



























 Химия
ХимияПохожие презентации:
, қызметі, май қышқылдары")









Липидтер метаболизмі
1.
“Биохимия” пәніТақырыбы: “Липидтер метаболизмі”
Дәріскер: химия ғылымдарының кандидаты, профессор С.М. Бәзілбаев
2. Дәрістің мақсаты:
Липидтердіңқұрылысы,
қасиеттері
және
молекуласының түзілуіндегі байланыстың түрлері
туралы жалпы түсінік бере отырып, метаболизмін
түсіндіру.
Негізі терминдер:
ЛИПИДТЕР,
ФОСФОЛИПИДТЕР,
ГЛИКОЛИПИДТЕР,
СУЛЬФОЛИПИДТЕР, ЛИПИАЗА, КОЛИПАЗА, ӨТ ҚЫШҚЫЛДАРЫ, ХОЛ
ҚЫШҚЫЛЫ,
ХЕНОДЕЗОКСИХОЛ
ҚЫШҚЫЛЫ,
ДЕЗОКСИХОЛ
ҚЫШҚЫЛЫ, ЛИТОХОЛ ҚЫШҚЫЛЫ.
3. Дәрістің жоспары:
1.2.
3.
4.
5.
6.
Майлардың қарында қорытылуы.
Майлардың ішекте қорытылуы.
Өт қышқылдары.
Май қышқылдарының бауырдағы метаболизмі.
Май қышқылдарының биосинтезі.
Май қышқылдарының -тотығу жолы
4. ЛИПИДТЕРДІҢ ҚОРЫТЫЛУЫ ЖӘНЕ СІҢІУІ
Липидтердің табиғи көзі:майлы өсімдектекті және жануартекті
тағам.
Қалыпты жағдайда липидтер
рационның калориясының 40%
құрайды: 80 г/тәулік –70 %-ға дейін
ҮАГ, 10% – ФЛ және ХС, 3%-кем
емес – ПҚМҚ.
5.
Ауыз қуысы – липидтерді қорытатын фермент жоқ.Асқазан (қарын) – үлкен адамдарда асқазан сөлінде
липаза өте аз болады және майлар іс жүзінде
қорытылмайды:
асқазанда рН ≈ 1,5, ал липаза рН = 5,5-7,5 активті;
Липазаның активтілігі майлы фазаның бетіне тура
пропорциональды, ал майлар асқазанда
эмульгирленбейді (эмульцияланбайды);
Балаларда (емшек сүтімен қоректенетін)
липидтердің асқазанда қорытылуы маңызды:
рН ≈ 5, сүттің майы жоғарыэмульгирленген,
липаза көп мөлшерде бөлінеді;
Он екі елі ішек – майлардың қорытылатын орны
он екі елі ішекке ашылады:
Ұйқы безінің өзегі (ферменттер және Н+ активтілігін
төмедететін сілтілік факторлар)
Өт өзегі (бауырдан өт түседі және майларды эмульциялайды)
6.
Майлардың ішекте қорытылуыМайлардың негізгі қорытылуы он екі елі ішекте өтеді,
оған әсер етуші ұйқы безінің липаза ферменті. Мұнда
қоректік заттар жиынтығы өттің, ұйқы безі сөлінің
және
ішек
секреттерінің
әсеріне
ұшырайды.
Липидтердің қорытылуында және олардың гидролизі
өнімдерінің сіңуінде өт қышқылдары маңызды роль
атқарады.
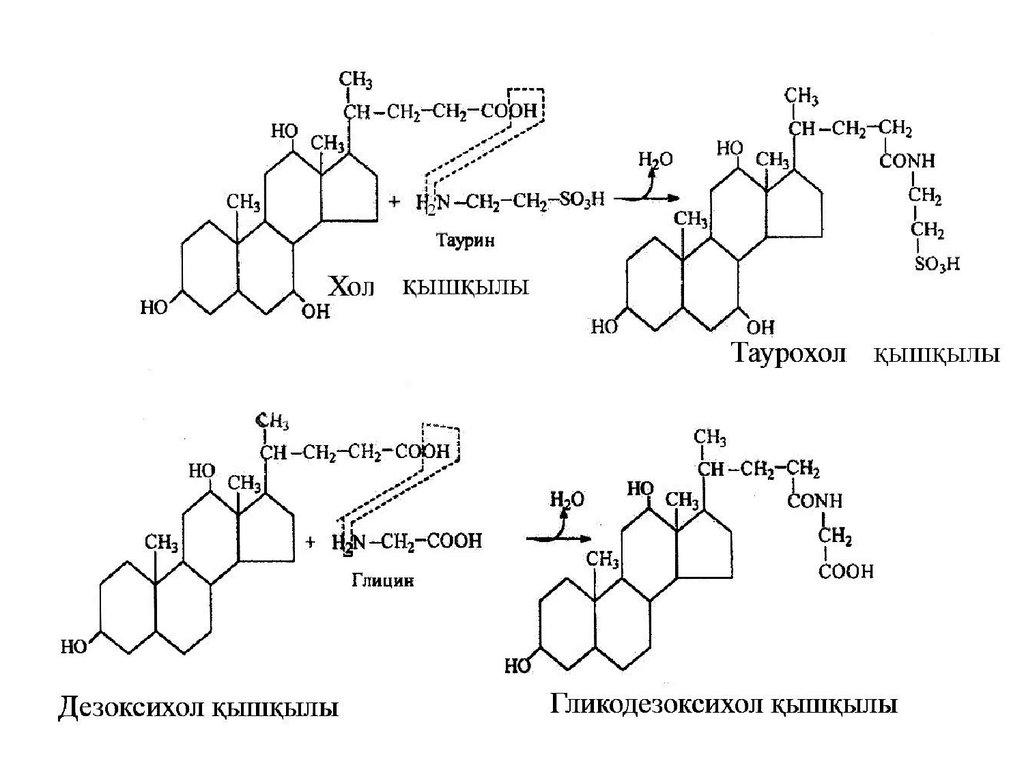
Өт қышқылдары
Өт қышқылдары өтпен бірге өт қалтасынан
шығады. Төрт түрлі өт қышқылы белгілі. Олар: хол
қышқылы, хенодезоксихол қышқылы, дезоксихол
қышқылы, литохол қышқылы.
7.
Бұл қышқылдар өт құрамында тиісінше 50,30, 15, 5% мөлшерінде кездеседі. Бұлар
пептидтік байланыс арқылы глицинмен
немесе тауринмен байланысқан. Соның
әсерінен осы қышқылдар ас қорытушы
барлық ферменттерге төзімді келеді.
Мұндай комплексті байланысу өт
қышқылдарының ерігіштігін жақсартады.
Өт қышқылдарының байланыспаған
тұздары рН көрсеткіші 7-ден төмен кезде
тұнбаға шөгеді.
8.
Өт қышқылдарының глицинмен немесе тауринменқосылған комплекстері натрий тұздары түрінде
кездеседі.
Өт қышқылдарының қызметі - липидтердің
эмульсияға айналуын және еруін дамыту. Сөйтіп
липаза ферментінің әсерін жеңілдету.
Төменде хол қышқылы мен тауриннің жұп
қосылысы таурохол қышқылы және дезоксихол
қышқылы мен глициннің жұп қосылысы
гликодезоксихол қышқылының түзілу реакциялары
берілген.
9.
10.
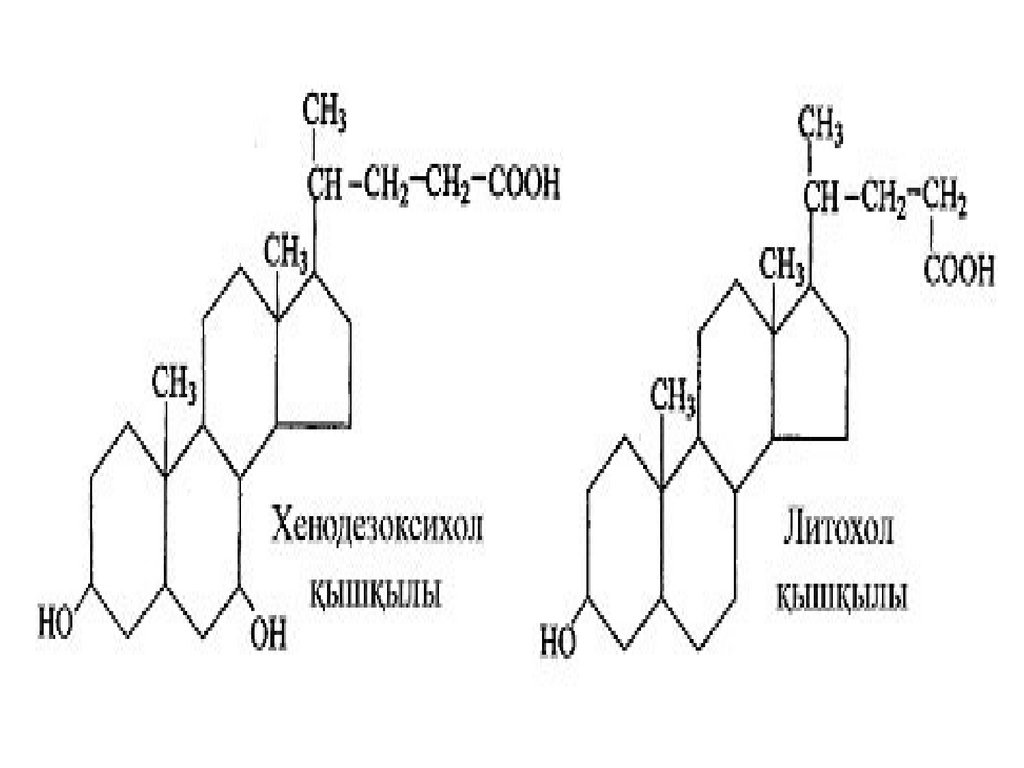
Өт қышқылдары құрылым жағынан бір біріне ұқсас, олар тек қана молекуласындагироксил
топарының
саны
және
орналасуымен ерекшеленеді.
Төменде
екі
қышқылдарының
–
хенодезоксихол қышқылының және
литохол қышқылының формулалары
келтірілген.
11.
12.
Майды эмульсияға айналдыру. Майларгидрофобты,
сулы
ортада
ерімейді.
Өт
қышқылдарының ерігіш амфипатиялық қасиеті
бар. Өт қышқылдары сырттай активті заттар.
Өйткені олардың стероидтық жағы гидрофобты,
ал полярлы «басы» (қышқылдық топ бар жері)
гидрофильді
келеді.
Өт
қышылдарының
гидрофобты жағы май түйіршіктерімен өзара
әрекеттеседі, ал олардың гидрофильдік тобы сулы
ортаға қараған жақ бетке жиналып топтасады да,
полярлы еріткіш сумен түйіседі.
13.
Өт қышқылдары, бос май қышқылдары менмоноацилглицеролдардың қатысында май
түйіршіктері - сыртына жинақталады да
жұқа қабықша түзеді. Осының нәтижесінде
екі заттың- су мен майдың арасындағы
сыртқы керіліс бірден азаяды. Осылайша
беттік керілістің азаюы және ішектің
жиырылып созылуы нәтижесінде үлкен
май түйіршіктері босайды да, бөлшектеніп,
ұсақ-ұсақ май түйіршіктеріне айналады.
14.
Осылай пайда болған майдың өте ұсақбөліктерінің сыртқы қабатында сол беттік
активті заттардан құралған жұқа қабықша
болады, ол қабықша май бөлшектерін
біріктірмейді. Май осылайша эмульсияланады да суда еритіндей қалыпқа түседі.
Эмульсияланғаннан кейін майдың беттік
аумағы өте ұлғаяды. Эмульсиялану деңгейі
жоғары болған сайын және май түйіршіктері
кішірейген сайын оған ферменттің әсер етуі
оңайланды. Сөйтіп ацилглицеролдар
гидролизінің жылдамдығы да артады.
15.
Өттің және ұйқы безі сөлінің реакциясы әлсіз сілтілік,ол қарынның қышқыл реакциясын бейтараптайды.
Триацилглицеролдар гидролизі. Май
түйіршіктерінің сыртына колипаза белогы (М 10 000)
тартылады. Сөйтіп сол жерде ұйқы безінің липаза
ферменті де жинақталады. Колипаза қызметі молярлық қатынасы 2 : 1 (липаза : колипаза) комплекс
түзу арқылы липаза ферментін активтендіру. Осы
кезде рН көрсеткіші 9-дан 6-ға дейін өзгереді. Бұл
липаза ферментінің әсері үшін ең қолайлы жағдай.
16.
Ал колипаза липазаны май түйіршіктерініңбетінде ұстап тұрады. Өт қышқылы, колипаза
және липаза үшеуі өзара әрекеттесіп, үш жақты
комплекс түзеді деген болжам бар. Липаза әсерін
күшейту үшін кальций иондары қажет.
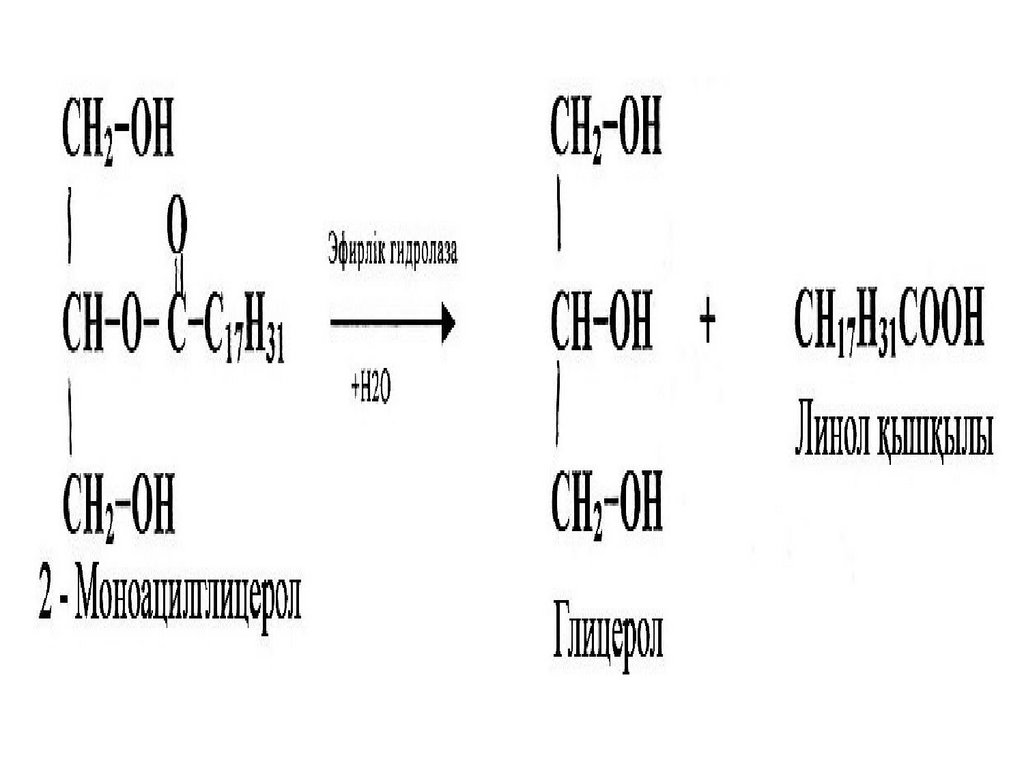
Ұйқы безінің липаза ферменті триацилглицеролдар құрамындағы негізінен 1 және 3
жағдайдағы май қышқылдарын босатады да, 2моноацилглицерол түзеді. 2-моноацилглидеролдар гидролизін ұйқы безінің басқа
ферменті – эфирлік гидролаза катализдейді.
17.
18.
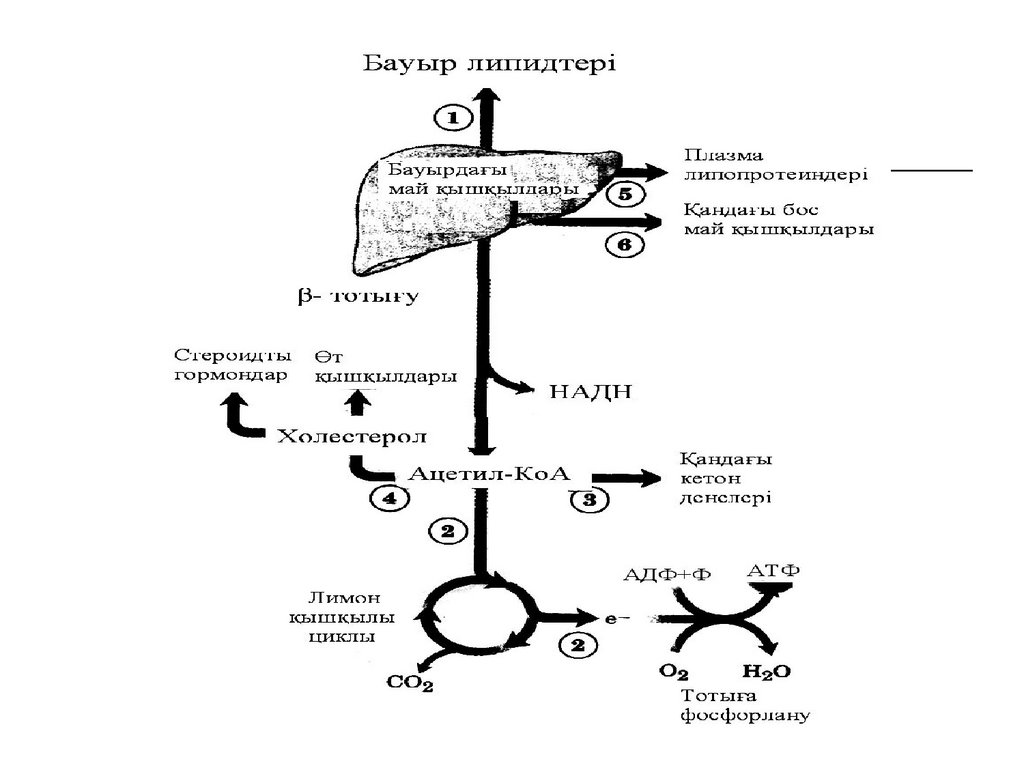
Май қышқылдарының бауырдағыметаболизмі
Қанмен келген липид негізінде бауырда
тоқтап қалады, әр түрлі өзгерістерге
айналады, катаболизмдік және анаболизмдік
реакцияларға
қатысады.
Жоғарыда
келтірілген
липидтерге
байланысты
химиялық процестер көбінесе бауырда
жүреді. Сол жазылған реакцияларға сүйене
отырып май қышқылдарының бауырдағы
метаболизмін қорытынды түрде берейік
19.
20.
1. Май қышқылдары липогенез процесінеқатысып, бауыр липидтерін құрайды.
2. -тотығуға ұшырап, ацетил-КоА молек
улаларына ыдыратады. Олар лимон
қышқылы циклымен тотығып, тотыға
фосфорлану процесінде АТФ, СО2 және
Н2О түзіледі.
21.
3. Ацетил-КоА-дан кетон денелерітүзіледі, қанға енеді.
4. Ацетил-КоА-дан холестерол синтезделеді,
одан
өт
қышқылдары,
стероидтық гормондар түзіледі. Қан
плазмасындағы
липопротеиндер
липидтерінің синтезіне қатысады.
5. Бос май қышқылдары бауырдан қанға
енеді
22. Липидтердің метаболизмі
Май қышқылдарының тотығуы митохондрияда жүреді, үшкезеңнен тұрады:
1) активтенген май қышқылы ацил-КоА β-тотығуға түсіп,
молекуласынан екі көміртек атомы бар ацетил-КоА-лар бөлінеді;
2) түзілген ацетил-КоА молекулалары лимон қышқылы
циклымен тотығып, электрондар, НАДН, ФАДН2 және СО2
ыдырайды;
3) электрондар, НАДН, ФАДН2 тыныстану тізбегіне кіреді де,
тотыға фосфорлану процесінде электрондардың энергиясы АТФ
молекулаларында шоғырланады.
1904 ж. Ф. Кнооп: май қышқылдары бірден көміртегінің екі
атомын бөліп шығару жолымен тотығады, сөйтіп, бастапқы
қышқыл көміртегінің екі атомына қысқаратындығын анықтады
және бұл процесті -тотығу деп атайды, өйткені әр жолы май
қышқылдарының -көміртегі атомы тотығады.
23. Май қышқылдарының -тотығуы
Май қышқылдарының -тотығуы-тотығудың механизмі алғаш рет 1904 ж. Кнопп ашты
МҚ С β-атомы арқылы:
МҚ молекулаларынан 2С-фрагменты Ас~КоА түрінде бөлінеді;
1 этап – ацил-КоА-синтетазамен (АТФ) МҚ активтелуі
RCOOH + HS-КоА + АТФ → RCO~SКоА + АМФ + ФФ + H2O
β-тотығу митохондрияда жүреді (бауыр, миокард, өкпе…).
Бос май қышқылдары да, олардың активті түрлері де
митохондриялық ішкі мембрана арқылы цитозольдан
матрикске өте алмайды. Оларды карнитин тасымалдайды. 810-ға дейін С атомдары бар МҚ МХ-ға карнитинсіз
тасымалданады.
24. Ацил-КоА дегидрогенденуі
Ацил-КоА транс-қанықпаған туындылары түзіледі жәнебұл реакцияны коферменті ФАД болатын ацил-КоАдегидрогенеза ферменті катализдейді:
Ацил-КоА
Транс-қанықпаған ацил-КоА
25. Транс-қанықпаған ацил-КоА гидраттануы
Қос байланыс бойынша транс-қанықпаған ацил-КоАгидраттанып,
-гидроксиацил-КоА
L-стереоизомері
түзіледі. Бұл реакцияны еноил-КоА-гидратаза ферменті
катализдейді:
Транс-қанықпаған ацил-КоА
L- -гидроксиацил-КоА
26. -Гидроксиацил-КоА дегидрогенденуі
-Гидроксиацил-КоА дегидрогенденуі-Гидроксиацил-КоА дегидрогенденуі, -кетоацилКоА түзіледі және бұл реакцияны коферменті НАД+
болатын
-гидроксил-КоА-дегидрогеназа
ферменті
катализдейді:
L- -гидроксиацил-КоА
-кетоацил-КоА
27. -Кетоацил–А-коферментінің ыдырауы
-Кетоацил–А-коферментінің ыдырауы-кетоацил-КоА
ацетил-КоА
және
ацил-КоА-ға
ыдырайды және бастапқы активтенген май қышқылы
молекуласымен салыстырғанда, пайда болған ацил-КоА
молекуласында көміртегінің екі атомы кем. Бұл реакция
кейде тиолиз деп аталады және тиолаза ферменті
катализдейді:
-кетоацил-КоА
КоА
ацил-КоА
ацетил-
28. 2 этап –β-тотығу
R-CH2-CH2-CO~SКоАацилКоА-дегидрогеназа
2АТФ
R-CH=CH-CO~SКоА
гидратаза
R-CHОН-CH2-CO~SКоА
β-гидроксиацил-ДГ
3АТФ
R-CО-CH2-CO~SКоА
тиолаза
R-CО~SКоА
+
CH3-CO~SКоА
Пальмитин қышқылы 16С=14С+2С
29. β-тотығудың энергетикалық маңызы
16С пальмитин қышқылында 7 (ФАДН2+ НАДН = 5 АТФ)
5 х 7 = 35 АТФ
Ас~КоА –катаболизмінің жалпы жолы
(ҮКҚА) Н2О мен СО2 және 12 АТФ
түзіледі
16С : 2 = 8 Ас~КоА → 8 х 12 = 96 АТФ
Барлығы
35 + 96 = 131 АТФ
1 сатыны есепке алсақ
131 – 1 = 130 АТФ
30. Бақылау сұрақтары:
1.2.
3.
4.
5.
Липидтер дегеніміз қандай қосылыс?
Липидтердің ішекте қорытылуына қандай
фермент әсет етеді?
Липидтердің метаболизмінде өт қышқылдарының
қызметі қандай?
Липидтер гидролизінен түзілген май қышқылдары
неге жұмсалады?
Липидтердің биологиялық маңызы неде?
31. 1. Сейітов З.С., Бейсебеков М.Қ. Физикалық және коллоидтық химия. –Алматы, 1993. 2. Қайырханов Қ.К. Жануарлар биохимиясы.
Әдебиеттер:1. Сейітов З.С., Бейсебеков М.Қ. Физикалық және
коллоидтық химия. –Алматы, 1993.
2. Қайырханов Қ.К. Жануарлар биохимиясы. –Алматы:«Ана
тілі», 1993.
3. Сеитов З.С. Биохимия.– Алматы: «Қазақ университеті»,
2001.
4. Салханова С.Н. Және т.б. Жануарлар биохимиясы пәнінен
студенттерге арналған оқу-әдістемелік кешен, 2013.
32.
Тапсырма:1.
“Липидтер” тақықырыбына глоссарий жазу
(тәжірбиелік сабаққа жазып келу керек).
Зейін қойып тыңдағандарыңызға рахмет!