



































































































 Биология
БиологияПохожие презентации:










Лекция 3. Фотосинтетические пигменты. Световая фаза фотосинтеза
1. ЦУР 2: «Ликвидация голода»
Фотосинтез – основапродуктивности
растений
2.
Лекция 3 Фотосинтетические пигменты. Световаяфаза фотосинтеза
Цель: Ознакомление со световой фазой фотосинтеза
1. Фотосинтетические пигменты.
2. Фотосистемы 1 и 2
3. Циклический и нециклический поток электронов.
фотосинтетическое фосфорилирование
4. . Хемиосмотическая теория П. Митчелла
(английский биохимик).
3.
ФОТОСИНТЕЗ1. Пигменты пластид.
2. Физико-хим. св-ва хлорофиллов.
3. Происхождение воды при фотосинтезе.
Фотосинтез
—
это
окислительновосстановительный
процесс,
в
котором
вода
окисляется до 02, а углекислый газ восстанавливается
до углеводов.
Свет
6С02 + 12Н20
----------С6Н1206 + 602+ 6Н20
Хлорофилл
Глюкоза
4.
Термины «окисление» и «восстановление»Окисление — это не только присоединение
кислорода, но и отнятие протонов, и потеря
электрона
Восстановление — это отнятие кислорода и
присоединение протонов или электронов.
5.
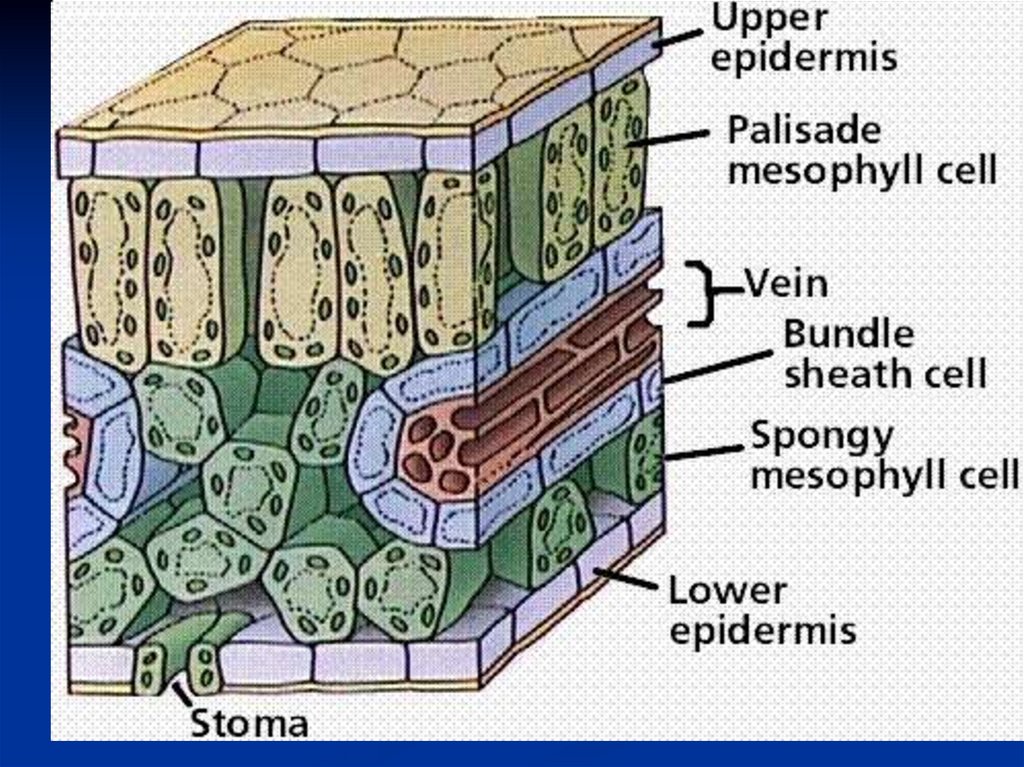
Анатомическое строение листа1. Лист с обеих сторон покрыт эпидермисом.
Клетки эпидермиса , за исключением
замыкающих клеток устьиц не содержат
хлоропластов.
Эпидермис защищает ткани листа,
регулирует газообмен и транспирацию.
2.
Основная ассимилционная ткань –
мезофилл.
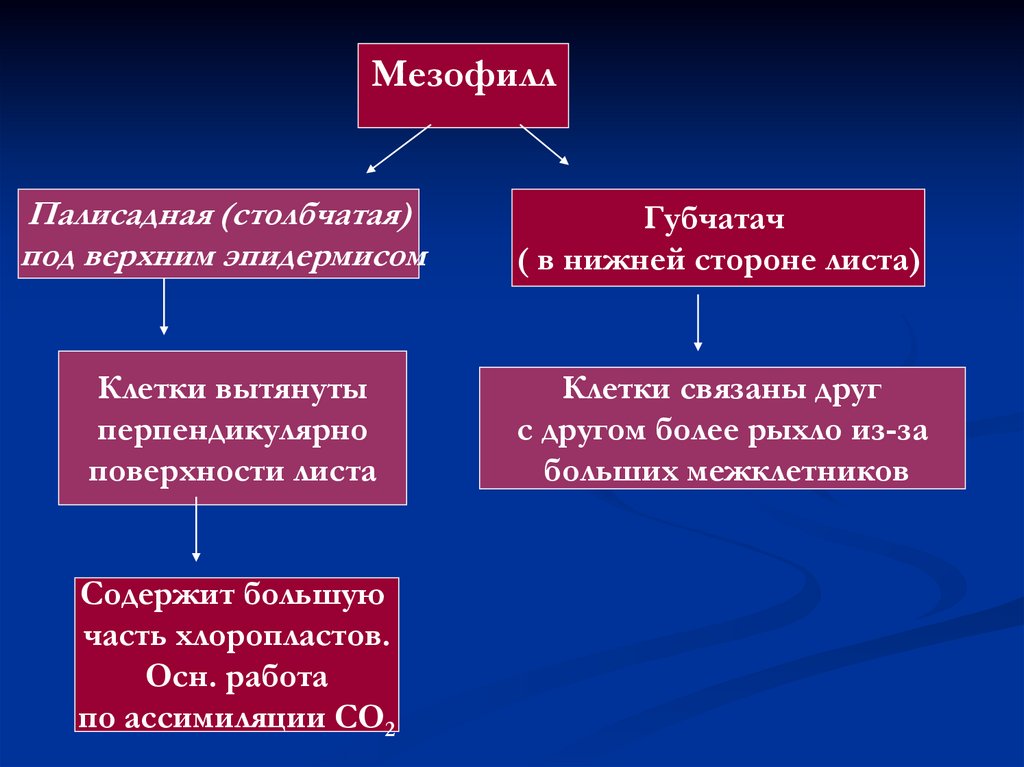
6.
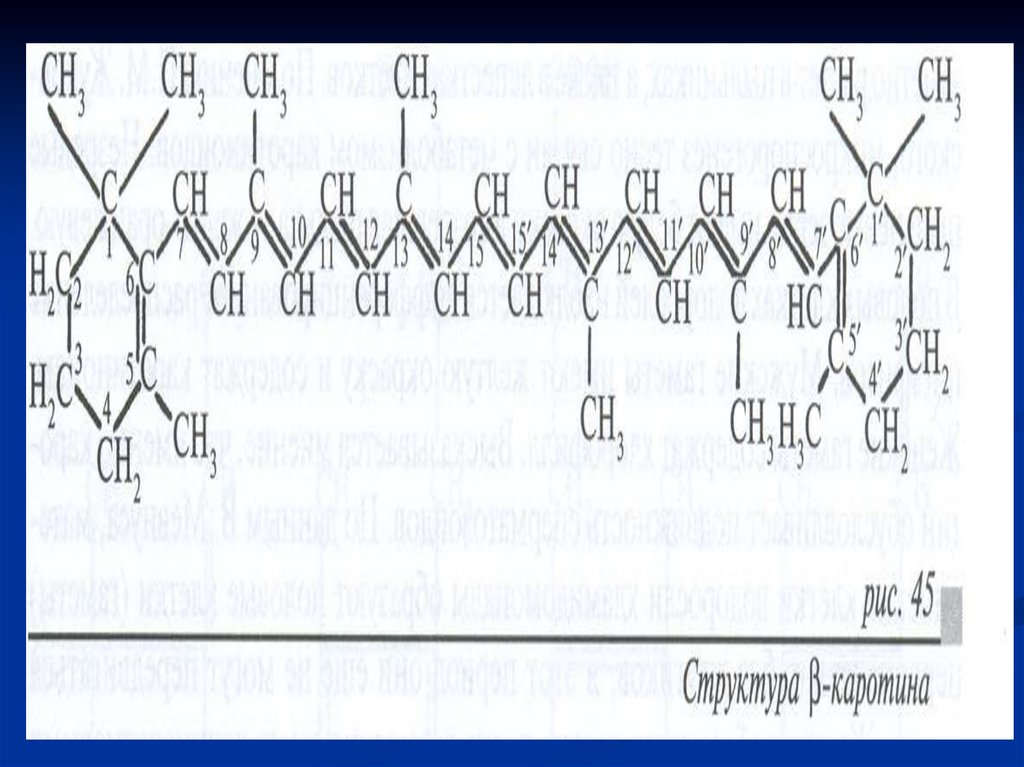
МезофиллПалисадная (столбчатая)
под верхним эпидермисом
Губчатач
( в нижней стороне листа)
Клетки вытянуты
перпендикулярно
поверхности листа
Клетки связаны друг
с другом более рыхло из-за
больших межклетников
Содержит большую
часть хлоропластов.
Осн. работа
по ассимиляции СО2
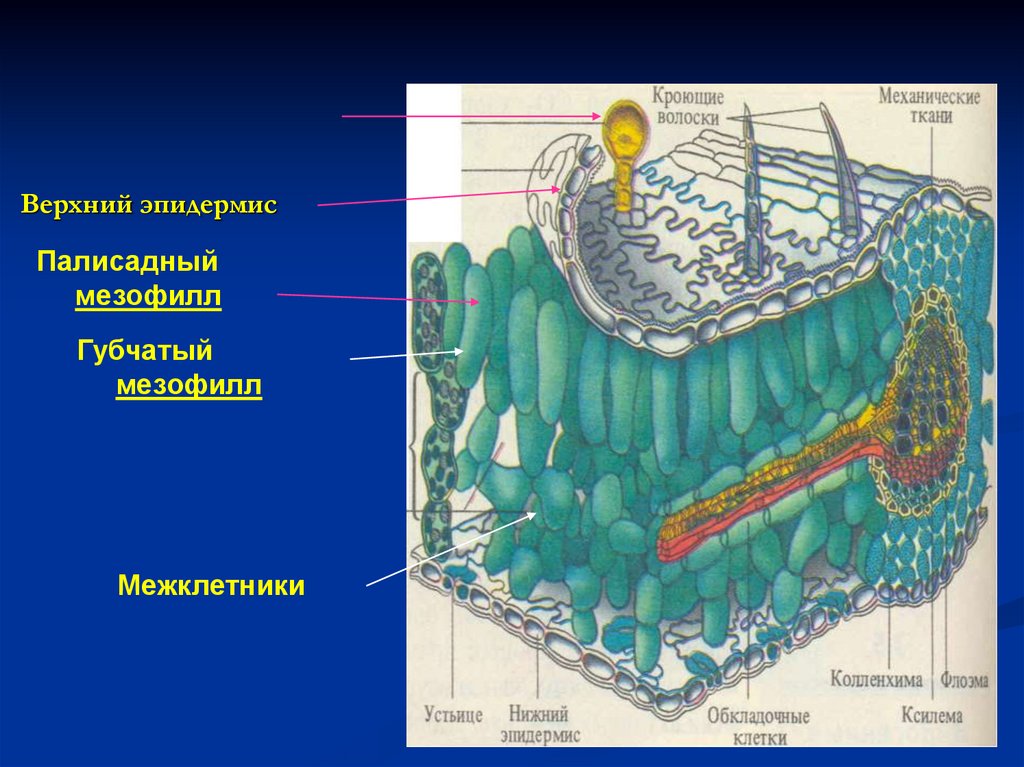
7.
Верхний эпидермисПалисадный
мезофилл
Губчатый
мезофилл
Межклетники
Железистый волосок
8.
Железистый волосок9.
10.
Пигменты хлоропластовРусский физиолог М.С. Цвет - 1901-1903 г.г.
Обнаружил 2 типа хлорофилла а и б , также желтые
пигмнты разделил на 3 фракции.
Пигменты пластид делятся на 3 класса:
хлорофиллы
фикобилины
каротиноиды.
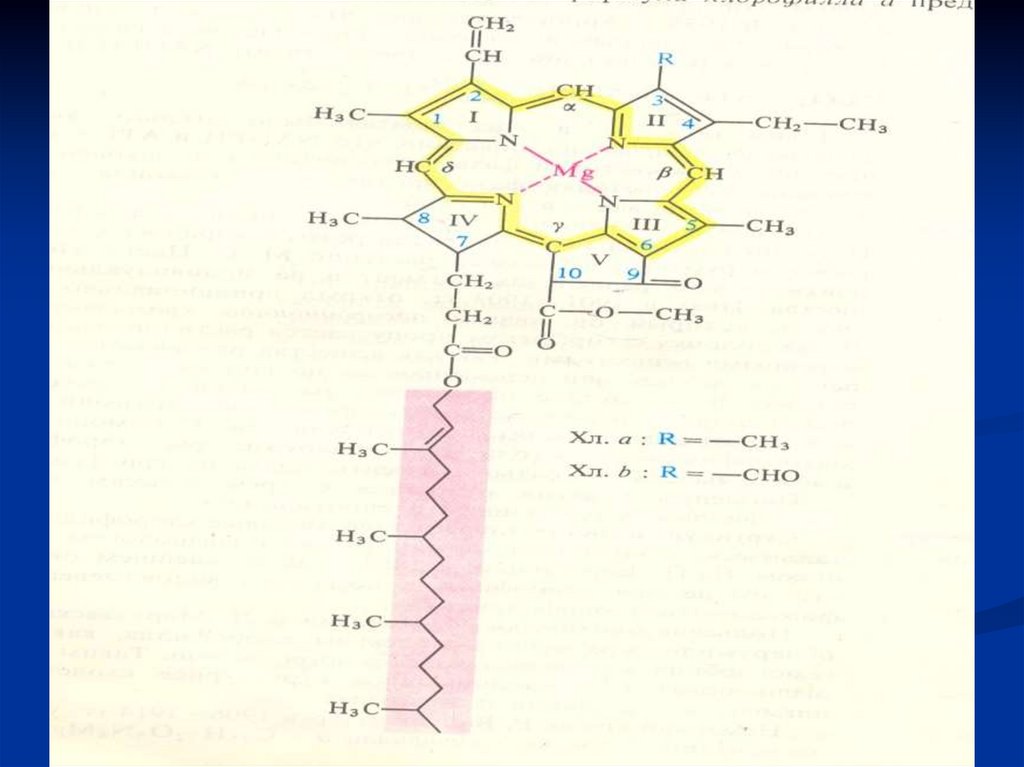
Строение хлорофиллов.
Хлорофиллы- осн. функция –поглощение солнечного света,
преобразование его в химическую энергию органических
соединений
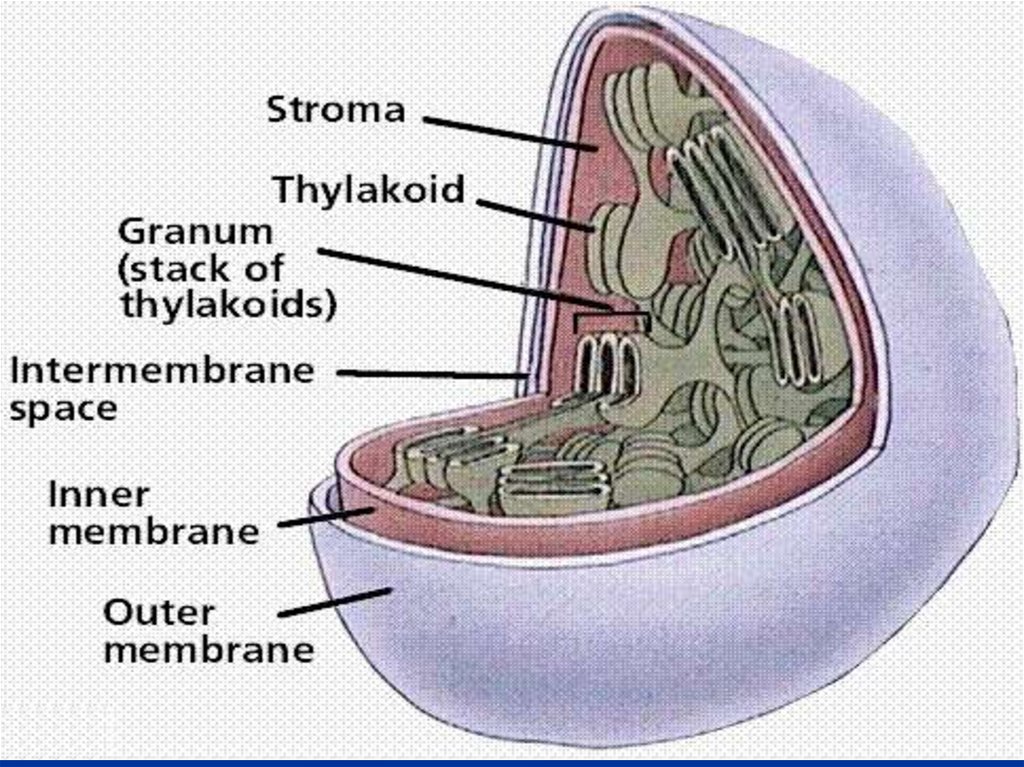
11.
Русский физиолог и ботаникИ. П. Бородин
- 1883 г.
Впервые выделил хлорофилл b
кристаллическом виде.
Немецкий
химик Р. Вильштеттер 19061914 г.г. – определил элементарный
состав хлорофилла.
хлорофилл а – C 55H7205N4Mg
хлорофилл б - C55H7006N4Mg
12.
1. Тетрапирролы:хлорофиллы — магний- порфирины;
фикобилины;
2. Полиизопреноидные пигменты каротиноиды.
Хлорофилл - сложный эфир дикарбоновой
кислоты хлорофиллина, одна карбоксильная
группа этерифицирована остатком
метилового спирта, а другая - остатком
одноатомного непредельного спирта фитола:
13.
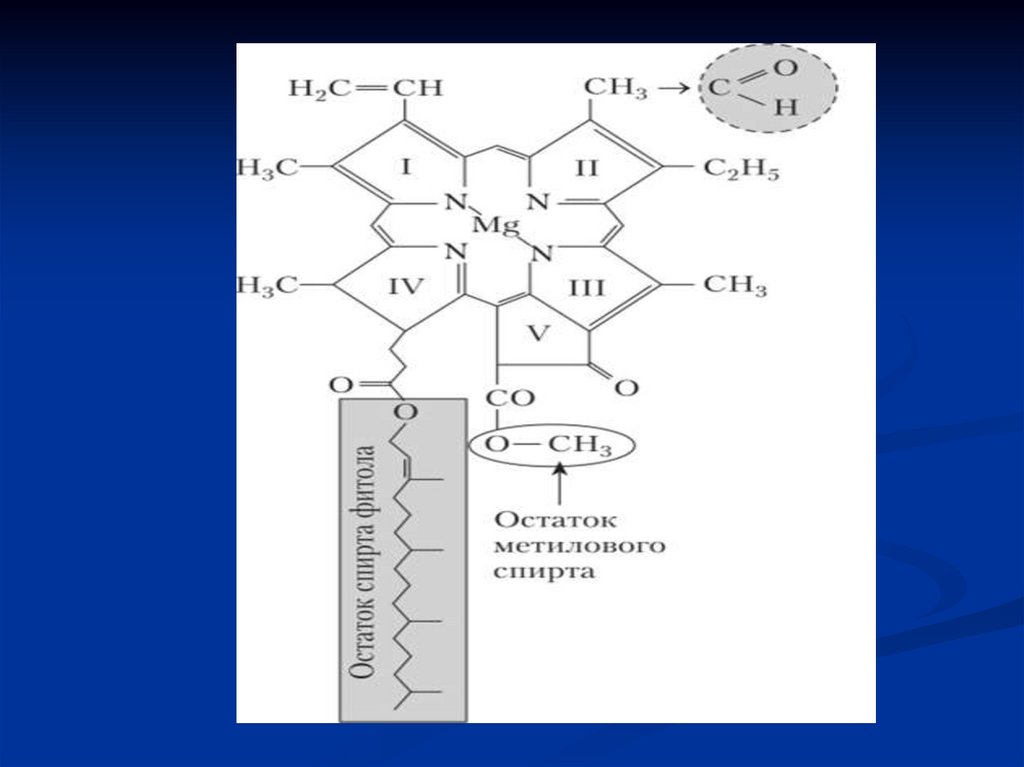
Хлорофилл b - отличается от хлорофилла а тем,что у 3-го углерода вместо метильной
находится формильная (-СНО) группа.
14.
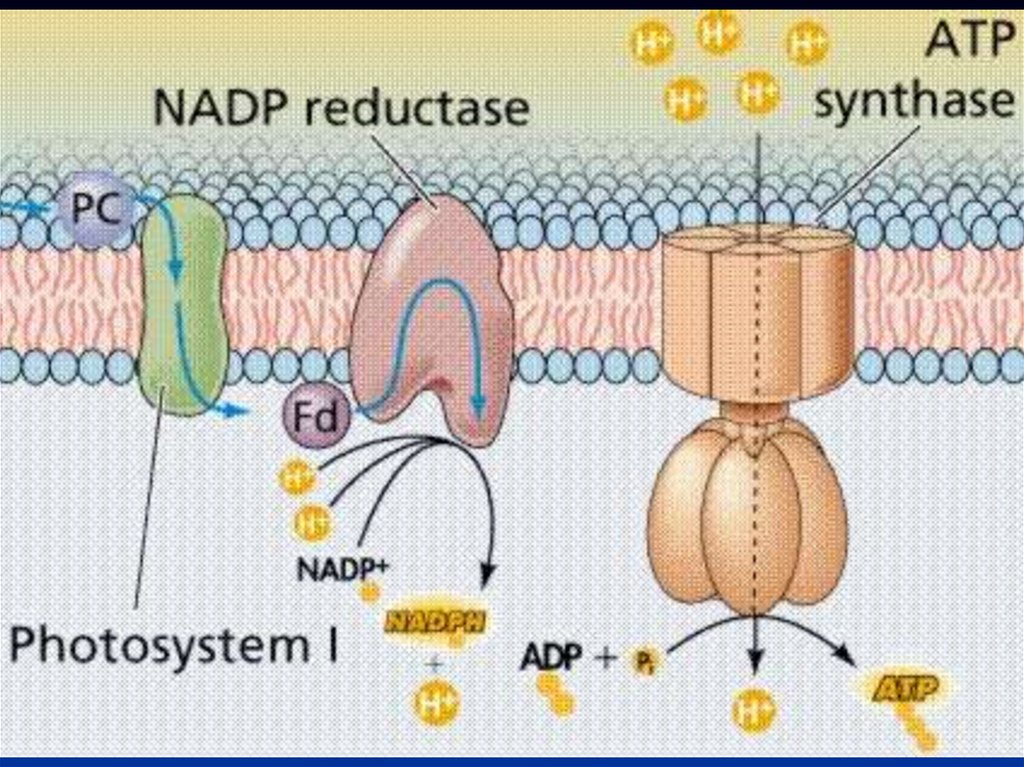
15.
Порфириновое кольцо - это плоская квадратнаяструктура, состоящая из четырех пиррольных
колец (1 - IV), каждое из которых содержит по
одному атому азота, способному
взаимодействовать с атомами металлов;
В хлорофиллах - это магний, в геме - железо.
К такой «голове» присоединен длинный
углеводородный «хвост» - сложноэфирная связь
образуется между спиртовой группой (-ОН) на
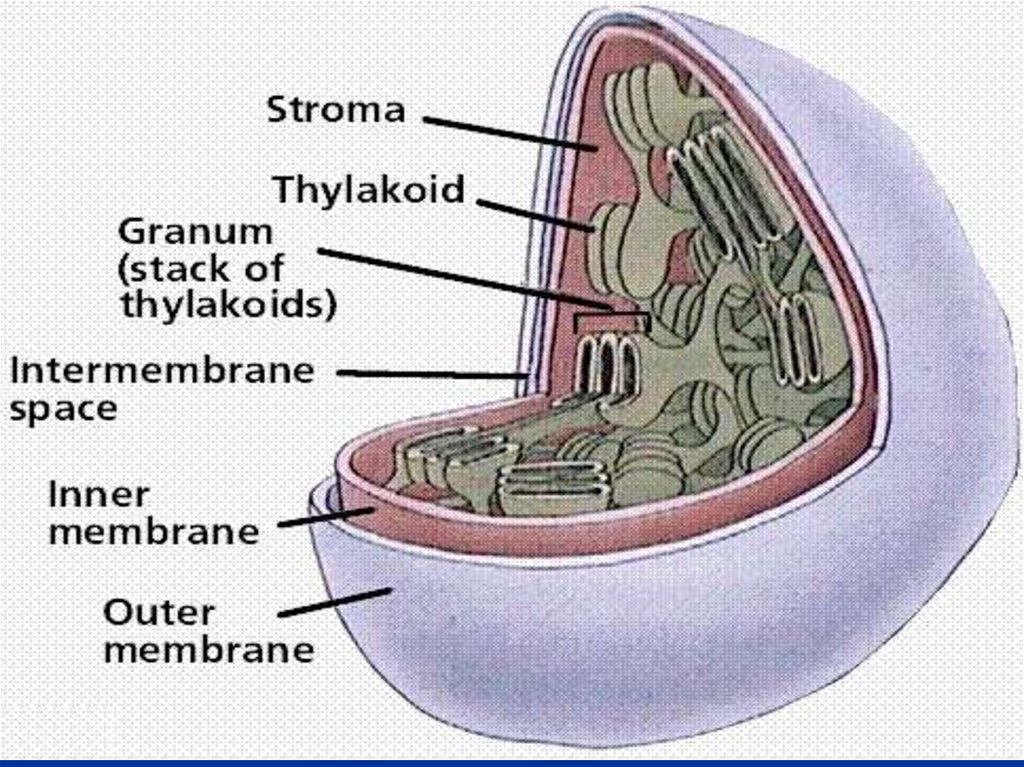
конце фитола и карбоксильной группой (-СООН)
на самой «голове»
16.
Связь структуры с функцией:а) длинный хвост растворим в липидах (т. е.
он гидрофобный) и таким образом
удерживает молекулу в мембране тилакоида;
б) голова гидрофильная (т. е. обладает
сродством к воде), и поэтому она обычно
лежит на той поверхности мембраны, которая
обращена к водной среде стромы;
17.
в) для лучшего поглощения света плоскость«головки» расположена параллельно
плоскости мембраны;
г) модификация боковых групп на
«головке» приводит к изменениям в
спектре поглощения, в результате чего
меняется и количество поглощаемой
энергии света;
18.
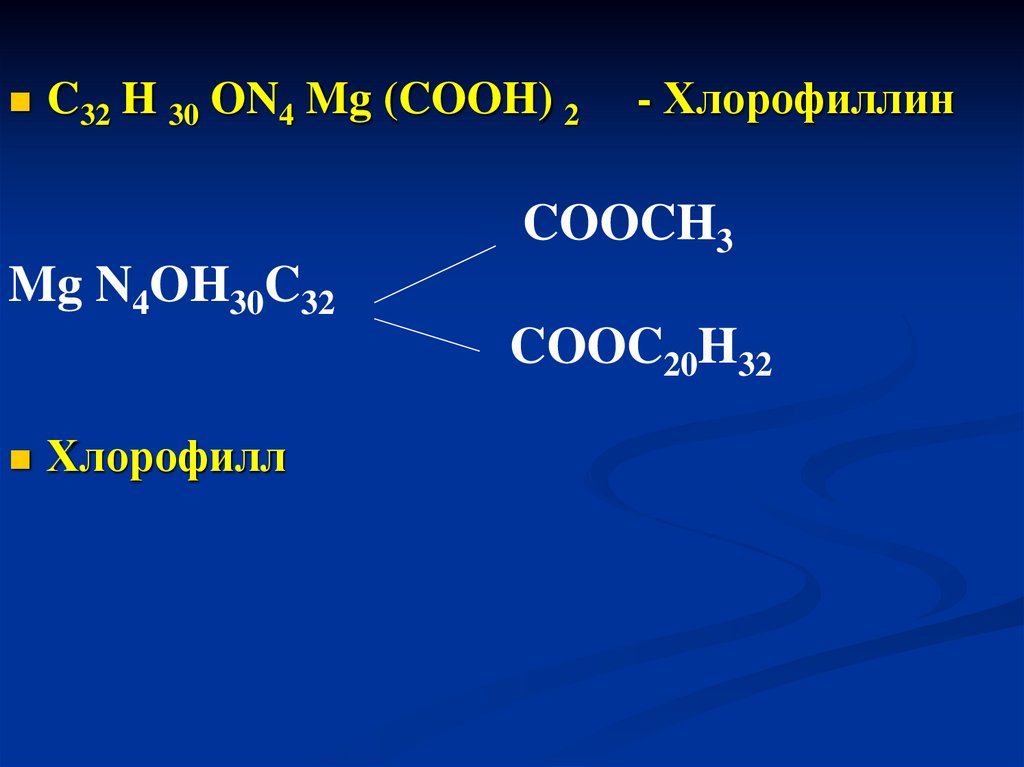
C32 H 30 ON4 Mg (COOH) 2- Хлорофиллин
COOCH3
Mg N4OH30C32
COOC20H32
Хлорофилл
19.
Форбин -тетрапиррол ициклопентановое
кольцо
Хлорофuллuд- структура хлорофилла,
лишенная фитола.
Феофитины образуются при замещении атома
магния протонами в молекуле хлорофиллов.
буро-зеленый цвет и ослабленный красный
максимум поглощения
20.
Хлорофиллы группы а:- фотосинтезирующие растения
- все группы водорослей,
- цианобактерии
Хлорофилл б:
- высшие растения
- зеленые водоросли и эвгленовые
Хлорофилл с – у бурых и диатомовых
водорослей
Хлорофилл d – у красных водорослей
21.
Хлорофилл а - фотосинтетический пигмент,представленный в наибольшем количестве; это
единственный пигмент, который имеется у всех
фотосинтезирующих растений и играет важную
роль в фотосинтезе.
Существует несколько форм этого пигмента,
которые различаются своим расположением в
мембране.
Каждая форма отличается от других и по
положению максимума поглощения в красной
области; например, этот максимум может быть
670, 680, 690 нм.
22.
В этиловомэфире максимумы
хлорофилла а:
В красной части спектра - 660-663 нм, в
синей - - 428-430 нм,
хлорофилл б
- 642 – 644, 452 - 455 нм.
У разных хлорофиллов разные боковые цепи, и это
несколько изменяет их спектры поглощения.
23.
Правило Стокса - максимумыфлуоресценции несколько сдвинуты в
более длинноволновую часть спектра по
отношению к максимумам поглощения.
Растворы хлорофиллов способны также
к фосфоресценции (т. е. длительному
послесвечению), максимум которого
лежит в инфракрасной области
24.
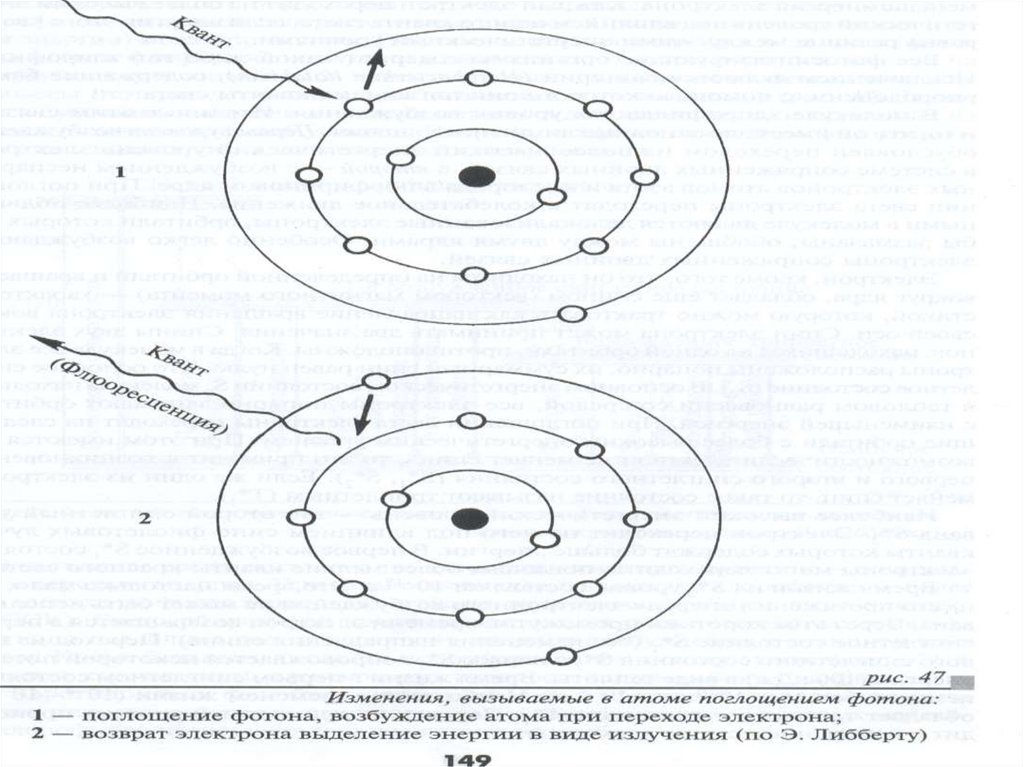
Физико-хим. св-ва хлорофилловСогласно законам фотохимии при поглощении
кванта света атомом или молекулой какоголибо вещества электрон переходит на другую,
более высокую орбиталь, т.е. на более высокий
энергетический уровень.
Наибольшей энергией обладает электрон,
отдаленный от ядра атома и находящийся на
большом расстоянии от него..
25.
Чем ближе к ядру, тем меньшеэнергии.
Каждый электрон переходит на более
высокий энергетический уровень под
влиянием одного кванта света, если энергия
этого кванта равна разнице между этими
энергетическими уровнями
26.
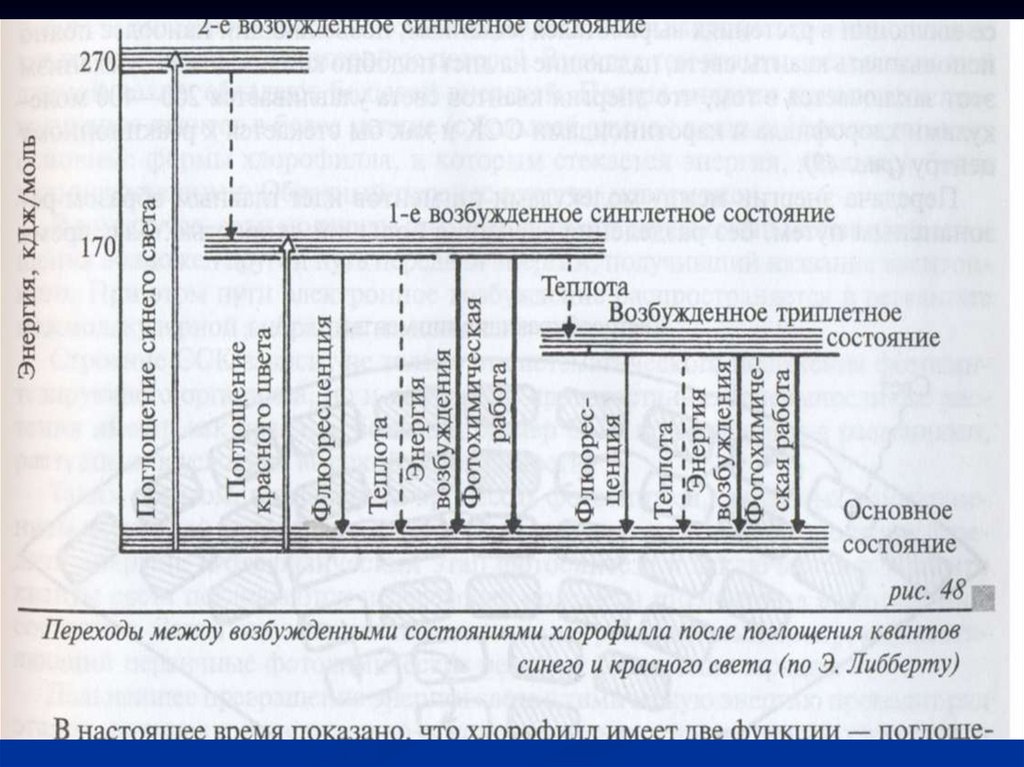
27.
В молекуле хлорофилла два уровнявозбуждения.
Именно с этим связано и то, что он имеет две
основные линии поглощения.
Первый уровень возбуждения обусловлен
переходом на более высокий энергетический
уровень электрона в системе сопряженных
двойных связей, а второй — с возбуждением
неспаренных электронов атомов азота и
кислорода в порфириновом ядре
28.
При поглощении света электроны переходят вколебательное движение.
Особенно легко возбуждаются электроны
сопряженных двойных связей.
Электрон находится на определенной орбитали и
вращается вокруг ядра, обладает спином
Спин - вектор магнитного момента, направление
вращения электрона вокруг своей оси.
Спины двух электронов, находящихся на одной
орбитали, противоположны.
29.
30.
Когда в молекуле все электронырасположены попарно, их суммарный спин
равен нулю.
Это основное синглетное состояние (S0).
В основном энергетическом состоянии S0
молекула находится в тепловом равновесии
со средой, все электроны попарно занимают
орбитали с наименьшей энергией.
При поглощении света электроны переходят
на следующие орбитали с более высоким
энергетическим уровнем.
31.
При этом имеются две возможности:если электрон не меняет спина, то это
приводит к возникновению первого и
второго синглетного состояния S*1, S*2.
Если же один из электронов меняет
спин, то такое состояние называют
триплетным (Т*).
32.
33.
Наиболее высокий энергетический уровень— это второй синглетный уровень S*2.
Электрон переходит на него под влиянием
сине-фиолетовых лучей, кванты которых
содержат больше энергии.
В первое возбужденное S*1 состояние
электроны могут переходить, поглощая более
мелкие кванты красного света.
Время жизни на S*2 уровне составляет 10-12
с. Это время настолько мало, что на его
протяжении энергия электронного
возбуждения не может быть использована.
34.
Через этот короткий промежуток времениэлектрон возвращается в первое синглетное
состояние S*1 (без изменения направления
спина).
Переход из второго синглетного состояния
S*2 →S*1 сопровождается потерей энергии
(100 кДж) в виде теплоты.
Время жизни в первом синглетном состоянии
немного больше - 10-9 или 10-8 с.
35.
Наибольшим временем жизни - 10-4 - 10-2с обладает триплетное состояние Т*1.
Переход на триплетный уровень
происходит с изменением спина
электрона.
Из возбужденного, первого синглетного и
триплетного состояния молекула хлорофилла
также может переходить в основное.
36.
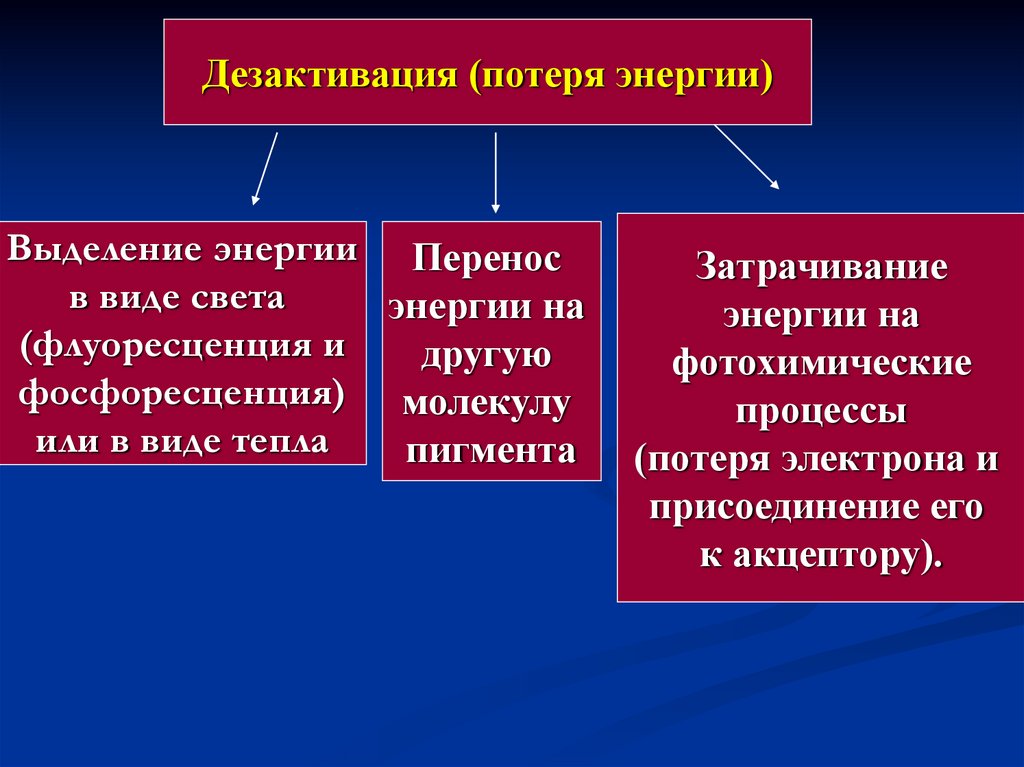
Дезактивация (потеря энергии)Выделение энергии Перенос
в виде света
энергии на
(флуоресценция и
другую
фосфоресценция) молекулу
или в виде тепла
пигмента
Затрачивание
энергии на
фотохимические
процессы
(потеря электрона и
присоединение его
к акцептору).
37.
В любом из указанных случаев молекулапигмента дезактивируется и переходит на
основной энергетический уровень.
Энергия, испускаемая в виде флуоресценции
или в виде теплоты, не может быть
использована.
В настоящее время показано, что хлорофилл
имеет две функции: поглощение и передача
энергии.
38.
Хлорофилл не существует в свободномвиде в клетке, так как он гидрофобен и
нестабилен.
Он связан с особыми белками, образуя
белок-пигментные комплексы.
Эта связь необходима по нескольким
причинам:
Стабилизация: белок удерживает
хлорофилл
в
определённой
ориентации,
защищает
его
от
разрушения.
39.
Фиксация в мембране: через взаимодействиебелка и фитольного хвоста хлорофилл
встроен в тилакоидную мембрану.
Оптимальное
расположение:
белок
обеспечивает
нужное
расположение
хлорофиллов в фотосистемах (I и II) для
эффективной передачи энергии.
Передача
энергии: белки участвуют в
энергетическом
взаимодействии
между
молекулами
хлорофилла
и
другими
пигментами (каротиноиды, фикобилины), а
также передают возбуждение к реакционному
центру фотосистемы.
40.
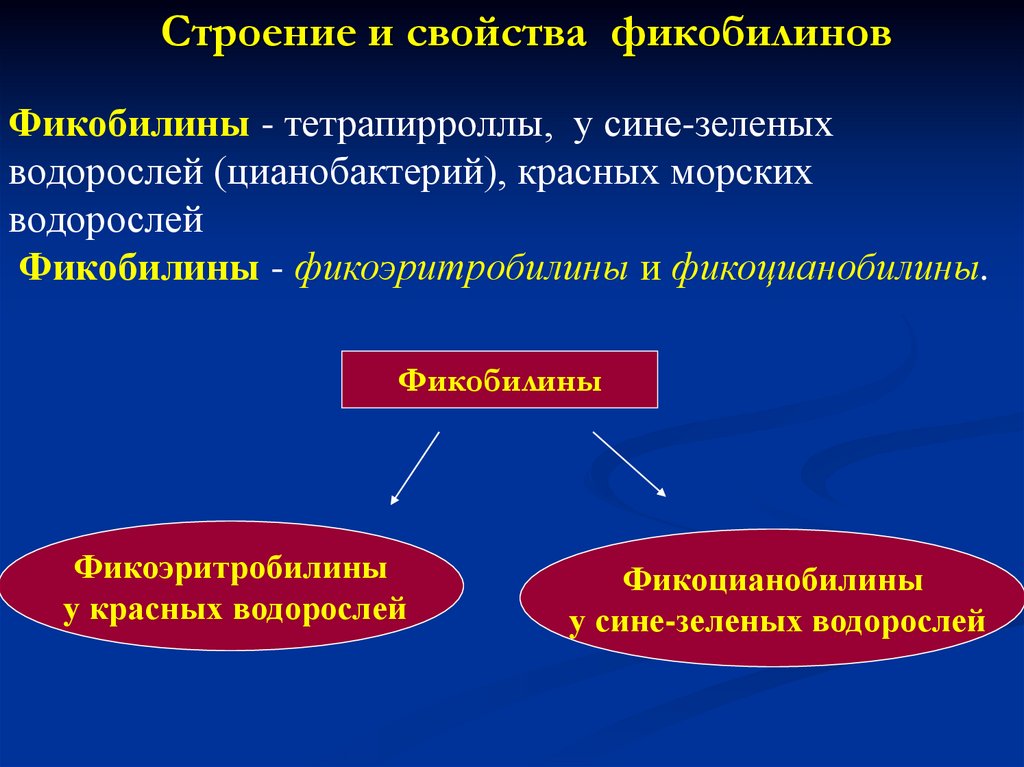
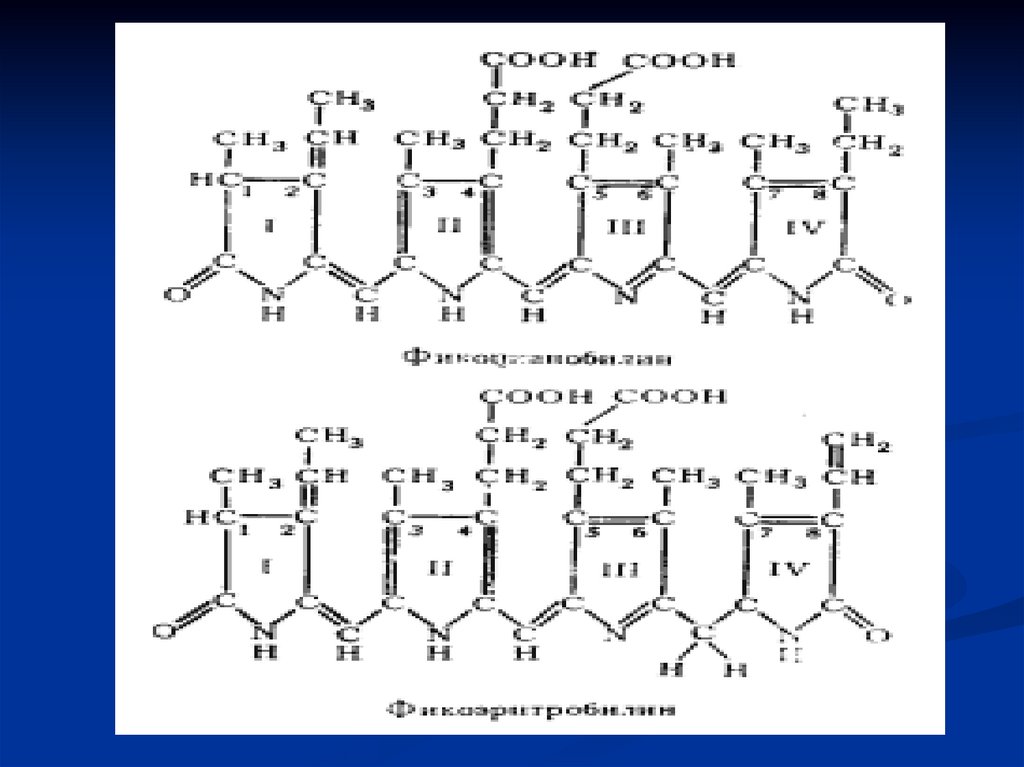
Строение и свойства фикобилиновФикобилины - тетрапирроллы, у сине-зеленых
водорослей (цианобактерий), красных морских
водорослей
Фикобилины - фuкоэрuтробилины и фuкоцuанобилины.
Фикобилины
Фикоэритробилины
у красных водорослей
Фикоцианобилины
у сине-зеленых водорослей
41.
Красные водорослиRhodophyta
Цианобакте́рии
Cyanobacteria
42.
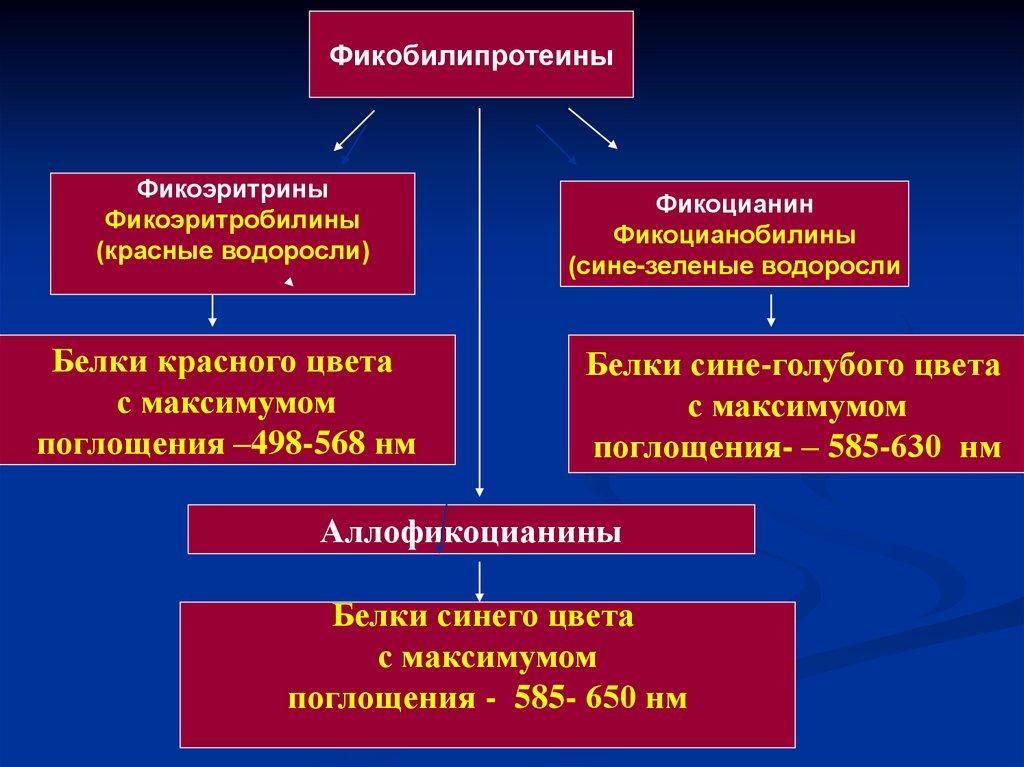
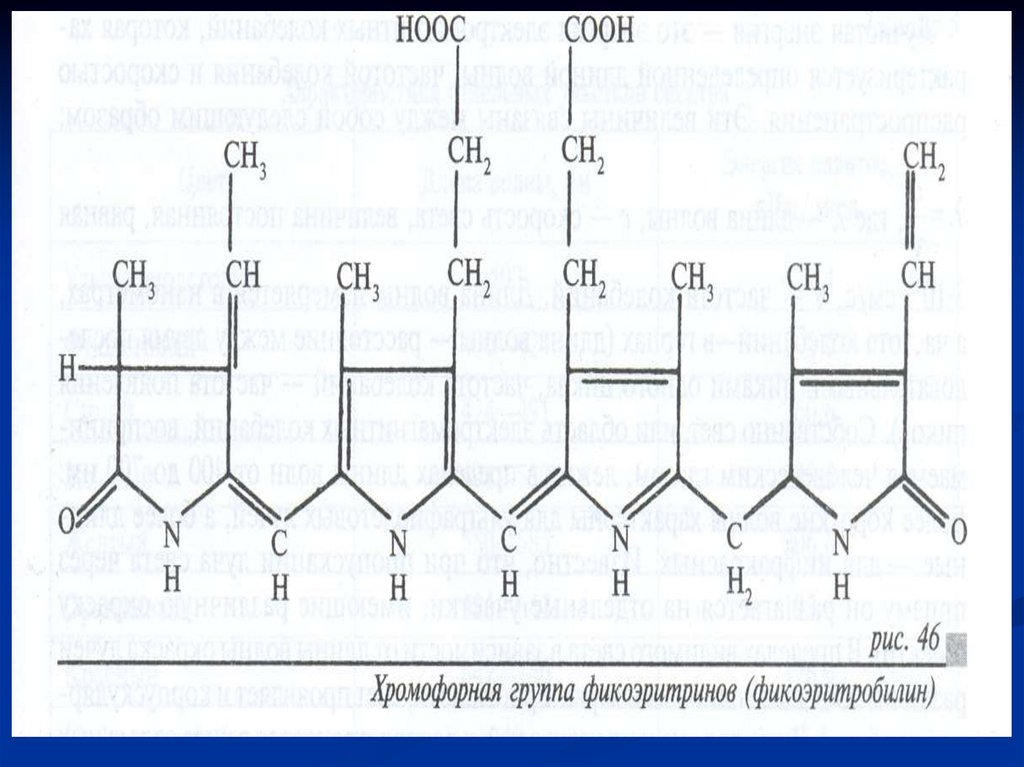
Фикобилины – хромофорные группыфикобилипротеинов.
Фикобилипротеины –белки, с которыми
фикобилины связаны прочными
ковалентными связями.
Фикобилипротеин = Фикобилин+ белок
43.
ФикобилипротеиныФикоэритрины
Фикоэритробилины
(красные водоросли)
Белки красного цвета
с максимумом
поглощения –498-568 нм
Фикоцианин
Фикоцианобилины
(сине-зеленые водоросли
Белки сине-голубого цвета
с максимумом
поглощения- – 585-630 нм
Аллофикоцианины
Белки синего цвета
с максимумом
поглощения - 585- 650 нм
44.
45.
46.
Максимумы поглощения света у фикобилиновнаходятся находятся между двумя максимумами
поглощения у хл-ла: в оранжевой, желтой и зеленой
частях спектра.
Это связано с оптическими свойствами воды.
Вода поглощает прежде всего длинноволновые лучи.
На глубине 34 м в морях исчезают полностью
красные лучи;
на глубине 177 м – желтые,
на глубине 322 м - зеленые,
на глубине свыше 500 м - синие и фиолетовые.
47.
В связи с изменением качественного состава света вверхних слоях морей обитают зеленые водоросли,
глубже – синезеленые водоросли
еще глубже – красные водоросли.
В. Т. Энгельман назвал это явление хроматической
комплементарной адаптацией водорослей.
У водорослей
фикобилины являются являются
дополнительными пигментами, выполняют вместо
хл-ла б роль светособирающего комплекса.
48.
КаротиноидыРастворимые в липидах пигменты, желтого,
красного цвета.
Встречаются в
хлоропластах, хромопластах –
моркови, латинское название -Daucus carota
L.
Из-за подавления
хлорофиллом, в листьях не
видно каротиноидов. Видны только осенью,
когда разрушается хлорофилл.
49.
Химическая природа -полиизопреноиды,
производные изопрена
Изопреноиды (СН2=С(СН3)—СН=СН2),
в основном содержат 40 атомов углерода.
50.
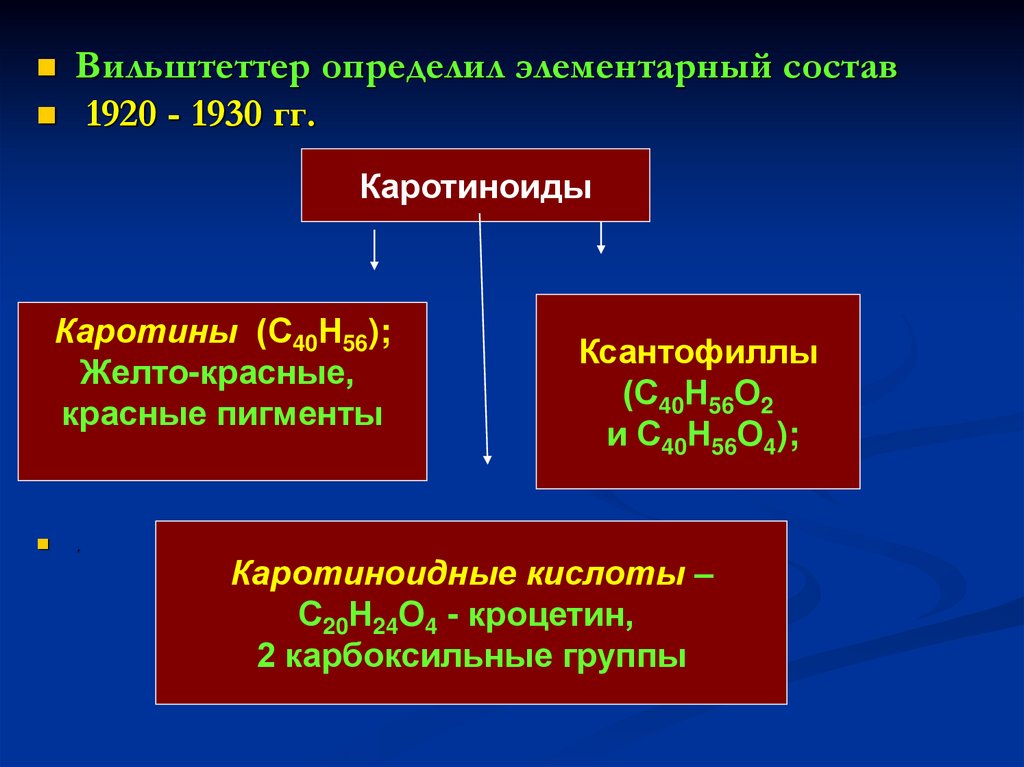
Вильштеттер определил элементарный состав1920 - 1930 гг.
Каротиноиды
Каротины (С40Н56);
Желто-красные,
красные пигменты
.
Ксантофиллы
(С40Н56О2
и С40Н56О4);
Каротиноидные кислоты –
С20Н24О4 - кроцетин,
2 карбоксильные группы
51.
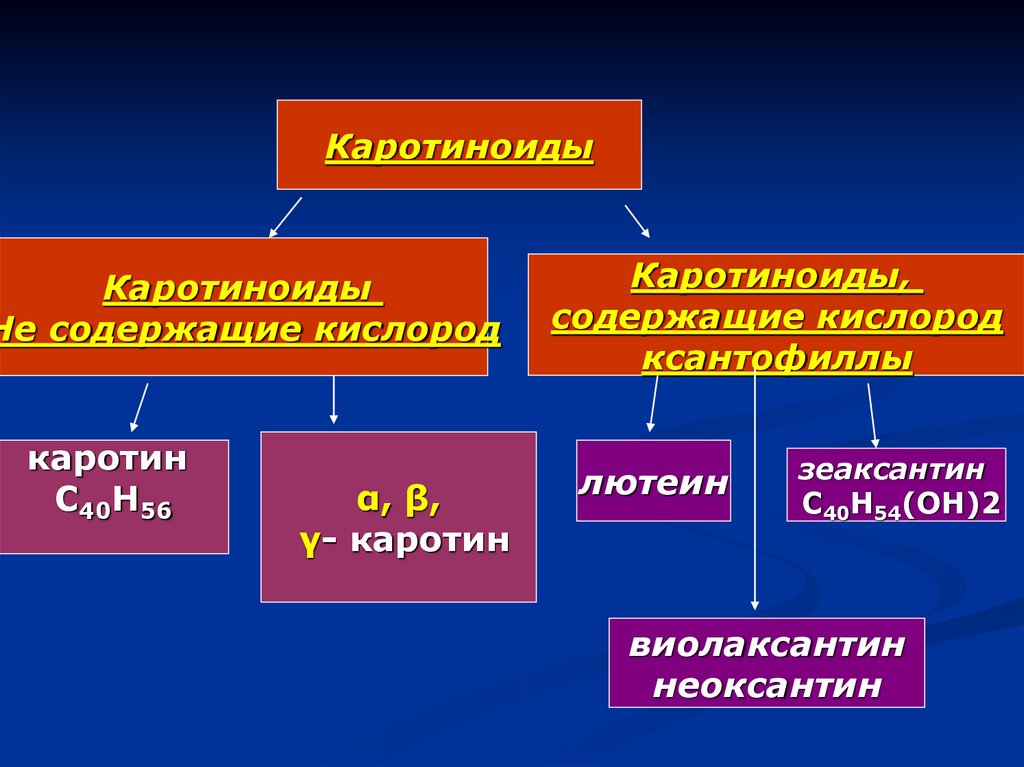
КаротиноидыКаротиноиды
Не содержащие кислород
каротин
С40Н56
α, β,
γ- каротин
Каротиноиды,
содержащие кислород
ксантофиллы
лютеин
зеаксантин
С40Н54(ОН)2
виолаксантин
неоксантин
52.
53.
Каротиноиды:- ациклические (алифатическими),
- моно- и бициклическими.
Циклы каротиноидов производные ионона.
Ациклический каротиноид
(С40Н56)
- ликопин
«Виолаксантин» произошло от названия
цветка анютины глазки (Viоlа tricolor)
Лютеин (от лат. luteus - желтый)-
желтке.
в яичном
54.
Фукоксанmин (С40Н60О6) – ксантофилл бурыхводрослей.
У высших растений и водрослей - β -каротин,
лютеин, виолаксантин и неоксантин.
55.
Синтез каротиноидовАцетил-СоА
Мевалоновая кислота
Геранилгеранилпирофосфат
Ликопин
Другие
каротиноиды
Синтезируется в
темноте, но при свете
усиливается синтез.
56.
Максимум поглощения в фиолетовой и синейчасти спектра - 400-500 нм.
Каротиноиды с белками
Роль
нековалентно связаны
каротиноидов в фотосинтезе.
Каротиноиды-участвуют в фотосинтезе
Функции:
1) Дополнительные пигменты
2) Предохраняет хлорофилл от фотоокисления.
57.
Д. И. Ивановский (1913) открыл защитнуюфункцию
Каротиноиды взаимодействуют с
хлорофиллом в триплетном состоянии,
спасает его от фотоокисления.
Хл Т* + Каротиноид -→ Хл Sо + Каротиноид
Т*
Каротиноид Т*-→ + Каротиноид Sо + теплота
Каротиноиды также могут взаимодействовать
с хлорофиллом в возбужденном синглетном
состоянии, переводит в основное состояние.
58.
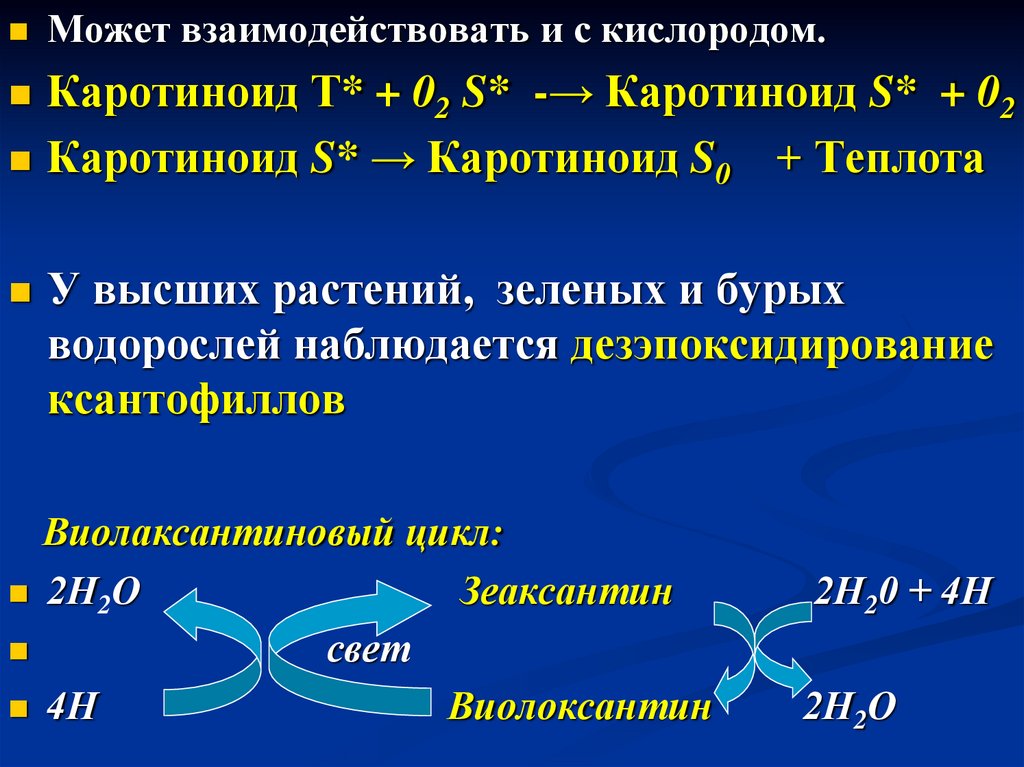
Может взаимодействовать и с кислородом.Каротиноид Т* + 02 S*
-→ Каротиноид S* + 02
Каротиноид S* → Каротиноид S0 + Теплота
У высших растений,
зеленых и бурых
водорослей наблюдается дезэпоксидирование
ксантофиллов
Виолаксантиновый цикл:
2Н2О
Зеаксантин
свет
4Н
Виолоксантин
2Н20 + 4Н
2Н2О
59.
60.
61.
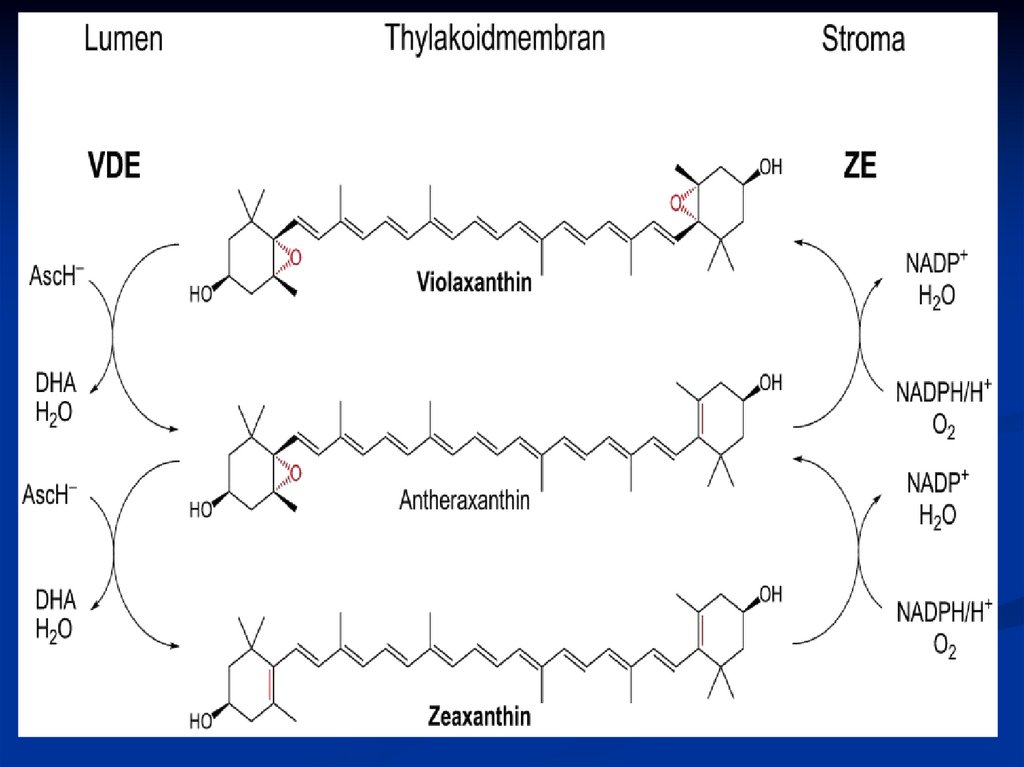
Виолоксантиновый цикл протекает в минорныхсубъединицах светособирающего комплекса
фотосистемы II .
При высокой интенсивности света вследствие
активной работы ЭТЦ фотосинтеза происходит
закисление люмена тилакоидов.
При падении pH до 5,0 активируются фермент
деэпоксидаза, которая с люменальной стороны
мембраны осуществляет восстанавление
эпоксидных групп виолоксантина, используя в
качестве восстановителя аскорбиновую
кислоту.
62.
Двукратноевосстановление приводит
образованию
зеаксантина,
который
выполняет фотопротекторную функцию.
При
к
и
снижении интенсивности освещения,
начинает преобладать обратная реакция
катализируемая эпоксидазой, расположенной
со стромальной стороны мембраны с pH
оптимумом 7,5.
63.
Для введения эпоксидных групп необходиммолекулярный кислород и восстановитель
(НАДФН).
В
результате образуется виолоксантин,
который
может
выполнять
роль
светособирающего пигмента
64.
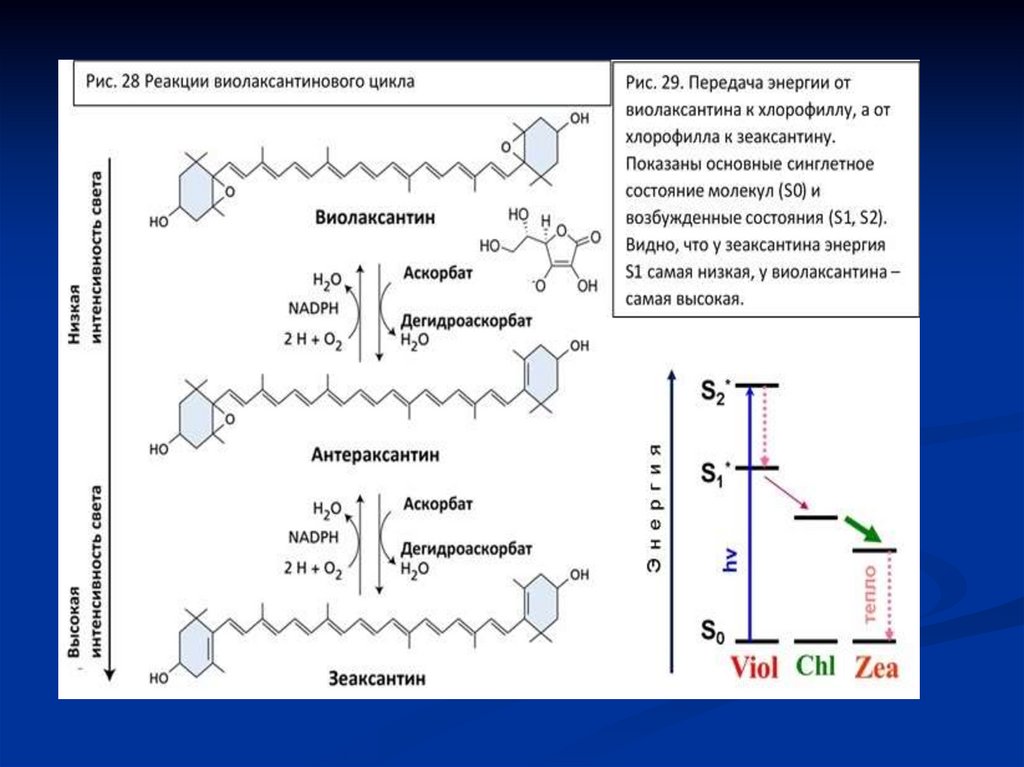
Суть виолоксантинового циклаЦикл включает обратимое превращение
трёх пигментов в тилакоидной мембране
хлоропластов:
При световом стрессе - интенсивное
освещение:
Виолаксантин (Vi)
(фермент: виолаксантин-депоксидаза)
Ананаксантин (An)↓
Зеаксантин (Ze)
65.
Зеаксантин— это активная форма,
которая помогает рассеивать лишнюю
энергию света в виде тепла (процесс
называется нефотохимическое тушение
—NPQ).
При уменьшении освещения:
Зеаксантин (фермент: зеаксантин-эпоксидаза
)
Ананаксантин
Виолаксантин
66.
Биологический смыслЗащита фотосинтетического аппарата от
повреждения высокоэнергетичными
фотонами.
Стабилизация работы фотосистем (особенно
ФСII) в условиях переменной освещённости.
Снижение образования опасных активных
форм кислорода
Быстрая адаптация к изменениям освещения.
67.
1. Химическая природа 3. Химическая природа фикобилиновхлорофилла
1. Изопрены
1. Сложный эфир
2. Тетрапирроллы
2. фосфолипид
3. Производное ионона
3. Феофетин
4. Производные ликопина
4. Дикарбоновая кислота 5. Хромопротеины
5. Полиизопрен
4. Тилакоид....- это
1. Внешняя мембрана хлоропластов
2. Химическая природа 2. . Внутренняя мембрана хлоропластов
каротиноидов
3. Инвагтинации внутренней мембраны
1. Полиеновое соединение4 Матрикс митохондрий
2. Витамин
5. Строма хлоропласт ов
3. Липопротеид
5. Что входит в состав хлорофилла? Выберите
4. Алифатическое
отрицательный ответ
Соединение
1. Фитол
5. Терпеноид
2.Гемпорфирин
3. Кольца порфирина
4.Тетрапиррол
5.Циклопентановое кольцо
68.
69.
70.
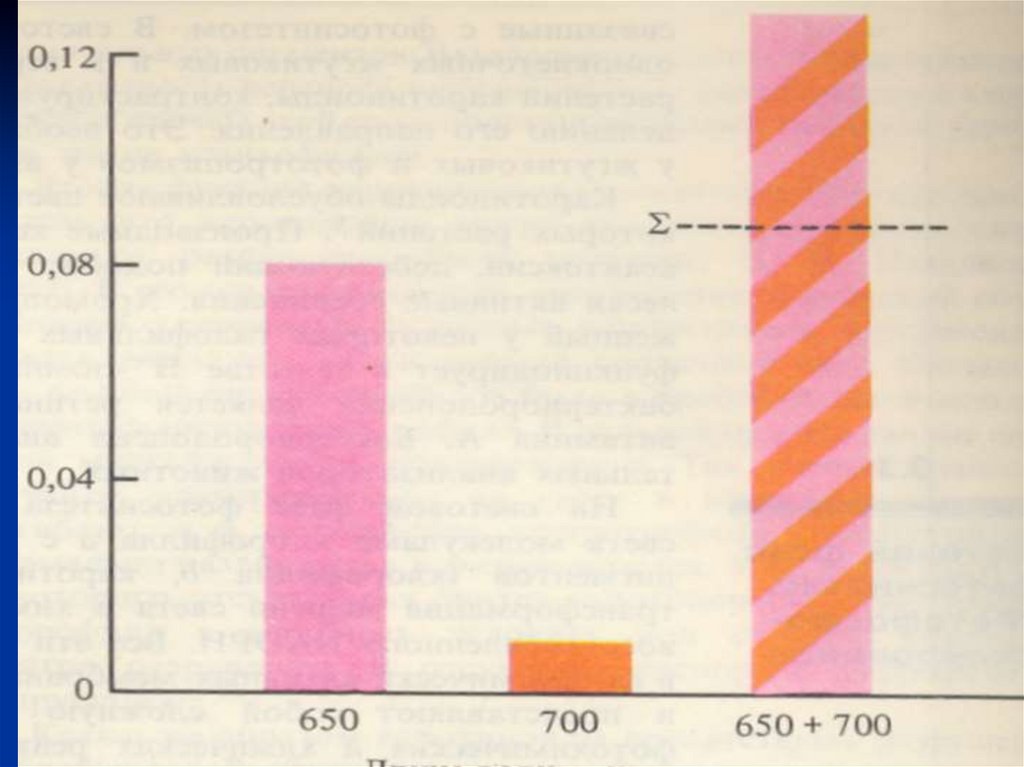
Эффект Эмерсона.Р. Эмерсон - эффективность света с длиной волны
680—700 нм может быть значительно повышена
добавлением света с более короткой длиной волны
(650—660 им).
Интенсивность ф/с при освещении смешанным
светом (с двумя длинами ) оказалась выше суммы
интенсивностей ф/с, наблюдаемой при освещении
светом каждой длины волны в отдельности.
Это явление называют эффектом усиления Эмерсона.
Вывод: необходимо одновременное световое
возбуждение пигментов, различающихся по спектру
поглощения.
71.
72.
Эффект усиления наблюдается при изучении ф/с увысших растений и цианобактерий. У других
бактерий этот эффект отсутствует и, соответственно,
в процессе фотосинтеза участвует одна фотосистема.
Фотосистема
(ФС)- совокупность
светособирающего комплекса (ССК),
фотохимического реакционного центра и связанных
с ним молекул - переносчиков электрона.
ФС является важнейшим структурно-
функциональным звеном ф/с-го аппарата.
В процессе фотосинтеза принимают участие две
фотосистемы.
73.
1. Фотосистемы 1 и 2. Циклический инециклический поток электронов.
фотосинтетическое фосфорилирование
Фотохимические реакции фотосинтеза - это реакции,
в которых энергия света преобразуется в энергию
химических связей, и в первую очередь в энергию
фосфорных связей АТФ.
АТФ – энергетическая валюта клетки, обеспечивающая
течение всех процессов.
Одновременно под действием света происходит
разложение воды, образуется восстановленный НАДФ
и выделяется кислород.
74.
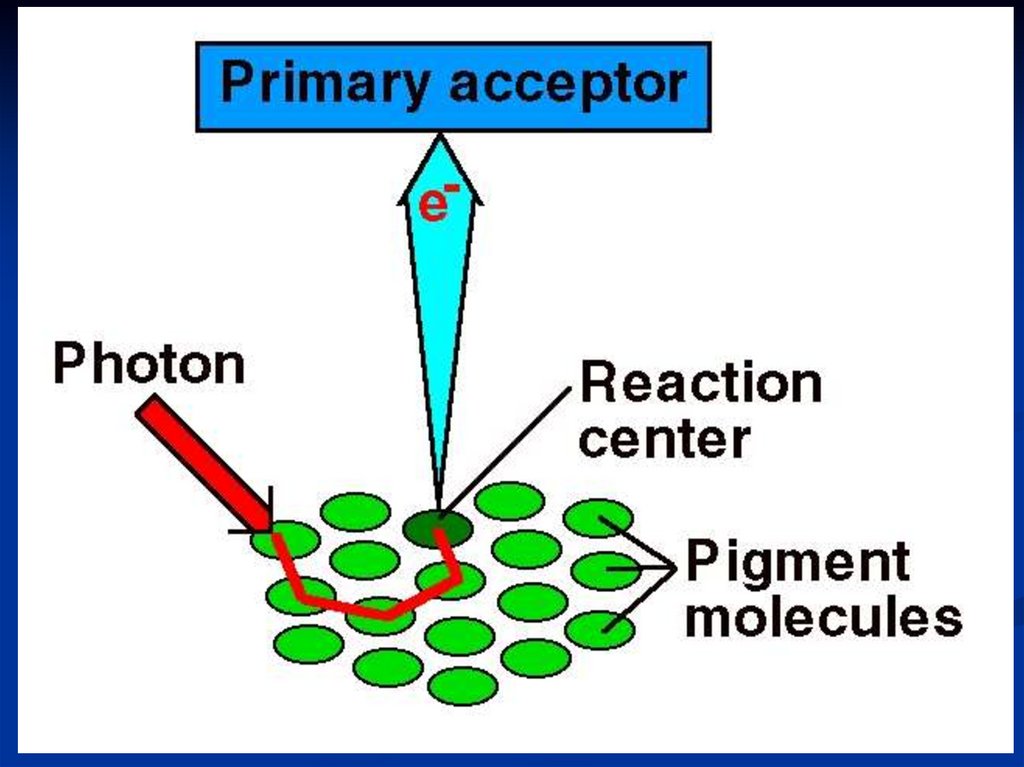
Поглощение кванта энергииСветособирающий комплекс (ССК)
/несколько сотен пигментов/
Фотохимический реакционный центр
Особая пара молекулы
хлорофилла а /димер/
75.
76.
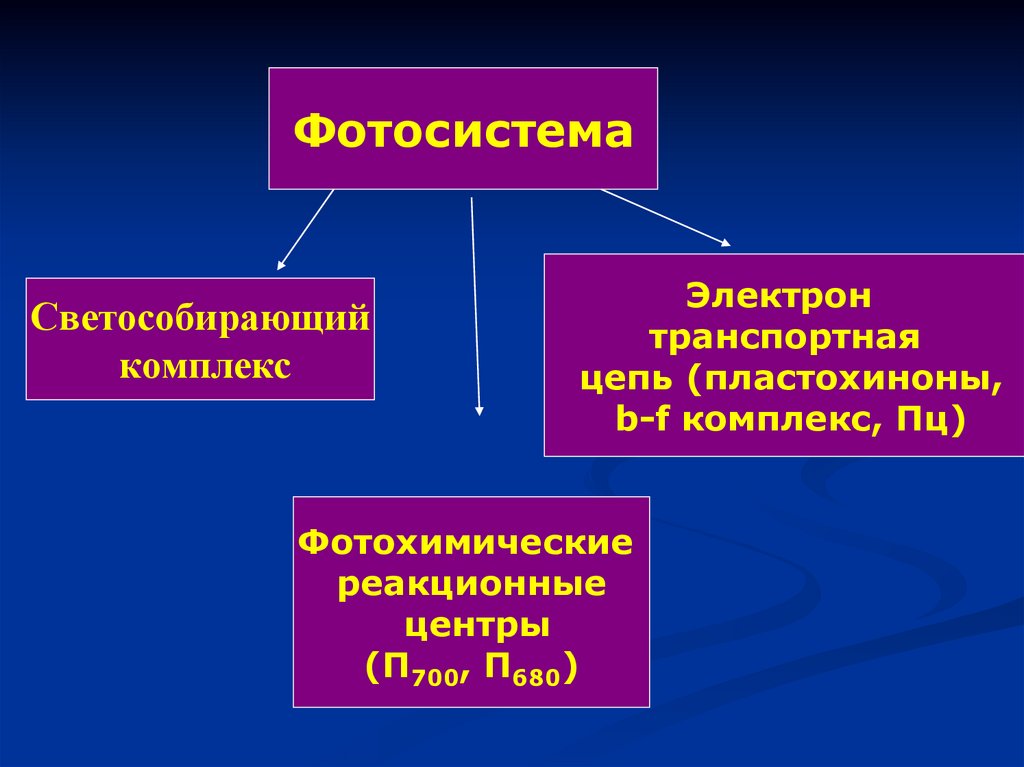
ФотосистемаСветособирающий
комплекс
Электрон
транспортная
цепь (пластохиноны,
b-f комплекс, Пц)
Фотохимические
реакционные
центры
(П700, П680)
77.
Энергия поглощенных квантов света стекаетсяот
сотен
молекул
пигментов
к
фотохимическому
реакционному
центру,
содержащему особую пару (димер) молекул
хлорофилла а, которые характеризуются
поглощением в наиболее длинноволновой
части солнечного спектра и выполняют роль
ловушки энергии возбуждения.
78.
Вфотосинтетический
комплекс
входят
молекулы первичного и вторичного акцепторов
электрона.
Молекула
хлорофилла, отдавая
первичному акцептору, окисляется.
электрон
Электрон поступает в электрон-транспортную
цепь (ЭТЦ).
79.
80.
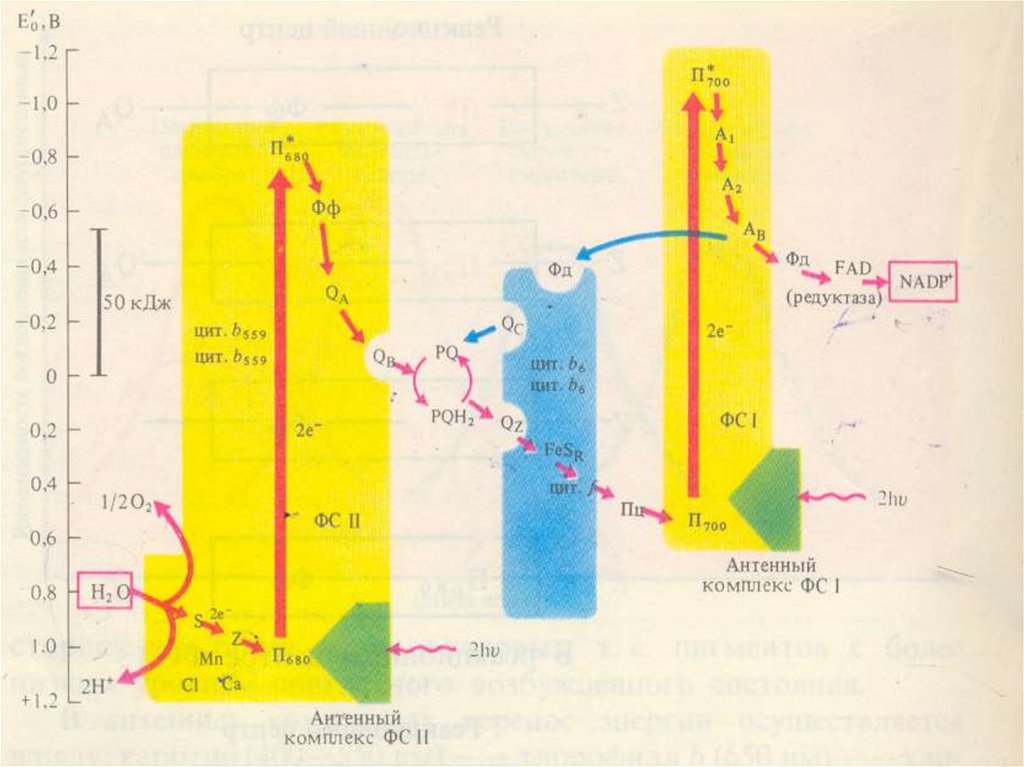
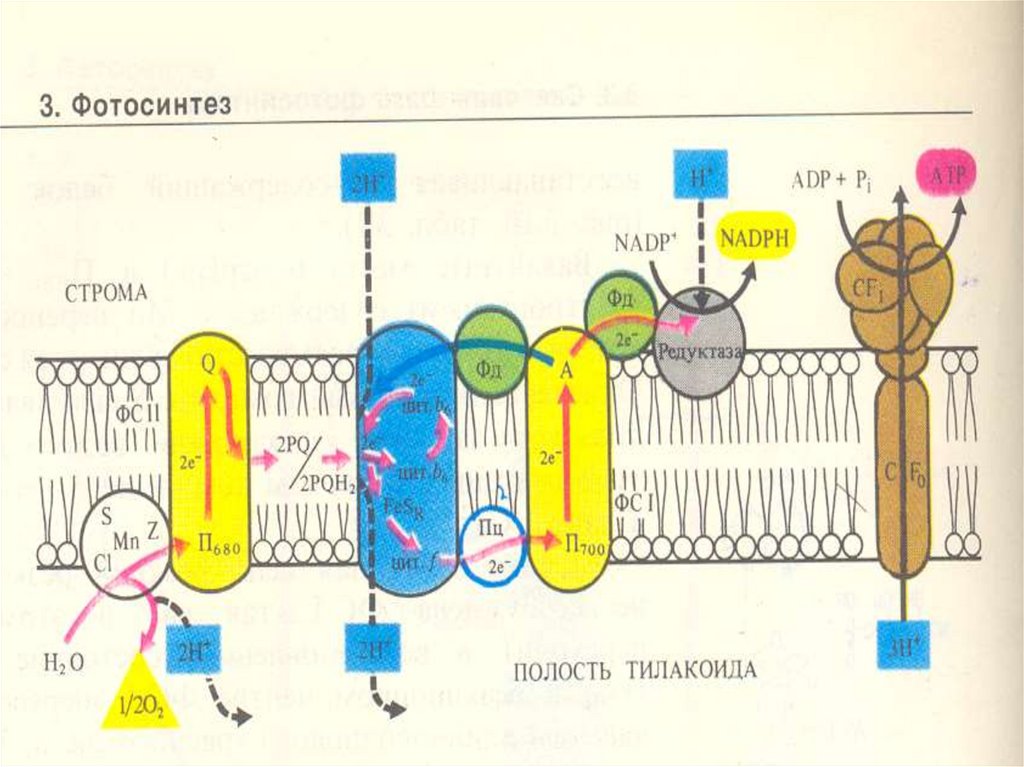
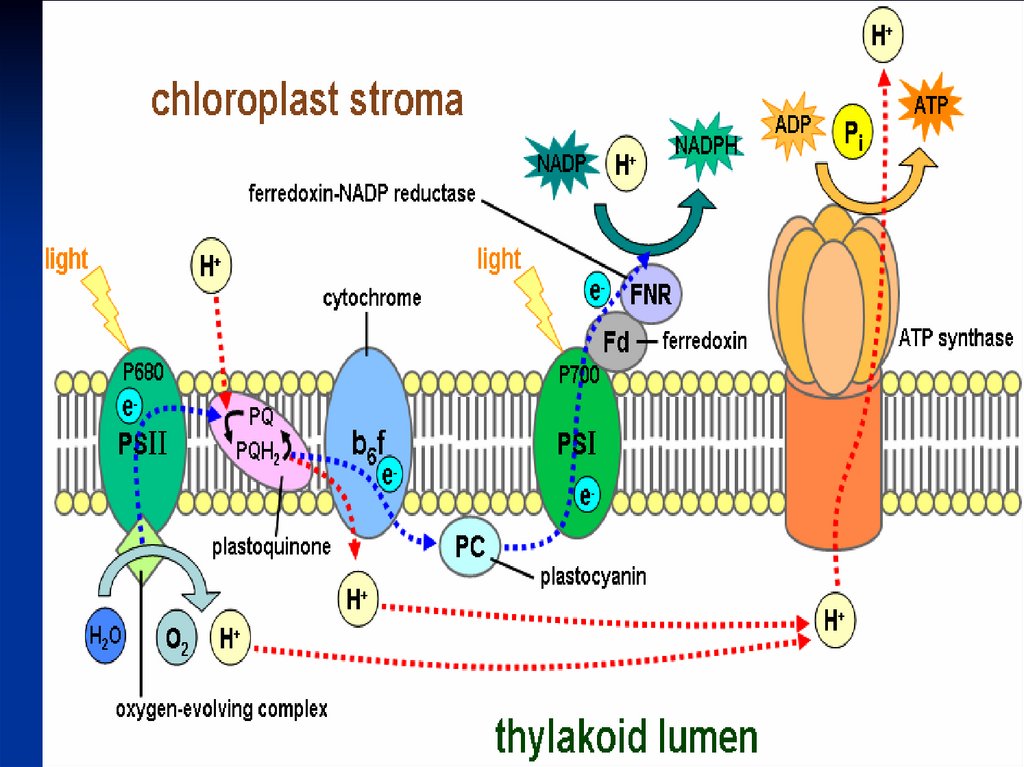
ФС 1Реакционный центр –димер пигмента Р700
(хлорофилл а с максимумом поглощения 700
нм)
Хлорофиллы а 675-695, компоненнты антенного
комплекса
хлорофилл а695 – мономерная форма (А1) –
первичный акцептор электрона
Вторичные акцепторы электронов - А2 и Ав
(железо-серные белки - FeS).
81.
ВФС
1
под
действием
растворимый в воде
FeS
ферредоксинд (Фд)
содержащий
пластоцианин
восстанавливается.
и
света
белок
Cuбелок
(Пц)
82.
ФС 2Реакционный центр - хлорофилл Р 680
Антенный комплекс - хлорофиллы
А 670-68З
Феофитин а (Фф) – первичный
акцептор электронов, передает эл-ны
первичному Пх-ну, ассоциированному с
FeS(QA), затем вторичному Пх-ну - (QB).
83.
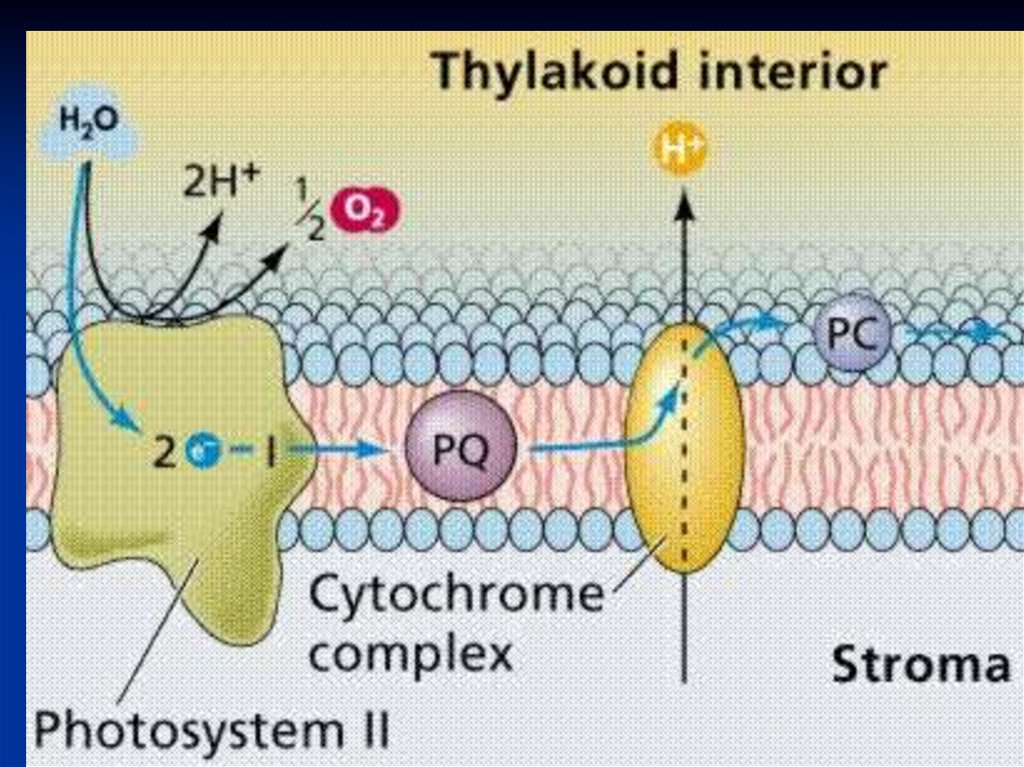
Переносчик эл-на Z является донором эл-надля П680 связан с окисляющим воду
комплексом,
В работе комплекса S принимают участие
Mn, Cl, Ca.
ФС2 :
Окисляет пластохиноны (PQ) и воду.
Выделяется кислород 02 и протоны.
ФС2 ответственна за разложение воды и
выделение кислорода
84.
Электронтранспортная цепь соединяет двеФС.
В нее входит:
- белковый комплекс (фонд) пластохинонов
- цитохромный б – f комплекс
-пластоцианин
85.
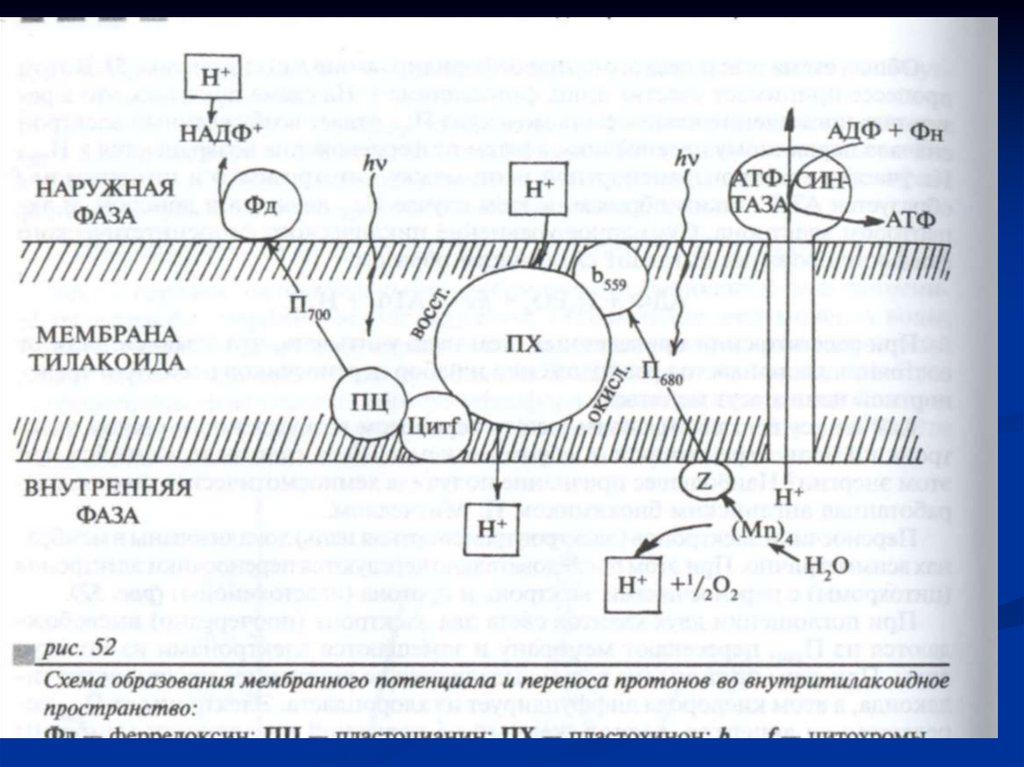
86.
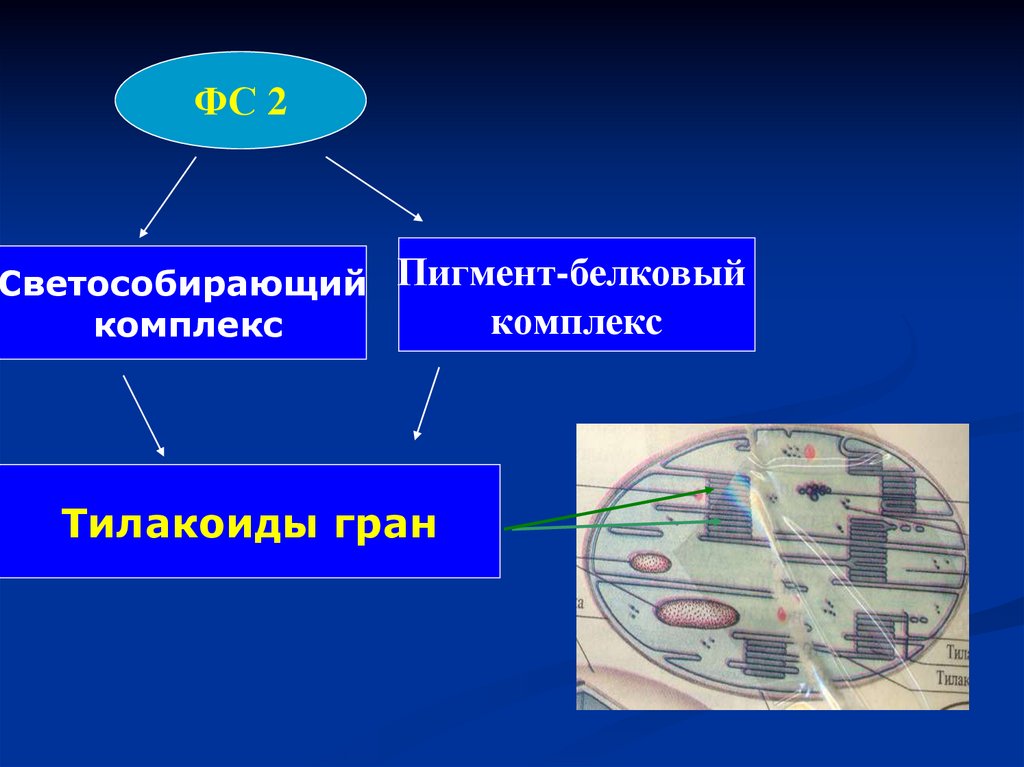
ФС 2Светособирающий Пигмент-белковый
комплекс
комплекс
Тилакоиды гран
87.
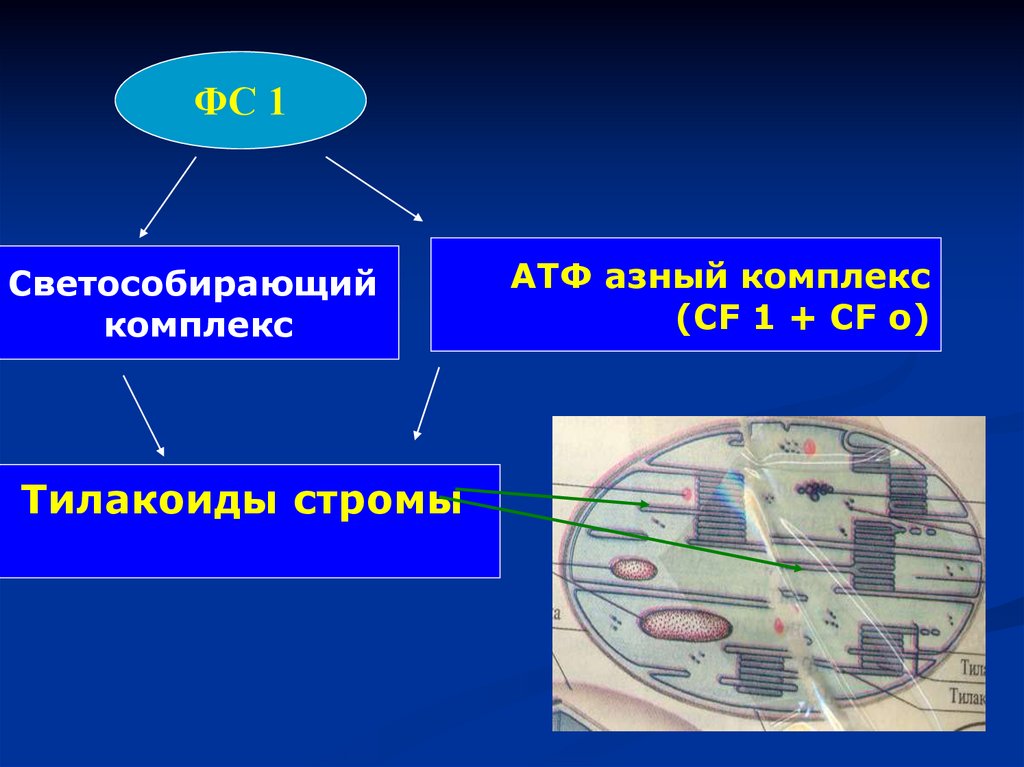
ФС 1Светособирающий
комплекс
Тилакоиды стромы
АТФ азный комплекс
(CF 1 + CF о)
88.
Цитохромный комплекс б - f расположенв тилакоидах стромы и гран.
Как они взаиодействуют?
Взаимодействие осуществляется с
помощью:
легкоподвижного липофильного
пластохинона PQ в липидной фазе,
водорастворимого Фд,
перемещающегося вдоль их наружных
поверхностей.
89.
Происходит перенос либо протонов иэлектронов, либо только электронов.
При этом молекула, отдающая протон или
электрон, окисляется, а молекула,
воспринимающая протон или электрон,
восстанавливается.
2 типа потока
электронов: циклический и
нециклический.
При нециклическом потоке - фотоокисление
воды и передача электрона от воды к НАДФ.
90.
Фотосинтетическое фосфорилированиепроцесс преобразования энергии света в
энергию АТФ (Д. Арнон).
Соответственно двум типам потока
электронов различают циклическое и
нециклическое фотофосфорилирование
.
-
91.
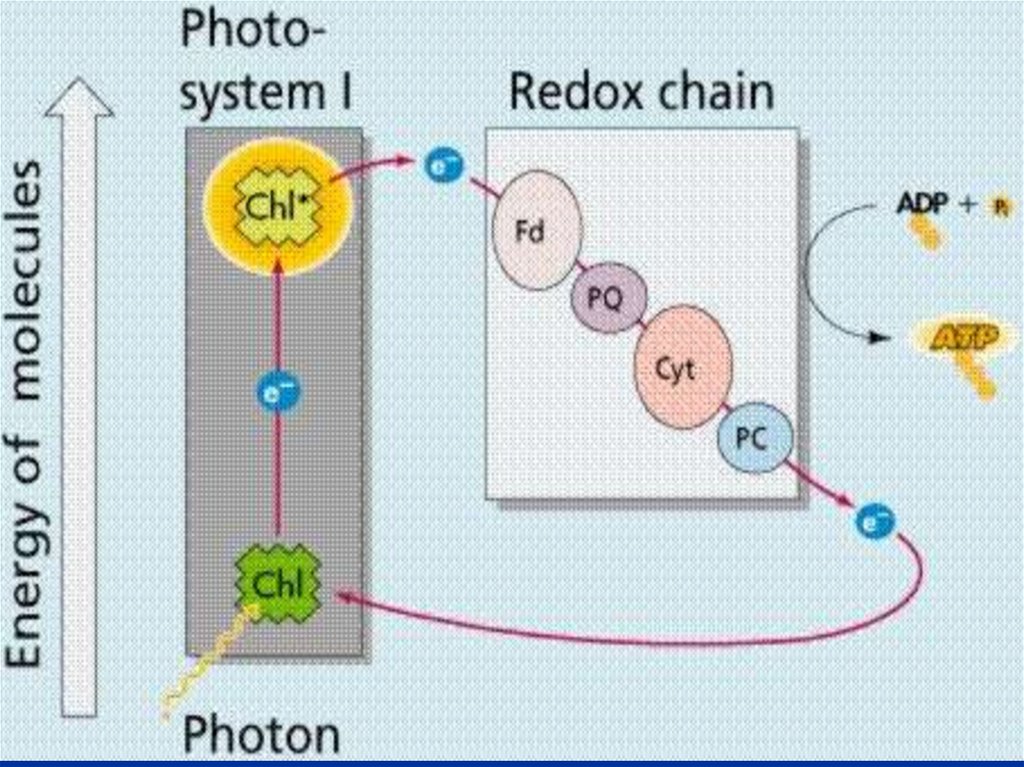
При нециклическом потоке электроновпринимают участие две ФС.
Энергия квантов света стекается к молекуле
пигмента П700, который входит в состав
реакционного центра ФС 1.
Хл + һν → Хл*,
Хл* → (Хл)+ + е,
где Хл* — возбужденная форма хлорофилла,
(Хл)+ — окисленная форма хлорофилла.
92.
Образовавшаяся электронная «дырка» вмолекуле П680 действует как сильный
окислитель и через ряд переносчиков при
участии ионов Mn и Cl отнимает электрон от
воды.
Этот электрон заполняет электронную
«дырку» в молекуле П680.
Происходит фотоокисление воды, и
выделяется молекула кислорода:
4 һν
2Н2О→4Н+ + 4ё + 02.
93.
ФС2П680
Фф
первичный акцептор
Пх
б/f-комплекс /от железо-серного белка к Цх f/
Цитохром окисляется Fe2+ - e → Fе 3+.
Следующий переносчик – Пц.
Пц – медь-содержащий
белок, в 1 мол-ле
белка 2 атома меди Сu2+ + е → Сu+.
Пц
- белок, соединяющий б/f-комплекс и ФС 1,
передают электроны П700.
П700 заполняет электронную «дырку».
94.
95.
В ФС2окисленный П680 - сильный
окислитель.
В результате в ФС2 вода разлагается,
выделяется кислород
Водоразлагающий комплекс (ВРК) -
в
активном центре - ионы марганца (Мn2+),
они являются донором электронов для
П680 :
2Мn 4+ +2Н20 → 2Мn 2+ +4Н+ + 4е + О2.
96.
После последовательной передачи четырехэлектронов от ВРК к П680 происходит
разложение сразу двух молекул воды,
сопровождающееся выделением одной
молекулы кислорода и четырех ионов
водорода, которые попадают во
внутритилакоидное пространство
хлоропласта.
Перенос электрона по цепи переносчиков от
ФС II к ФС1 сопровождается образованием
АТФ из АДФ и неорганического Фн (АДФ +
Фи → АТФ).
97.
Суммарное уравнение процесса нециклическогофотофосфорилирования может быть выражено
следующим образом:
2НАДФ + 2Н20 + 2АДФ + 2Н3Р04 → 2НАДФН + 2Н+
+ 2АТФ + 02.
Отличительные особенности нециклического
фотосинтетического фосфорилирования:
1) окисление двух молекул воды;
2) участие двух фотосистем;
3) передача электронов от молекул воды
(первичный донор) через электронтранспортную
цепь на НАДФ (конечный акцептор).
98.
Продукты нециклическогофотофосфорилирования:
1. Никотинамидадениндинрнуклеотидфосфат
(НАДФН + Н)
2.
АТФ.
Эти соединения в дальнейшем используются в
темновой фазе фотосинтеза.
99.
При циклическом фотофосфорилированиипринимает участие фотосистема 1.
В результате поглощения кванта света
молекула П700 отдает возбужденный
электрон сначала первичному
переносчику, а затем от Фд возвращается
к П700.
На участке электронтранспортной цепи
между цитохромом б и цитохромом f
образуется АТФ.
100.
П700 является донором и акцепторомэлектронов.
Циклическое фотофосфорилирование –
суммарное уравнение:
АДФ+Н3РО4+ һν →АТФ+Н20.
101.
Хемиосмотическая теория П. Митчелла(английский биохимик).
Переносчики электронов
(электронтранспортная цепь)
локализованы в мембранах
асимметрично.
Существует 2 типа переносчиков
электронов:
- переносчики электронов – цитохромы
переносчики электронов и протонов пластохиноны .
102.
При поглощении 2-х квантов света дваэлектрона (поочередно) высвобождаются из
П680, пересекают мембрану и замещаются
электроны из молекул воды.
При этом протоны (из воды) выделяются во
внутреннее пространство тилакоида, а атом
кислорода диффундирует из хлоропласта.
Электроны от П680 передаются на акцептор,
расположенный на наружной поверхности
мембраны тилакоида. Этим акцептором
является пластохинон (ПХ).
103.
104.
105.
106.
Пластохинон — производное хинона, вполностью окисленном состоянии
содержит два атома кислорода,
соединенных с углеродным кольцом
двойными связями.
В полностью восстановленном состоянии
атомы кислорода в бензольном кольце
соединяются с протонами:
Гидрохинон
+2е+2Н
Хинон
107.
При переносе пары электронов от Хл680 на Хл700 вовнутреннем пространстве тилакоидов
накапливаются протоны.
В результате создается электрохимаческий
потенциал водорода (∆ψН), имеющий две
составляющие: химическую ∆рН
(концентрационную), и электрическую ∆μН,
обусловленную противоположным зарядом разных
сторон мембраны.
Чтобы использовать эту энергию, необходимо
обеспечить обратный поток протонов.
Энергия, выделяющаяся при этом, и используется
для образования АТФ.
108.
Накопление протонов внутритилакоида происходит не
только при окислении Пх, но и в
результате фотоокисления воды
(∆μН).
109.
Сопряжение диффузии протонов черезмембрану осуществляется ферментным
комплексом, называемым АТФ-синтазой или
сопрягающим фактором.
Этот комплекс состоит из двух частей -
факторов сопряжения:
- круглой «шляпки» F1, выступающей с
наружной стороны мембраны (в ней
располагается каталитический центр
фермента),
- «ножки» F0, погруженной в мембрану.
110.
Мембранная часть состоит изполипептидных субъединиц (а, б, с) и
формирует в мембране протонный канал,
по которому ионы водорода попадают к
фактору сопряжения F1.
Белок F1 представляет белковый
комплекс, который состоит из девяти
субъединиц пяти типов: α, β, γ, δ, ε.
111.
Белковый комплекс можно сравнительнолегко отделить от мембраны, при этом он
сохраняет способность катализировать
гидролиз АТФ, т.е. является АТФазой.
Направленный транспорт возможен
только в случае, если АТФ-синтаза
встроена в мембрану.
112.
113.
Существуют две гипотезы относительно механизмафосфорилирования (прямой механизм и косвенный).
1-я гипотеза - фосфатная группа и АДФ
связываются с ферментом в активном участке
комплекса F1.
Два протона перемещаются через канал по
градиенту концентрации и соединяются с
кислородом фосфата, образуя воду.
Это делает фосфатную группу весьма
реакционноспособной, и она присоединяется к АДФ
с образованием АТФ.
114.
Вторая гипотеза, которая в последние годыполучила экспериментальные подтверждения
(косвенный механизм) - АДФ и Ф, соединяются
в активном центре фермента (субъединицы α и
β фактора F1) спонтанно.
Однако образовавшаяся АТФ прочно связана с
ферментом, и для ее освобождения требуется
энергия.
Энергия доставляется протонами, которые,
связываясь с ферментом (протонирование),
изменяют его конформацию, после чего АТФ
высвобождается.
115.
Вторая гипотеза, которая в последние годыполучила экспериментальные подтверждения
(косвенный механизм) - АДФ и Ф, соединяются в
активном центре фермента (субъединицы α и β
фактора F1) спонтанно.
Однако образовавшаяся АТФ прочно связана с
ферментом, и для ее освобождения требуется
энергия. Энергия доставляется протонами, которые,
связываясь с ферментом (протонирование),
изменяют его конформацию, после чего АТФ
высвобождается.
Продукты световой фазы, в которых аккумулирована
энергия света, — АТФ и НАДФН
Д. Арнон назвал их ассимиляционной силой,
поскольку они используются в темновую фазу, в
ходе которой СО2 восстанавливается до углеводов.
116.
1.Реакционный центр - это...1. Это место в тилакоиде
2. Участок граны
3. Комплекс пигментов П 700 и
П 680 с белками
4.Антенный комплекс
5. Водоокисляющий комплекс
2. Источник кислорода в
процессе фотосинтеза...
1.
Углекислый газ
2.Глюкоза
3. Вода
4.Крахмалдан
5.Фосфоглицериновая кислота
3.Где проходит световая фаза фотосинтеза?
1.В строме хлоропластов
2. Внутренняя мембрана хлоропластов
3.Тилакоиды хлоропластов
4. Внешняя мембрана хлоропластов
5. Строма хлоропластов
4. Что не входит в фотосистему І
1.НАДФ
4. Пластоцианин
2.П700
5.Пластохинон
3. Железо-серный белок
5. Продукт световой фазы фотосинтеза...
1. Углевод
2. Кислород
3.Вода
4. Углекислый газ
5. НАДН
117.
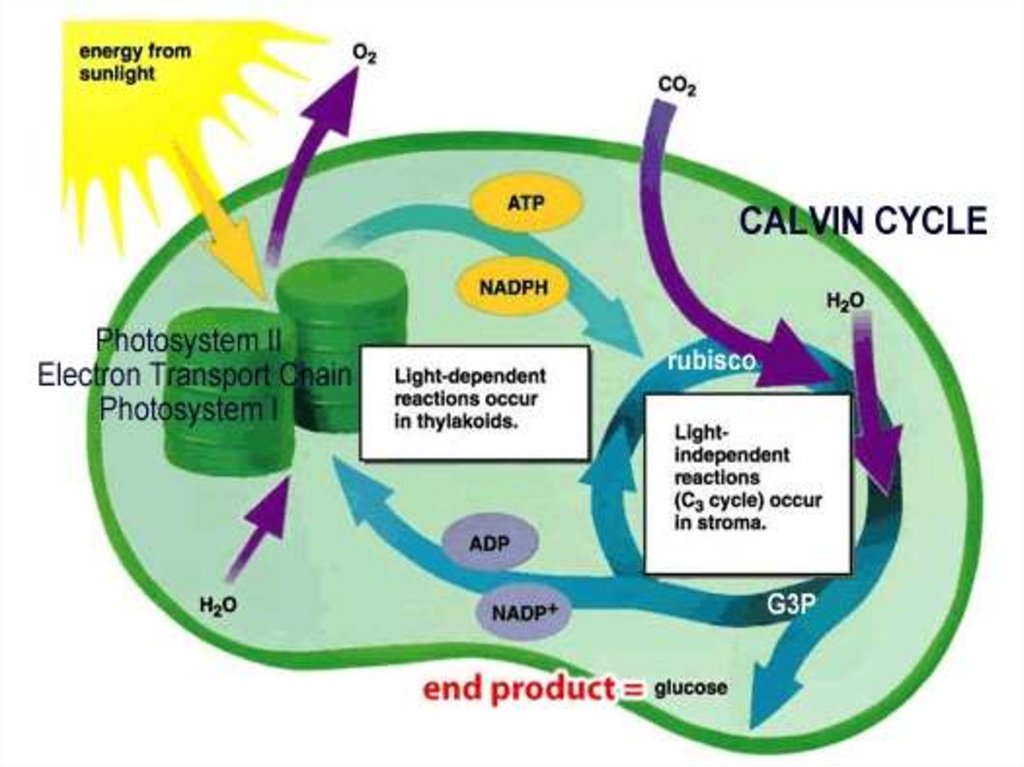
Темновая фаза фотосинтеза1. Цикл Кальвина - С3-путь фотосинтеза.
2. Фотодыхание
3. С4 – путь фотосинтеза. Цикл Хэтча и Слэка
4. САМ-метаболизм.
118.
Свет (hv)6С02 + 12Н20 → С6Н1206 + 602+ 6H2O
Глюкоза
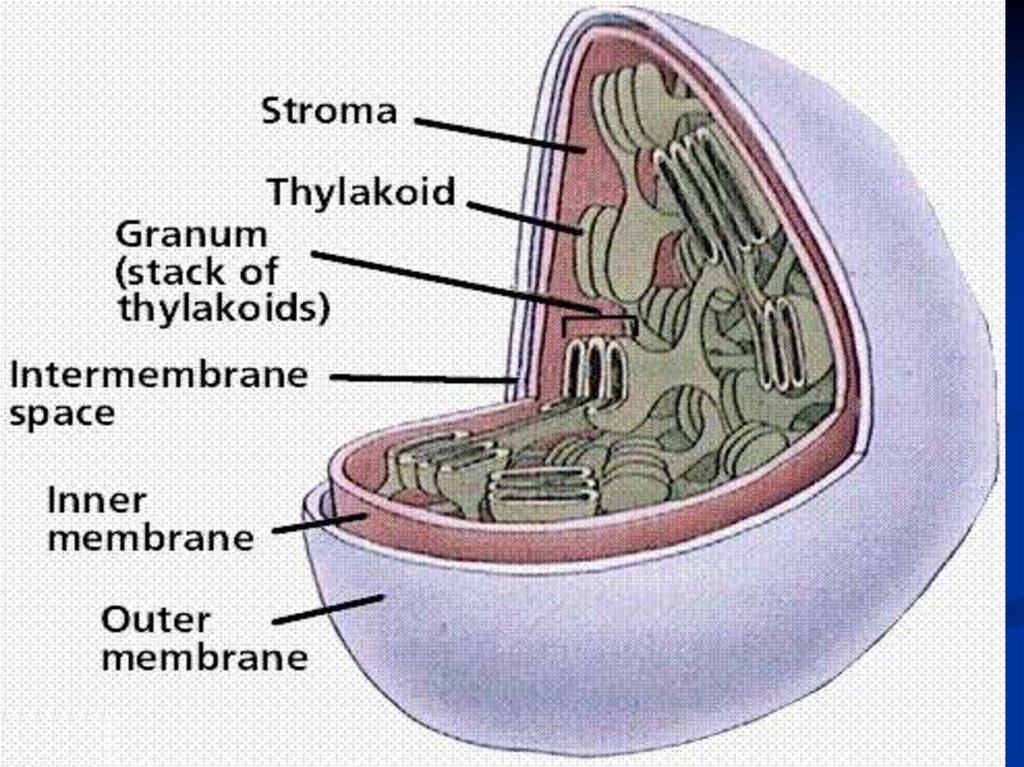
Световая фаза проходит в тилакоидах,
Темновая фаза – в строме.
119.
120.
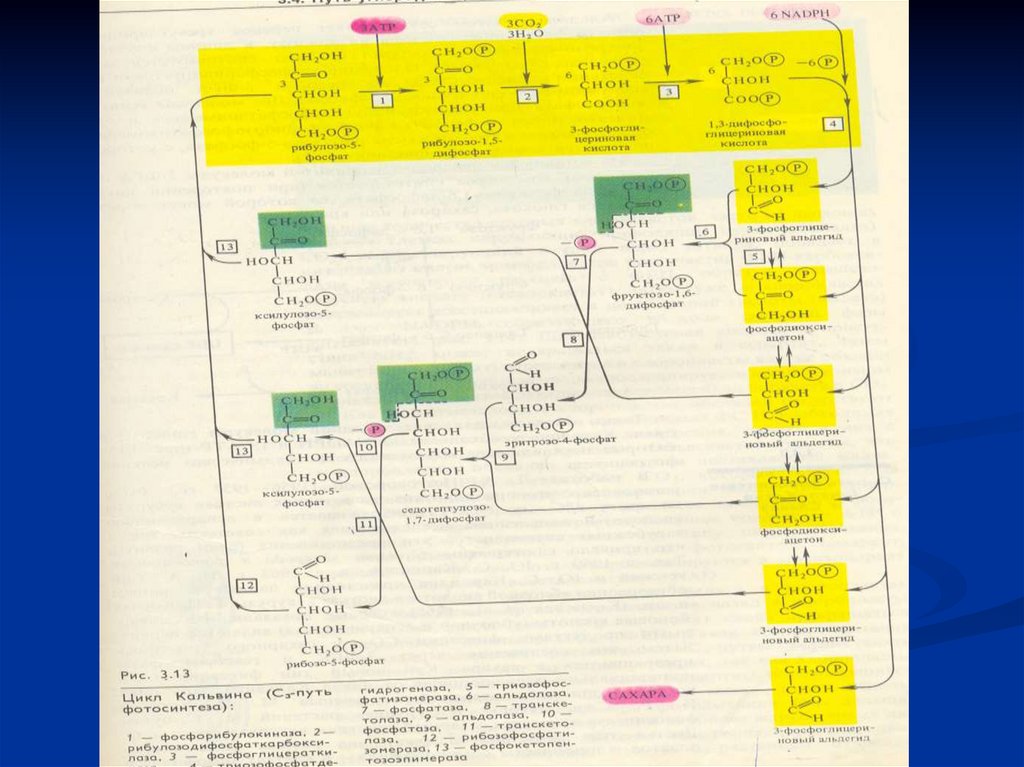
Пути ассимиляции СО21946-1956 г.г. - М.Кальвин
С3 - путь, цикл Кальвина
С3 – у растений, растущих в умеренных зонах
продукты световой фазы
6АТФ+6НАДФ.Н2
3СО2+3РБФ+3Н2О→6ФГК→→→→6ФГА→1ФГА
углеводы
3АТФ →→В←Б←А ←←←←←← 5ФГА
из световой фазы
121.
С3 - путь, цикл Кальвина3 этапа:
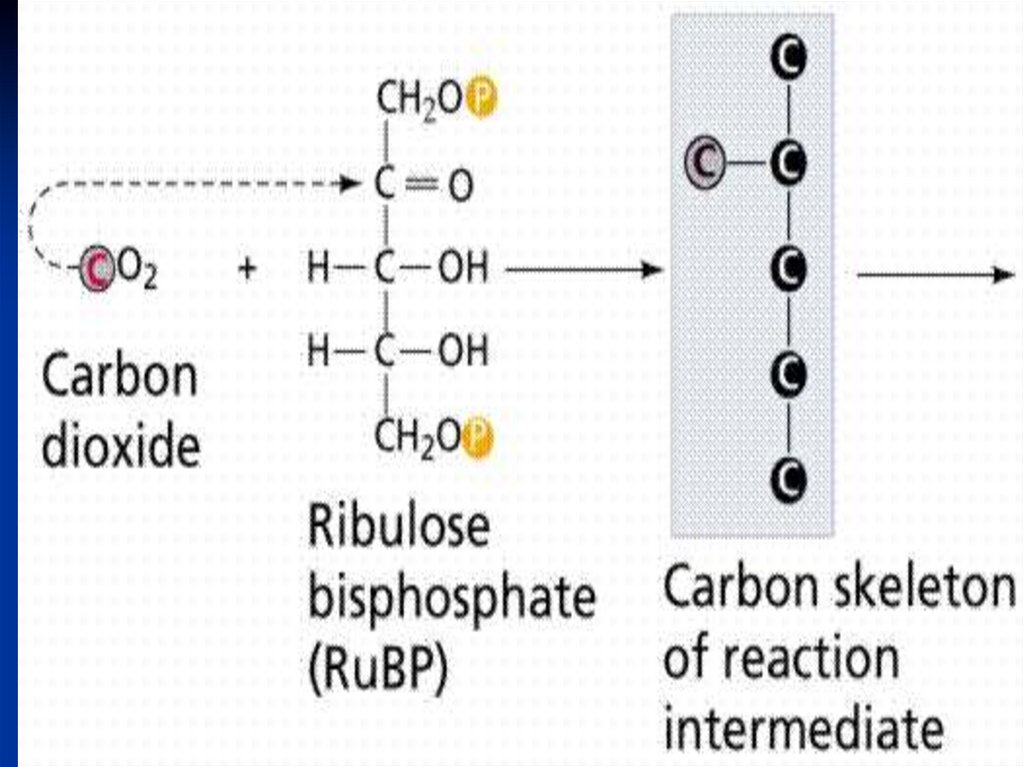
I Карбоксилирование.
АТФ
РБФ- карбоксилаза
Риб-5Ф → → → Риб-1,5 диФ + СО2 → 3-ФГК
Фосфорибулозокиназа
Первый прдукт карбоксилирования
– соединение содержащее 3 атома
углерода – фосфоглицериновая
кислота - ФГК
122.
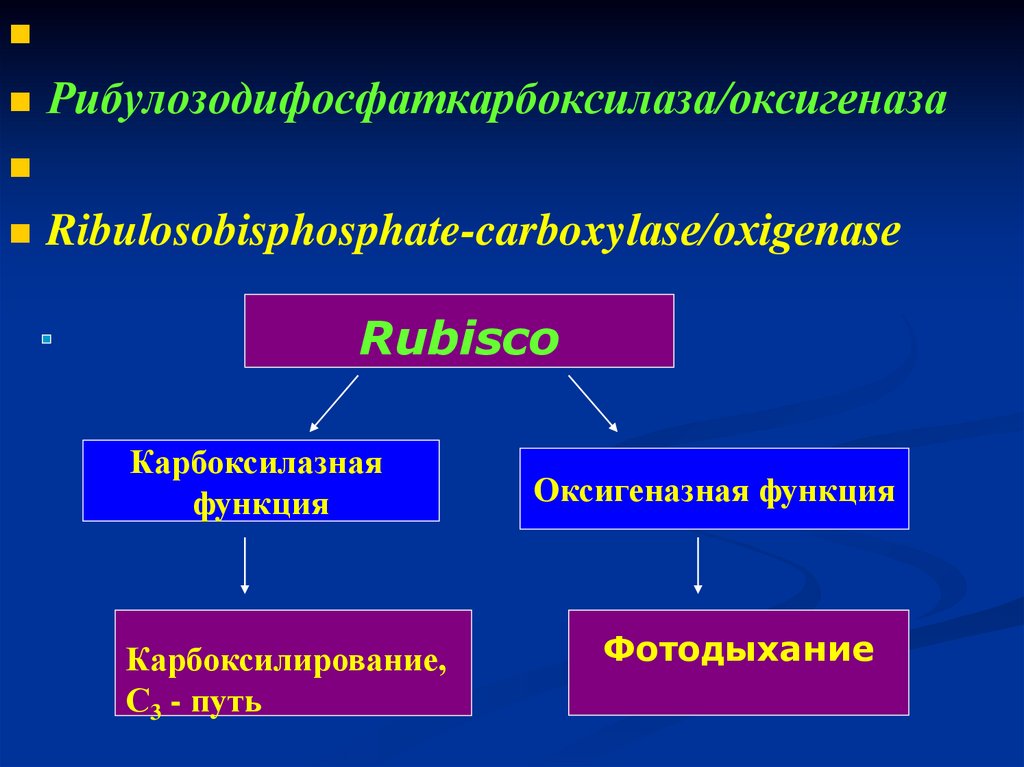
Рибулозодифосфаткарбоксилаза/оксигеназаRibulosobisphosphate-carboxylase/охigenase
Rubisco
Карбоксилазная
функция
Карбоксилирование,
С3 - путь
Оксигеназная функция
Фотодыхание
123.
124.
II Фаза восстановления.АТФ
НАДФН
3-ФГК →→→→→→→ 1,3 ФГК →→→→ ФГА
фосфоглицераткиназа
восстановление
III фаза регенерации
6ФГА
1 ФГА
углеводы
5 ФГА
РБФ
125.
126.
2 мол.3-ФГА ( 2 цикла Кальвин)↓
2 мол. Фруктозо-1,6-дифосфат
↓
↓
Гл-1-Ф
Фр-6-ф
↓ + УТФ
УДФГл
Крахмал
↓+ Фр-6-ф
сахарозо-Ф
↓
сахароза
↓
глюкоза + фруктоза
127.
128.
Реакции темновой фазы:6РБФ+6СО2 +12НАДФН+12 Н+ + 18 АТФ
6РБФ+С6Н12О6 +Н2О+12НАДФ+18АДФ+Фнг.
Цикл Кальвина присущ всем растениям, но
пути доставки СО2 к ц. Кальвину могут быть
различными.
129.
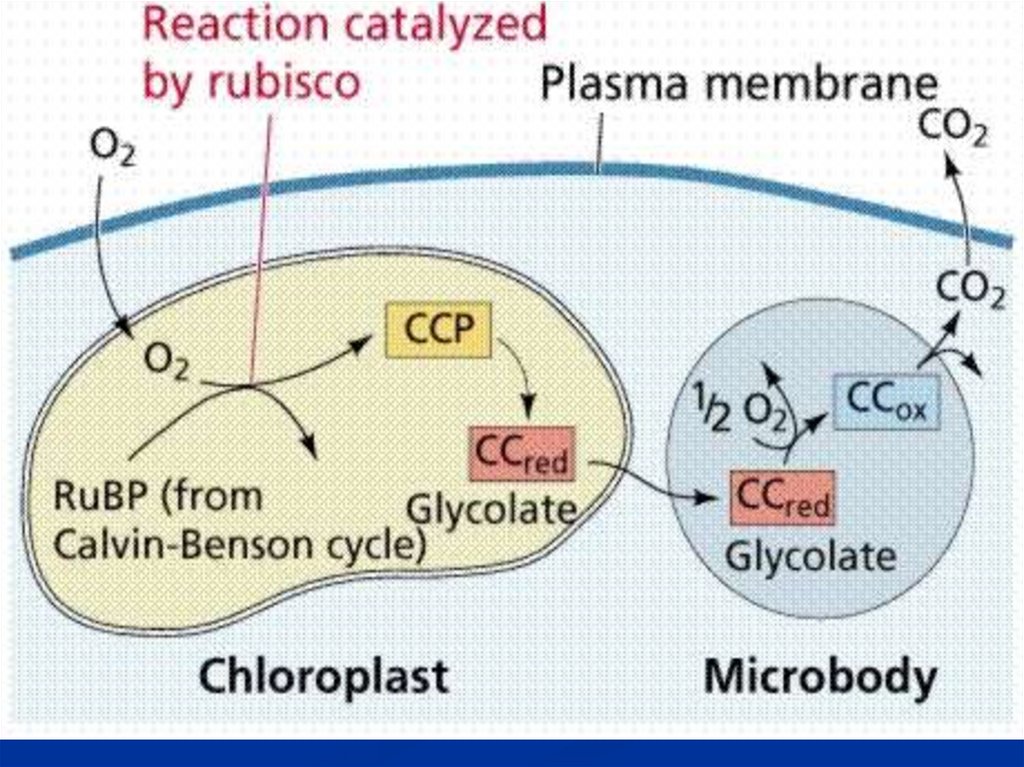
Фотодыхание(гликолатный цикл).
В этом процессе связывается О2, освобождается
СО2↑.
Первичный продукт фотодыхания – гликолевая
кислота.
Поэтому цикл называют гликолатным.
СО2 - С3 – путь фотосинтеза.
СО2 О2 - Фотодыхание.
Rubisco выполняет оксигеназную функцию.
130.
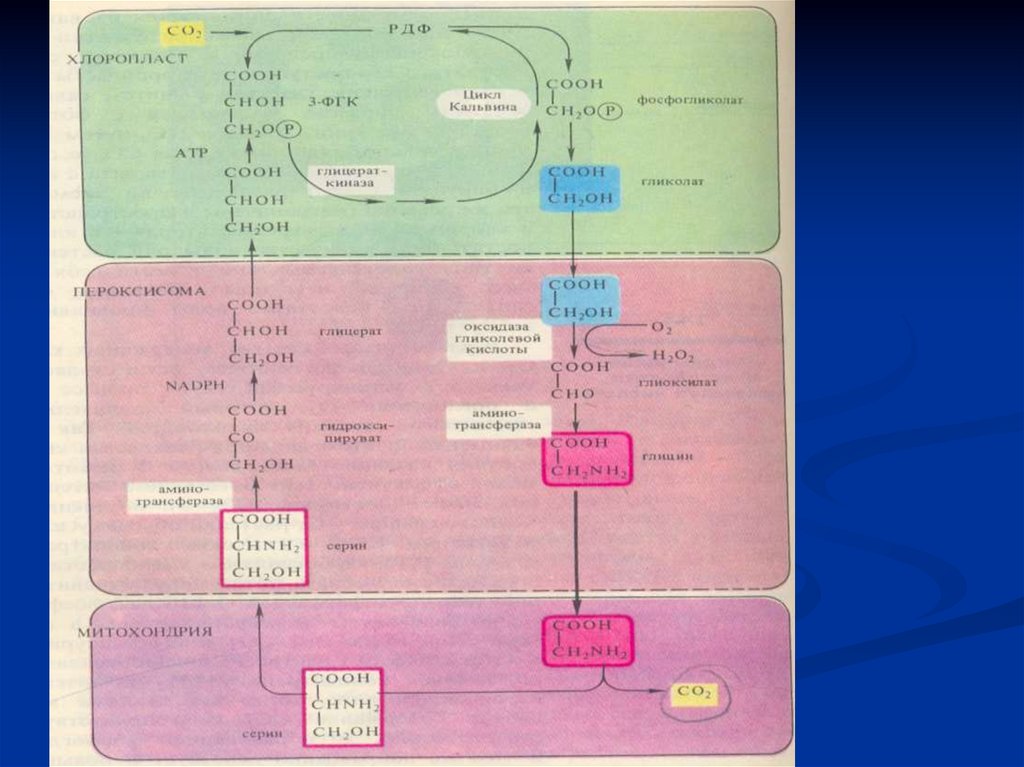
В процессе фотодыхания образуются 3ФГК и 2 – фосфогликолевая кслота.Фотодыхание проходит в 3-х органеллах:
1.Хлоропласт
- РБФ
гликолевая
кислота /гликолат/
2.Пероксисома – Гликолат
глицин
3.Митохондрия – Глицин
серин
131.
132.
133.
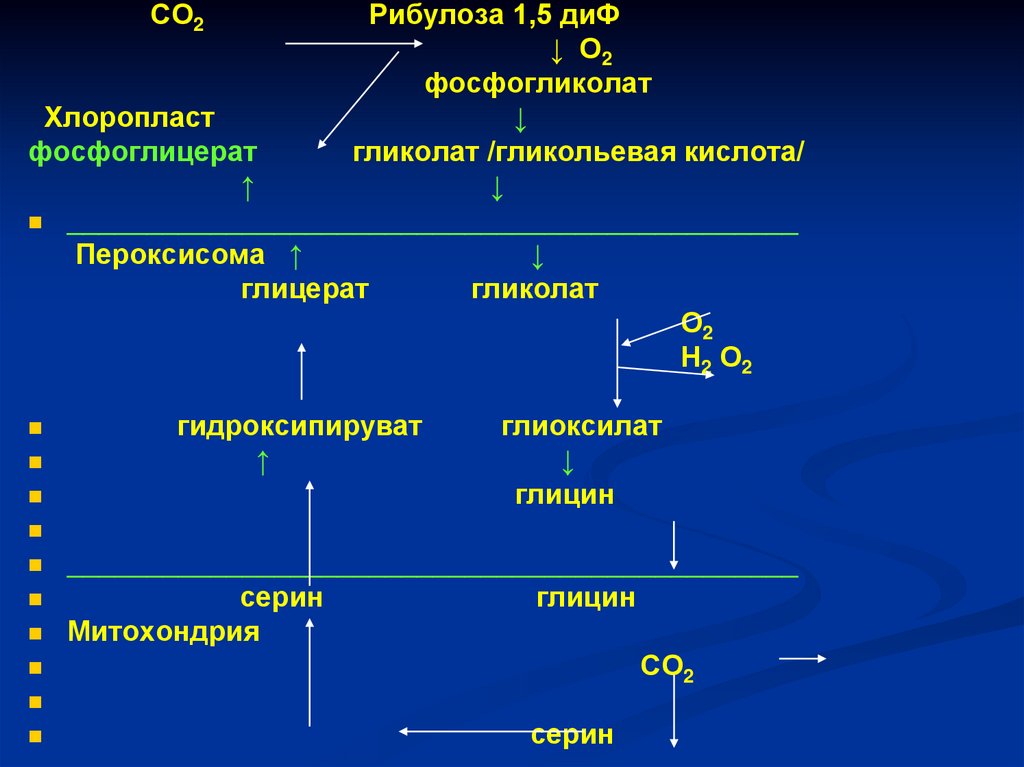
СО2Рибулоза 1,5 диФ
↓ О2
фосфогликолат
Хлоропласт
↓
фосфоглицерат
гликолат /гликольевая кислота/
↑
↓
______________________________________________
Пероксисома ↑
↓
глицерат
гликолат
О2
Н2 О2
гидроксипируват
↑
глиоксилат
↓
глицин
______________________________________________
серин
глицин
Митохондрия
СО2
серин
134.
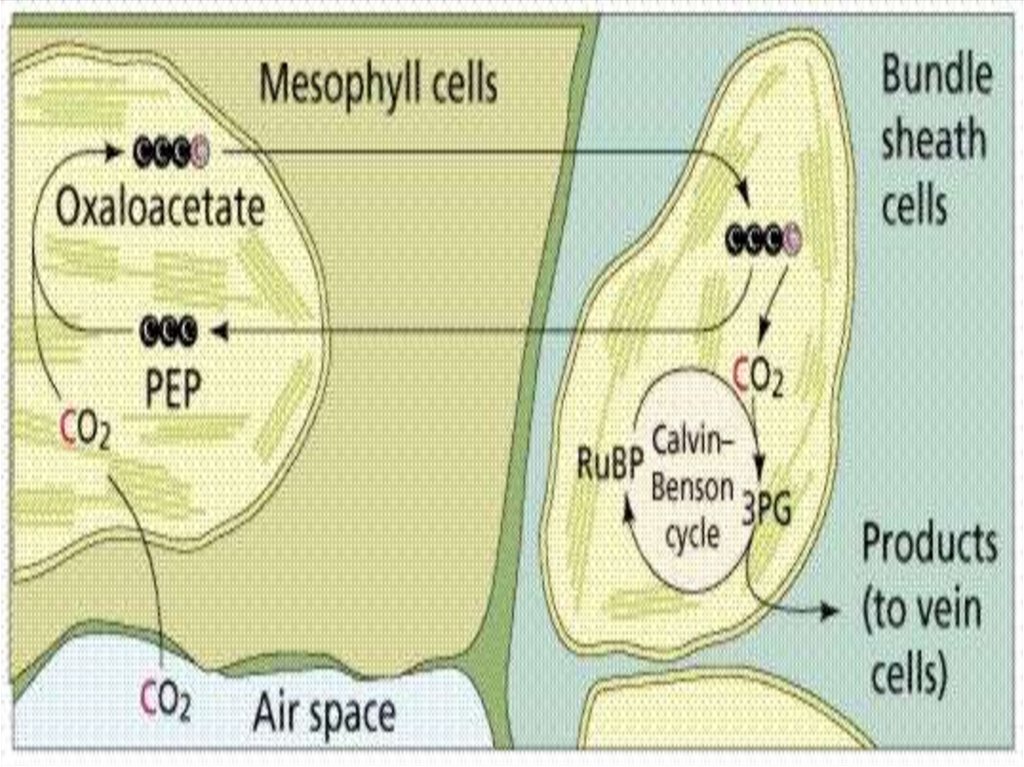
C4 –путь фотосинтезаЦикл Хэтча и Слэка
Появился в процессе адаптации к изменниям
условий окружающей среды
1966 г. – М.Д.Хетч и К.Р.Слэк
1960 г. Ю.С.Карпилов
У некоторых субтропических и тропических растений
Aмарант, кукуруза, сахарный тростник
Первый продукт карбоксилирования –
4 атома углерода.
135.
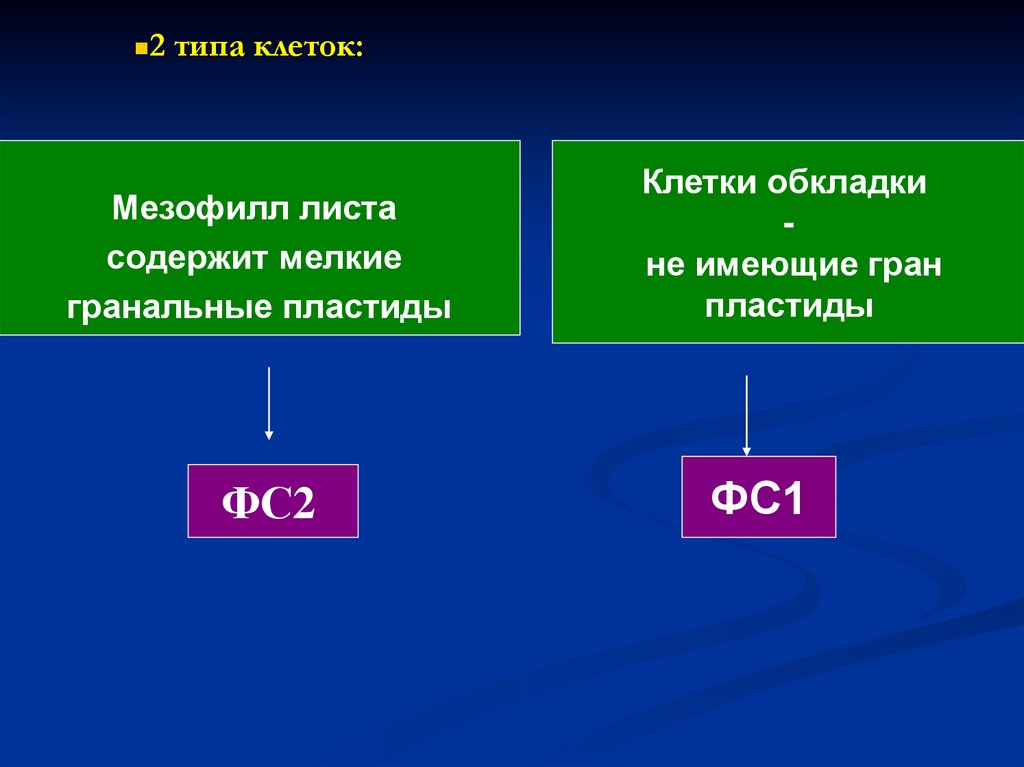
2 типа клеток:Мезофилл листа
содержит мелкие
гранальные пластиды
ФС2
Клетки обкладки
не имеющие гран
пластиды
ФС1
136.
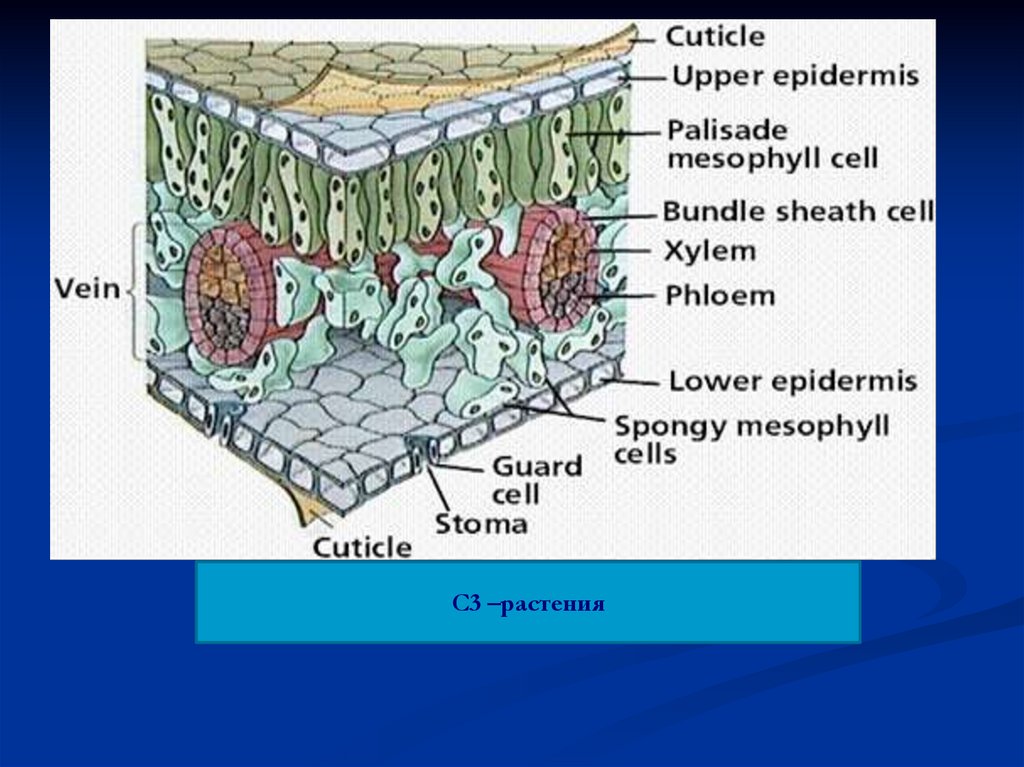
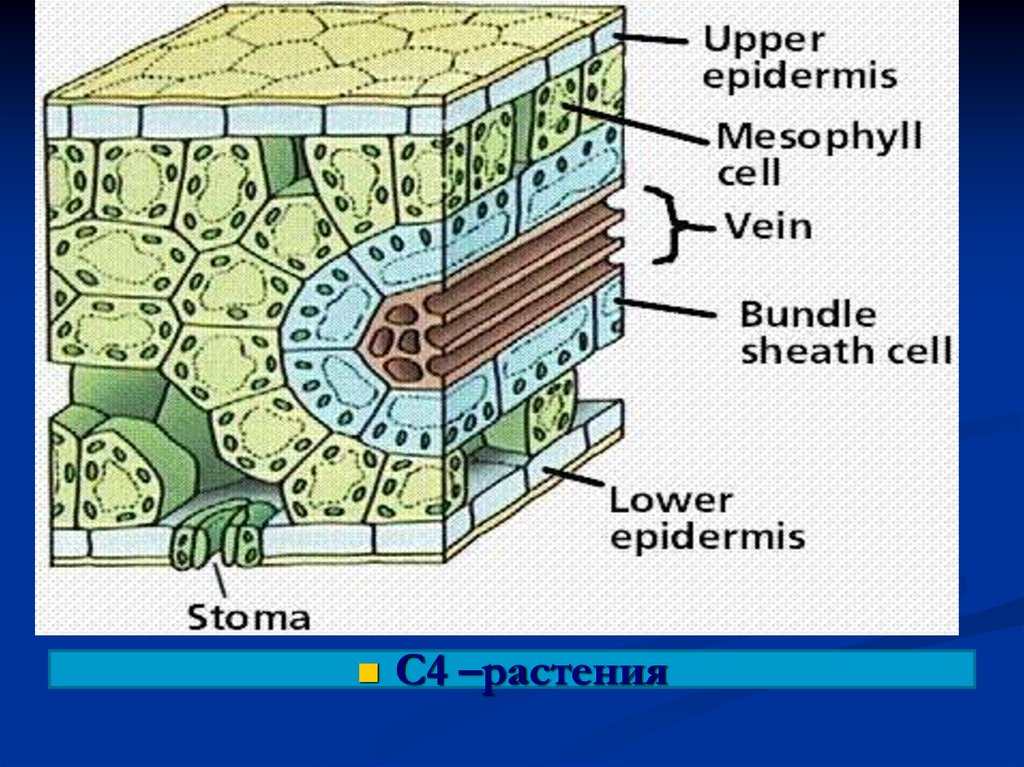
С3 –растения137.
С4 –растения138.
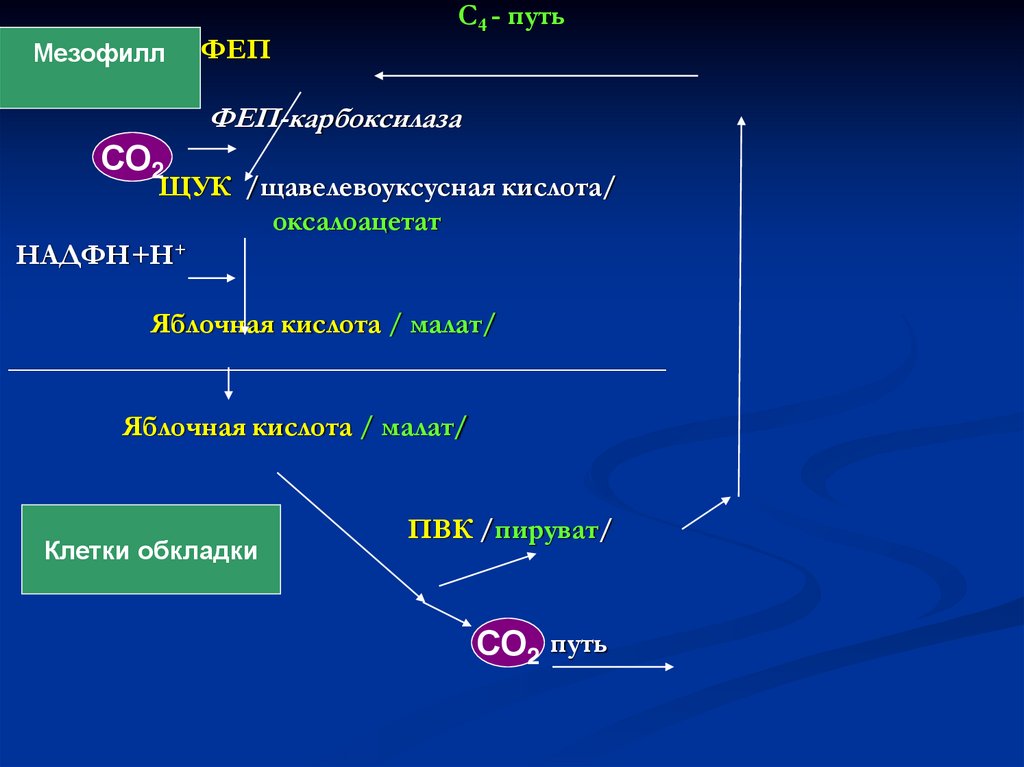
МезофиллФЕП
C4 - путь
ФЕП-карбоксилаза
СО2
ЩУК /щавелевоуксусная кислота/
оксалоацетат
НАДФН+Н+
Яблочная кислота / малат/
______________________________________________
Яблочная кислота / малат/
Клетки обкладки
ПВК /пируват/
С32- путь
СО
139. Мезофилл Клетки обкладки
140.
141.
Преимущества С4 – пути:•У С3-растений
интенсивность фотодыхания высокая, у
С4 – растений – низкая.
•Rubisco участвует
и в процессе фотодыхания:
•РБФ + СО2 →2ФГК – фосфоглицериновая кислота
•РБФ + 02→ ФГК – фосфогликолевая кислота.
•Реакции оксигеназаная и карбоксилазная конкурируют
друг с другом
•У С4-растений в клетках обкладки проходит цикл
Кальвина. Там концентрация 02 всегла будет низкой, так
как в этих клетках работает только ФС1, где не
происходит выделения 02
•Оксигеназная и карбоксилазная функция Rubisco
зависит от концентрации 02 и СО2-.
142.
2. Температурный оптимум фотосинтеза/ у С3растений 20-25 °С, у С4 – 30-45°С.3. С4-растений светонасыщение фотосинтеза
происходит прибольшей интенсивности солнечного
света по сравнению с С3-растениями.
Например, у С3-растений при использовании 50%
полной солнечной энергии дальше уже интенсивность
фотосинтеза не увеличивается.
У С4-растений при увеличении интенсивности
солнечной энергии, интенсивность фотосинтеза
упродолжает увеличиваться.
Поэтому, у С4-растений интенсивность фотосинтеза
при высокой температуре и солнечной энергии может
увеличиваться.
143.
4. У С4-растений продукты цикла Кальвинаобразуются в клетках обкладки. Это
способствует быстому распространению
ассимилятов, и способствует усилению
интенсивности фотосинтеза.
5. С4-растения устойчивы к засухе, высокой
температуре
144.
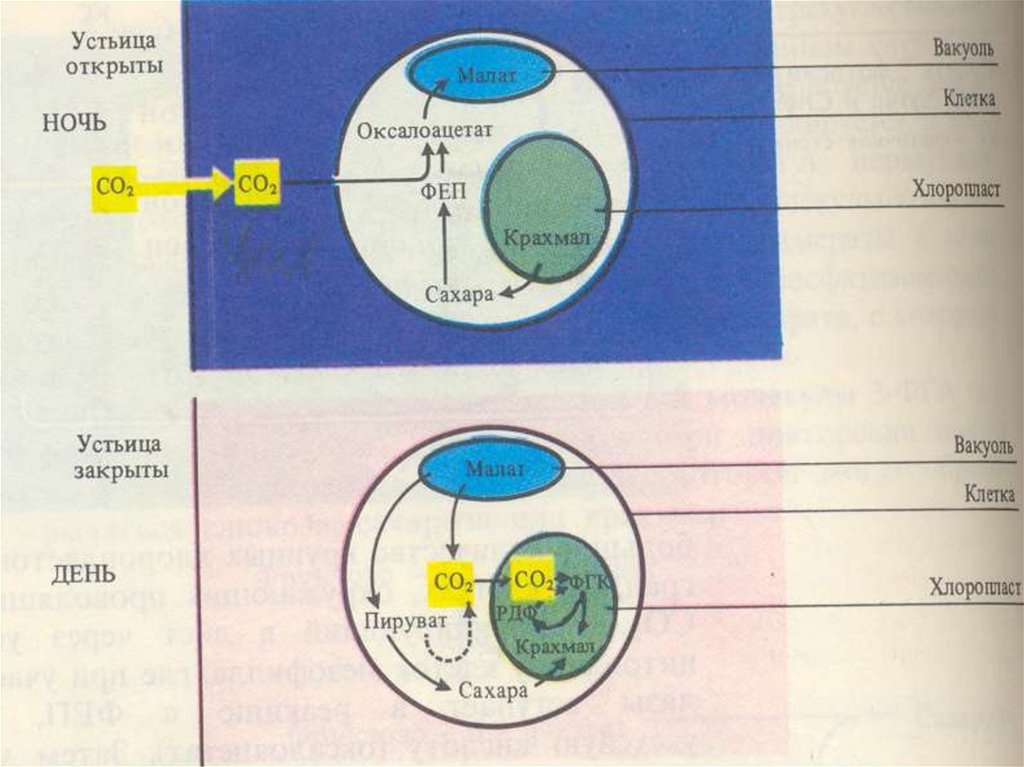
САМ /Crassulacean acid metabolism/-метаболизм
У тропических растений /суккуленты/.
Устьица ночью открыты, днем - закрыты.
Теряют мало влаги, устойчивы к засухе.
145.
Crassula_Ovata146.
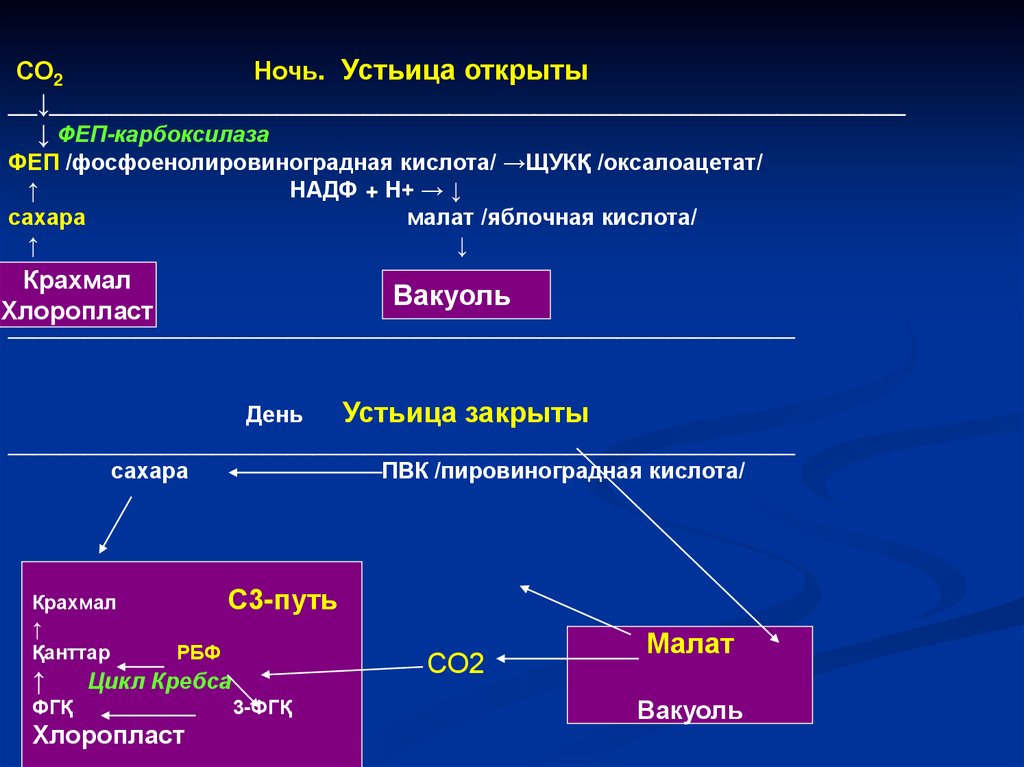
СО2Ночь. Устьица открыты
__↓____________________________________________________________
↓ ФЕП-карбоксилаза
ФЕП /фосфоенолировиноградная кислота/ →ЩУКҚ /оксалоацетат/
↑
НАДФ + Н+ → ↓
сахара
малат /яблочная кислота/
↑
↓
Крахмал
Хлоропласт
Вакуоль
______________________________________________________________
День
Устьица закрыты
______________________________________________________________
сахара
ПВК /пировиноградная кислота/
Крахмал
↑
Қанттар
↑
С3-путь
РБФ
СО2
Цикл Кребса
ФГҚ
Хлоропласт
3-ФГҚ
Малат
Вакуоль
147.
148.
САМ- путь сходен с С4-путем фотосинтеза.Особенность: у САМ-растений реакции разделены
пространственно, у С4–растений – во времени.
1.В клетках имеется и ФЕП-карбоксилаза, и РБФкарбоксилаза/оксигеназа.
2. Синтезируемые днем в цикле Кальвина углеводы
необходимы для повторения цикла, являются
источником метаболитов (например, ФЕП).
3. САМ –путь приспособлен для максимальной
экономииводы, способствует прохождению
фотосинтеза в условиях засухи.
Но САМ-растения не являются высокоурожайными,
они ратсут медленно. Некоторые растения при
достаточном количестве воды могут переходить на
С3-путь фотосинтеза (хрустальная травка).
149.
1.Почему С3-путь фотосинтеза такназывается?
1. Из-за образования 3 молекул глюкозы
2. Состоит из 3 этапов
3.Первичный продукт 3фосфоглицериновая кислота является
трехуглеродным (триоза) 4. Из-за
присоединения СО2 к триозе
5 Из-за образования
фосфоенолпирувата
2. У САМ растений первичный акцептор
СО2
1.Оксалоацетат
2.Фосфоглицерат
3.Фосфоэнолпируват
4.Рибулозодифосфат
5.Эритрозофосфат
3. Где проходит присоединение СО2 у С3растений
1. В цитоплазме
2. В хлоропластах
3. В клетках обкладки
4. В строме хлоропластов
5. В митохондриях
4. Где проходит присоединение СО2 у С4растений?
1. В клетках обкладки
2.Вакуолях
3. В цитоплазме мезофилла
4. Хлоропластах мезофилла
5.В хлоропластах клеток обкладки
5. Первичный акцептор СО2 у С3-растений…
1.фосфоэнолпируват
2.рибулозо-1,5-дифосфат
3.фосфоглицерат
4.рибулозо-5-фосфат
5.рибозо-5-фосфат