











")
")
")
")
")
 и")
 расположена ростральнее ВРГ Анатомически и функционально нейроны пФРГ отличаются от")

















 Биология
БиологияПохожие презентации:







")


Регуляция дыхания. Дыхательная система
1. Регуляция дыхания
Лекция проф. Н. П. Ерофеевадля пульмонологов и всех, живущих в
воздушном океане
Отдельную благодарность выражаю
академику Александру Григорьевичу
Чучалину за неугомонный поиск новых
знаний в огромном объеме современной
информации
2. Дыхательная система обеспечивает оптимальный уровень метаболизма, определяемый физиологическим состоянием организма (покой,
нагрузки, болезнь)Дыхательная система
реализует свое
предназначение путем
регуляции вентиляции
легких и просвета
дыхательных путей
В результате дыхательная
система создает полное
соответствие между количеством
О2, поступающего через лёгкие в
кровь и скоростью его потребления
в тканях и такое же соответствие
между продукцией в тканях СО2 и
удалением его из лёгких
Итог: вентиляция лёгких
подгоняет газообмен в лёгких к
метаболизму в тканях и делает его
адекватным функциональному
состоянию человека
3. Но кажется зачем необходима регуляция?
Когда:Не успел подумать, а уже сделал
вдох
По команде или
самопроизвольно можно
задержать дыхание или провести
гипервентиляцию
Более того - во сне у человека
изменяется ритм дыхания
При общем наркозе человек
способен самостоятельно
дышать
С другой стороны: никогда нельзя по
своей воле надолго задержать дыхание
так, чтобы произошел обморок
К тому же существуют
общепризнанные факты:
• — у здорового человека ритм дыхания,
как и ритм сердечных сокращений,
задается автоматической
деятельностью систем клеток и не
требуют сознательного участия
человека;
• —периодичность дыхательных
движений по воле человека может
быть изменена, хотя добровольный
контроль ограничен по времени;
• — сильные механизмы нейронного
контроля достаточно быстро подавляют
сознательный контроль после того, как
человек намеренно прекратит
дыхание;
— здоровым людям не нужно
беспокоиться о том, чтобы забыть
дышать во время сна или наркоза;
— включение произвольной регуляции
дыхания необходимо для разговора,
пения, смеха и других действий
человека
4. Система контроля дыхания включает три основных звена для управления вентиляцией – потоками воздуха
1- Рецепторы (хемо + механо)Аппарат дыхания как и сердце
человека работают в
определенном постоянном, но
изменяющемся ритме
Помните! Ритм сердца возникает
автоматически-спонтанно и
непрерывно. Генератор заложен в
самом сердце
Дыхание похоже на сердцебиение с
точки зрения автоматического и
непрерывного возникновения даже во
время сна или общей анестезии.
Однако скелетные мышцы,
управляющие вентиляцией, не
сокращаются спонтанно, как это делает
сердечная мышца.
К тому же генератор ритма вдоха и
выдоха лежит вне легких, а в стволе
мозга
2- Центральный регулятор: Medulla
Мотонейроны сегментов спинного
мозга
3- Эффекторы (дыхательные
мышцы)
Обычно повышение активности
дыхательных мышц приводит к
снижению импульсации от
рецепторов в дыхательный центр –
отрицательная обратная связь.
5. Три основных звена дыхательного рефлекса
Основные регуляторы О2 и СО2в артериальной крови
Основные параметры
вентиляции и их резервы
Частота дыхания ЧД – 12-18/мин,
резерв - до 50/мин
Дыхательный объём ДО – 300500 мл, резерв – 1л
Минутный объём дыхания МОД –
5-6 л, резерв – 120 л
Химические параметры крови:
Ро2, Рсо2, рН, резерв этих
показателей очень мал
6. История изучения регуляции дыхания
Долго время представлялось, что особогодыхательного центра не существует, а
дыхание регулирует совокупность нейронов,
находящихся на различных уровнях ЦНС
В 1873 году Ж. Легалуа (J. Legallois)
определил, что ядро контроля
дыхания находится на уровне ствола
мозга, дыхание продолжается даже
после удаления головного мозга
Следует отметить вклад
отечественного физиолога
Н.А.Миславского (1854-1928),
который в своей докторской
диссертации «О дыхательном
центре» ( 1885) описал разнородную
группу ядер дыхательных нейронов,
сосредоточенных в
дорсомедиальной и
вентролатеральной областях
продолговатого мозга
В 80-х годах 20-го века область
регуляции респирацией была сужена до
Medulla ствола мозга В основу организации
дыхания положен автоматический
бессознательный процесс
• Гипотеза водителя ритма
Комплекс preBötzinger
(впервые обнаружен 25 лет
назад) Приблизительно 20%
нейронов preBötC
классифицируются как
пейсмекеры дыхательного
ритма
Комплекс preBötzinger у
взрослого человека
устанавливается
доминирующим
генератором дыхательного
ритма
Нейроны ДРГ исключены
из генераторов ритма
дыхания
7. На сегодняшний день «исчез» их центров регуляции апнейстический центр
Такой представлялась локация центров
управления вдохом и выдохом: центры
рассредоточены по разным уровням
головного мозга
8. Современная схема отделов ствола мозга, управляющих дыханием
Основной генератор ритма дыханиясосредоточен в стволе мозга на уровне
мedulla
Он задает частоту дыхания, хотя
ЧД может модулироваться ПД,
исходящими из отделов коры, и ПД с
периферии, например, от
проприорецепторов
Эти центральные и периферические входы
могут изменять активность дыхательных
мотонейронов и дыхательных центров
Нейроны в мedulla – генераторы
дыхательного ритма создают базисный
автоматический ритм дыхания – паттерн
чередования вдохов и выдохов путем
проекции своих аксонов на дыхательные
мышцы
Люди дышат примерно 20 000
раз в день и сотни миллионов
раз в течение средней
продолжительности жизни
9. Дыхательные движения создающие дыхательный ритм вдохов и выдохов можно разделить на три последовательных фазы: Инспирация –
фаза I, активный процесс вдоха,создаваемый активностью диафрагмального нерва и
нервов наружных межреберных мышц и последующих
сокращений диафрагмы и наружных межреберных
мышц грудной клетки;
Постинспирация – фаза PI, пассивная
экспирация (выдох) – диафрагмальный нерв
постепенно перестает генерировать ПД и после вдоха
диафрагма постепенно прекращает быть активной;
Активная экспирация (выдох) – фаза Е2,
диафрагмальный нерв «молчит», ПД генерирует
внутренний межреберный нерв и нервы брюшной
стенки - сокращаются внутренние межреберные и
брюшные мышцы
Дыхательный ритм и его
фазы прямо отражают нервный
ритм, то есть электрическую
активность- генерацию ПД
моторными нервами диафрагмы,
грудной клетки и брюшной стенки
в фазах I , PI и Е2.
Диафрагмальный нерв и нервы
грудной клетки и брюшной стенки
получает импульсы от групп
медуллярных нейронов
пребётцингеровского комплекса
10. Классы респираторных нейронов и их вклад в формирование фаз дыхательного ритма – А + активность моторных нервов диафрагмы и
экспираторныхмышц - Б
11. Ключевые нейронные элементы для генерации паттерна дыхания различны, компактны и образуют медуллярно-понтинную сеть
Ключевые нейронныеэлементы для генерации
паттерна дыхания
различны, компактны и
образуют медуллярнопонтинную сеть
Первичным центральным
генератором дыхательного
ритма является комплекс
Пребётцингера (preBötC),
который состоит из возбуждающих
ритмогенных интернейронов и
возбуждающих и ингибирующих
интернейронов, которые вместе
производят необходимый
периодический импульс для вдоха и
последующий паттерн дыхательных
фаз.
preBötC координирует не только
все фазы дыхательного цикла, но и
координирует дыхание с глотанием,
вокализацией и т.п. и сильно влияет
на эмоции и поведение
Центральный генератор паттернов
расположен в Medulla рядом с
другими дыхательными центрами,
которые интегрируют
афферентную информацию для
вентиляторных рефлексов c целью
точной настройки моторного
выхода на вентиляционные мышцы
12. Я рассказываю вам самую последнюю информацию о нейронном субстрате и моделях, лежащих в основе управления дыханием
Контроль скорости, ритма и глубиныразличных фаз дыхания
необходим не только для
газообмена, но и формировании не
дыхательного двигательного
поведениия и эмоциональных
состояний.
Сеть генерирующих ритм и
паттерн нейронных групп в понтомедуллярной области мозга
контролирует активность
инспираторных, экспираторных и
вспомогательных мышц дыхания.
Эта сеть тесно связана с высшими
подкорковыми и корковыми
областями мозга для модуляции
дыхания и поведения.
Сеть также получает
афферентную информацию от
периферии, чтобы модулировать
текущую дыхательную активность,
так чтобы дыхание – вентиляция
соответствовала метаболическим
потребностям
Я рассказываю вам самую
последнюю информацию о
нейронном субстрате и
моделях, лежащих в основе
управления дыханием
13. Вентральная респираторные группа нейронов(ВРГ)
Нейроны компактнораспределены в виде колоннстолбов в n. Ambiguus и n.
Retroambiguus , состоящих из
четырех сегментов
• Комплекс Бётцингера
Комплекс пре-Бётцингера
Ростральный сегмент ВРГ
(рВРГ)
Каудальный сегмент ВРГ (кВРГ)
Анатомическая карта
дыхательных генераторов в
вентральном столбе
14. Комплекс Бётцингера (Бётц. К)
— этопередний регион ВРГ, анатомически
перекрывающийся с
ретротрапециевидными ядрами и
парафациальной респираторной
группой (РТЯ/пФРГ)
Комплекс Бётцингера содержит
популяции глицинергических
экспираторных нейронов,
которые вносят вклад в
дыхательный ритм, но не
считаются независимыми
генераторами ритма
Их аксональные проекции,
оканчивающиеся на медуллярных и
спинномозговых мотонейронах,
активируют экспираторные мышцы
при нормальном дыхании
Бётц. К, тормозя вдох, участвует в
формировании фазы постинспирации
(РI) и при необходимости усиливает
экспираторную активность в фазе
активного выдоха (Е2)
Комплекс Бётцингера (Бётц. К)
15. Пре-Бётцингера комплекс (пре-БК)
Пре-БК был локализован в 1991 г. коллективом подруководством Джека Фелдмана в составе ВРГ.
Нейроны пре-БК обладают спонтанной допороговой
электрической активностью даже в изолированном
виде; их удаление или блокирование их связей
ведет к остановке дыхания
Ритм активности пре-БК порождается
эндогенными колебаниями возбудимости, поэтому
респираторные нейроны пре-БК относят к типу
пейсмекерных нейронов
Сеть нейронов, образующих пре-БК,
признана центральным генератором
ритмогенеза — создателем базового ритм
дыхания – инспираций и контролирует
функции расширения дыхательных путей
гортани, глотки и положение языка
В дальнейшем созидаемый пейсмекерными клетками преБК нервный ритм дыхания напрямую (моносинаптически)
передается на мотонейроны спинного мозга,
иннервирующие диафрагму для ее сокращения, тем самым
создавая двигательный ритм дыхания
Ритмогенез в пре-БК может варьироваться в зависимости от
метаболических, поведенческих и экологических условий
организма. Кроме пейсмекерных нейронов пре-БК содержит
большое количество проприобульбарных нейронов,
имеются бульбоспинальные и черепные мотонейроны
Пре-Бётцингера
комплекс (пре-БК)
16. Ростральная часть ВРГ (рВРГ)
Ростральная часть ВРГ содержитбольшое количество
диафрагмальных премоторных
бульбоспинальных нейронов,
которые напрямую
(моносинаптически) усиливают
возбуждение мотонейронов,
иннервирующих диафрагму
В качестве моносинаптического
нейротрансмиттера в этой
передаче возбуждения
используется глутамат
Доказано, что нейроны
рВРГ не влияют на
дыхательный ритм
Группа нейронов рВРГ подавляется
Бётц.К. во время выдоха и
возбуждается пре-БК во время вдоха
17. Каудальная часть ВРГ (кВРГ)
кВРГ в основном содержит экспираторныепремоторные нейроны. Их аксоны, образуя
синапсы на двигательных нейронах грудного и
поясничного отделов спинного мозга,
иннервируют дополнительные мышцы выдоха
(брюшные и внутренние межреберные.
Модуляция нейронов кВРГ может происходить из
возбуждающих входов от РТЯ/пФРГ, и,
возможно, из комплекса Бётцингера
По общему мнению, каудальные
нейроны ВРГ не участвуют в
респираторном ритмогенезе во
время нормального дыхания
В то же время кВРГ (как и комплекс
Бётцингера) участвует в важных рефлексах с
верхних дыхательных путей, которые требуют
модуляции фазы выдоха, например, удлинение
выдоха при кашле и рвоте
Каудальные нейроны ВРГ, получая нисходящие
сигналы от нейронов околоводопроводного серого
вещества в среднем мозге, участвуют в координации
дыхания при разговоре и пении преимущественно во
время активного выдоха
Интересно, что нейроны кВРГ управляют краткой
остановкой дыхания для улучшения
целенаправленной деятельности человека,
например, при прицеливании во время стрельбы
из лука.
Каудальная часть ВРГ (кВРГ)
18. РТЯ/пФРГ— это две функционально не однородные и отдельные области, расположенные вентрально к ядру лицевого нерва (VII) и
лежащих рядомдруг с другом, с некоторым
анатомическим перекрытием с ним
РТЯ/пФРГ играет ключевую роль в
регуляции дыхания, особенно для
усиления экспираторной активности при
гиперкапнических состояниях
РTЯ являются основным сайтом
центральных хеморецепторов в стволе
мозга, модулируя входы от
каротидного тельца и обеспечивают
важный “драйв” вентиляции при
изменениях Ро2 и Рсо2 во вдыхаемом
воздухе
На нейронах РТЯ конвергирует множество
афферентных сигналов от нескольких центральных
хеморецепторных зон (высокочувствительных к CO2 и
pH), а также от периферических хеморецепторов
каротидного тельца (чувствительных
преимущественно к низкому уровню O2 и в меньшей
степени — к обнаружению CO2 и pH).
Эта поступающая в РТЯ информация рассматривается как
«побуждение» к активации вдоха
Совсем недавно было установлено, что хемочувствительные
нейроны РТЯ являются глутаматергическими
проприобульбарными интернейронами, которые избирательно
через нейроны ВРГ иннервируют гладкую мускулатуру
дыхательных путей и другие области продолговатого мозга,
участвующие в регуляции дыхания
Ретротрапециевидное
ядро / парафациальная
респираторная группа
(РТЯ/пФРГ) расположена в
ростральной и вентральной
частях ретикулярной
формации Medulla
19. Парафациальная респираторная группа (пФРГ) расположена ростральнее ВРГ Анатомически и функционально нейроны пФРГ отличаются от
Латеральное парафациальноеядро (лпФРГ), является
ритмогенным
лпФРГ - это условный
генератор, который управляет
активным выдохом и усиливает
вентиляцию во время высоких
метаболических и
дыхательных потребностей за
счет включения экспираторных
мышц брюшной полости. В
покое и под анестезией
нейроны лпФРГ «кажутся
молчаливыми», но когда они
расторможены и/или
активированы, они вызывают
активный выдох фазу Е2.
Кроме того, модуляция лпФРГ,
по-видимому, имеет решающее
значение для экспираторной
активности мышц брюшной
стенки во время быстрого сна
Вентральное парафациальное
ядро (впФРГ) не считается
ритмогенным
Парафациальная респираторная группа (пФРГ) расположена
ростральнее ВРГ
Анатомически и функционально нейроны пФРГ отличаются от
смежных и частично перекрывающихся с ними
хемочувствительных нейронов ретротрапециевидного ядра
(РТЯ)
Основываясь на функциональных различиях, нейроны,
расположенный рядом с лицевым ядром, разделили на две
отдельные популяции: латеральную парафациальную
респираторную группу –лпФРГ и вентральную
парафациальную респираторную группу впФРГ
20. Постинспираторный комплекс
(пост. К) – этонебольшая группа
холинергических нейронов
расположенная в области
смежной с N. ambiguus (NA)
пост. К обладает
автономными свойствами
генерации ритма и
необходим и достаточен
для постинспираторной
активности in vivo
Установлено, что пост.К
связан с двумя группами
нейронов, участвующих в
процессе дыхания, то есть
с комплексом преБетцингера (preBötC, вдох)
и лпФРГ, активный выдох),
вместе составляя тройной
дыхательный генератор
Постинспираторный
комплекс
21.
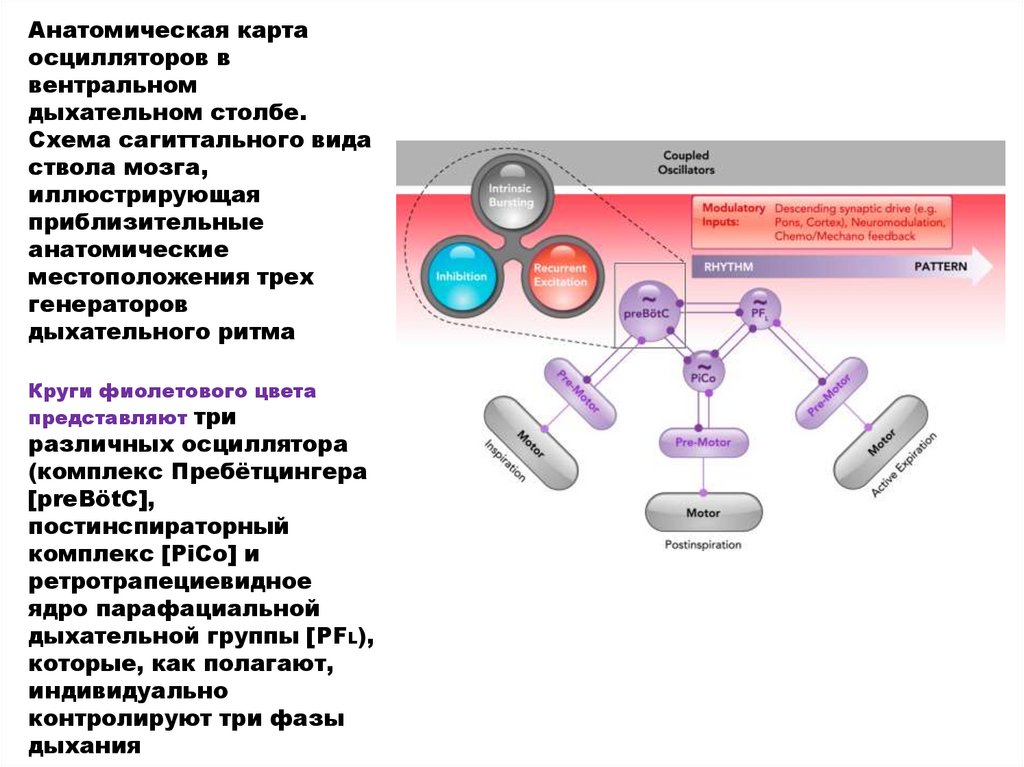
Анатомическая картаосцилляторов в
вентральном
дыхательном столбе.
Схема сагиттального вида
ствола мозга,
иллюстрирующая
приблизительные
анатомические
местоположения трех
генераторов
дыхательного ритма
Круги фиолетового цвета
представляют три
различных осциллятора
(комплекс Пребётцингера
[preBötC],
постинспираторный
комплекс [PiCo] и
ретротрапециевидное
ядро парафациальной
дыхательной группы [PFL),
которые, как полагают,
индивидуально
контролируют три фазы
дыхания
22. Дорсальная респираторная группа ДРГ
Дорсальная респираторная группанейронов занимает каудальную треть ядра
одиночного тракта (ЯОТ и является местом
расположения релейных
(переключательных, второго порядка,
нейронов.
В ДРГ, поступающая от рецепторов
растяжения легких и периферических
хеморецепторов аортальных и каротидных
телец информация переключается на
респираторные нейроны пре-БК и другие
нейронные сети, модулирующие паттерн
дыхания в зависимости от состояния
метаболизма, деятельности человека, а
также состояния окружающей среды.
Таким образом, нейронная сеть ДРГ не
участвует в ритмогенезе
ДРГ является главным фокусом, в
котором сосредотачивается важная
афферентная информация о степени
раздувания легких, просвете дыхательных
путей и уровне артериального Ро2 и Рсо2.
Эти сигналы приходят в ДРГ по
блуждающим и языкоглоточным нервам от
сенсорных нейронов
Дорсальная
респираторная группа
ДРГ
23. Механизм чередования дыхательных фаз или координация работы респираторных нейронных сетей
Чередование инспираций ипостинспираций при обычном
дыхании происходит только за счет
взаимоингибирующих синаптических
связей между нейронами преБётцингерова комплекса и
нейронами постинспираторного
комплекса
Фаза активного выдоха отсутствует
В случае необходимости усилить
выдох – сделать его активным,
включаются нейроны
латеральной парафациальной
респираторной группы –лпФРГ и
кВРГ
Мышцы выдоха включаются в
дыхательный ритм только тогда,
когда требуется высокий уровень
вентиляции легких, например, во
время физических упражнений,
гипоксии или воздействия
аномально высоких уровней СO2
Механизм чередования
дыхательных фаз или
координация работы
респираторных нейронных
сетей
24. Вклад коры больших полушарий и других отделов мозга в регуляцию дыхания
Гипоталамусосуществляет связь с
работой внутренних
органов
Лимбическая система
изменяет дыхание во
время эмоций
Спинальный контроль.
Мотонейроны спинного
мозга на уровне CIII-CV
управляют диафрагмой, а
TI-TXII – наружными
межреберными мышцами
Во время бодрствования,
выполнения физических
упражнений, разговоре, пении
дыхание регулируется
непроизвольно и осознается
исключительно в качестве
двигательной функции
И в этом смысле дыхание
обучаемо (это используется в
лечебных и др. целях)
Самопроизвольный контроль
дыхания носит защитный характер
– вспомните игру в прятки! –
затаить дыхание
Человек использует
произвольное усиление дыхания гипервентиляцию при надобности
Условные рефлексы,
упреждающее усиление дыхания
(предстартовые состояния)
25. Современное представление о структуре ДЦ
26. Респираторные паттерны нормального и патологического дыхания
27. Химическая регуляция дыхания Периферические и центральные хеморецепторы
Периферические илиартериальные хеморецепторы
обладают высокой
чувствительностью к сдвигам
концентрации дыхательных газов
(преимущественно кислорода)
в артериальной крови
Периферические хеморецепторные
клетки расположены в каротидных
(glomera carotica) и аортальных
(glomera aortica) тельцах
артериального русла
системного круга
кровообращения
Glomus carotica локализованы
билатерально в месте бифуркации
общей сонной артерии на наружную
сонную артерию и внутреннюю
сонную артерию и иннервируются
ветвью языкоглоточного нерва
Glomus aortica расположены вокруг
дуги аорты и правой подключичной
артерии и иннервируются ветвью
блуждающего нерва.
Химическая регуляция
дыхания
Периферические и
центральные
хеморецепторы
28.
Чувствительность каротидных иаортальных телец к О2 настолько
высока, что они возбуждаются уже
при нормальном артериальном Ро2,
равном 95—100 мм рт. ст.
Это и понятно: данные хеморецепторы
расположены в непосредственной близости к
левому желудочку, из которого в системный
кровоток (в норме) выбрасывается высоко
оксигенированная кровь. Поэтому главной
функцией хеморецептивного каротидного и
аортального телец является немедленное
усиление вентиляции легких на снижение
оксигенации крови в системной циркуляции
Хемочувствительные клетки
каротидных и аортальных телец
являются рецепторами О2: они
буквально измеряют его парциальное
давление в артериальной крови и
поддерживают гомеостатическое
регулирование его уровня
посредством рефлекторного контроля
вентиляции. Клетки телец обладают
высокой чувствительностью к
недостатку О2, так что резко
увеличивают частоту афферентных
импульсов при снижении Ро2 в
пределах 30—60 мм рт. ст., то есть в
диапазоне, при котором насыщение
гемоглобина кислородом
стремительно падает
Каротидные тельца состоят из
иннервируемых скоплений
хеморецепторных клеток типа I и
синаптически связанных с ними глииподобных клеток типа II (не служат
хеморецепторами). Внутри каждого
каротидного тельца клетки окружены
большим количеством фенестрированных
кровеносных капилляров с очень
интенсивным кровотоком в них. Клетки
клубочка связаны друг с другом щелевыми
контактами и образуют пресинаптические и
постсинаптические связи с афферентными
и эфферентными симпатическими и
парасимпатическими нервными волокнами
29. Механизмы функции артериальных хеморецепторов, обеспечивающих изменения вентиляции легких в ответ на снижение Ро2в артериальной
кровиНейротрансмиттер - АТФ
30. Центральные хеморецепторы
Центральные хеморецепторы расположены менее чем в1 мм от вентральной поверхности продолговатого мозга
мозга в ретротрапециевидном ядре (РТЯ) внутри
гематоэнцефалического барьера и реагируют только на
изменения концентрации H+
Их реакция на CO2 является косвенной, происходящей
через изменения [H +] в спинномозговой жидкости
(ЦСЖ). Артериальная кровь и ликвор разделены
полупроницаемой мембраной, которая проницаема
дляCO2, но не для ионов H+. Повышение парциального
давления CO2 (PCO2) артериальной крови вызывает
диффузию молекул CO2 в ликвор, где они реагируют с
водой с образованием углекислоты (H2CO3). H + как
часть H2CO3 стимулирует рост активности медуллярных
хеморецепторов.
Гиперкапния вызывает быстрое и энергичное
увеличение минутной вентиляции
Молекулярные механизмы, лежащие в основе
чувствительности центральных хеморецепторов к
изменениям в PaCO2, остаются неясными. Считается,
что изменения РаCO2 обнаруживаются из - за
результирующих изменений рН в интерстициальной
жидкости.
Предполагается, что клетки хеморецепторов содержат
чувствительные к кислоте ионные каналы, которые
действуют как протонные датчики, чувствительные к
изменениям PaCO2 мозга или pH. Увеличение
артериального Pco2 приводят к увеличению частоты
дыхания и глубины дыхания до тех пор, пока через
несколько минут не будет достигнута стационарная
гипервентиляция
Хотя гипоксия угнетает метаболизм нервной
ткани и влияет на активность мозга, она
напрямую не стимулирует
центральную химическую сенсорную
систему, регулирующую вентиляцию
31. Механорецепторный контроль
Другие рецепторыМедленно адаптирующиеся рецепторы
растяжения лёгких находятся в ГМК ВП. Это
вагусные афференты, они раздражаются в
результате увеличения объёма лёгких и
связаны с ДРГ, т.е. тормозят вдох – рефлекс
Геринга-Брейера. При спокойном дыхании (ДО
< 1л) этот рефлекс не работает
Быстро адаптирующиеся рецепторы
расположены в эпителии и субэпителии
вплоть до альвеол, работают кратковременно,
вызывают кашель, чихание. В зависимости от
уровня раздражения ВП включаются
тройничный, блуждающий, языкоглоточный
нервы.
Лёгочные J- рецепторы расположены в
альвеолах в месте контакта их с капиллярами
и связаны с ДЦ тонкими
немиелинизированными С-волокнами,
реагируют на токсические вещества,
интерстициальный отёк.
Проприорецепторы (в том числе мышечные
веретена и γ- мотонейроны) в
периферических суставах и мышцах
усиливают вентиляцию при пассивной и
активной работе
Висцеральные и кожные
афференты (прикосновение к коже,
температурные, болевые
раздражения ) вызывают
гипервентиляцию.
Барорецепторы – при повышении
артериального давления урежается
дыхание
32.
ВздохиКашель
Чихание
Простые вентиляционные реакции
Вздохи в значительной степени зависят от
негативных (паника, тревога, и боль) и
положительных эмоций (удовольствие,
любовь и облегчение)
Дыхание модулируется различными
циркадными, когнитивными и
эмоциональные состояния
33. Зевота от скуки, гипоксии?
Вряд ли: зевает уже 20-недельный человеческий плод, трудно
предположить , что от скуки и сонливости
Зевота в среднем продолжается 5 с и состоит из непроизвольного
широкого открытия рта с максимальным расширением челюстей, а
также длительного и глубокого вдоха через рот и нос, с последующим
медленным выдохом, связанных с чувством комфорта.
Зевота очень заразительна.
Одна из наиболее убедительных версий состоит в том, что зевота помогает оттоку
перегретой крови от головы, то есть служит для терморегуляции. Новое исследование
подтверждает эту гипотезу.
Как обнаружили ученые, зевание вызвано именно повышением температуры внутри
черепа.
И цель этого процесса - как спонтанного, так и "заразительного" - поддерживать
оптимальную температуру мозга для его наиболее эффективного функционирования
Заразительность зевоты сильно меняется в разные времена года и прямо зависит от
температуры окружающего воздуха.
В северном полушарии, например, зимой мы значительно реже испытываем потребность
зевнуть, чем летом.
При этом наиболее заразительная зевота начинается примерно при 20 градусах Цельсия.
Дальнейшее повышение температуры уже почти не влияет на частоту и
продолжительность зевков.
34. Гипоксемия и гиперкапния
Гипоксемия подразумевает низкий Рo2 в крови - менее 60
мм рт.ст. Показатель дефицита растворенного кислорода в
артериальной крови. Симптомы - основным симптомом
гипоксемии является одышка, но в зависимости от того, как
быстро развивается гипоксемия, человек может испытывать
снижение способности к физическим нагрузкам, усталость и
спутанность сознания
Гиперкапния - это состояние, при котором слишком много
двуокиси углерода в крови (РCO2 более 47 мм рт.ст).
Гиперкапния, как правило, вызвана гиповентиляцией,
болезнями легких, а также может быть вызвана
воздействием сред, содержащих аномально высокие
концентрации двуокиси углерода.Клинические проявления:
вазодилятирующий эффект - расширение сосудов, багрово-синюшное лицо с
видимым расширением отдельных кожных сосудов, особенно на скулах, носу,
гиперемия сосудов коньюктивы, багровосинюшные, теплые конечности,
потливость. Влияние углекислого газа на нервную систему: эйфория,
агрессивность, тревога или наоборот безразличие, апатия очень характерный
симптом – сонливость
35. Дыхание в необычных условиях
Пониженное атмосферноедавление:
Проявляется с высоты около
3,5 км, появляются симптомы
горной болезни – результат
гипоксемии. Возникает
гипервентиляция (при Ро2 < 60
мм рт.ст.) – «вымывается
СО2» - остановка дыхания
Как вы думаете, почему
СО2 быстро
диффундирует из крови
человека при подъеме на
высоту в не герметичной
кабине летательного
аппарата?
Повышенное атмосферное давление
Столб воды высотой 10 м создает давление в 1
атм.
Повышение давления не вызывает изменений
вентиляции
После погружения на глубину важно медленно
возвращаться к нормальному давлению, т.к. азот
не может быстро удаляться через лёгкие.
Пузырьки азота могут закупорить сосуды – газовая
эмболия, развивается кессонная болезнь
Декомпрессия – медленный переход к
нормальному атмосферному давлению, например,
при погружении на 40 м время подъёма должно
составлять 40 мин
36. Физическая работа сопровождается усилением вентиляции
Вентиляция усиливаетсябуквально в первые 20 с
под влиянием двигательных зон
коры и проприорецепторов
сокращающихся мышц
Хеморецепторный контроль
связан с увеличением в плазме
молочной кислоты (ацидоз),
катехоламинов, повышении
температуры тела
Оксигенобаротерапия
Лечение кислородом под
повышенным давлением до 130140 мм рт.ст (в барокамерах)при
норме 100 мм рт.ст.
Увеличивает доставку кислорода к
тканям, физически растворенного
кислорода в плазме крови и
других жидкостях организма