


")








")









 Э.Ф.В.Пфлюгера")

")


")
")












")
")
")











")




")
 Биология
БиологияПохожие презентации:

")



")




Физиология возбудимых тканей. Подготовка к итогу №1
1. Подготовка к итогу №1 Физиология возбудимых тканей
2. 1. Понятие «биомембрана»
Биологическаямембрана – морфофункциональноеобразование,отграничивающеесодержимоеклеткиживыхорганизмовотвнешнейдлянеё
средыи жидкостныекомпартментывнутриклетки, имеющиеобщийплан
строенияи сходныефункции.
Понятие ввелв1963году Дж. Робертсон.
3. 2. Эволюция представлений о биомембране
2. Эволюция представленийобиомембранеРобертГук – 1665год.
Усовершенствовалмикроскоп
(установилклеточноестроение
тканей, исследуясрезпробкового
растения), ввелтермин«клетка».
КарлНегели– 1855год.
Ввелтермин«мембрана»для
объясненияплазмолиза
(гипертоническийраствор).
ДеФриз– 1877год.
Ввелпонятиеплазмолиз(отделение
протопластаотклеточнойстенкив
гипертоническомрастворе)
4.
5. 3. Жидкостно-мозаичная концептуальная модель биомембраны Сингера Николсона (1972 г.)
3. Жидкостно-мозаичнаяконцептуальнаямодельбиомембраныСингераНиколсона (1972г.)
В состав биологической мембраны входят липиды (в основном
фосфолипиды), белки (в основном гликопротеины) и углеводы (в
основном мукополисахариды).
Согласно этой модели мембрана представлена бислоем (двойным
слоем) фосфолипидов, ориентированных таким образом, что
гидрофобные хвосты (их два в каждой молекуле!) находятся внутри
бислоя, а гидрофильные головки направлены в водную фазу, т.е.
снаружи.
Белки бывают трех видов:
Интегральные (по топологической классификации – политопические) –
полностью пронизывают два слоя фосфолипидов и выполняют
функцию ионных каналов и насосов;
Полуинтегральные (по топологической классификации –
монотопические) – пронизывают только один слой фосфолипидов и
выполняют функцию белков-переносчиков и рецепторную;
Поверхностные или периферические (по топологической
классификации –монотопические) – лежат на поверхности
гидрофильных головок и участвуют в рецепторопосредованном
эндоцитозе, также выполняют ферментативную, сократительную
функции.
Углеводы представлены цепочками моносахаридов, в основном лежат
на поверхности мембраны и носят название гликокаликс,
участвующий в рецепторной функции биомембраны.
6. 4. Предметные модели биомембраны
4. Предметные моделибиомембраныМодели биологической мембраныможноподелитьна 2 вида:
1) Физические– исследованиепроводитсяс неживойприродой(вода,
жир).
а) плоская;
б) сферическая(липосома).
2) Биологические– исследованиепроводитсяна структурах, взятыхиз
живогоорганизма.
а) «тени» эритроцитов;
б) «гигантский» аксонкальмара.
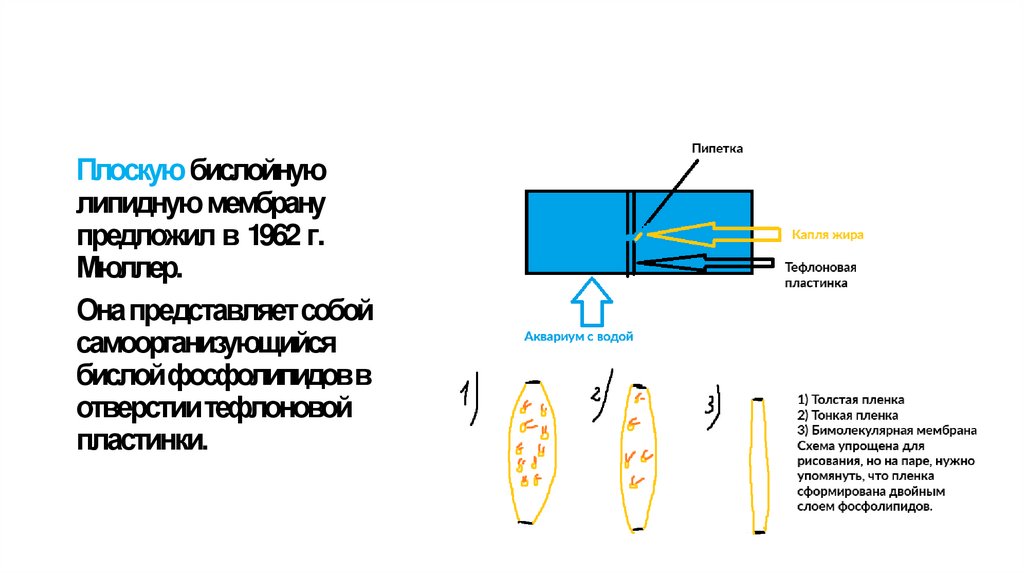
7.
Плоскую бислойнуюлипидную мембрану
предложил в 1962 г.
Мюллер.
Онапредставляетсобой
самоорганизующийся
бислойфосфолипидовв
отверстиитефлоновой
пластинки.
8. Сферическая модель мембраны – липосома.
Липосома представляет собой 2 слоя липидов,внутри вода, снаружи тоже вода. Главное не
путайте с мицеллой, которая не является
моделью мембраны (1 слой липидов), а любая
мембранаимеет2 слоялипидов.
Суть опыта сферической модели: взяли стакан
с водой и капнули жир. Затем посмотрели, как
располагаются хвостики и головки в липосоме,
тоестьстрогоупорядоченно.
Плоские модели мембраны помогают изучать
электрические свойства мембраны, её
проницаемость.
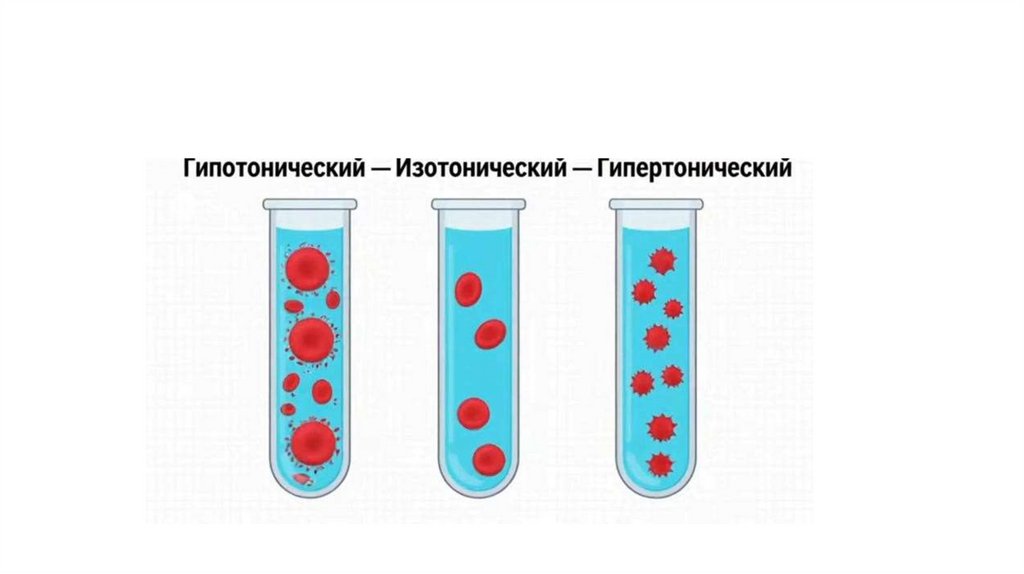
9.
«Тени»эритроцитов– берутклеткиэритроцитови опускаютихвгипотонический(менее0,9%NaCl)раствор, эритроцитынабухают
(явление гемолиза), мембранылопаются,содержимоевыходит,
остаютсятолькомембраны.Напомню,вгипертоническомрастворе–
водавыходитизклеткии эритроцитысморщиваются(явление
плазмолиза).
«Гигантский» аксонкальмараимеетдиаметроколо0,5 мм,чтов1001000разбольше,чемдиаметраксоначеловека. Являетсяоднимиз
модельныхобъектовдляизучениябиопотенциалов.
10. 8. Свойства и функции биомембраны
8. Свойства и функциибиомембраныМембранаимеет3 основныхсвойства:
1) Замкнутость– мембраназамкнута
гидрофильнымиголовками;
2) Текучесть– белки,находящиесяв
составемембраны,могутпереходить
изодногослояфосфолипидовв
другой, явлениеflip-flop
взаимодействия;
3) Ассиметричность– фосфолипиды
ассиметричны,снаружигидрофильные
головкибольше,чемвнутри
гидрофобныехвосты.
Функции мембраны:
1)
Отграничивающая(барьерная)– отделяет
внутреннююсредуклеткиотвнешней;
2)
Защитная– защищаетвнутреннеесодержимое
клетки;
3)
Рецепторная – реагированиена сигналывнешней
средыи передачаихотоднойклеткик другой;
4)
Каталитическая – изменениеактивности
ферментныхсистем;
5)
Компартментационная– разделениена отсеки
органеллклетки(двумембранные– митохондрии,
пластиды,ядро;одномембранные– ЭПС,аппаратГольджи,
вакуоли, лизосомы)
6)
Образованиемежклеточныхконтактов–
десмосома,замыкающий,адгезионный;
7)
Транспортная– движениеионовчерез мембрану.
11. 9. Подмембранные и надмембранные структуры
Подмембранныекомпоненты клетки—этоструктурыбелковойприроды,которыеобразуютцитоскелет—опоруклетки. К ним
относятсямикротрубочкии микрофиламенты.Цитоскелетвыполняет
опорнуюфункцию, способствуетзакреплениюорганеллв
определённомположениии ихперемещениювклетке.
Надмембранныеструктуры– периферическийбелоки гликокаликс.
Углеводыпредставленыцепочкамимоносахаридов, восновномлежат
на поверхностимембраныи носятназваниегликокаликс, участвующий
врецепторнойфункциибиомембраны.
12. 10. История изучения биоэлектрических явлений
10.История изучениябиоэлектрическихявлений
ПервыйопытЛ. Гальванис металлом(1786):
Пришелк выводу, что существует«живое
электричество» (биотоки), которое возникает
вспинноммозгеи передается к нимпо
металлическимпроводникам.
13. 10. История изучения биоэлектрических явлений
10.История изучениябиоэлектрическихявлений
ВторойопытЛ. Гальванис металлом(1797):
Вольтсказал, что выводхуйняи провелсериюэкспериментовс
разнороднымиметаллами, где доказал разность потенциаловмежду
медьюи железомс электролитом(ткань лягушки) – возникло
электричество. Создал«вольтовстолб» основанныйна этомпринципе.
Гальванинеостановилсяи провелопытбезметалла. Иобосновалдо
концасвоепредположение.
14. 10. История изучения биоэлектрических явлений
10.История изучениябиоэлектрическихявлений
ОпытМаттеуччиc вторичнымтетанусом(1838):
Окончательное доказательство существования
электрических явленийвживыхтканях былополученов
опыте«вторичного тетануса» Маттеуччи.
Тетанус - состояние длительногосокращения,
непрерывногонапряжениямышцы(опистотонус). Тетанус
возникает припоступлениик мышценервныхимпульсовс
такой частотой(более20 Гц), что расслаблениямежду
последовательнымиодиночнымисокращениями не
происходит.
Сутьопыта: берется2 нервно-мышечныхпрепарата
лягушки. Нерводногопрепаратанакладываетсяна мышцу
другого. Нерв1препаратастимулируетсятоком. Т.о. один
нервно-мышечныйпрепаратвозбуждалсятоком, а биотоки
сокращающейсямышцыраздражаютнерввторого нервномышечногопрепарата.
15. 11. Понятие «мембранный потенциал» (МПП)
11.Понятие «мембранныйпотенциал»(МПП)Мембранныйпотенциал(МП) – эторазность
потенциалов, существующуюмежду
цитоплазмой(φвн) и окружающимклетку
наружнымраствором(φнар). МП=φвн - φнар
В1943 ДавидГолдманвывелуравнение
мембранногопотенциалаоснованноена
формулеНернстаи учитывающее
концентрациювсехионов,а также
проницаемостьмембраныдляних.
RT PK[K]BH PNa[Na]BH PCl[Cl]H
EM ln
nF PK[K]H PNa[Na]H PCl[Cl]BH
16. 12. Регистрация ПП. Изменения ПП
12.РегистрацияПП. Изменения ПП1. Методповреждения- на ткани делаютнадрез, помещаютактивный
электрод. Методповреждения чащеиспользуется прирегистрации
потенциалапокояна макропрепарате – мышце,нерве. Неоченьточный,
т.к. содержимоеклетки и смешивается с окружающимраствором
2. Методвнутриклеточногоотведения - клетку помещаютв жидкость.
Активныйэлектрод-микроэлектрод-это тонкая серебрянная нить,
котораянаходится вмикро-пипетке, которуювводятвклетку, приэтом
мембранане повреждается. Приудачномвведении микроэлектрода
мембранаплотноохватываетегокончики клетка сохраняет
способность функционироватьв течение нескольких часов, не проявляя
признаковповреждения. Допрокола мембранымикроэлектродом
разностьпотенциаловмежду активными индифферентнымэлектродом
равнанулю. Kак толькомикроэлектродпрокалываетповерхностную
мембрануклетки, регистрируется разностьпотенциаловмежду
поверхностьюи содержимымклетки, равнаяпотенциалу покояклетки.
17. 13. Механизм формирования ПП
13.МеханизмформированияППЗначениепотенциала покояклетки определяется двумя
основнымифакторами:
1.
соотношениемконцентраций проникающихчерез
покоящуюсяповерхностнуюмембранукатионови
анионов
2.
соотношением проницаемостеймембраныдля
этих ионов.
Основныммеханизмомформированияпотенциалапокоя
являются:
1.
созданиеконцентрационнойасимметрии K+ при
работекалий-натриевогонасоса (калий-натриевой
АТФазы)
2.
выходK+изклеткипоградиенту концентрации
18. 14. Фазы ПД, возбудимость при возбуждении
14.ФазыПД, возбудимость привозбужденииПД– изменениеМПпривозбуждении.
19. 15. Ионные механизмы формирования ПД
15.Ионные механизмыформированияПДА.Потенциалпокоя– двухворотныйзакрытый
натриевыйканали одноворотныйзакрытый
калиевыйканал.
Б.Деполяризация– открытнатриевыйканал, натрий
поступаетвклетку, достигаетсначалКУД, затем
овершут(положительныймембранныйпотенциал).
В.Реполяризация– закрытиенатриевогоканала,
открытиекалиевого(восстановлениеисходного
отрицательногозначениямембранногопотенциала)
Г. Гиперполяризация– медленныйкалиевыйканал
неуспеваетзакрыться,поэтомумембранный
потенциалопускаетсянижепотенциала покоя.
20. 16. Понятия «раздражитель», «раздражение» в физиологии возбудимых тканей (Типы раздражителей, Электрический стимул при
16.Понятия«раздражитель», «раздражение» вфизиологиивозбудимыхтканей(Типыраздражителей, Электрический
стимулприэлектрофизиологическихисследованиях)
Раздражитель - фактор внешнейиливнутреннейпоотношениюк возбудимойструктуре
среды, которыйпридействииилиизменениидействия, способенвызватьвозбуждение.
Раздражение– (Раздражимость) – это способностьклеток, тканей, организма вцелом
переходить подвоздействиемфактороввнешнейиливнутреннейсредыиз состояния
физиологическогопокояв состояние активности.
Признаки, покоторымразличаются раздражители:
• Природе(модальность – вкус; температура, валентность– горький, кислый,сладкий,
соленый):физические, химическиеи т.п.
• Биологическому значению(адекватные – стимулкоторому требуется меньшая
энергиядлявозникновениявозбуждения, например– определенныерецепторы
сетчаткиглаза длявосприятиясвета, неадекватные– НАДАВИНАГЛАЗ).
Электрическийток является адекватнымраздражителем.
• Отношениюсилывоздействия к порогувозбуждения (подпороговые – не вызовут
возбуждение, пороговые– минстимулдлявозбуждения, сверхпороговые– болеечем
достаточныйдлявозбуждения).
• Одиночныеилисерийные
Возбуждениевозникает приаппликациикатодом(-) и инъекции анодом(+) за счет
разностипотенциаловвнеи внутриклетки.
21. 17. Законы раздражения: силы, времени, градиента
17.Законыраздражения:силы,времени,градиента
Законыраздраженияотвечаютна вопрос,каким
долженбытьраздражитель, чтобывозникло
возбуждение.
Раздражительможетвызватьвозбуждение,
еслиондостаточносильный(пороговымили
вышепорогового)(законсилы),длительный
(законвремени)ибыстронарастает(закон
градиента).
С– произошлааккомодация(клеткапривыклак
действиюстимула)
22. 18. Закон «силы-времени» Гоорвега-Вейса- Лапика
18.Закон«силы-времени» Гоорвега-ВейсаЛапика23. 19. Законы возбуждения: «всё или ничего», «силы»
19.Законывозбуждения:«всёилиничего»,«силы»
Законывозбужденияотвечаютна вопрос,каким
образомможетответитьвозбудимаяструктурана
действие раздражителя. Как клетка будет
возбуждаться.
Закон«всеилиничего»силаответнойреакции
возбудимойструктурыприпрочихравныхусловиях
даётмаксимальнуюответнуюреакцию(«всё») при
любойсиле пороговогоилисверхпорогового
раздраженияи недаётникакогоответа («ничего»)при
подпороговомраздражении(нервноеволокно,
мышечноеволокно,миокард.
Закон«силы»с увеличениемсилыстимула
увеличиваетсясилаответнойреакциивозбудимой
структуры(нерв,мышца).
24. 20. Действие постоянного подпорогового тока на возбудимые структуры
20. Действие постоянногоподпороговоготока на возбудимыеструктуры
Возбудимость определяется значением порога
раздражения:
ПР=КУД-МП
В 1859 г. немецкий физиолог Пфлюгер Э.Ф.В.
установил, что если на нерв воздействовать слабым
(подпороговым) постоянным током, то его возбудимость под катодом повышается, а под анодом
снижается.
В 1883 г. российский (пермский) физиолог Б.Ф.Вериго
назвал явления Пфлюгера временными. Если ток
действует достаточно долго, то под катодом возбудимость снижается, становясь меньше исходной
(в состоянии покоя), а под анодом может
повыситься.
25. 21. Замыкательно-размыкательные законы (полярный закон) Э.Ф.В.Пфлюгера
21.Замыкательно-размыкательныезаконы(полярныйзакон)Э.Ф.В.Пфлюгера
Катодно-замыкательное, анодно –
размыкательное возбуждение.
1859 г
Если с неповрежденным участком
соприкасается катод, возбуждение
возникало в момент замыкания
тока. Если же катод устанавливали
на поврежденном участке, а анод —
на неповрежденном, возбуждение
возникает только при размыкании
тока.
26. 22. Длительная сверхпороговая деполяризация возбудимых структур
22. Длительная сверхпороговаядеполяризациявозбудимыхструктур
Длительная пороговая деполяризация
преобразуется в возбудимых клетках в
ритмический разряд потенциалов
действия (генерирование ритмических
потенциалов действия (импульсов);
повторные (повторяющимся) ответы
при одном, но длительном стимуле .
При сверхпороговой деполяризации
частота ПД увеличивается, но при
длительном раздражении возникает
адаптация (частота ПД возвращается
частоте пороговой деполяризации).
27. 23. Функциональная подвижность возбудимых структур (лабильноть)
23. Функциональная подвижностьвозбудимыхструктур(лабильноть)
Н.Е.Введенский на нервно-мышечном
препарате установил, что нерв,
мионевральные синапсы и мышца по-разному
реагируют на сверхпороговые раздражения
различной частоты. Отсюда был сделан вывод,
что разные структуры имеют неодинаковую
функциональную подвижность. Синонимом
термина «функциональная подвижность»
является «лабильность».
Лабильность – скорость возникновения
возбуждения и определяется длительностью
рефрактерных периодов (время, когда клетка
не возбуждена).
28. 24. Парабиоз Н.Е.Введенского
Парабиоз – общая, универсальнаяреакция возбудимых тканей на
сильное или длительное
воздействие. Суть парабиоза в
том, что под влиянием
раздражителя в возбудимых
тканях изменяются их
физиологические свойства, в
первую очередь резко снижается
лабильность.
29. 25. Описание процессов распространения автоволн. Тау-модель распространения возбуждения.
25. Описаниепроцессовраспространенияавтоволн. Тау-модельраспространения
возбуждения.
Автоволны–
cамоподдерживающиесяволныв
активнойсреде, сохраняющиесвои
характеристикипостояннымиза счет
распределенныхвсреде источников
энергии.
Декрементпроведения–
постепенноеослабление
возбуждения(затуханиеволны
деполяризации)помереего
распространенияповозбудимой
структуре(КАМЕНЬВОЗЕРО).
Автоволнынезатухают
(бездекрементно).
30. 26. Циркуляция возбуждения в замкнутых возбудимых структурах (кольце)
26. Циркуляция возбуждениявзамкнутыхвозбудимыхструктурах(кольце)
31. 27. Повторный вход возбуждения (re-entry)
27. Повторныйвходвозбуждения(re-entry)32. 28. Распространение электротона
28. РаспространениеэлектротонаЭлектротон – возникает в клетках не генерирующие ПД
(глия, Пуркинье мозжечка), и распространение ПД идет, как
электрическое возбуждение, т.е. декрементно. Причем,
чем больше расстояние, тем больше затухание.
33. 29. Распространение возбуждения в нервных волокнах
29. Распространениевозбуждениявнервныхволокнах
Работают по закону «все или ничего».
34. 30. Явление перескока при проведении возбуждения в нервных волокнах
30. Явление перескокаприпроведениивозбуждениявнервныхволокнах
• «Перепрыгивание» потенциала действия через
межперехватный участок возможно потому, что
амплитуда ПД в 5-6 раз превышает величину,
необходимую для возбуждения соседнего
перехвата. ПД может «перепрыгивать» не только
через один, но и через два—четыре
межперехватных промежутка. Это явление может
наблюдаться, например, при снижении
возбудимости соседнего перехвата под
действием какого-либо фармакологического
вещества (например, новокаина).
• Для определения возможности перескока
используют понятие гарантийный фактор –
отношение АПД раздражающего участка к порогу
раздражения раздражаемого участка. Эффект
перескока возникнет при значении гарантийного
фактора 5-7.
35. 31. Сальтаторное проведение потенциалов действия
31.Сальтаторное проведениепотенциаловдействия
В перехватах Ранвье происходят весьма большие траты
энергии вследствие большой плотности здесь натрийкалиевых каналов.
36. 32. Классификация нервных волокон по Эрлангеру-Гассеру
32. КлассификациянервныхволоконпоЭрлангеру-Гассеру
37. 33. Законы проведения возбуждения в нервных волокнах
33. Законыпроведениявозбуждениявнервныхволокнах
• Возбуждение может
распространяться по нервному
волокну только при его
морфологической и функциональной
целостности
38. 34. Классификация синапсов (по характеру контактирующих клеток, по способу передачи сигнала, по конечному эффекту, по форме
34. Классификациясинапсов(похарактеруконтактирующих клеток, поспособупередачисигнала,
поконечномуэффекту,поформеконтакта, поразвитию
вонтогенезе).
39. 35. Типы межнейрональных синапсов
35. Типымежнейрональных синапсов• По характеру контактирующих частей нейронов
• аксо-аксональные,
• аксо-дендритические (дендритные),
• аксо-соматические,
• дендро-дендритические,
• дендро-соматические
• По локализации в нервной системе
• центральные (расположены в головном и спинном мозге, т.е. в ЦНС)
• периферические (в периферической нервной системе,
нервномышечный, нейросекреторный синапс вегетативной нервной
системы)
40. 36. Химический синапс: общий план строения. Типы химических синапсов
36. Химическийсинапс:общийпланстроения.Типыхимическихсинапсов
41. 37. Этапы передачи сигнала в химическом
1.2.
3.
4.
5.
6.
7.
37. Этапыпередачисигналавхимическом
синапсе
Деполяризация
предсинаптической
мембраны
приходящим по аксону потенциалом действия (ПД).
Открытие потенциалзависимых Сa++-каналов на
предсинаптической мембране и поступление Сa++ в
предсинапс (пассивный транспорт).
Увеличение концентрации Сa++ в предсинапсе
запускает слияние синаптических пузырьков с
мембраной
и
выброс
предсинаптической
нейромедиатора в синаптическую щель (экзоцитоз).
Диффузия медиатора к постсинаптической мембране.
Взаимодействие
медиатора
с
рецепторами
постсинаптической мембраны.
Реакция постсинапса на действие медиатора.
Инактивация медиатора.
Первый механизм - типичный экзоцитоз: полное слияние везикулы и встраивание
ее мембраны в пресинаптическую. Все содержимое везикулы оказывается в
синаптической щели.
Второй механизм - экзоцитоз без полного слияния, с частичным освобождением
(шутливо названный "кратковременным поцелуем"). Формируется временная
пора (канал) между внутренностью везикулы и окружающей средой. Через
образованную пору по градиенту концентрации медиатор диффундирует в
синаптическую щель. Количество выделенного медиатора (величина кванта)
зависит от времени, в течение которого пора находится в открытом состоянии.
Третий механизм -специфический белок (медиатофор) или канал, встроенный в
пресинаптическую мембрану, освобождает медиатор из цитоплазмы нервного
окончания.
Везикулы выполняют не связанную с экзоцитозом функцию (содержат
резервный медиатор, который при необходимости поступает в цитоплазму
нервного окончания).
42. 38. Медиаторы химического синапса
38. МедиаторыхимическогосинапсаМедиаторы– биологически активные вещества, выделяемые пресинаптической мембранойи
являющиеся посредником в процессе синаптической передачи возбуждения.
ОттоЛёви установил химическуюприроду передачивозбуждения через синапсыи рольв ней
ацетилхолина.
В1921году ОттоЛёвипровёл эксперимент с сердцамилягушек, чтобыпродемонстрировать
химическуюприроду передачинервных импульсов.
Ходопыта:
Лёвиизвлёк сердца у двух живыхлягушек. Водномизсердецонвыделилблуждающий нерв,
другое оставилнетронутым.
Обасердца учёный омывалфизиологическимрастворомРингера.
Блуждающий нервпервогосердца Лёви возбуждал электрическимтокомдлязамедления
сердечных сокращений, послечегос помощьюпипетки собираломывающуюего жидкость.
Добавление этойжидкости в питательныйраствор, чтоомывалвторое (невозбуждённое) сердце,
приводилок замедлениюего ритма без воздействия электрического тока.
Лёви высказал предположение, чтовжидкости, омывающейпервое сердце, присутствует некое
вещество, которое способно передаватьсигналотблуждающего нерва сердечной мышце. Такое
«замедляющее» вещество Лёвиназвал«vagustoff» (cубстанцией блуждающего нерва), или
парасимпатином.
ЗатемЛёви произвёлпохожийэксперимент. Онвозбуждал уже другой, симпатическийнерв
сердца, в результате чего сердечныйритмускорялся. Перенесённая оттакогосердца
омывающая жидкость вызывала ускорение ритма другого сердца. Обнаруженное
«ускорительное» вещество Лёвиназвалсимпатином.
43. 39. Комедиаторы, модуляторы, агонисты, антагонисты
КомедиаторыВ одном и том же синапсе возможно выделение более чем одного медиатора.
Причём и действие первого (М1) и второго (М2) вызывает эффект в постсинапсе.
Наиболее частая совместная локализация медиаторов:
Ацетилхолин + гамма-аминомасляная кислота ГАМК (мотонейроны спинного мозга, амакриновые клетки сетчатки, нейроны
нижней оливы).
ГАМК + глицин (нейроны спинного мозга и мозжечка).
ГАМК + глутамат (нейроны гиппокампа, зубчатой фасции, пирамидные нейроны).
Модуляторы
Влияют на интенсивность и продолжительность действия классических медиаторов.
Самостоятельно непосредственно не меняют проводимость синаптических мембран и выделяются, как правило, вместе
с медиатором.
Модуляторы не осуществляют синаптическую передачу, но могут значительно усиливать как синаптическую передачу,
так и возбудимость нервных клеток.
Агонисты (ацетилхолин и мускарин)
Каждый рецептор постсинаптической мембраны взаимодействует со своим специфическим медиатором.
Однако такая специфичность не абсолютна – практически все рецепторы способны связываться с другими веществами.
Такие вещества для медиаторов являются АГОНИСТАМИ (полными или частичными).
Полные а. – полностью воспроизводят эффекты медиатора.
Частичные а. – частично воспроизводят эффекты медиатора.
Антагонисты (атропин, стрихнин антагонист глициновых рецепторов)
Если вещество, связываясь с рецепторами не вызывает возбуждения, а при этом мешает действию медиатора, то его
называют АНТАГОНИСТОМ.
44. 40. Ионотропные рецепторы (ПСП)
40. Ионотропные рецепторы(ПСП)Рецептор на поверхности ионного канала.
Постсинаптические потенциалы ВПСП (вызывает
деполяризацию) – ацетилхолин, норадреналин,
дофамин, серотонин. ТПСП (гиперполяризация)глицин, ГАМК
45. 41. Метаботропные синапсы (структура G- белка)
41.Метаботропные синапсы(структураGбелка)Е – АДЕНИЛАТЦИКЛАЗА
ВП - цАМФ
46. 42. Нервно-мышечный синапс скелетного мышечного волокна (этапы передачи сигнала)
42. Нервно-мышечныйсинапсскелетногомышечного волокна(этапыпередачисигнала)
Этапы передачи возбуждения в НМС
Деполяризация предсинаптической мембраны пришедшим по аксону потенциалом
действия (ПД).
Открытие потенциалзависимых Сa++-каналов на предсинаптической мембране и
поступление Сa++ в предсинапс (пассивный транспорт).
Выход в синаптическую щель ацетилхолина путем экзоцитоза.
Диффузия медиатора к постсинаптической мембране.
Взаимодействие ацетилхолина с Н-холинорецепторами постсинаптической мембраны
мышечного волокна.
Открытие никотиновых рецепторных каналов постсинаптических каналов, пассивный
вход Na+ в мышечное волокно.
Образование
потенциала
концевой пластинки
- ПКП
(возбуждающего
постсинаптического потенциала - ВПСП) в области постсинаптической мембраны.
Электротоническое распространение ПКП в околосинаптическую область.
Формирование ПД мышечного волокна на сарколемме околосинаптической области.
47. 43. Электрические синапсы. Смешанные синапсы.
43. Электрические синапсы.Смешанныесинапсы.
• Электрический синапс — место более близкого прилегания пары
клеток, где их мембраны соединяются с помощью особых
белковых образований — коннексонов (щелевой контакт).
• Смешанный синапс — это синапс, сочетающий элементы как
химической, так и электрической передачи.
48. 44. Типы мышечной ткани: морфо- функциональные различия
44. Типымышечнойткани: морфофункциональныеразличияРазличают 2 типа тканей: поперечнополосатая (скелетная и сердечная) и гладкая (сосуды, внутренние органы и кожа).
Структурно-функциональными единицами для:
• Скелетной мышцы – мышечное волокно (симпласт – крупные многоядерные образования, появляющиеся, как результат
слияния большого количества клеток), которые образуют моторную единицу – группа мышечных волокон и
иннервирующий их мотонейрон (передние рога спинного мозга – альфа-мотонейрон);
• Сердечной мышцы – кардиомиоциты, соединенные друг с другом при помощи вставочных дисков, образуют клеточный
комплекс синцитий или функциональное волокно;
• Гладкой мышцы – гладкий миоцит (клетка).
• Сократительный аппарат для скелетной – длинные миофибриллы; сердечной – короткие миофибриллы; гладкой –
миофиламенты.
• Адекватный раздражитель (вид раздражителя, характерный для данной возбудимой структуры и требующий
минимальной затраты энергии):
• скелетная – ацетилхолин (возбуждающий постсинаптический потенциал – ВПСП);
• сердечная – диастолическая деполяризация клеток пейсмейкера (атипичные кардиомиоциты в проводящей системе
сердца, способные самостоятельно генерировать импульс) –> ПД соседнего волокна –> через нексусы возбуждение всех
кардиомицитов;
• гладкая – ацетилхолин или норадреналин (ВПСП), либо гуморальные факторы (адреналин, ангиотензин), либо
механическое растяжение.
49.
45. Физическиеи физиологическиесвойствамышц.Скелетнаямышца:иерархияструктурныхсократительныхкомпонентов
50. 46. Структурная организация миофибриллы. Саркомер.
46. Структурная организациямиофибриллы.Саркомер.
51. 47. Классификация скелетных мышечных волокон и мышц
47. Классификацияскелетныхмышечныхволокони мышц
Делятся по расположению на экстрафузальные (мышечные волокна образуют
основную массу мышцы и выполняют всю работу, необходимую для движения и
поддержания позы) и интрафузальные (прикрепляются нервные окончания, которые
передают информацию в ЦНС сокращена или расслаблена мышца в данный момент
времени).
По характеру сокращения: тонические и фазические.
• Тонические находятся постоянно в тонусе, имеют низкий потенциал покоя,
подчиняются закону силы раздражения, имеют несколько синаптических контактов
с несколькими аксонами разных мотонейронов. Для возникновения ПД
необходимо несколько импульсов. Например, наружные мышцы глаза.
• Фазические: белые (за счет отсутствия миоглобина, быстрые, легко утомляются,
большая сила сокращения, гликолиз), красные (за счет миоглобина медленные,
устойчивые к утомлению, небольшая сила сокращения, окисление) и
промежуточные (быстрые, устойчивые к утомлению, с большой силой, окисление и
гликолиз).
52. 48. Структурно-функциональная организация скелетной мышцы. Механизм мышечного сокращения и расслабления
48. Структурно-функциональнаяорганизацияскелетноймышцы. Механизммышечного
сокращения и расслабления
В структурно-функциональную организацию входит двигательная единица = мотонейрон + группа иннервируемых им
мышечных волокон. Мотонейронный пул мышцы: группа мотонейронов, иннервирующих данную мышцу
Чем больше точных движений, тем меньше волокон в ДЕ, в то время как в мышцах, участвующих в поддержании позы,
двигательные единицы включают несколько сотен и даже тысяч мышечных волокон.
Механизм мышечного сокращения объясняется моделью скользящих нитей, авторами которой принято считатьХ.Хаксли и
Дж.Хансона (1954 г.).
Теория скольжения нитей – во время сокращения мышцы длина толстых и тонких миофиламентов не изменяется, происходит
их скольжение друг относительно друга.
Механизм мышечного сокращения:
Мышечное волокно составляет триада (две цистерны и одна Т-трубочка), при возникновении нервного импульса
1.
возникает деполяризация частей саркоплазматического ретикулума;
2.
Потенциал действия приводит к открытию кальциевых каналов;
3.
Кальций связывается с тропонином C, который входит в состав актина и меняет конформацию тропомиозина
(блокиратор актомиозинового комплекса);
После этого происходит связывание головок миозина к актиновым центрам;
После связывания под воздействием молекул АТФ происходит открепление головки миозина от актина и изменение
конформации;
После гидролиза АТФ на АДФ и Ф произошло связывание миозиновой головки с актиновым центром и после
отсоединения Ф происходит протягивание (как в канате) актина, прикрепленного к Z-линии. Z-линия становится ближе к
миозиновому комплексу, вследствие чего исчезает I-диск и H-зона. Несколько таких процессов обусловливают
сокращение мышцы.
4.
5.
6.
53. 49. Электромеханическое сопряжение
54. 50. Рабочий цикл миозиновых мостиков поперечнополосатого миоцита, гладкого миоцита
50. Рабочийциклмиозиновыхмостиковпоперечнополосатогомиоцита, гладкогомиоцита
Фермент-киназа легкого миозина
55. 51. Расслабление. Энергетика мышечного сокращения.
51.Расслабление.Энергетика мышечногосокращения.
Расслабление поперечно-полосатого миоцита:
При уменьшении концентрации кальция в саркоплазме тропонин освобождается от него и восстанавливает
прежнюю конформацию молекулы.
Тропомиозин возвращается на прежнее место и уже не позволяет большинству головок миозина
прикрепляться к актиновой нити.
Когда большинство головок миозина начинает отделяться от актиновых нитей, наступает расслабление
мышечного волокна.
Расслабление гладкого миоцита:
Cа2+АТФаза саркоплазматического ретикулума закачивает Cа2+ из саркоплазмы в цистерны ретикулума
Cа2+ связывается с кальсеквестрином.
При низкой саркоплазматической концентрации Cа2+ тропомиозин закрывает миозинсвязывающие участки
и препятствует их взаимодействию с миозином.
Единственным прямым (непосредственным) источником энергии для мышечного сокращения служит
аденозинтрифосфат (АТФ).
Ресинтез АТФ происходит 2 основными путями:
анаэробным
аэробным
Химические (энергетические) системы:
фосфогенная, или АТФ-КФ-система; (КФ – креатинфосфат)
лактацидная, или гликолитическая;
кислородная, или окислительная.
Cистемы синтеза и ресинтеза АТФ при мышечном сокращении: 1 – фосфогенная, 2 –
гликолитическая, 3 – окислительная.
56. 52. Типы мышечного сокращения
52. Типымышечного сокращенияРазличают два основных типа мышечных
сокращений
• Если оба конца мышцы неподвижно
закреплены, происходит изометрическое
сокращение, и при неизменной длине
напряжение увеличивается.
• Если один конец мышцы свободен, то в процессе
сокращения длина мышцы уменьшится,
происходит изотоническое сокращение, и при
укорочении напряжение не изменяется.
Ауксотонический тип мышечных сокращений
• смешанный режим
• характеризуется изменением длины и тонуса
мышцы, при сокращении которой происходит
перемещение груза.
57. 53. Режимы мышечного сокращения
53. Режимымышечного сокращения• Одиночное
• Тетаническое (зубчатый тетанус, гладкий тетанус)
58. 54. Одиночные сокращения: фазы (тетанус)
54. Одиночныесокращения: фазы(тетанус)• Зубчатый тетанус возникает, если
вторая фаза каждого следующего
сокращения начинается тогда, когда
продолжается третья фаза
предыдущего сокращения.
• Сплошной или гладкий тетанус
возникает, если вторая фаза каждого
следующего сокращения начинается
тогда, когда продолжается вторая фаза
предыдущего сокращения.
59. 55. Оптимум и пессимум частоты раздражения скелетной мышцы
55. Оптимуми пессимумчастотыраздраженияскелетноймышцы
Частоты раздражений, при
которых достигаются
максимальные частоты ответов
называются оптимальными
частотами.
Частоты раздражений выше
оптимальных, при которых
снижаются частоты ответов
называются пессимальными
частотами.
60. 56. Сила и работа мышц
56. Сила и работамышцСила – это способность за счет мышечного сокращения
преодолевать внешнее сопротивление.
61. 57. Закон средних нагрузок
57. Законсреднихнагрузок• Мышца совершает максимальную работу при нагрузках
средних значений.
62. 58. Кривая изометрических максимумов
58. Криваяизометрических максимумов• Кривая изометрических максимумов - это график зависимости
максимумов изометрического сокращения мышцы или
саркомера от длины, при которой они измерялись.
• Изометрическое сокращение - развитие силы (напряжения) при
постоянной длине мышцы.
Характерный максимум при изометрическом сокращении одиночного мышечного волокна — при длине саркомера 2,0–2,2 мкм (примерно соответствует
длине мышцы в состоянии покоя). При уменьшении длины саркомера сила снижается, поскольку направленные навстречу друг другу актиновые филаменты
двух его половин начинают перекрываться, а толстые миозиновые филаменты упираются в Z-пластинки.
Кривая помогает определить соотношение между активной сократительной силой и длиной мышцы - для этого из максимумов вычитают пассивные
напряжения.
63. 59. Оценка функционального состояния мышечной системы у человека (Динамометрия, Электромиография, Стабилография, Эргометрия)
59. Оценкафункциональногосостояниямышечнойсистемыу человека(Динамометрия,
Электромиография, Стабилография,Эргометрия)