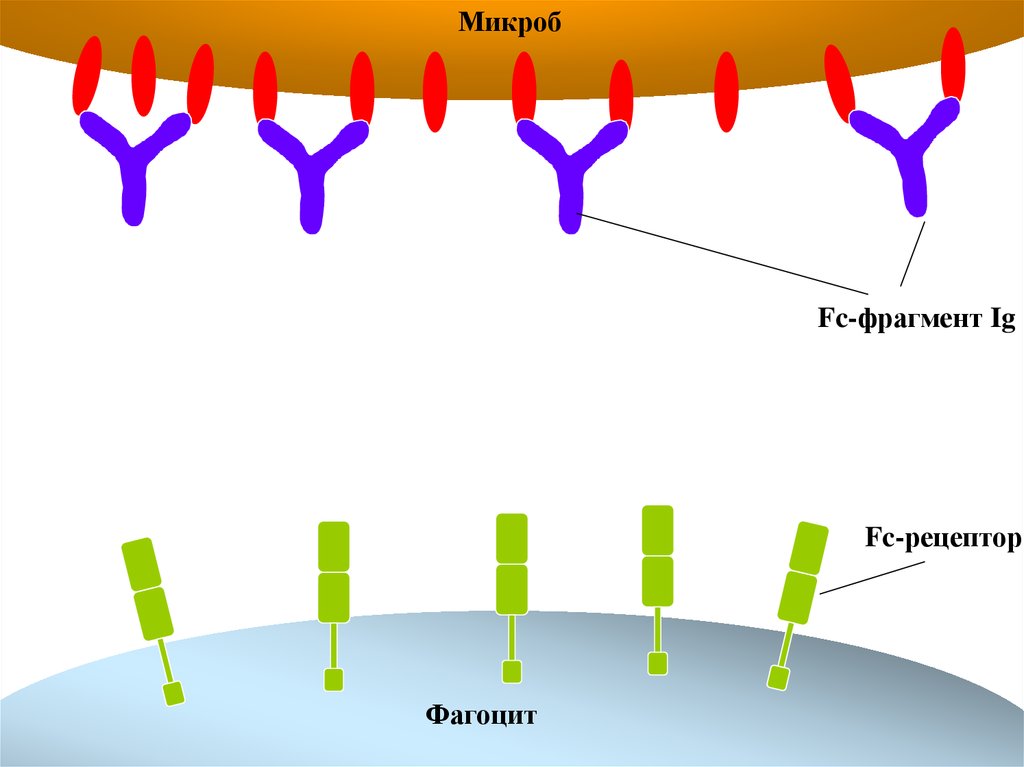
Антитела опсонизируют поверхность микробов28.
МикробFc-фрагмент Ig
Fc-рецептор
Фагоцит
29.
Fc-рецепторы фагоцитов облегчают фагоцитозчастиц, покрытых антителами
30.
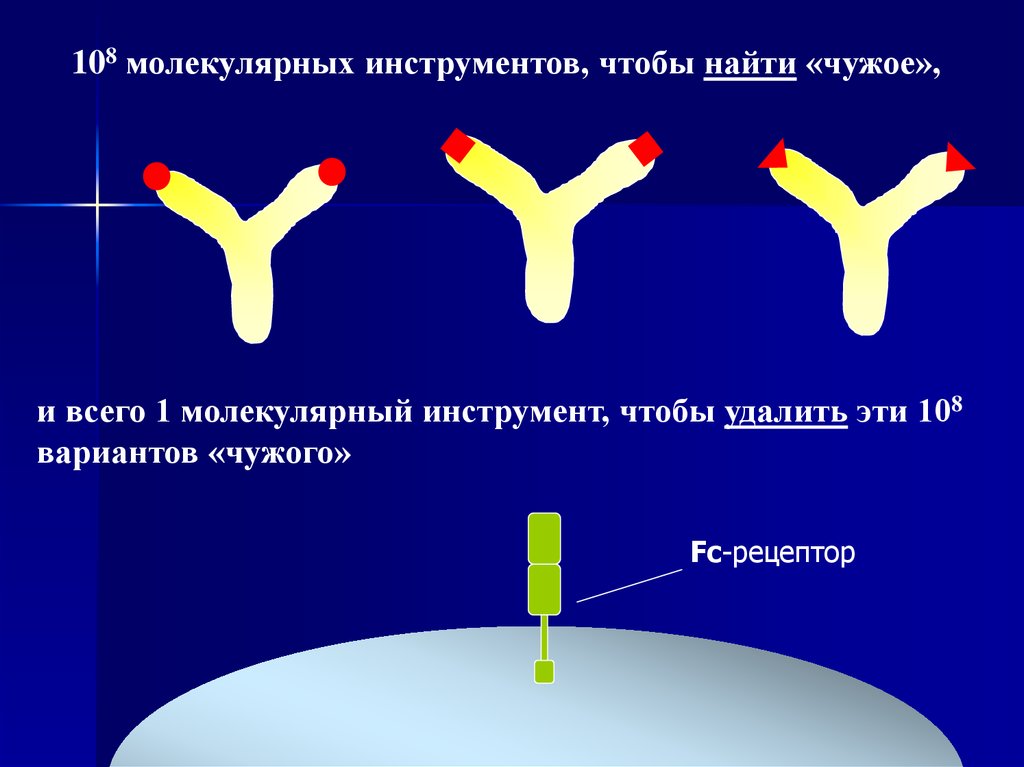
31.
108 молекулярных инструментов, чтобы найти «чужое»,и всего 1 молекулярный инструмент, чтобы удалить эти 108
вариантов «чужого»
Fc-рецептор
32.
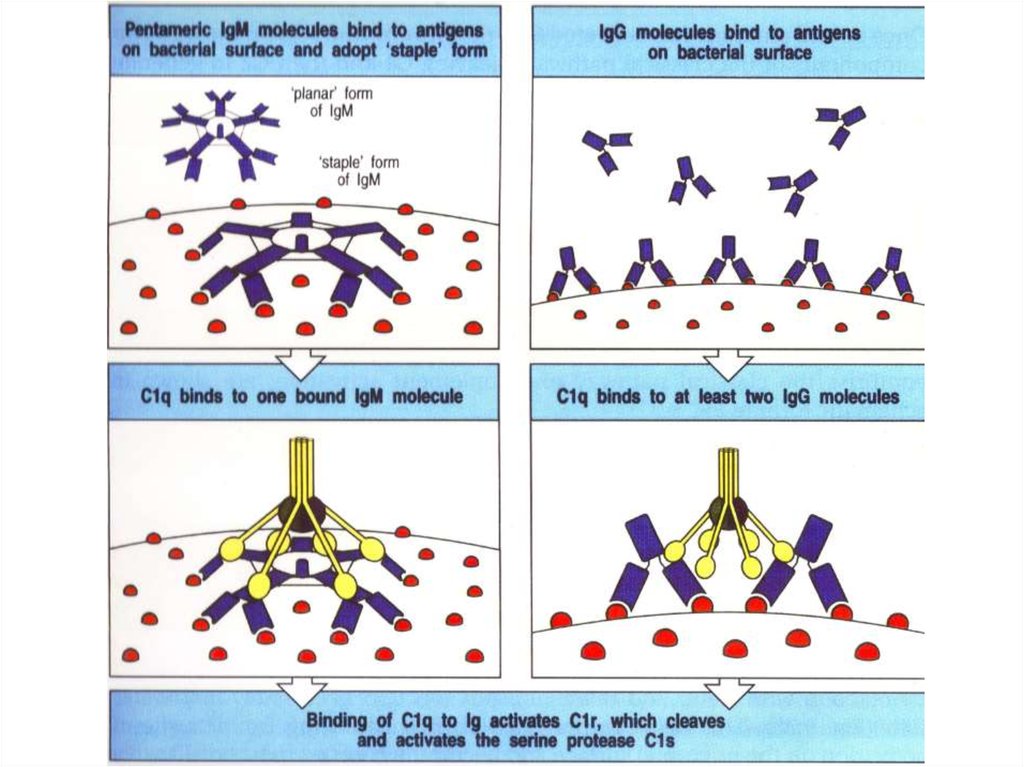
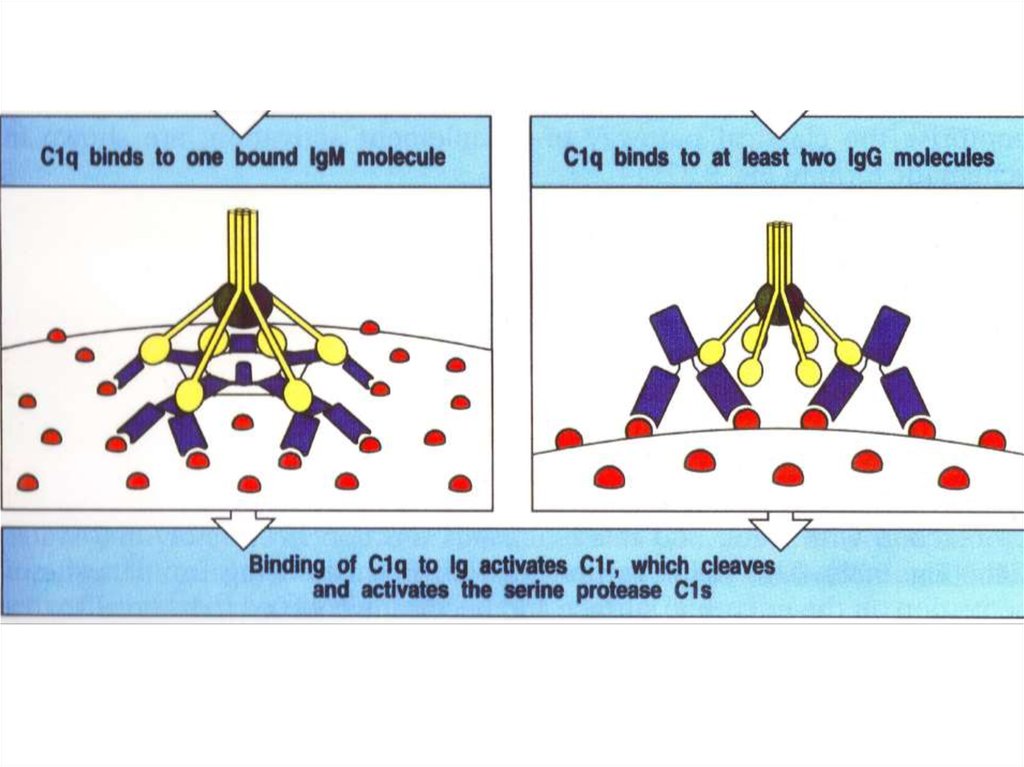
Комплексы [антиген + антитело] активируют комплемент33.
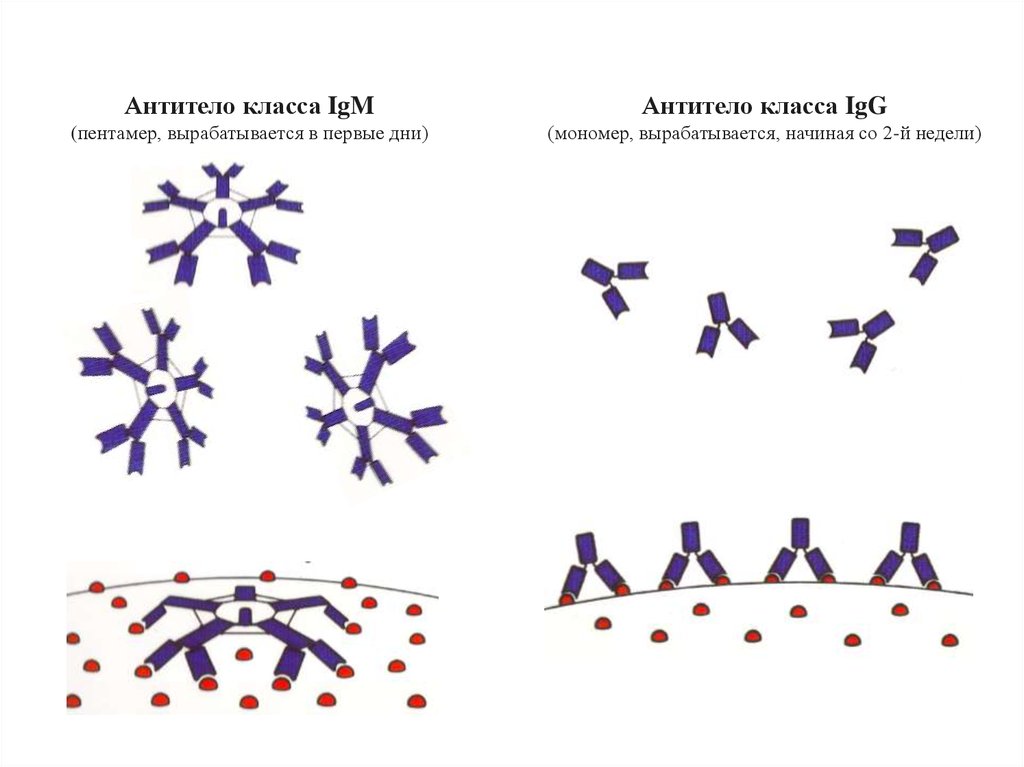
Антитело класса IgMАнтитело класса IgG
(пентамер, вырабатывается в первые дни)
(мономер, вырабатывается, начиная со 2-й недели)
34.
35.
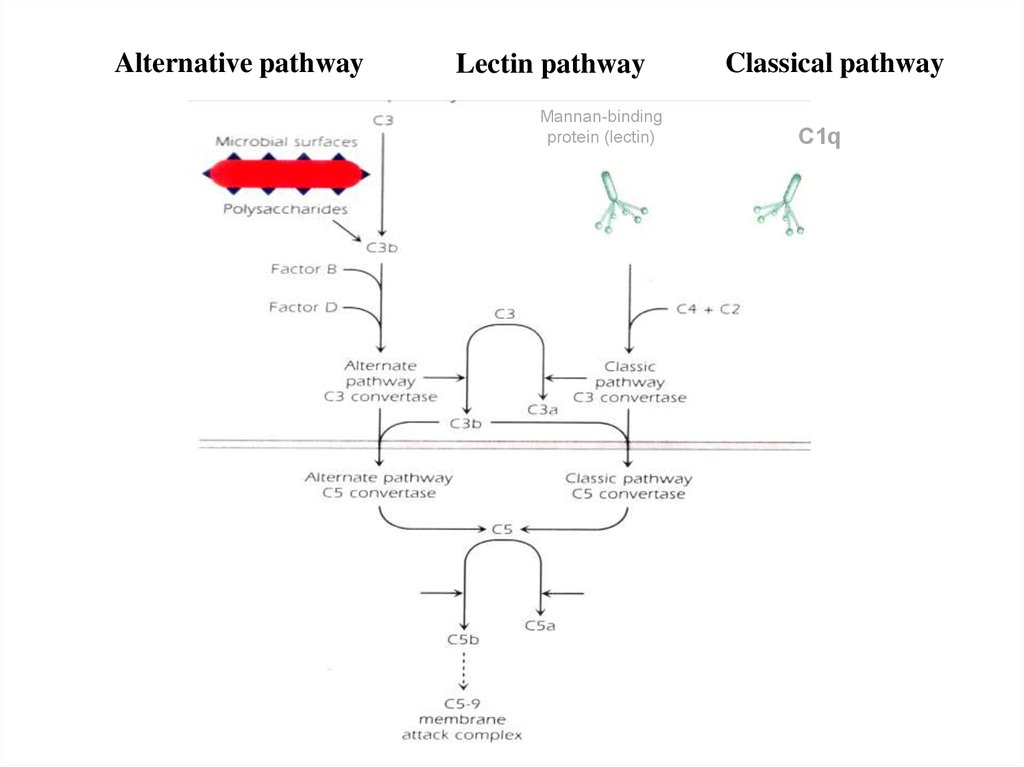
Alternativepathway
Lectin
pathway
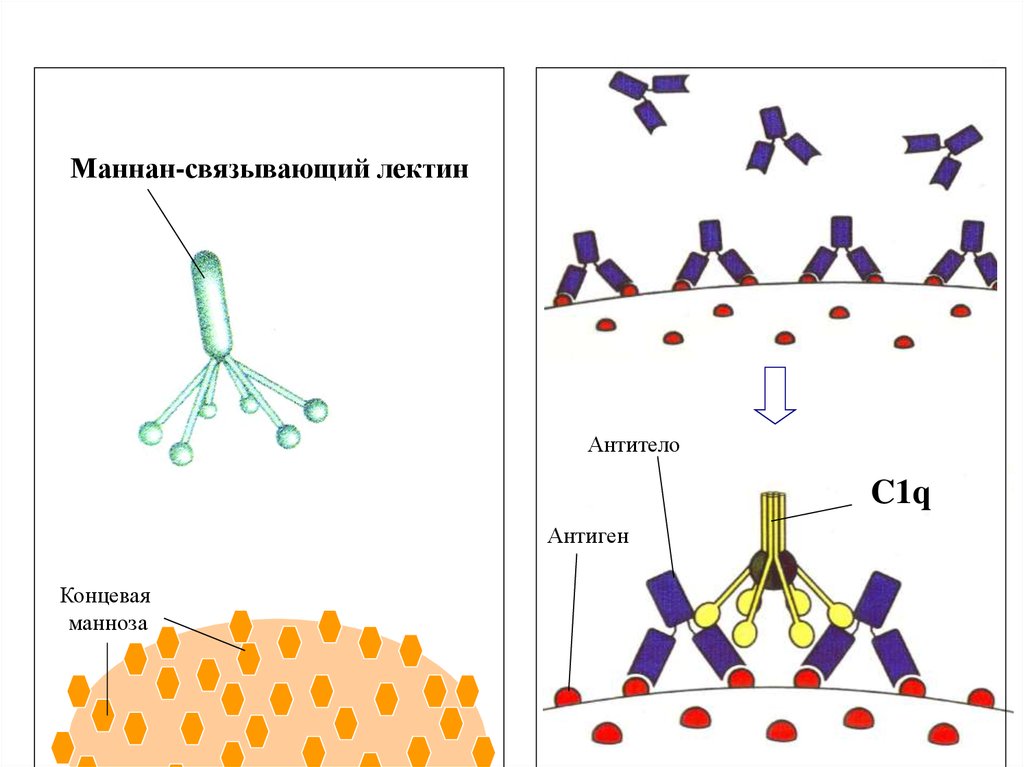
Маннан-связывающий лектин
C4
Концевая
манноза
36.
Маннан-связывающий лектинАнтитело
C1q
Антиген
Концевая
манноза
37.
38.
39.
40.
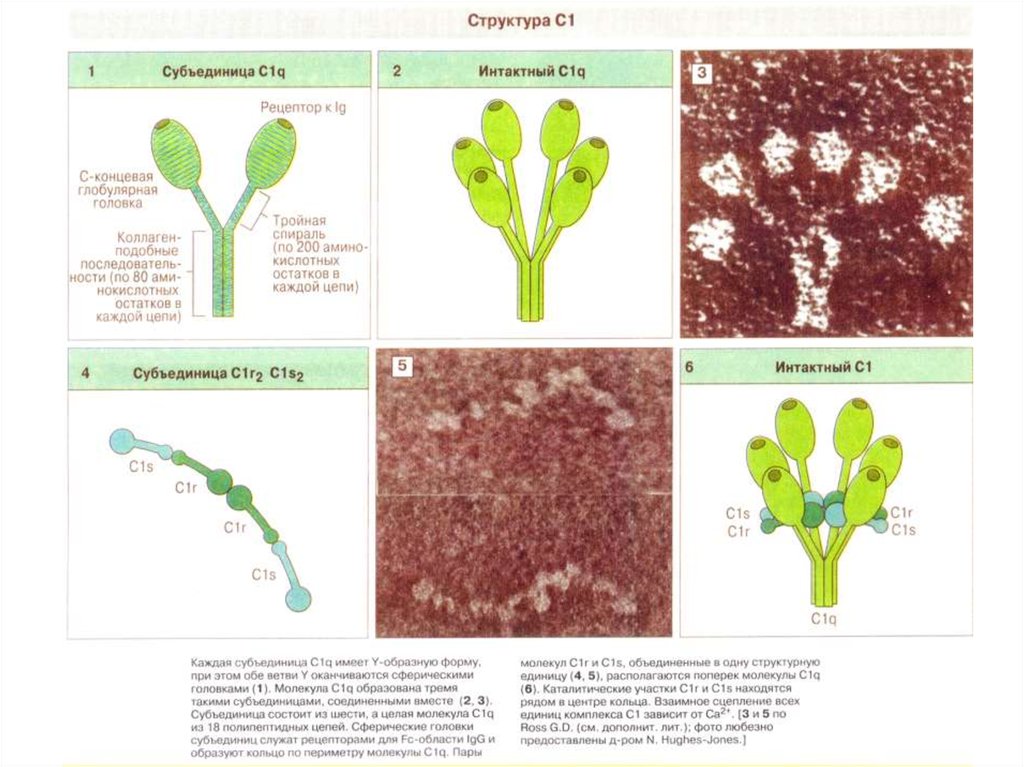
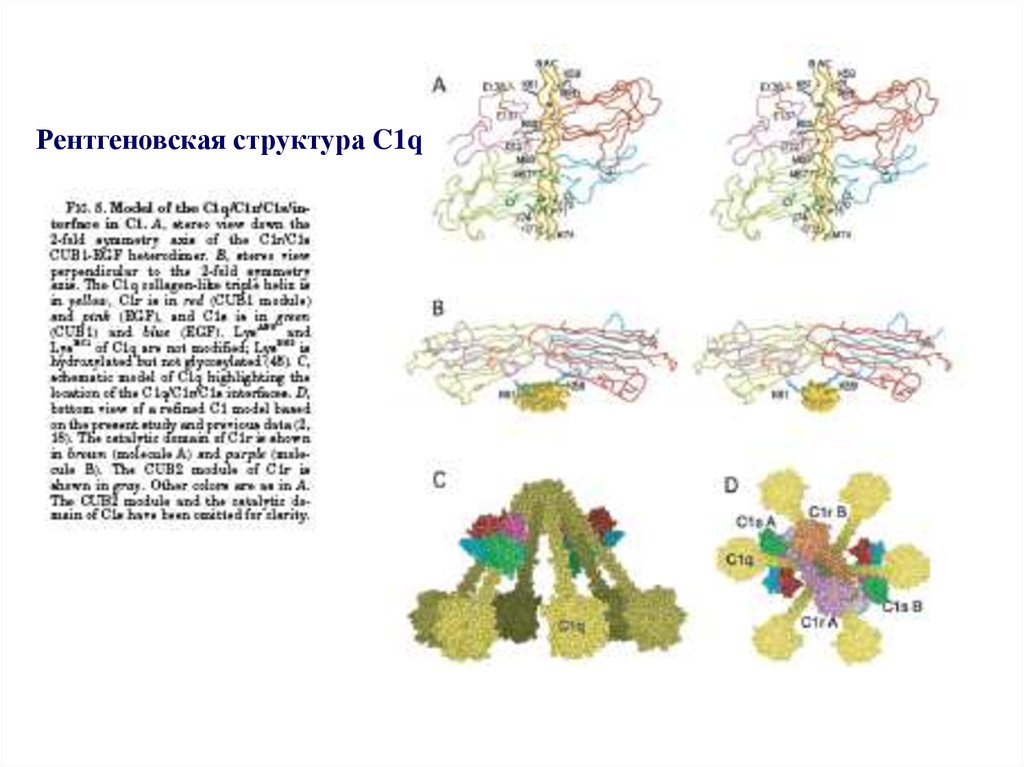
Рентгеновская структура C1q41.
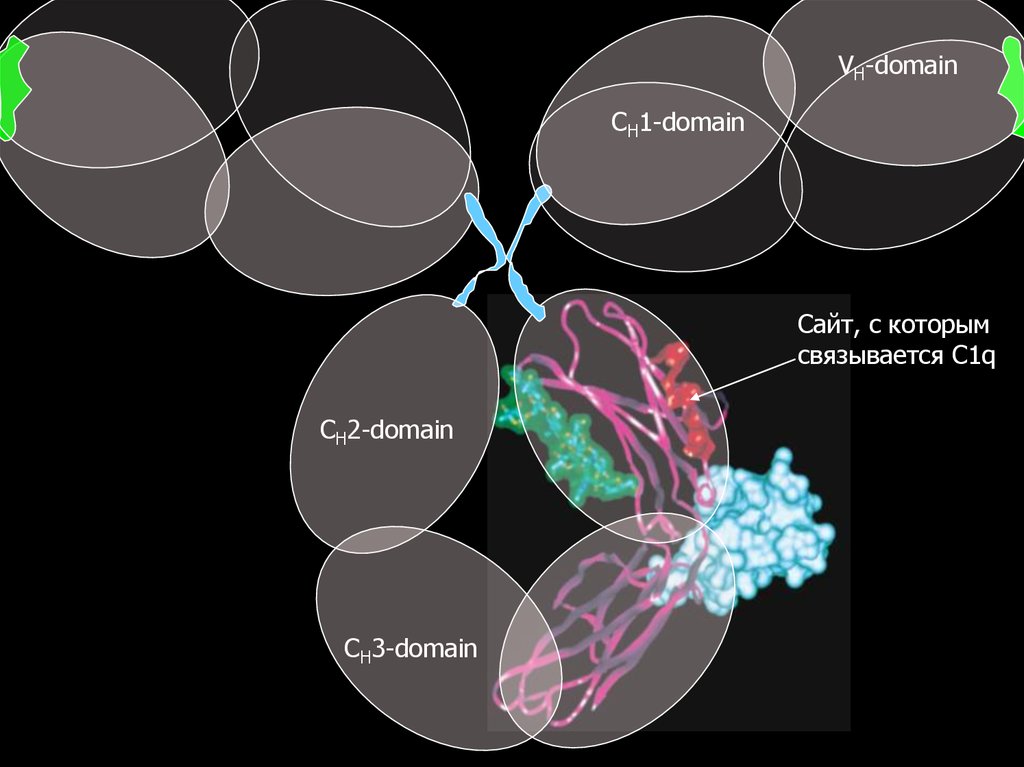
VH-domainCH1-domain
Сайт, с которым
связывается C1q
CH2-domain
CH3-domain
42.
Посадку C1q на две молекулы Ig можно реконструироватьс учетом рентгеновской структуры молекул
(см. следующие два слайда)
43.
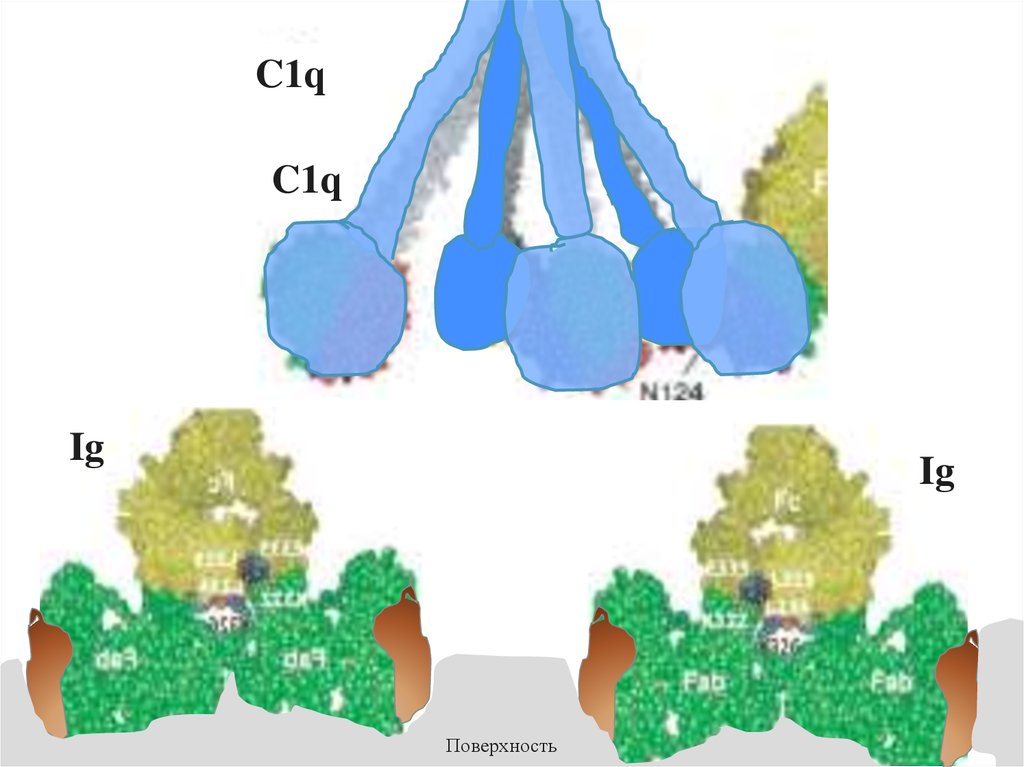
Модель взаимодействия C1q c Fc-фрагментом антитела (IgG)C1q
Ig
44.
C1qC1q
Ig
Ig
Поверхность
45.
Движение коллагеноподобных стержнеймолекулы маннан-связывающего лектина
перемещает субъединицы MASP, что приводит к
их суперпозиции и активации
Figure 6 Structural Basis of Complement Activation in the Lectin Pathway (A) Superimposition of the structures of the MBL/MASP-1 complex
with the human CUB1-EGF-CUB2 (PDB: <ce:inter-ref href="pdb:3DEM" type="simple"> 3DEM</ce:inter-ref> ) and extension of...
Alexandre R. Gingras , Umakhanth Venkatraman Girija , Anthony H. Keeble , Roshni Panchal , Daniel A. Mitchell , Pet...
Structure Volume 19, Issue 11 2011 1635 - 1643
Structural Basis of Mannan-Binding Lectin Recognition by Its Associated Serine Protease MASP-1: Implications for Complement
Activation
http://dx.doi.org/10.1016/j.str.2011.08.014
46.
C2 C2a +C2bC4 C4a + C4b
C3 C3a +C3b
C1r, C1s
C5 C5a + C5b
C1q
C5
C9
C2aC4b
(конвертаза С3)
C2aC4bC3b
(конвертаза С5)
C6 C5b
C7
C9C9
C9 C9C9
C8
C9
C9
Пора
47.
Alternative pathwayLectin pathway
Mannan-binding
protein (lectin)
Classical pathway
С1q
48.
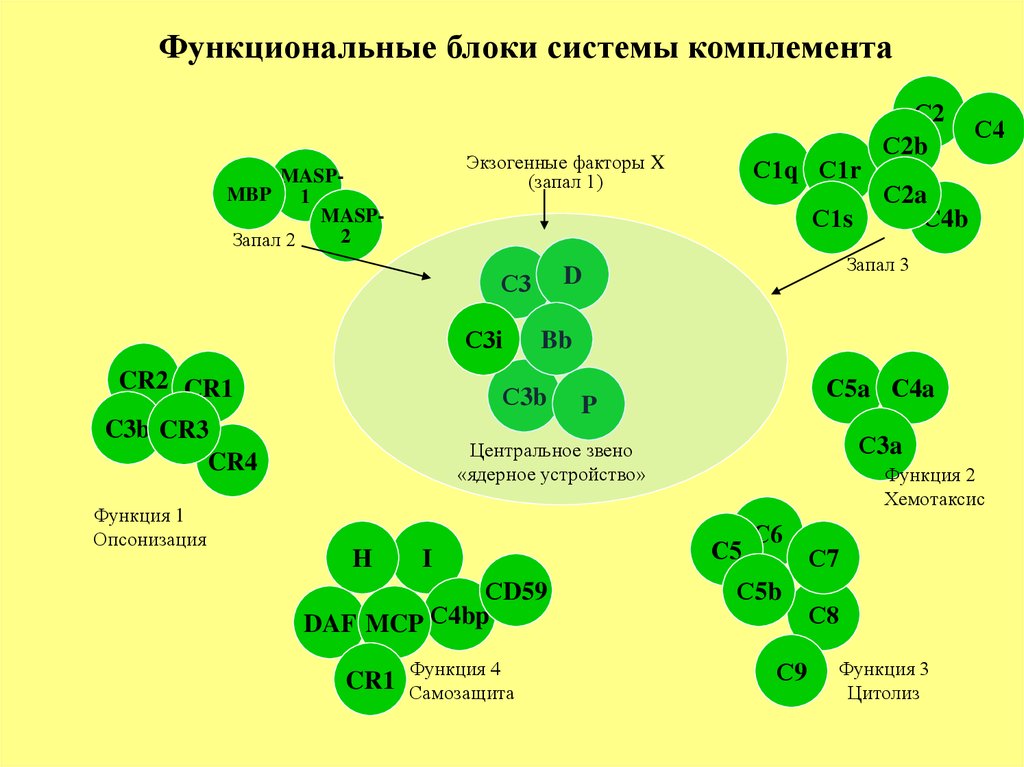
Функциональные блоки системы комплементаЭкзогенные факторы X
(запал 1)
MASPMBP 1
MASP2
Запал 2
С1s
С3i
CR2 CR1
С5a С4a
P
С3a
Центральное звено
«ядерное устройство»
H
I
СD59
DAF MCP С4bp
CR1
Функция 4
Самозащита
С2a
С4b
Bb
С3b
C3b CR3
CR4
С4
Запал 3
D
С3
Функция 1
Опсонизация
С1q С1r
С2
С2b
Функция 2
Хемотаксис
С5
С6
С5b
С9
С7
С8
Функция 3
Цитолиз
49.
50. ЗАЩИТНЫЕ ФУНКЦИИ АНТИТЕЛ
Нейтрализация токсинаУдаление токсина в виде иммунных комплексов
Нейтрализация вируса
Препятствие адгезии бактерий
Активация комплемента на поверхности вирусов,
бактерий и других существ
Опсонизация бактерий, вирусов и других патогенов
(за счет антител и комплемента)
Нацеливание киллеров (через FcR)
Защита границ за пределами организма (IgA)
51.
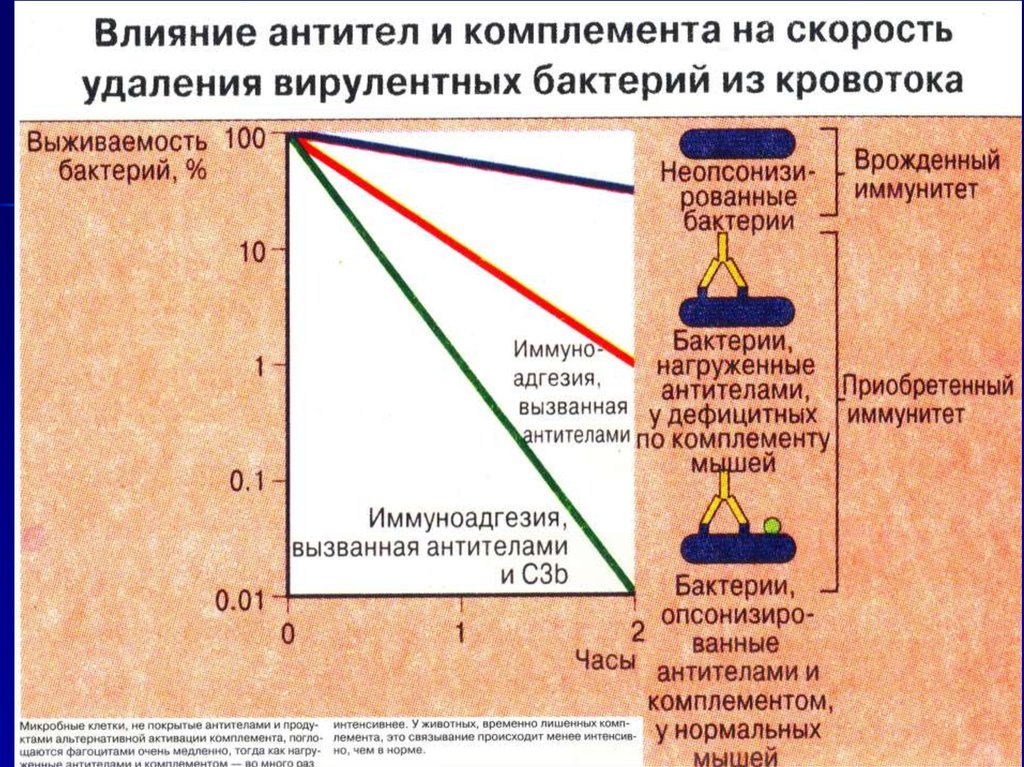
Антитела и комплемент синергичныпри опсонизации микроорганизмов
52.
53.
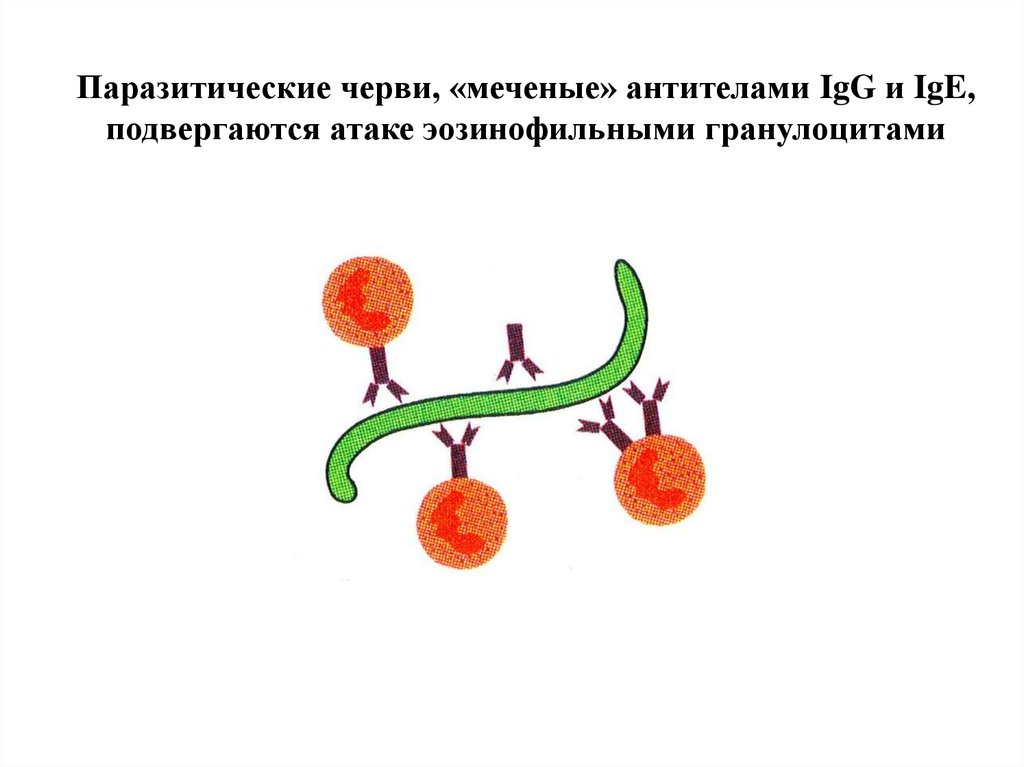
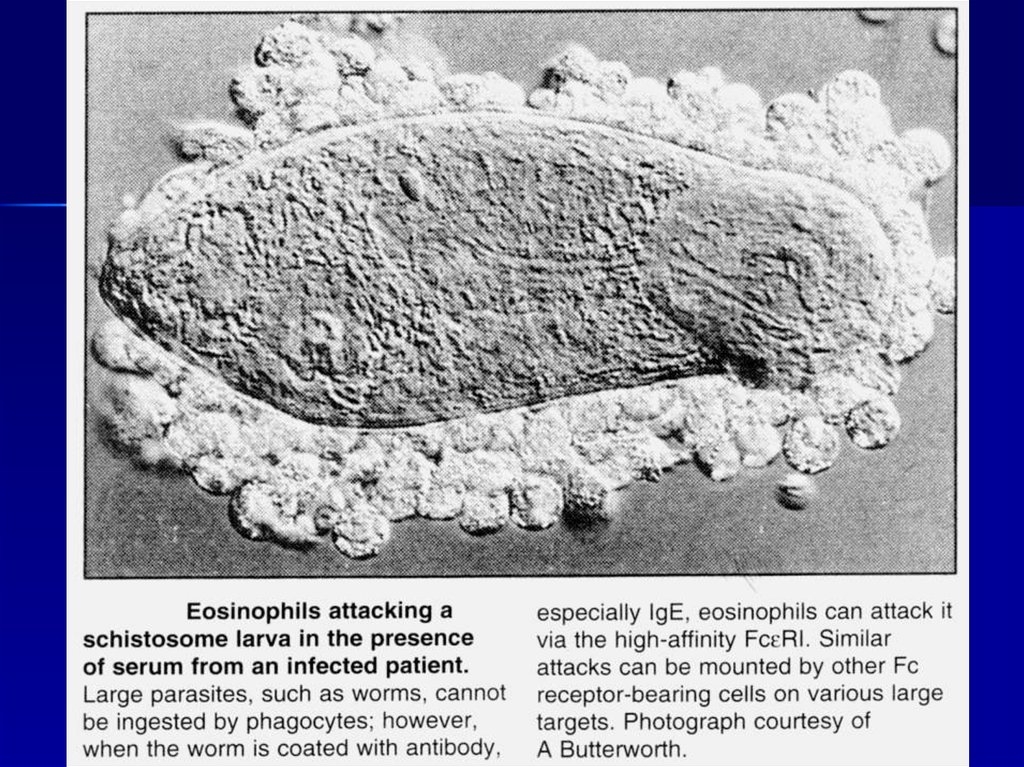
Антитела и комплемент повреждают даже многоклеточных паразитов54.
55.
Антитела в роли «черной» метки,по которой клетки-киллеры атакуют паразитов
56.
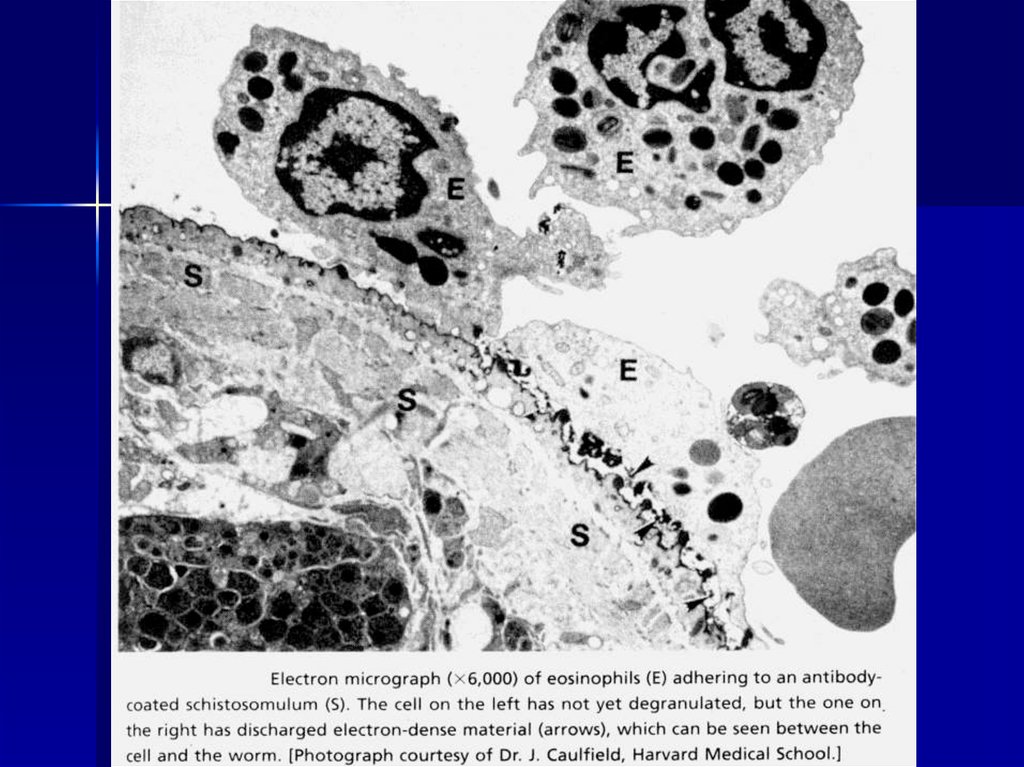
Паразитические черви, «меченые» антителами IgG и IgE,подвергаются атаке эозинофильными гранулоцитами
57.
ЭОЗИНОФИЛ58.
Содержимое гранул эозинофила токсичнодля многоклеточных паразитов
и некоторых бактерий:
Eosinophil granule major basic protein
//en.wikipedia.org/wiki/Major_basic_protein
Образует кристаллическое ядро эозинофильной гранулы,
участвует в защите от паразитов как цитотоксин. 117 аа мощный
фермент против гельминтов, токсичен в отношении некоторых
бактерий.
Может оказывать токсические эффекты на организм хозяина:
высвобождение гистамина из тучных клеток и базофилов,
активация нейтрофилов и макрофагов, повреждение
эпителиальных клеток, бронхоспазм
Eosinophil peroxidase
окисилитель, образует гипохлорит анион и другие активные
субстанции с бактерицидной активностью
RNAse, DNAse, Collagenase
Defensins
Histamine
http://en.wikipedia.org/wiki/Eosinophil_granulocyte
MBP
59.
60.
61.
62.
Антитела защищают нас не только внутри,но и снаружи,
на границе с внешним миром
63.
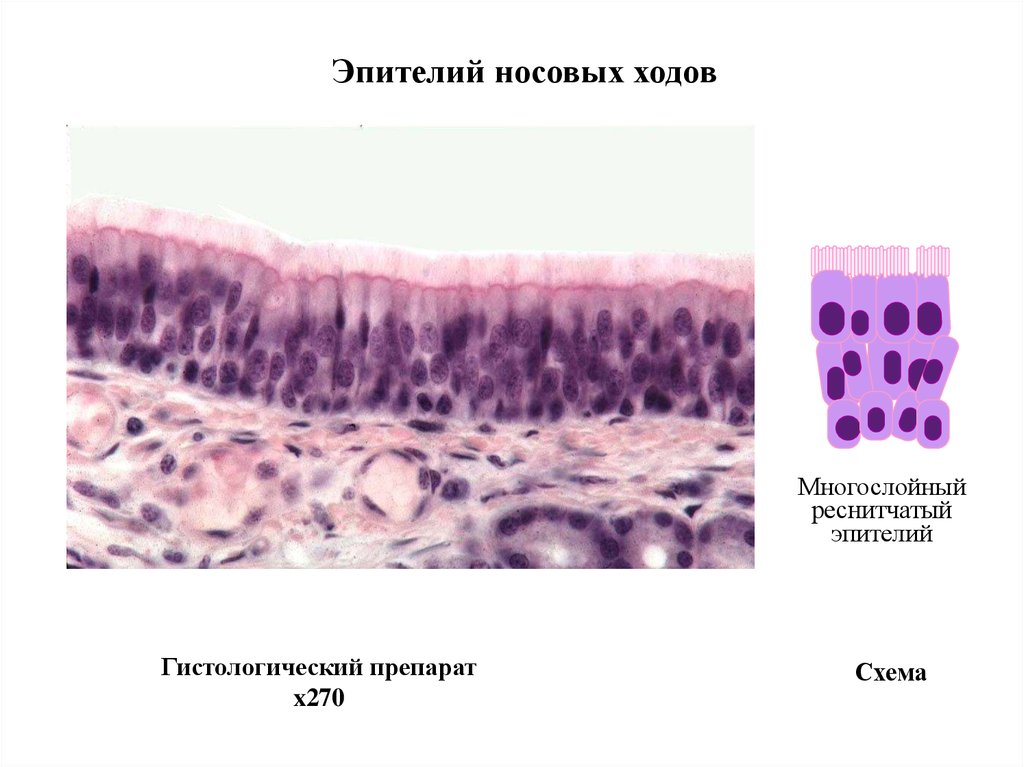
Эпителий носовых ходовМногослойный
реснитчатый
эпителий
Гистологический препарат
x270
Схема
Схема
64.
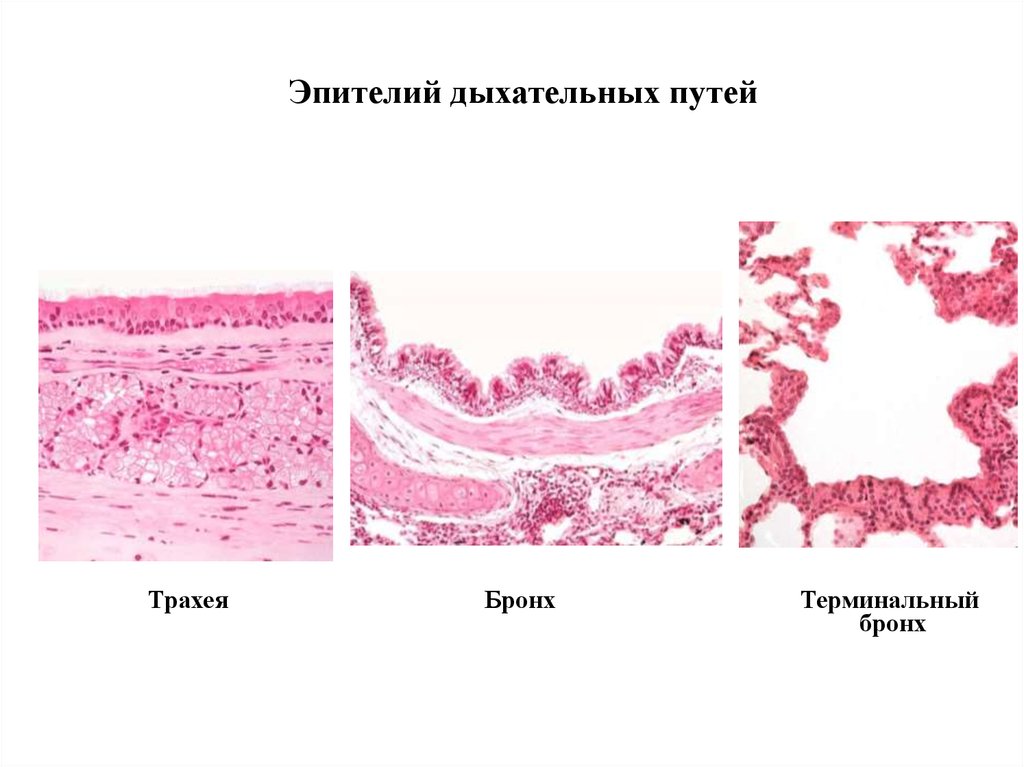
Эпителий дыхательных путейТрахея
Трахея
Бронх
Терминальный
бронх
65.
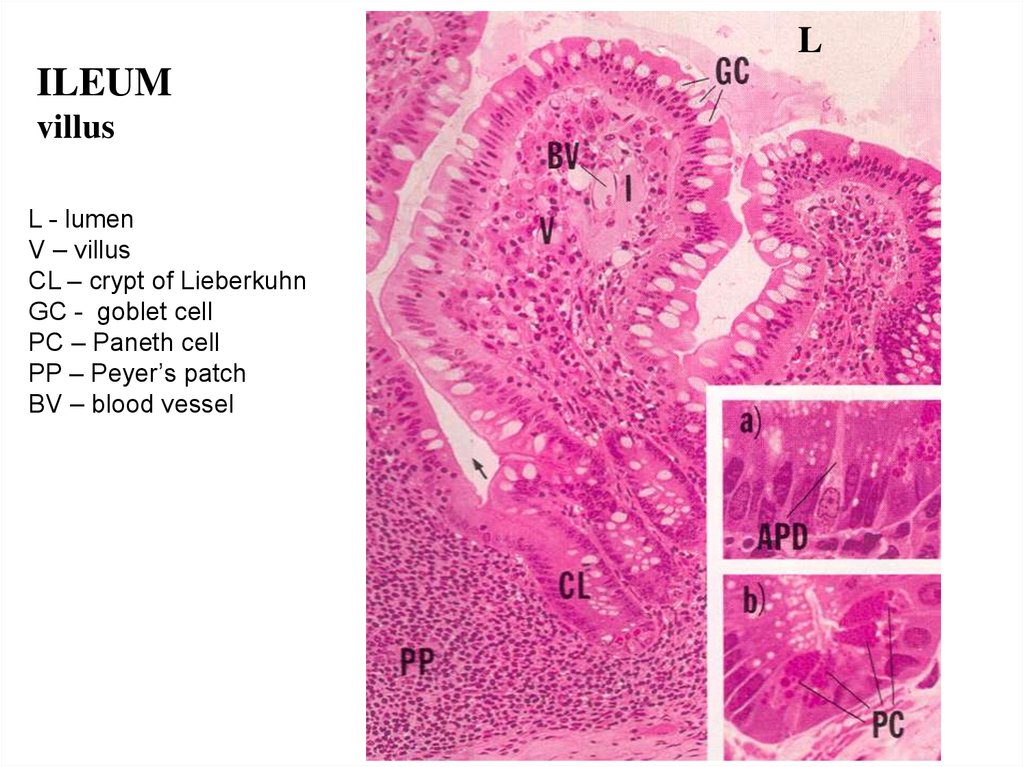
LILEUM
villus
L - lumen
V – villus
CL – crypt of Lieberkuhn
GC - goblet cell
PC – Paneth cell
PP – Peyer’s patch
BV – blood vessel
66.
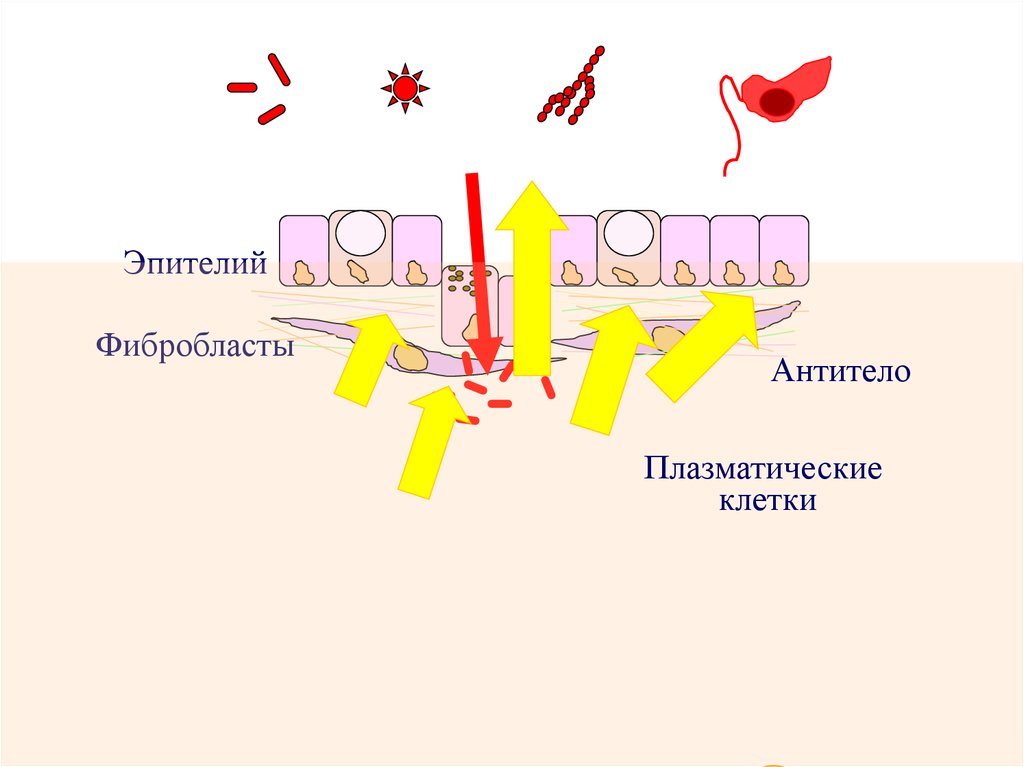
БактерииВирусы
Грибы
Простейшие
Эпителий
Фибробласты
Антитело
Плазматические
клетки
67.
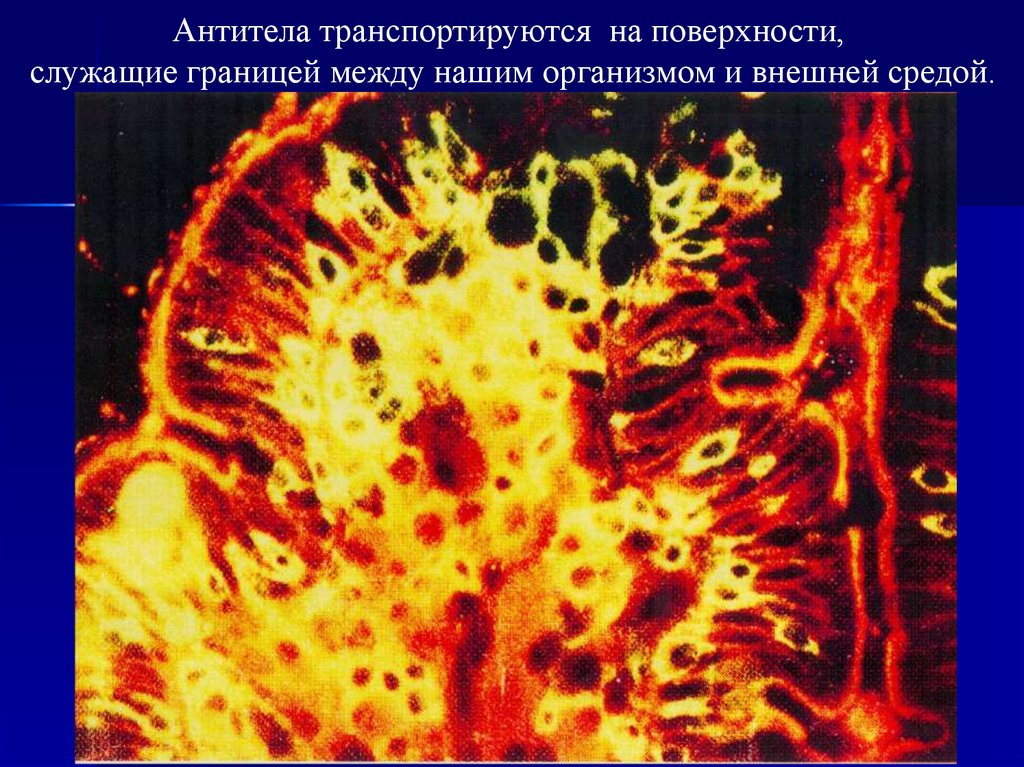
Антитела транспортируются на поверхности,служащие границей между нашим организмом и внешней средой.
68.
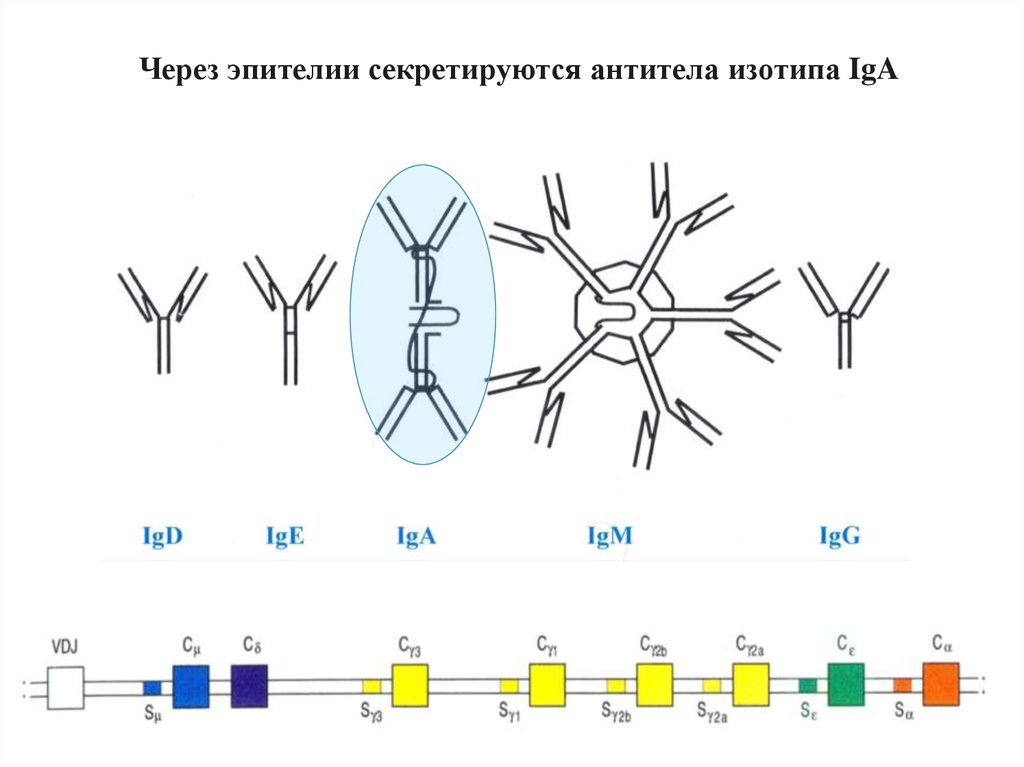
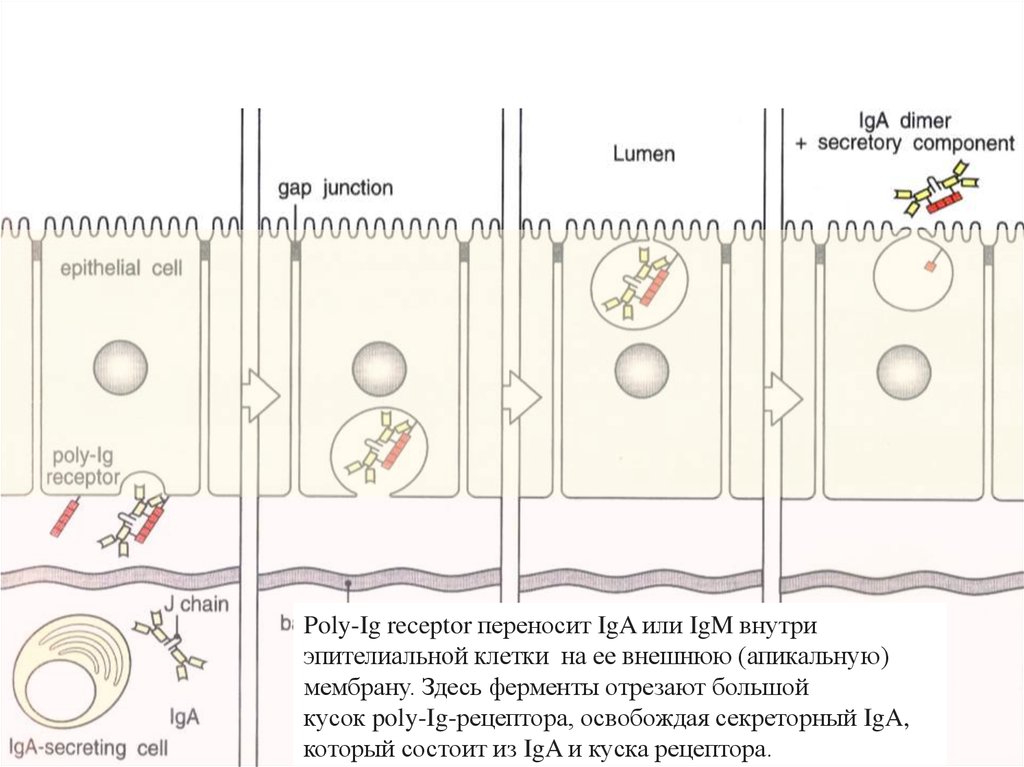
Через эпителии секретируются антитела изотипа IgA69.
Poly-Ig receptor переносит IgA или IgM внутриэпителиальной клетки на ее внешнюю (апикальную)
мембрану. Здесь ферменты отрезают большой
кусок poly-Ig-рецептора, освобождая секреторный IgA,
который состоит из IgA и куска рецептора.
70.
Особенно эффективно poly-Ig-рецептор переносит антитела черезэпителий молочной железы и печени.
В случае молочной железы это обеспечивает ребенка молоком, в котором
есть антитела матери.
В случае печени – секретирующиеся антитела попадают вместе с желчью в
кишечник.
71.
Что они делают?Они очищаются от паразитов
72.
Антитела сообщают собаке (лошади):1. О том, что на теле поселился паразит
2. Точно указывают место, где этот паразит сидит
3. Заставляют неистово избавляться от паразита
73.
Антитела изотипа IgE синтезируются в очень малых количествахв ответ на вторжение малых количеств чужих антигенов,
их концентрация в крови не выше 50-100 нг/мл.
Антитела IgG производятся в больших количествах (до 1-10 мг/мл)
в ответ на вторжение больших количеств чужих антигенов.
В этом случае IgG в 100 000 раз больше, чем IgE.
74.
Зуд и воспаление кожи направлены на механическое удаление паразита.Те же механизмы
вызывают
усиленное антигена
выделение слизи
При малых
концентрациях
на поверхности кишечника и дыхательных путей,
производится
мало антител
класса IgG,
доминируют
антитела
класса IgE
а также импульсивное
сокращение
гладких
мышц кишечника
и бронхов.
Задача та же – удалить паразита.
Паразит
Эпителий
Антигены паразита
IgE
[Fc-рецептор + IgE]
Ка ~1010
Тучная клетка,
эозинофил
Клетка, секретирующая
антитела класса IgE
75.
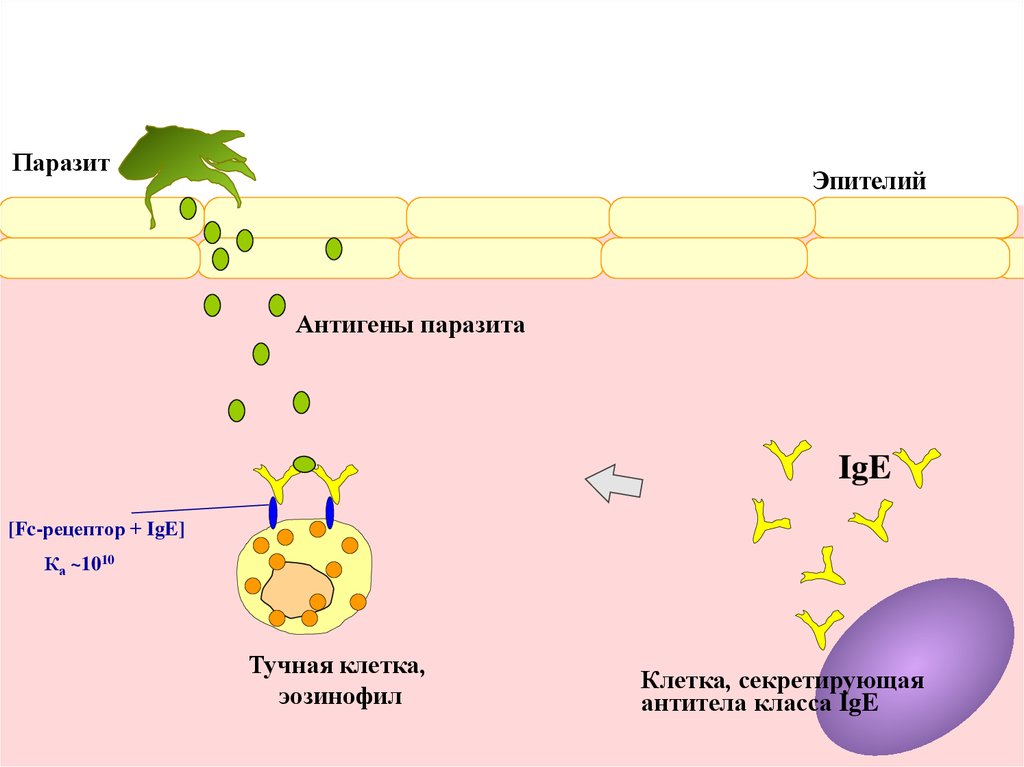
Зуд и воспаление кожи направлены на механическое удаление паразита.Те же механизмы вызывают усиленное выделение слизи
на поверхности кишечника и дыхательных путей,
а также импульсивное сокращение гладких мышц кишечника и бронхов.
Задача та же – удалить паразита.
Паразит
Эпителий
Антигены паразита
IgE
[Fc-рецептор + IgE]
Ка ~1010
Тучная клетка,
эозинофил
Клетка, секретирующая
антитела класса IgE
76.
Антитела IgE служат рецепторами антигенов вмембране тучных клеток.
Это позволяет тучным клеткам запускать следующие
защитные реакции:
• воспаление в ткани, где паразитирует червь или
насекомое,
• зуд, который точно указывает локализацию паразита
и побуждает к действиям, направленным на его
удаление
77.
Изотипы антител имеют разные защитные функцииМономеры IgD и IgM служат рецепторами на поверхности B-клеток
Пентамеры IgM – ранние антитела с низкой аффинностью
и высокой авидностью
Димеры IgА – секретируются на эпителиях, чтобы защищать
внешние границы тела
Мономеры IgE имеют удлиненный «хвост», которым они прочно
присоединяются к Fcε-рецепторам на тучных клетках.
Если антиген попадает вновь, то тучные клетки дегранулируются,
выбрасывая гистамин и другие вещества гранул, что вызывает сильный зуд
и воспаление кожи, выделение слизи и сокращение гладких мышц в стенке
слизистых оболочек.
78.
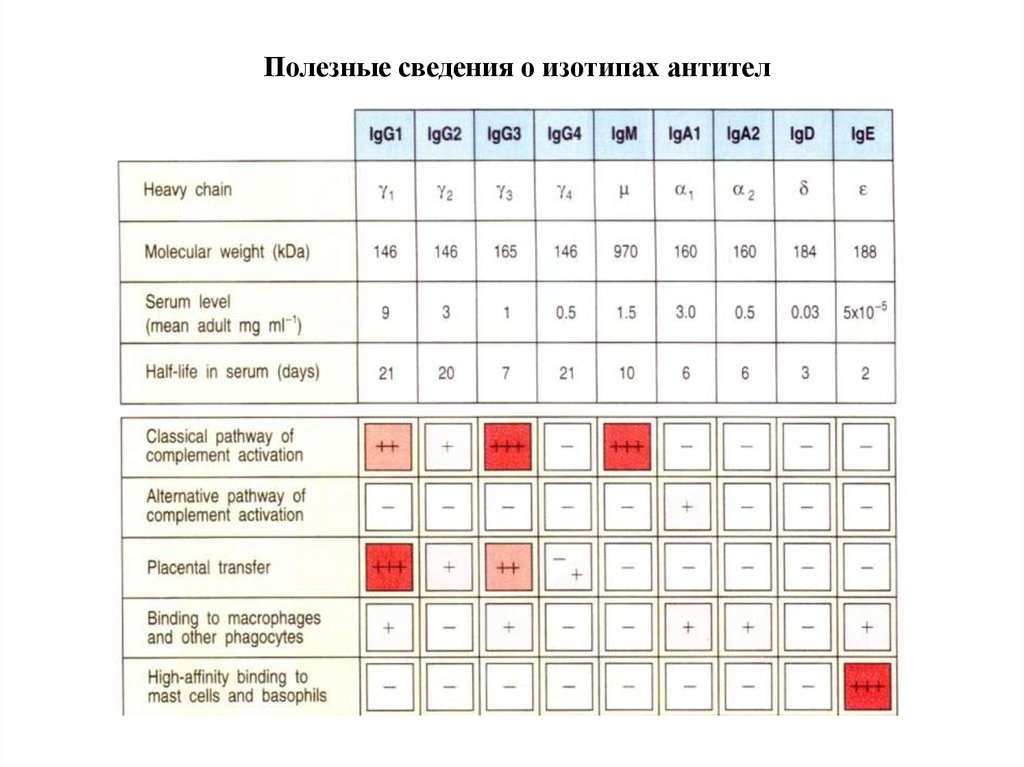
Полезные сведения о изотипах антител79. ЗАЩИТНЫЕ ФУНКЦИИ АНТИТЕЛ
Нейтрализация токсинаУдаление токсина в виде иммунных комплексов
Нейтрализация вируса
Препятствие адгезии бактерий
Активация комплемента на поверхности вирусов,
бактерий и других существ
Опсонизация бактерий, вирусов и других патогенов
(за счет антител и комплемента)
Нацеливание киллеров (IgG, IgE)
Защита границ за пределами организма (IgA)
Зудящие реакции (IgE)