

























 Биология
БиологияПохожие презентации:










Физиология возбудимых тканей
1.
ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ1. Понятие о раздражимости и возбудимости живых структур
2. Понятие о мембранном потенциале покоя и его природе
3. Типы электрического ответа возбудимых структур. Природа
потенциала действия
4. Основные электрофизиологические параметры возбудимых
структур
2.
ПОНЯТИЕ О РАЗДРАЖИМОСТИ И ВОЗБУДИМОСТИЖИВЫХ СТРУКТУР
3.
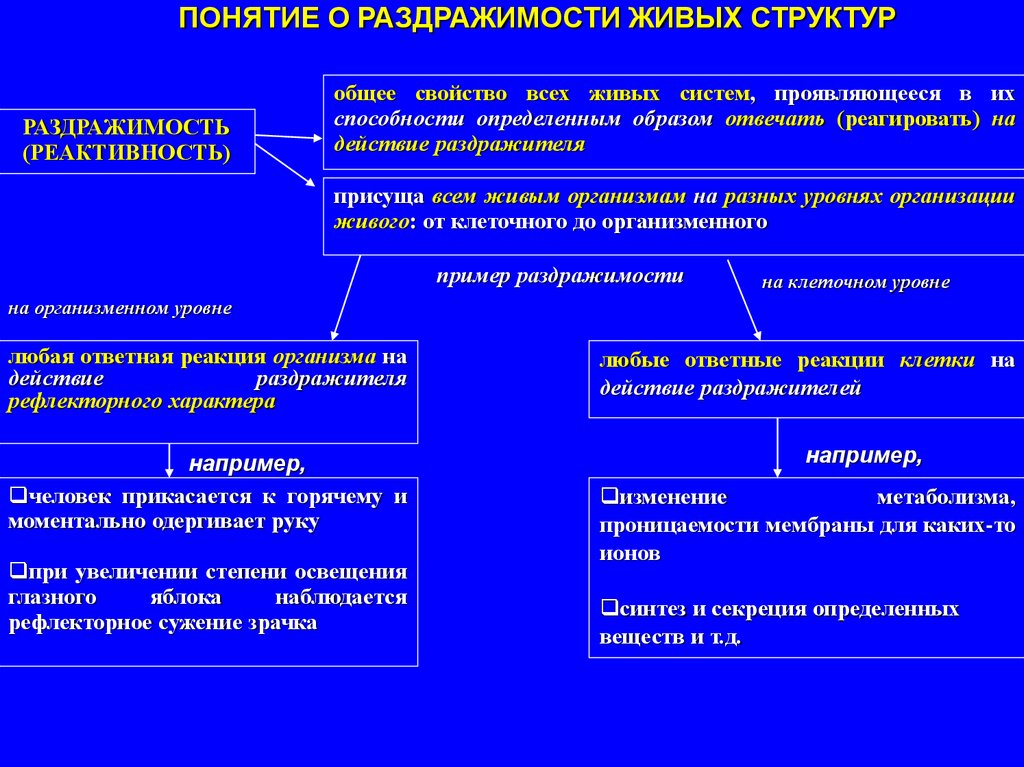
ПОНЯТИЕ О РАЗДРАЖИМОСТИ ЖИВЫХ СТРУКТУРРАЗДРАЖИМОСТЬ
(РЕАКТИВНОСТЬ)
общее свойство всех живых систем, проявляющееся в их
способности определенным образом отвечать (реагировать) на
действие раздражителя
присуща всем живым организмам на разных уровнях организации
живого: от клеточного до организменного
пример раздражимости
на клеточном уровне
на организменном уровне
любая ответная реакция организма на
действие
раздражителя
рефлекторного характера
например,
человек прикасается к горячему и
моментально одергивает руку
при увеличении степени освещения
глазного
яблока
наблюдается
рефлекторное сужение зрачка
любые ответные реакции клетки на
действие раздражителей
например,
изменение
метаболизма,
проницаемости мембраны для каких-то
ионов
синтез и секреция определенных
веществ и т.д.
4.
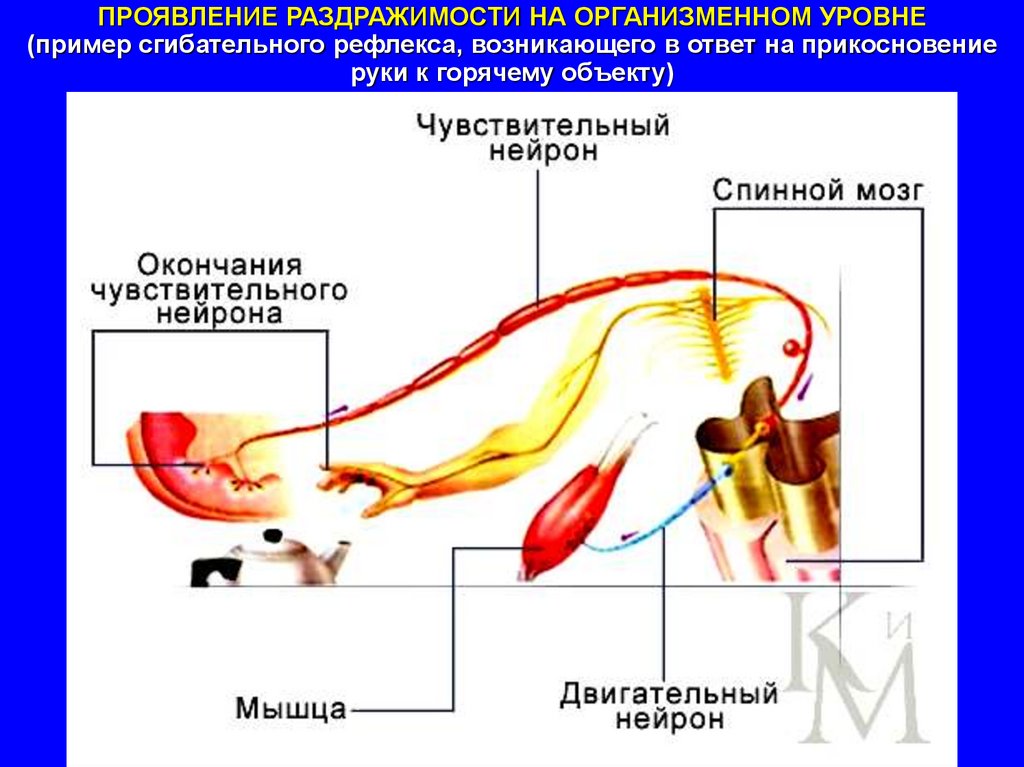
ПРОЯВЛЕНИЕ РАЗДРАЖИМОСТИ НА ОРГАНИЗМЕННОМ УРОВНЕ(пример сгибательного рефлекса, возникающего в ответ на прикосновение
руки к горячему объекту)
5.
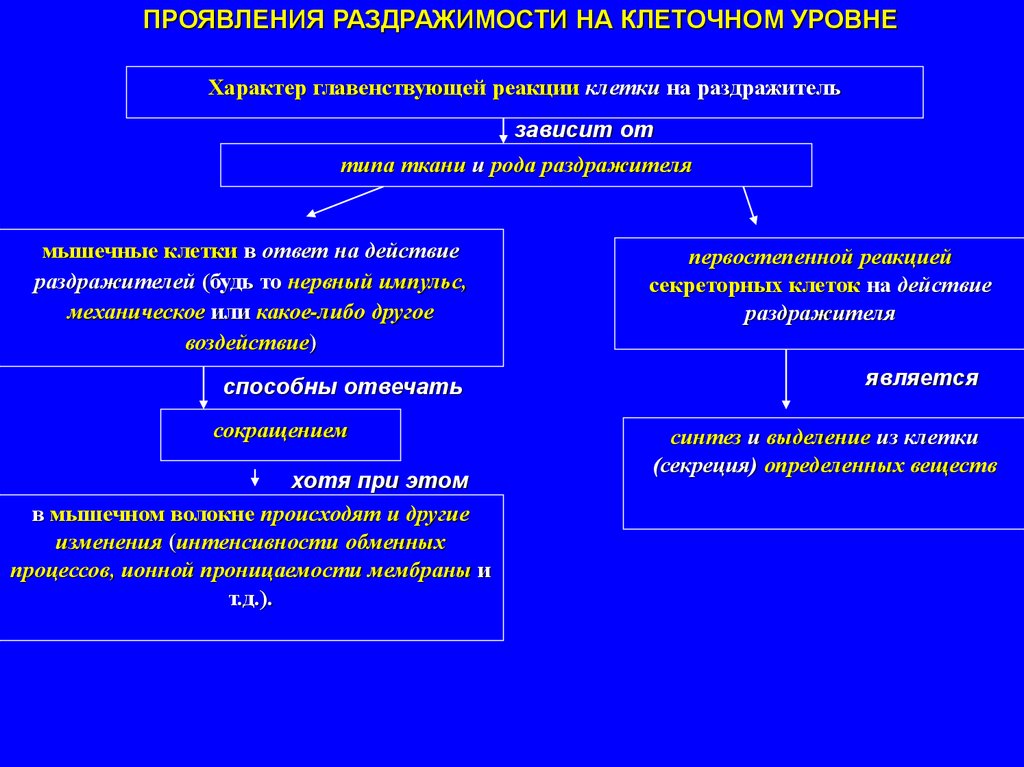
ПРОЯВЛЕНИЯ РАЗДРАЖИМОСТИ НА КЛЕТОЧНОМ УРОВНЕХарактер главенствующей реакции клетки на раздражитель
зависит от
типа ткани и рода раздражителя
мышечные клетки в ответ на действие
раздражителей (будь то нервный импульс,
механическое или какое-либо другое
воздействие)
первостепенной реакцией
секреторных клеток на действие
раздражителя
способны отвечать
является
сокращением
хотя при этом
в мышечном волокне происходят и другие
изменения (интенсивности обменных
процессов, ионной проницаемости мембраны и
т.д.).
синтез и выделение из клетки
(секреция) определенных веществ
6.
ПРОЯВЛЕНИЕ РАЗДРАЖИМОСТИ НА КЛЕТОЧНОМ УРОВНЕ(на примере активированного фагоцита)
7.
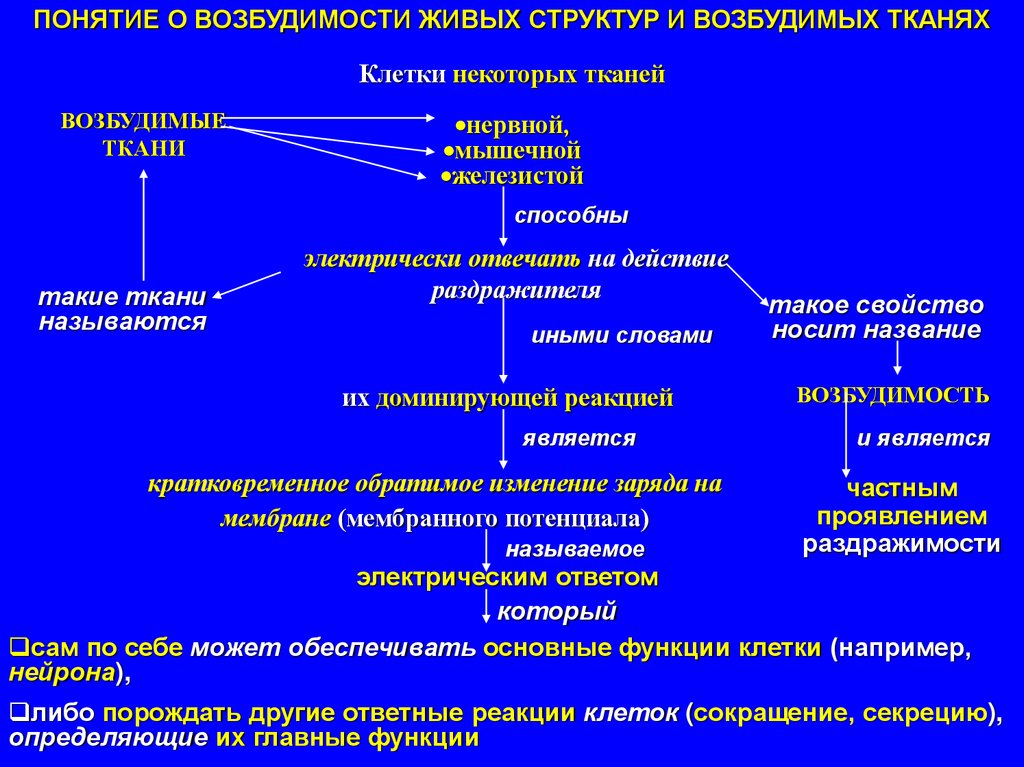
ПОНЯТИЕ О ВОЗБУДИМОСТИ ЖИВЫХ СТРУКТУР И ВОЗБУДИМЫХ ТКАНЯХКлетки некоторых тканей
ВОЗБУДИМЫЕ
ТКАНИ
нервной,
мышечной
железистой
способны
такие ткани
называются
электрически отвечать на действие
раздражителя
иными словами
их доминирующей реакцией
является
кратковременное обратимое изменение заряда на
мембране (мембранного потенциала)
называемое
такое свойство
носит название
ВОЗБУДИМОСТЬ
и является
частным
проявлением
раздражимости
электрическим ответом
который
сам по себе может обеспечивать основные функции клетки (например,
нейрона),
либо порождать другие ответные реакции клеток (сокращение, секрецию),
определяющие их главные функции
8.
ГИСТОСТРУКТУРА ВОЗБУДИМЫХ ТКАНЕЙнервные клетки
мышечная ткань
железистый
эпителий
9.
ПОНЯТИЕ О МЕМБРАННОМ ПОТЕНЦИАЛЕ ПОКОЯ,ЕГО ПРИРОДА
10.
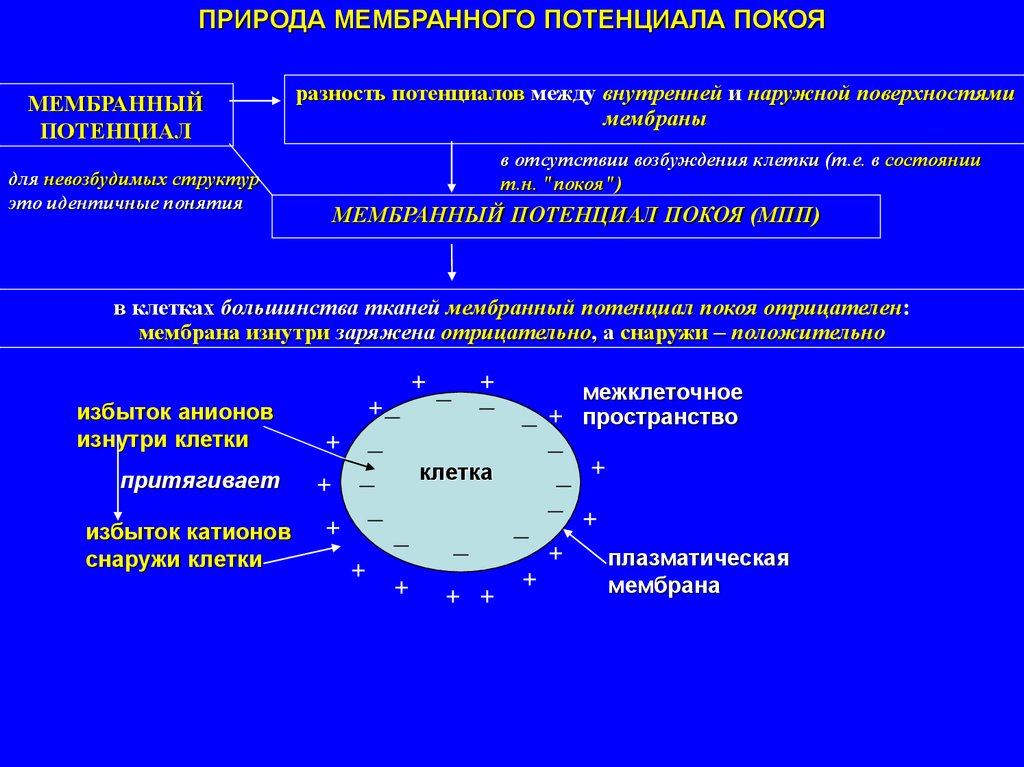
ПРИРОДА МЕМБРАННОГО ПОТЕНЦИАЛА ПОКОЯМЕМБРАННЫЙ
ПОТЕНЦИАЛ
для невозбудимых структур
это идентичные понятия
разность потенциалов между внутренней и наружной поверхностями
мембраны
в отсутствии возбуждения клетки (т.е. в состоянии
т.н. "покоя")
МЕМБРАННЫЙ ПОТЕНЦИАЛ ПОКОЯ (МПП)
в клетках большинства тканей мембранный потенциал покоя отрицателен:
мембрана изнутри заряжена отрицательно, а снаружи – положительно
+ _
+
_
межклеточное
+_
_ + пространство
+ _
_
клетка
_ +
притягивает
+ _
_
_
+
_
избыток катионов +
_
_
+
плазматическая
снаружи клетки
+
+
мембрана
+ + +
избыток анионов
изнутри клетки
11.
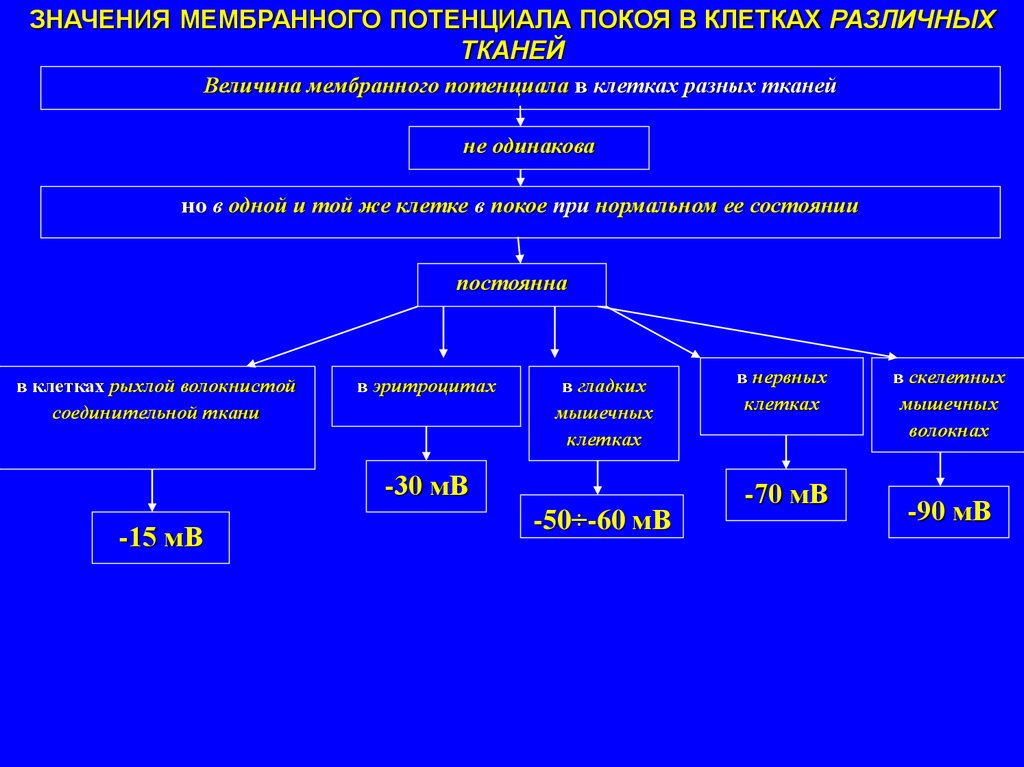
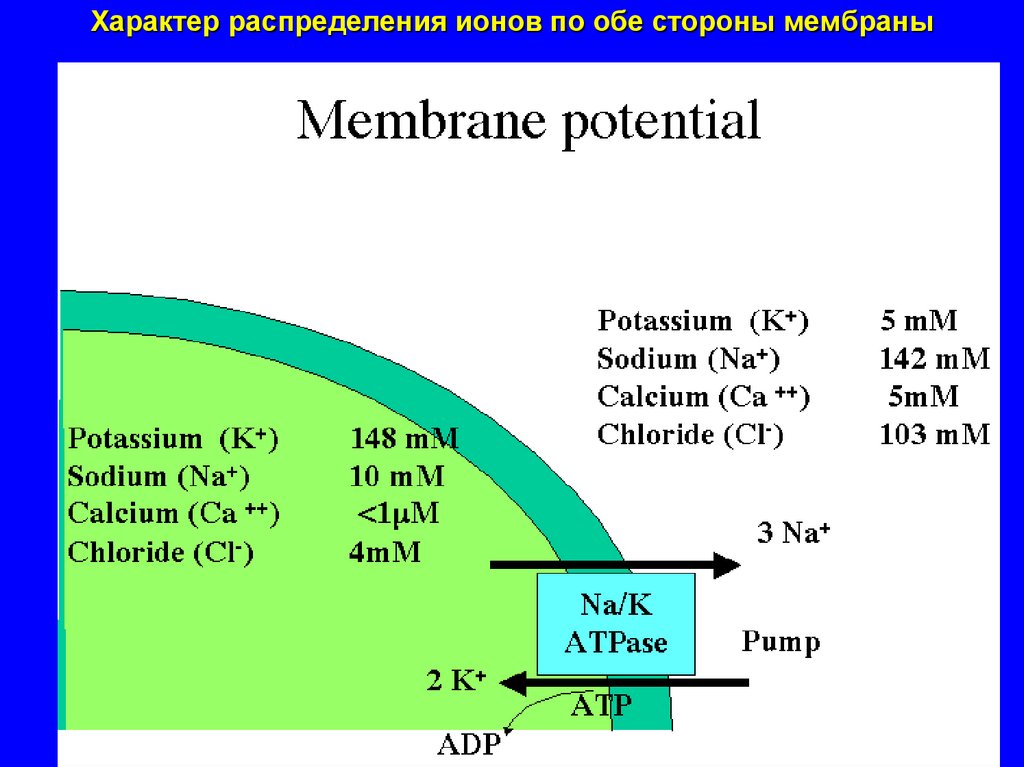
ЗНАЧЕНИЯ МЕМБРАННОГО ПОТЕНЦИАЛА ПОКОЯ В КЛЕТКАХ РАЗЛИЧНЫХТКАНЕЙ
Величина мембранного потенциала в клетках разных тканей
не одинакова
но в одной и той же клетке в покое при нормальном ее состоянии
постоянна
в клетках рыхлой волокнистой
соединительной ткани
в эритроцитах
в гладких
мышечных
клетках
-30 мВ
-15 мВ
-50÷-60 мВ
в нервных
клетках
-70 мВ
в скелетных
мышечных
волокнах
-90 мВ
12.
ПРИРОДА МЕМБРАННОГО ПОТЕНЦИАЛАсогласно мембранно-ионной теории Бернштейна-Ходжкина
ПРИЧИНЫ ВОЗНИКНОВЕНИЯ мембранного потенциала
1. ассиметричное распределение
ионов по обе стороны мембраны
2. неодинаковая проницаемость
мембраны для различных ионов
иными словами
полупроницаемость или
неодинаковая концентрация определенных ионов
избирательная проницаемость
по обе стороны мембраны (т.е. существование
мембраны
трансмембранных градиентов для определенных
ионов)
в большинстве клеток
ионов калия больше
внутри, чем
снаружи клетки
например,
ионов натрия,
кальция, хлора
больше снаружи, чем
внутри клетки
что обусловлено
деятельностью ионных насосов
систем активного ионного транспорта,
которые транспортируют ионы против
концентрационных градиентов
из области с меньшей концентрацией в
область с большей концентрацией
а, следовательно,
с затратой энергии в форме АТФ
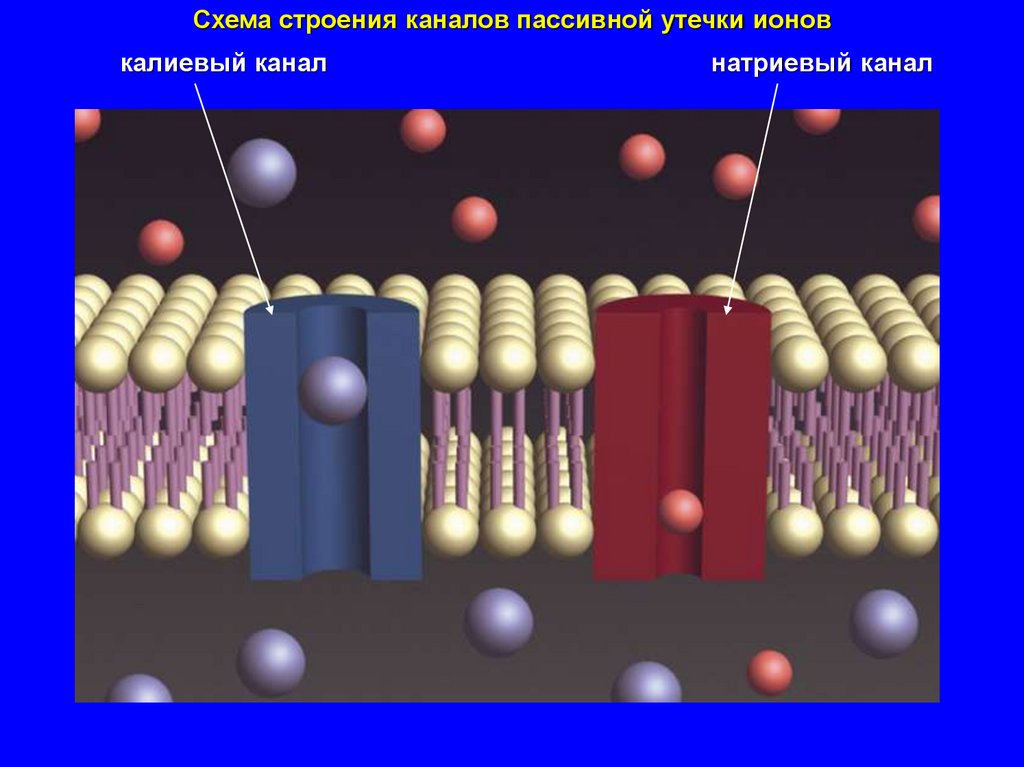
мембраны большинства
клеток в покое
хорошо
относительно
проницаемы плохо проницаемы
для калия
для органических
анионов, натрия,
кальция
13.
Схематическое представление природы мембранного потенциала покоя1. Ионы по обе стороны мембраны распределены асимметрично
что создает предпосылку для
пассивного движения ионов через мембрану по концентрационному
градиенту
межклеточное вещество
клетка
К+> 40-50 раз
Na+> 7-10 раз
Cl->
Ca2+>
14.
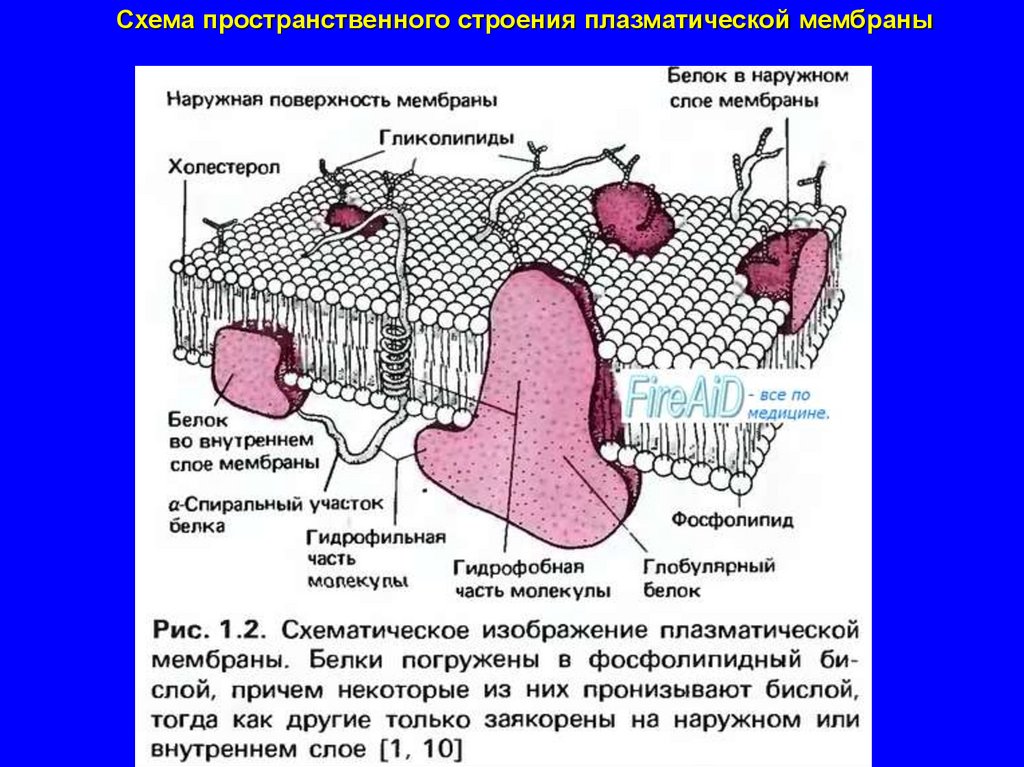
Схема пространственного строения плазматической мембраны15.
Характер распределения ионов по обе стороны мембраны16.
Схема строения каналов пассивной утечки ионовкалиевый канал
натриевый канал
17.
2. Мембрана избирательно проницаема для различных ионов(полупроницаема)
например,
в состоянии покоя соотношение проницаемостей мембраны нервного
волокна для ионов калия, натрия и хлора
составляет
рК+: рNa+: рCl- = 1:0,04:0,45
следовательно,
преобладающим в состоянии покоя в нервном волокне
является
выходящий К+-ток
тогда как
токи всех остальных ионов будут мизерны по сравнению с выходящим
калиевым током
межклеточное вещество
клетка
К+> 40-50 раз
Na+> 7-10 раз
Cl->
Ca2+>
преобладающий ток
мизерные токи
18.
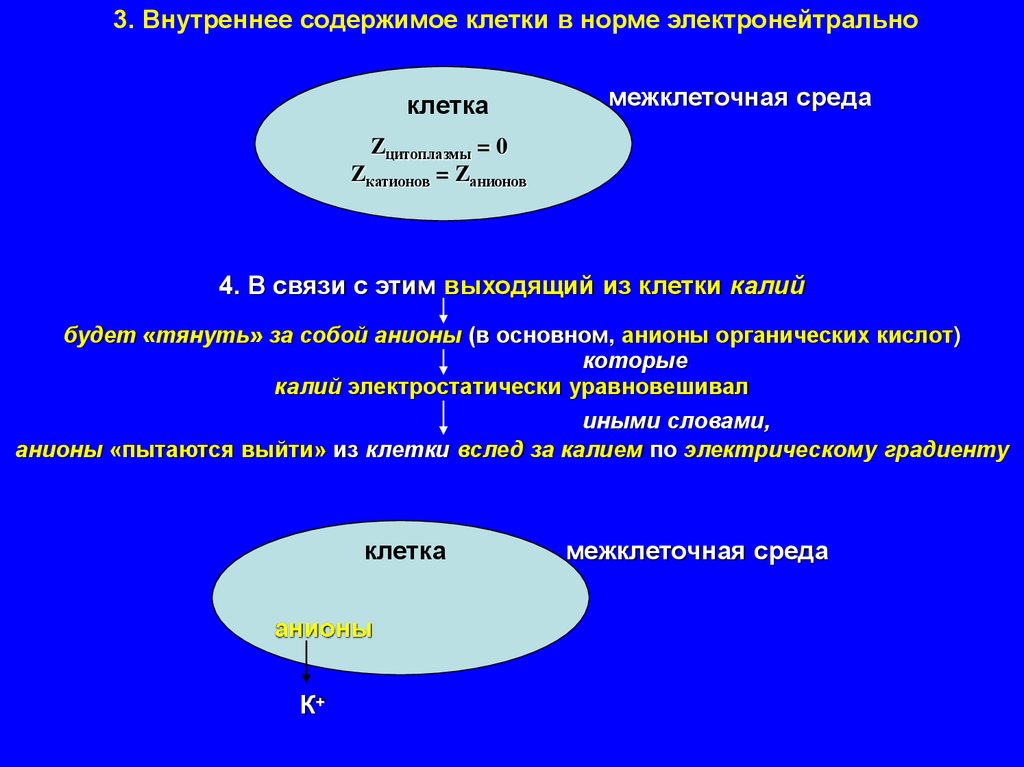
3. Внутреннее содержимое клетки в норме электронейтральноклетка
межклеточная среда
Zцитоплазмы = 0
Zкатионов = Zанионов
4. В связи с этим выходящий из клетки калий
будет «тянуть» за собой анионы (в основном, анионы органических кислот)
которые
калий электростатически уравновешивал
иными словами,
анионы «пытаются выйти» из клетки вслед за калием по электрическому градиенту
клетка
анионы
К+
межклеточная среда
19.
5. Но мембрана клеток не проницаема для органических анионовпоскольку
в ней содержится много отрицательно заряженных групп
в связи с чем
избыток анионов, следующих по электрическому градиенту за
выходящими из клетки ионами калия
«покинуть» клетку не может и скапливается у внутренней поверхности
мембраны,
при этом
притягивая положительные заряды со стороны наружной ее поверхности
(преимущественно ионы Na+)
как следствие,
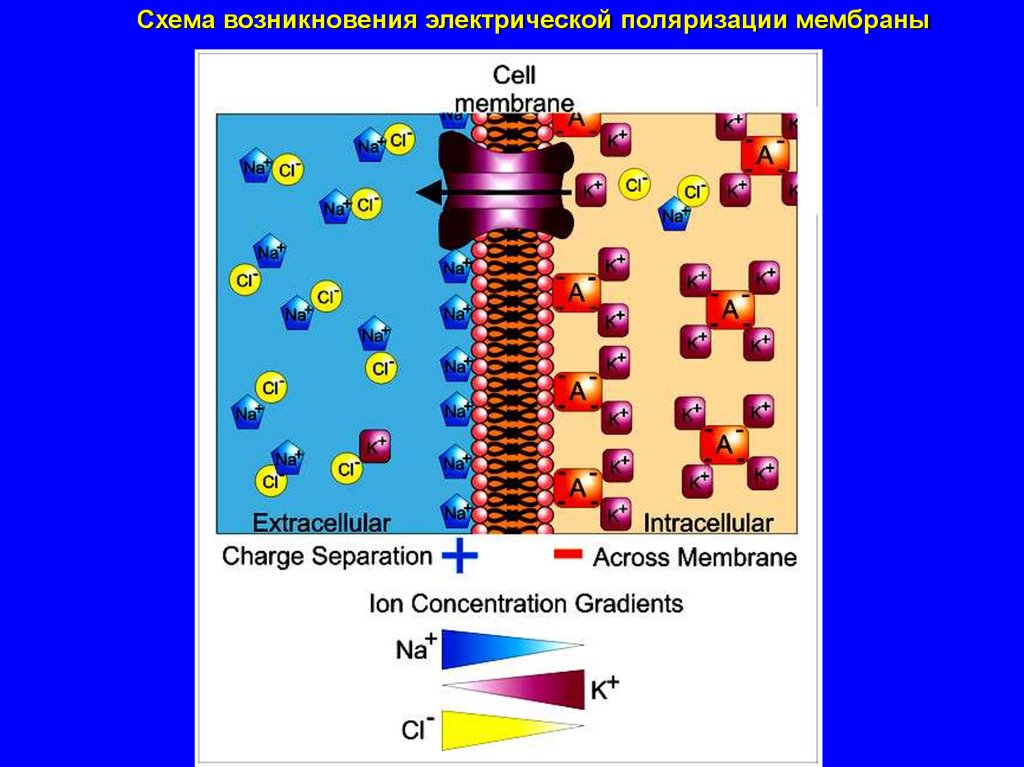
возникает электрическая поляризация мембраны
а именно,
мембрана изнутри заряжается отрицательно, а снаружи – положительно
+ _ + _ + + межклеточное
_
избыток анионов
пространство
+ _
_
изнутри клетки
+
клетка
_
притягивает
_
+
+ _
_
избыток катионов
_
_
плазматическая
снаружи клетки
+
+
мембрана
+
20.
Схема возникновения электрической поляризации мембраны21.
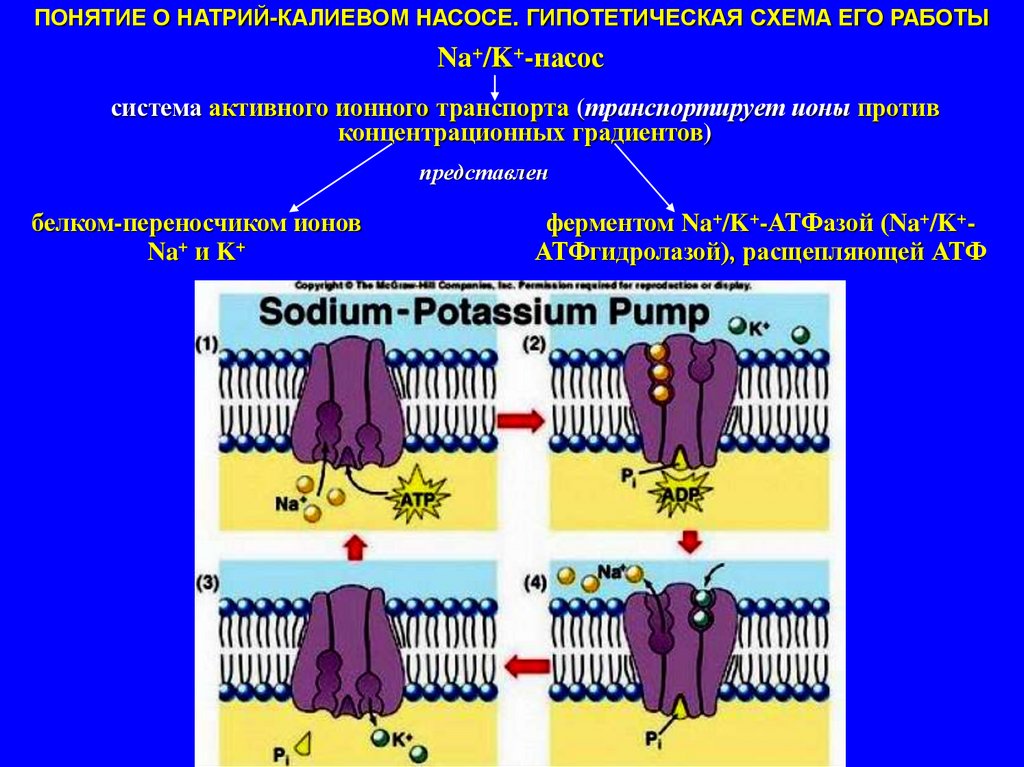
ПОНЯТИЕ О НАТРИЙ-КАЛИЕВОМ НАСОСЕ. ГИПОТЕТИЧЕСКАЯ СХЕМА ЕГО РАБОТЫNa+/K+-насос
система активного ионного транспорта (транспортирует ионы против
концентрационных градиентов)
представлен
белком-переносчиком ионов
Na+ и K+
ферментом Na+/K+-АТФазой (Na+/K+АТФгидролазой), расщепляющей АТФ
22.
ОСНОВНЫЕ ПРИНЦИПЫ РАСЧЕТА МЕМБРАННОГО ПОТЕНЦИАЛА ПОКОЯВ самом простом случае (если только учитывать диффузию ионов калия через
мембрану)
потенциал покоя
можно рассчитать по
формуле Нернста для калиевого равновесного потенциала:
К наружн
Емембр.= ЕК+= RT
Ln
F
К внутр
вместе с тем
если принять отношение К+наружн/К+внутрен, равным 1/50
то
мембранный потенциал, рассчитанный по формуле Нернста с учетом
только калиевого тока
составит
- 97,5 мВ
что
не достижимо ни для одной живой клетки
завышенное значение мембранного потенциала,
рассчитанное по формуле Нернста
связано с тем, что
в данном случае не учитывается движение через мембрану других ионов
(натрия, кальция, хлора), накладывающее «отпечаток» на величину
мембранного потенциала
23.
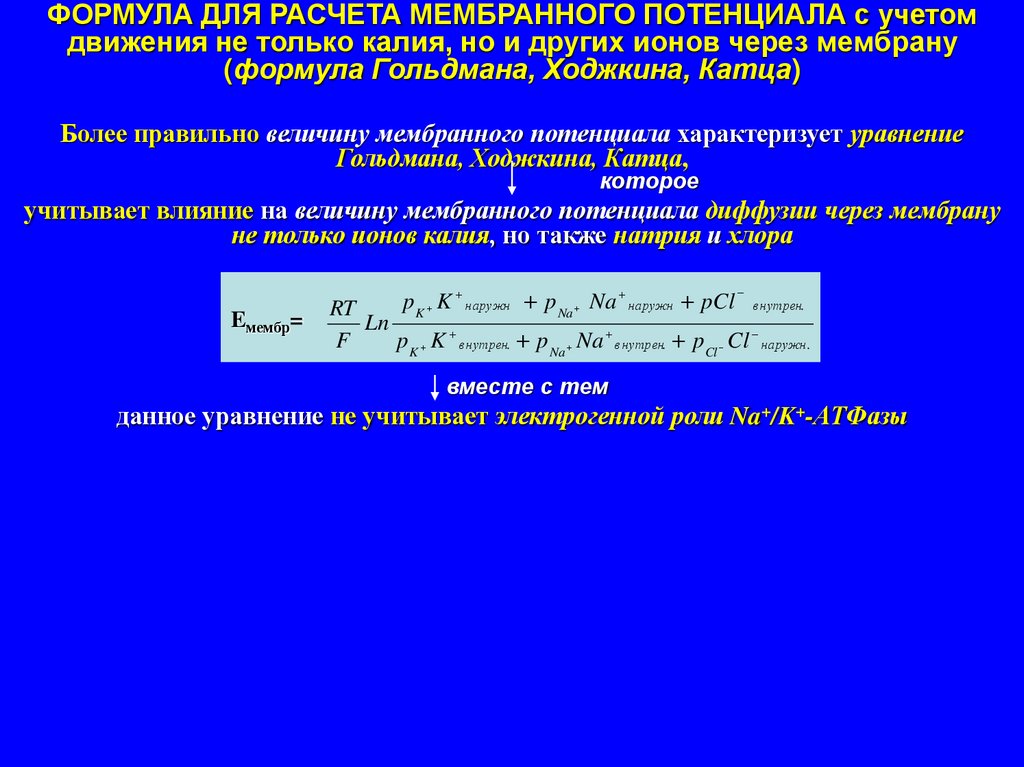
ФОРМУЛА ДЛЯ РАСЧЕТА МЕМБРАННОГО ПОТЕНЦИАЛА с учетомдвижения не только калия, но и других ионов через мембрану
(формула Гольдмана, Ходжкина, Катца)
Более правильно величину мембранного потенциала характеризует уравнение
Гольдмана, Ходжкина, Катца,
которое
учитывает влияние на величину мембранного потенциала диффузии через мембрану
не только ионов калия, но также натрия и хлора
Eмембр=
p K K наружн p Na Na наружн pCl внутрен.
RT
Ln
F
p K K внутрен. p Na Na внутрен. pCl Cl наружн.
вместе с тем
данное уравнение не учитывает электрогенной роли Na+/K+-АТФазы
24.
ЭЛЕКТРОГЕННАЯ РОЛЬ НАТРИЙ-КАЛИЕВОГО НАСОСАцикл работы натрийкалиевого насоса
натрий-калиевый насос работает так, что
клетка
за каждый цикл своей работы
выносит из клетки больше положительных зарядов (3 иона
натрия), чем возвращает их в клетку (2 иона калия)
в связи с чем
генерирует дополнительную разность потенциалов,
суммирующуюся с
потенциалом, создаваемым диффузией ионов по
концентрационным градиентам (т.н. концентрационным
потенциалом).
3 Na+
2 К+
_
_ _
_ клетка
_
_
некомпенсированные анионы,
которые ранее были
компенсированы натрием
25.
ОСНОВОПОЛАГАЮЩИЕ ФАКТОРЫ В СОЗДАНИИ МЕМБРАННОГО ПОТЕНЦИАЛАМЕМБРАННЫЙ ПОТЕНЦИАЛ
является суммой
концентрационного
потенциала, создаваемого в
потенциала мембраны
результате электрогенной работы
Na+/K+-насоса
создаваемого преимущественно
диффузией через мембрану ионов
калия по концентрационному
градиенту (из клетки наружу)
что обуславливает
появление некомпенсированных
анионов (преимущественно
органических) со стороны
внутренней поверхности
мембраны
что усиливает
в связи с тем, что мембрана для
них не проницаема
они скапливаются у внутренней
поверхности мембраны что обуславливает
электрическую поляризацию
и притягивают
мембраны (создание
положительные заряды со
мембранного потенциала)
стороны наружной
поверхности мембраны
26.
ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ МЕМБРАННОГО ПОТЕНЦИАЛА ПОКОЯМЕМБРАННЫЙ ПОТЕНЦИАЛ ПОКОЯ
определяет
определяет наличие в мембране электрического поля определенной напряженности
что обуславливает
определенную ориентацию заряженных групп макромолекул мембраны,
а, следовательно,
определенную их пространственную структуру
необходимую для:
проявления активности ферментов мембраны;
нормального состояния рецепторов мембраны;
нормального состояния ионных каналов мембраны, а, следовательно, и нормального
ионного транспорта и т.д.
27.
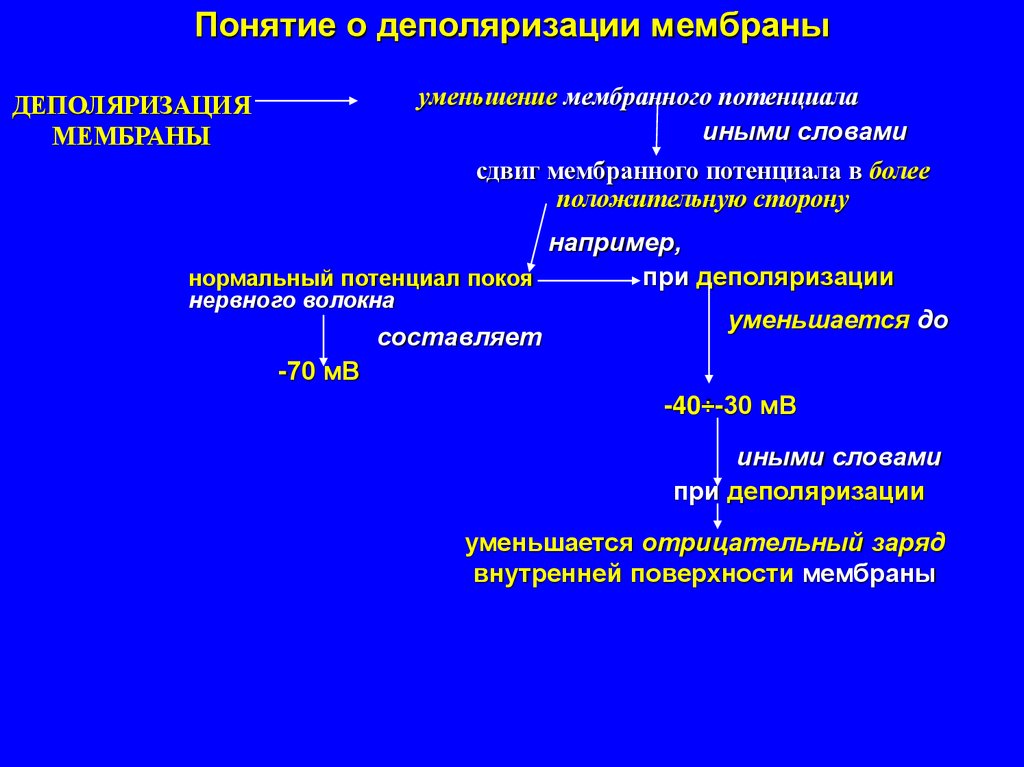
Понятие о деполяризации мембраныуменьшение мембранного потенциала
иными словами
ДЕПОЛЯРИЗАЦИЯ
МЕМБРАНЫ
сдвиг мембранного потенциала в более
положительную сторону
например,
при деполяризации
нормальный потенциал покоя
нервного волокна
составляет
уменьшается до
-70 мВ
-40÷-30 мВ
иными словами
при деполяризации
уменьшается отрицательный заряд
внутренней поверхности мембраны
28.
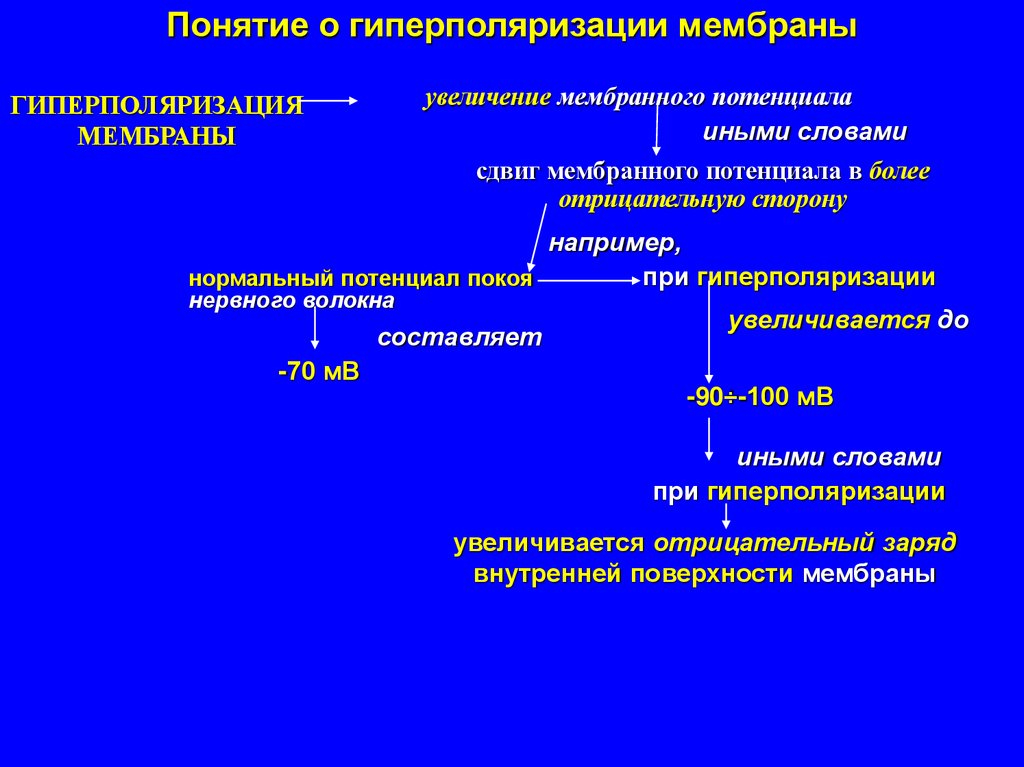
Понятие о гиперполяризации мембраныувеличение мембранного потенциала
иными словами
ГИПЕРПОЛЯРИЗАЦИЯ
МЕМБРАНЫ
сдвиг мембранного потенциала в более
отрицательную сторону
например,
при гиперполяризации
нормальный потенциал покоя
нервного волокна
составляет
-70 мВ
увеличивается до
-90÷-100 мВ
иными словами
при гиперполяризации
увеличивается отрицательный заряд
внутренней поверхности мембраны
29.
ЭЛЕКТРИЧЕСКИЕ ОТВЕТЫ ВОЗБУДИМЫХ СТРУКТУР30.
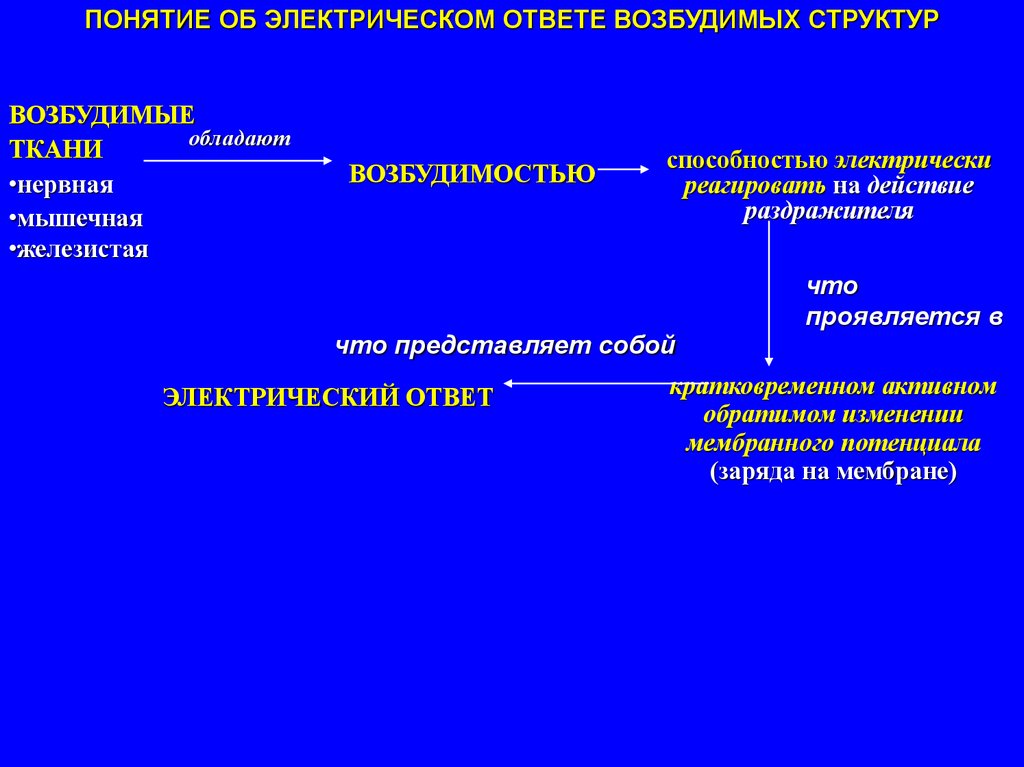
ПОНЯТИЕ ОБ ЭЛЕКТРИЧЕСКОМ ОТВЕТЕ ВОЗБУДИМЫХ СТРУКТУРВОЗБУДИМЫЕ
обладают
ТКАНИ
•нервная
•мышечная
•железистая
ВОЗБУДИМОСТЬЮ
способностью электрически
реагировать на действие
раздражителя
что
проявляется в
что представляет собой
ЭЛЕКТРИЧЕСКИЙ ОТВЕТ
кратковременном активном
обратимом изменении
мембранного потенциала
(заряда на мембране)
31.
ФАКТОРЫ, ОБУСЛАВЛИВАЮЩИЕ СПОСОБНОСТЬ ВОЗБУДИМЫХ СТРУКТУР КЭЛЕКТРИЧЕСКОМУ ОТВЕТУ
Электрический ответ клеток возбудимых тканей
возможен только
благодаря тому, что
в их мембранах наряду с каналами пассивной утечки ионов
(присущими мембранам всех живых клеток)
имеются
специальные электровозбудимые каналы
(потенциалзависимые или электрогенные каналы)
не имеют ворот и открыты
постоянно независимо от
величины мембранного
потенциала
которые
в отличие от каналов пассивной утечки ионов имеют
своеобразные ворота (образованные белковыми
молекулами)
состояние которых (открыты они или закрыты)
зависит от
величины мембранного потенциала
движение ионов через электровозбудимые каналы,
подобно таковому через каналы пассивной утечки
ионов
осуществляется
пассивно (по градиенту концентрации иона)
путем простой диффузии
потенциалзависимость
(электровозбудимость) ворот
и соответственно каналов
32.
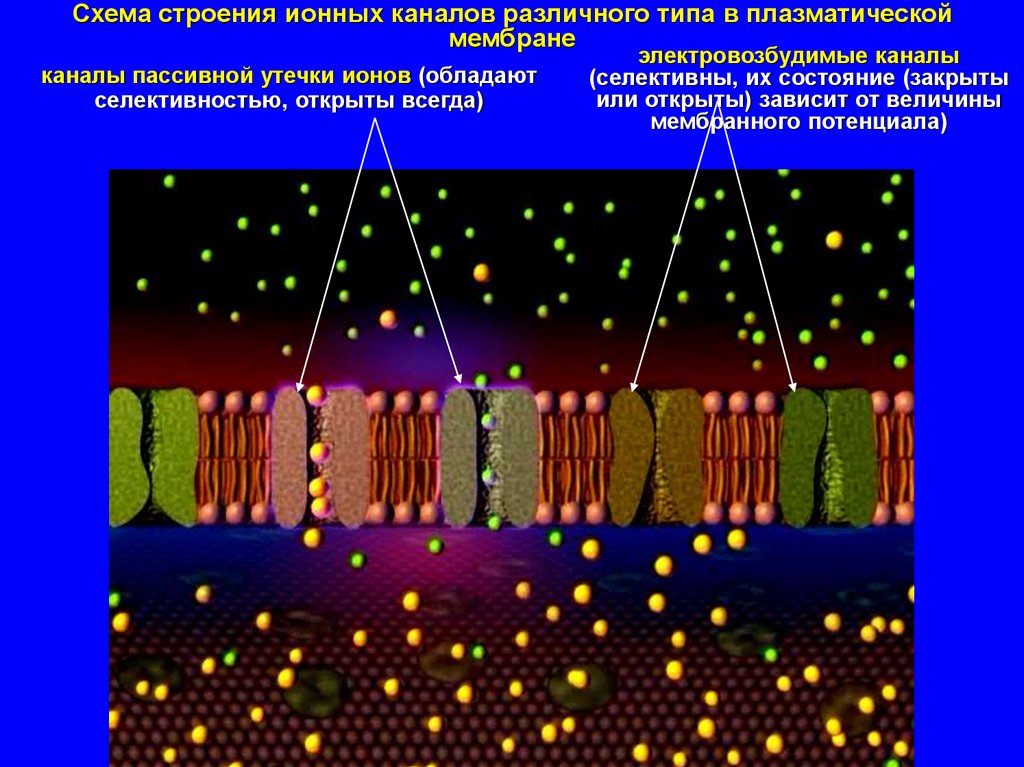
Схема строения ионных каналов различного типа в плазматическоймембране
каналы пассивной утечки ионов (обладают
селективностью, открыты всегда)
электровозбудимые каналы
(селективны, их состояние (закрыты
или открыты) зависит от величины
мембранного потенциала)
33.
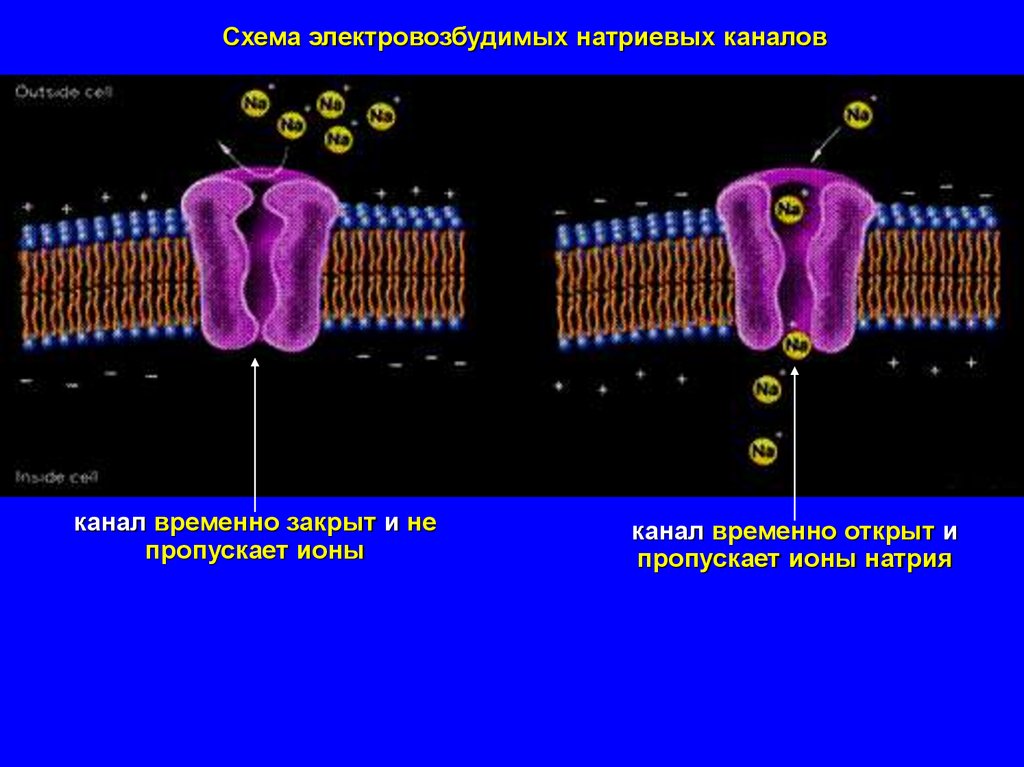
Схема электровозбудимых натриевых каналовканал временно закрыт и не
пропускает ионы
канал временно открыт и
пропускает ионы натрия
34.
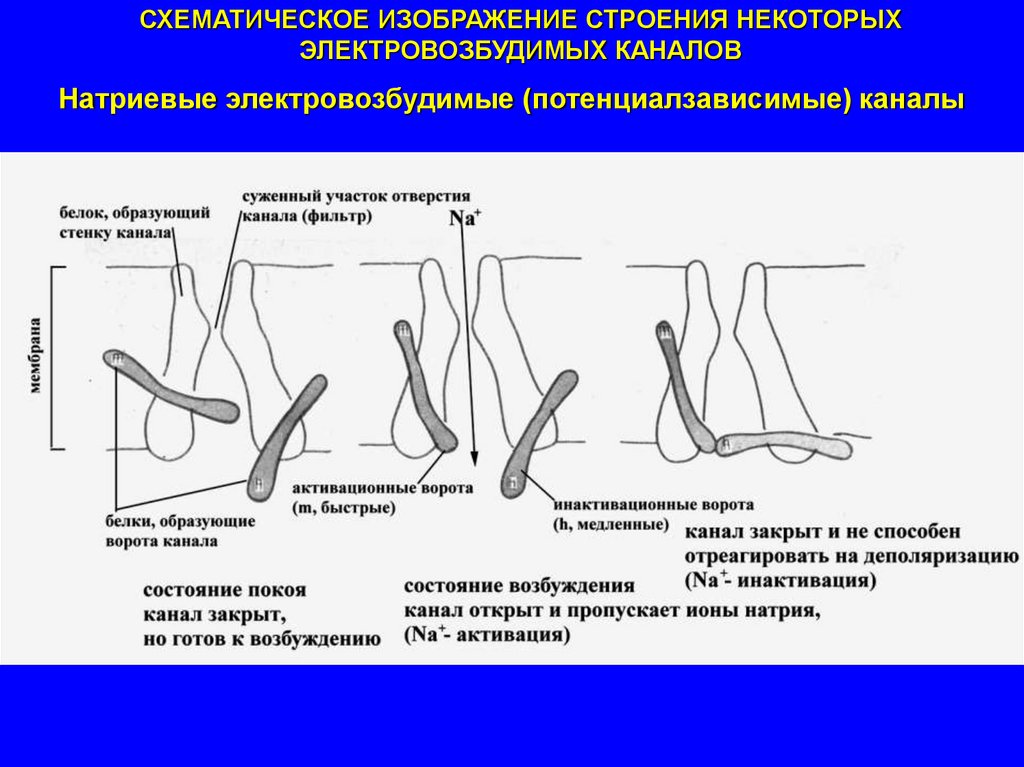
СХЕМАТИЧЕСКОЕ ИЗОБРАЖЕНИЕ СТРОЕНИЯ НЕКОТОРЫХЭЛЕКТРОВОЗБУДИМЫХ КАНАЛОВ
Натриевые электровозбудимые (потенциалзависимые) каналы
35.
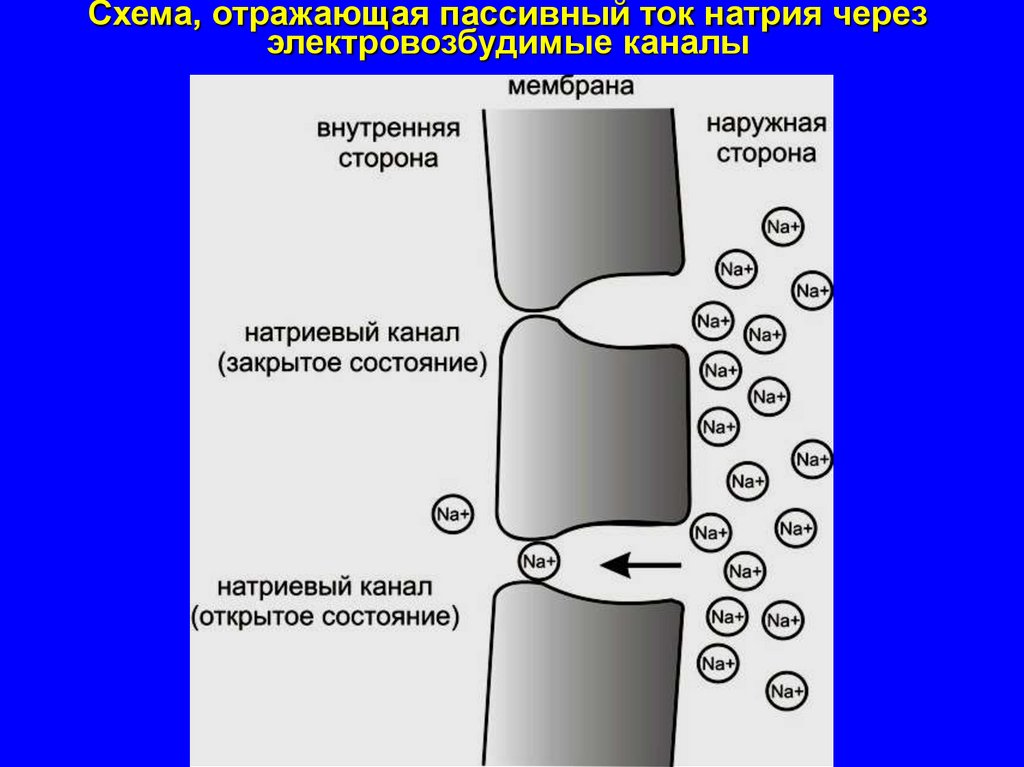
Схема, отражающая пассивный ток натрия черезэлектровозбудимые каналы
36.
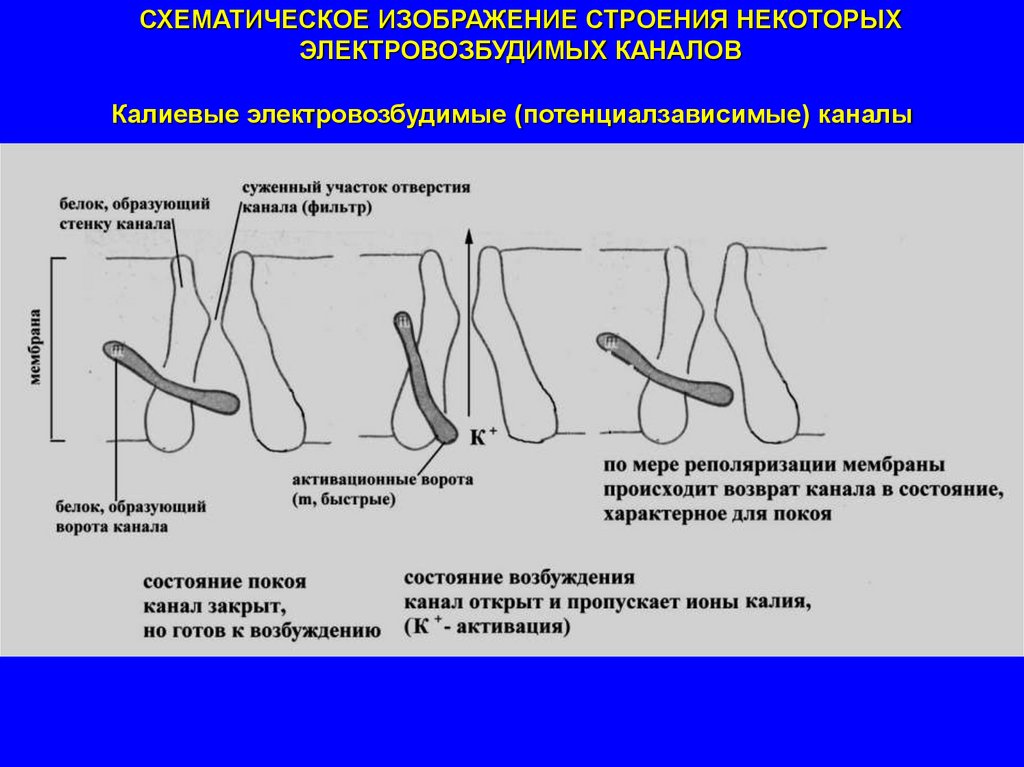
СХЕМАТИЧЕСКОЕ ИЗОБРАЖЕНИЕ СТРОЕНИЯ НЕКОТОРЫХЭЛЕКТРОВОЗБУДИМЫХ КАНАЛОВ
Калиевые электровозбудимые (потенциалзависимые) каналы
37.
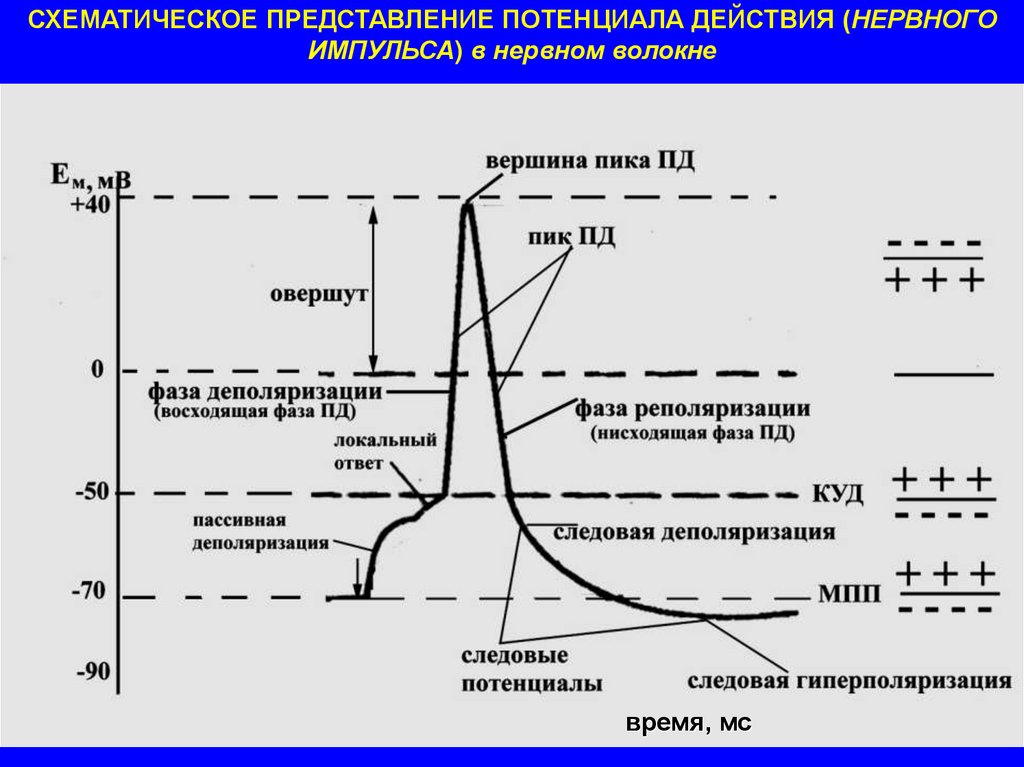
СХЕМАТИЧЕСКОЕ ПРЕДСТАВЛЕНИЕ ПОТЕНЦИАЛА ДЕЙСТВИЯ (НЕРВНОГОИМПУЛЬСА) в нервном волокне
время, мс
38.
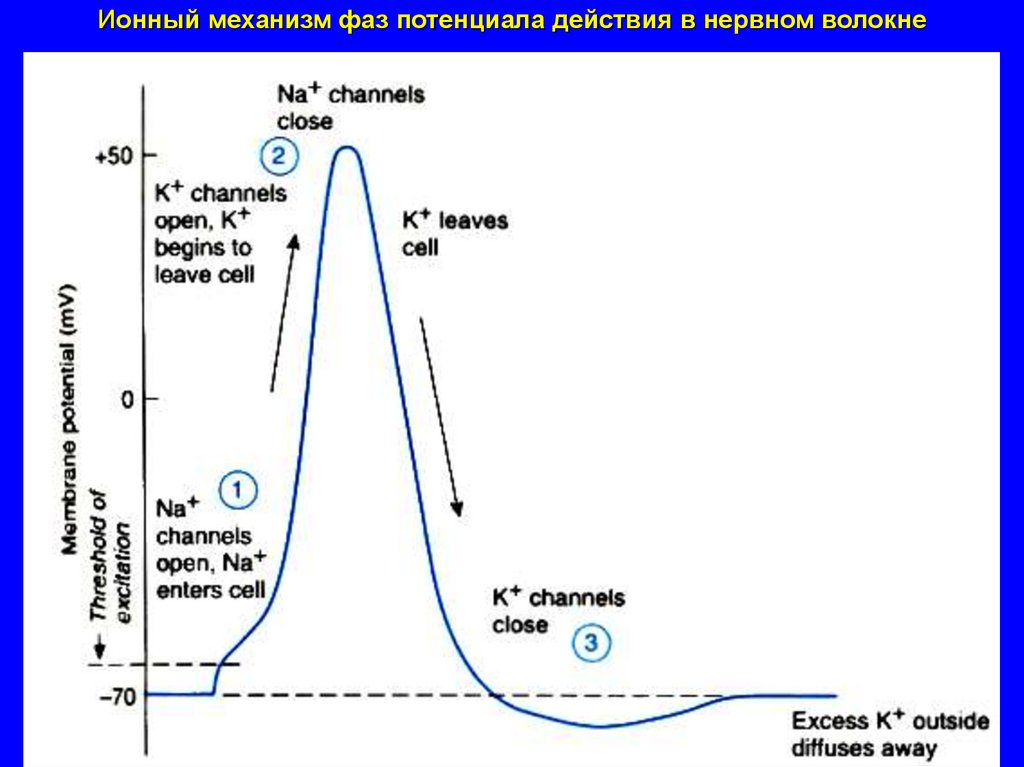
Ионный механизм фаз потенциала действия в нервном волокне39.
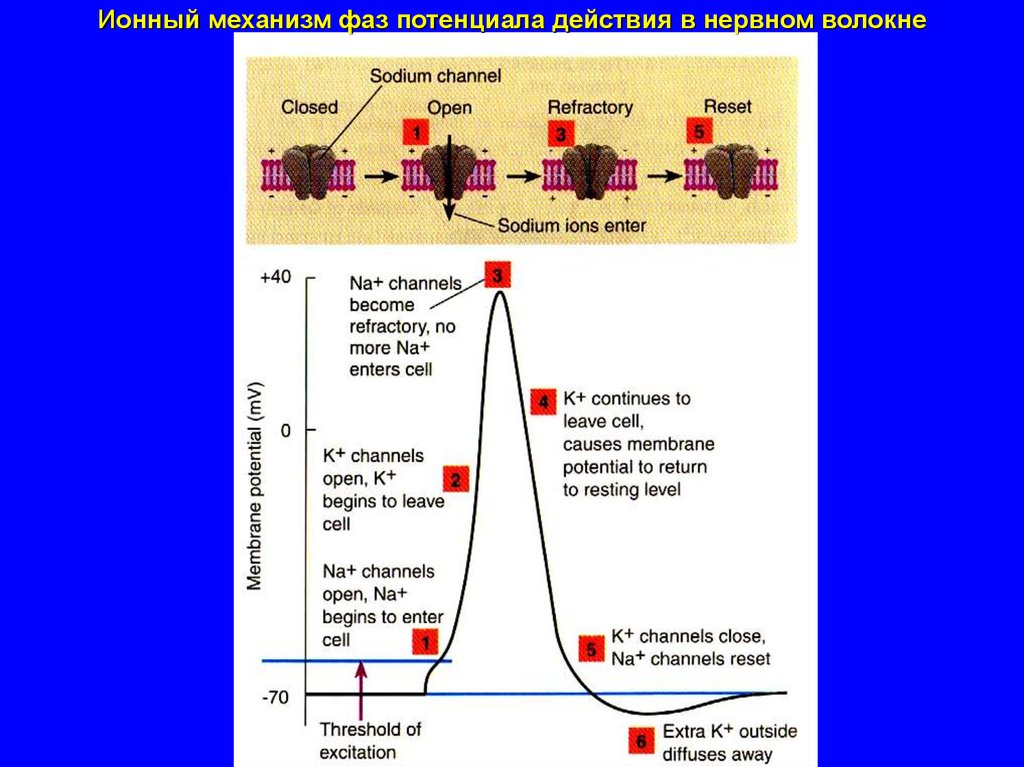
Ионный механизм фаз потенциала действия в нервном волокне40.
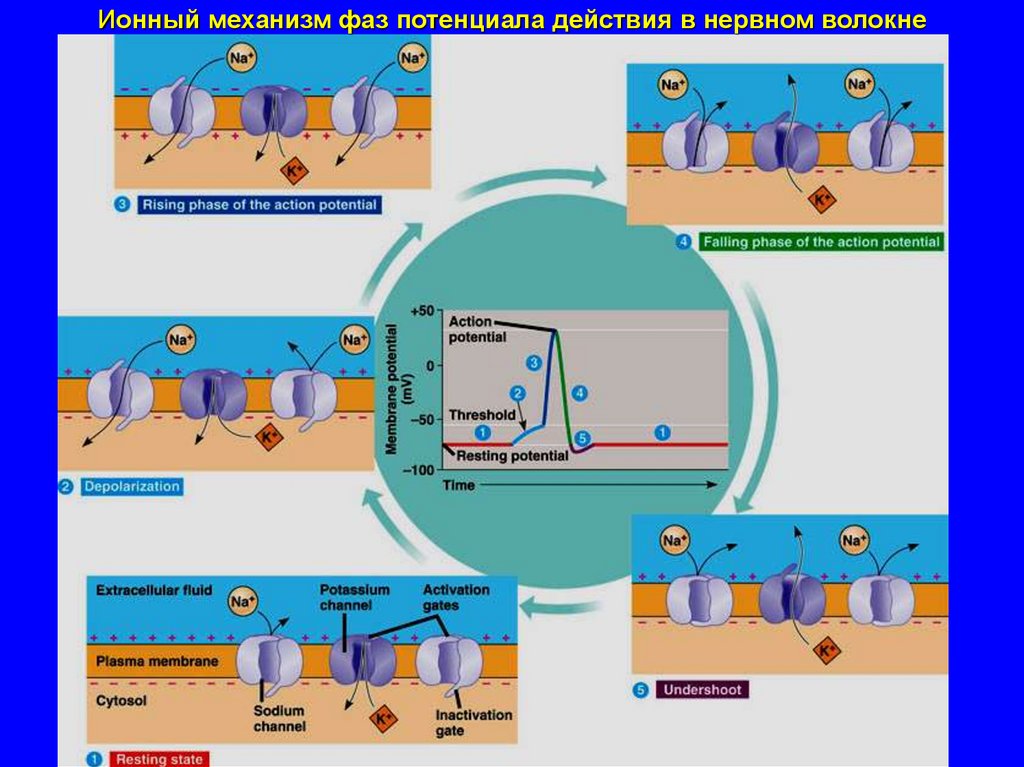
Ионный механизм фаз потенциала действия в нервном волокне41.
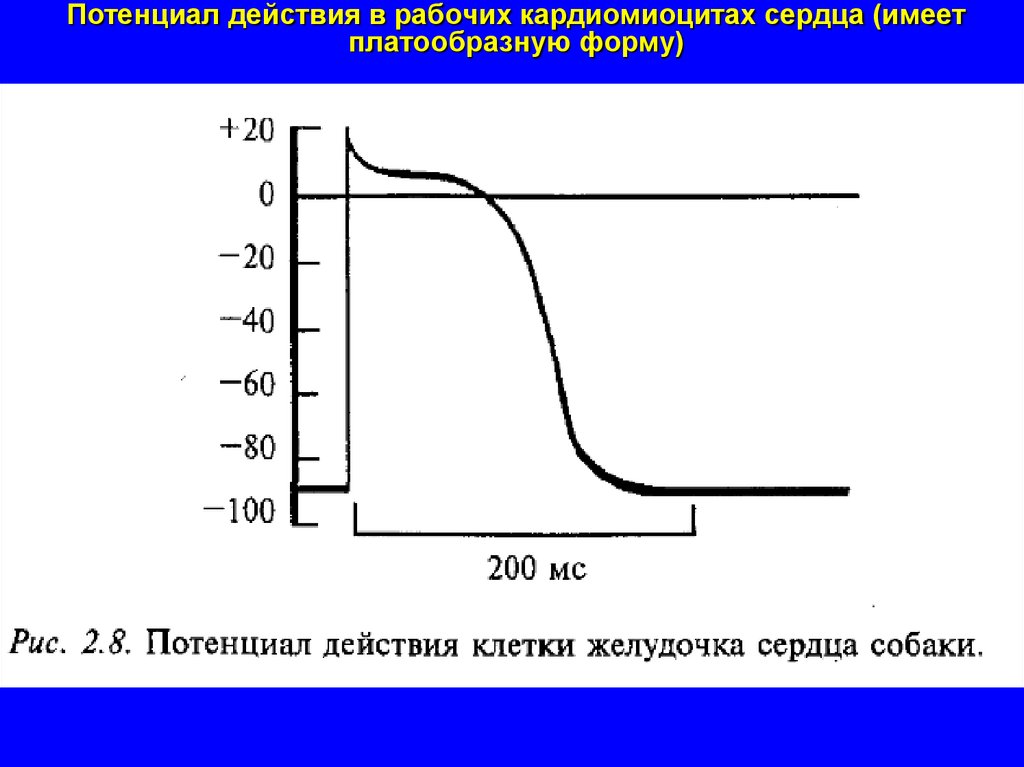
Потенциал действия в рабочих кардиомиоцитах сердца (имеетплатообразную форму)
42.
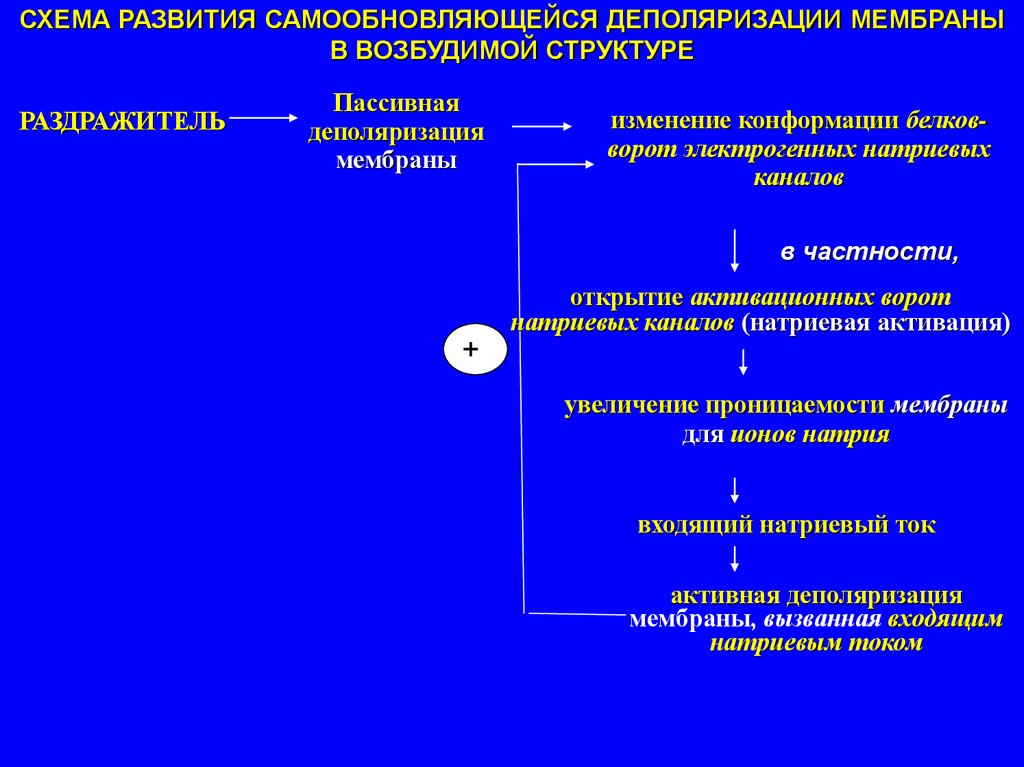
СХЕМА РАЗВИТИЯ САМООБНОВЛЯЮЩЕЙСЯ ДЕПОЛЯРИЗАЦИИ МЕМБРАНЫВ ВОЗБУДИМОЙ СТРУКТУРЕ
РАЗДРАЖИТЕЛЬ
Пассивная
деполяризация
мембраны
изменение конформации белковворот электрогенных натриевых
каналов
в частности,
открытие активационных ворот
натриевых каналов (натриевая активация)
+
увеличение проницаемости мембраны
для ионов натрия
входящий натриевый ток
активная деполяризация
мембраны, вызванная входящим
натриевым током
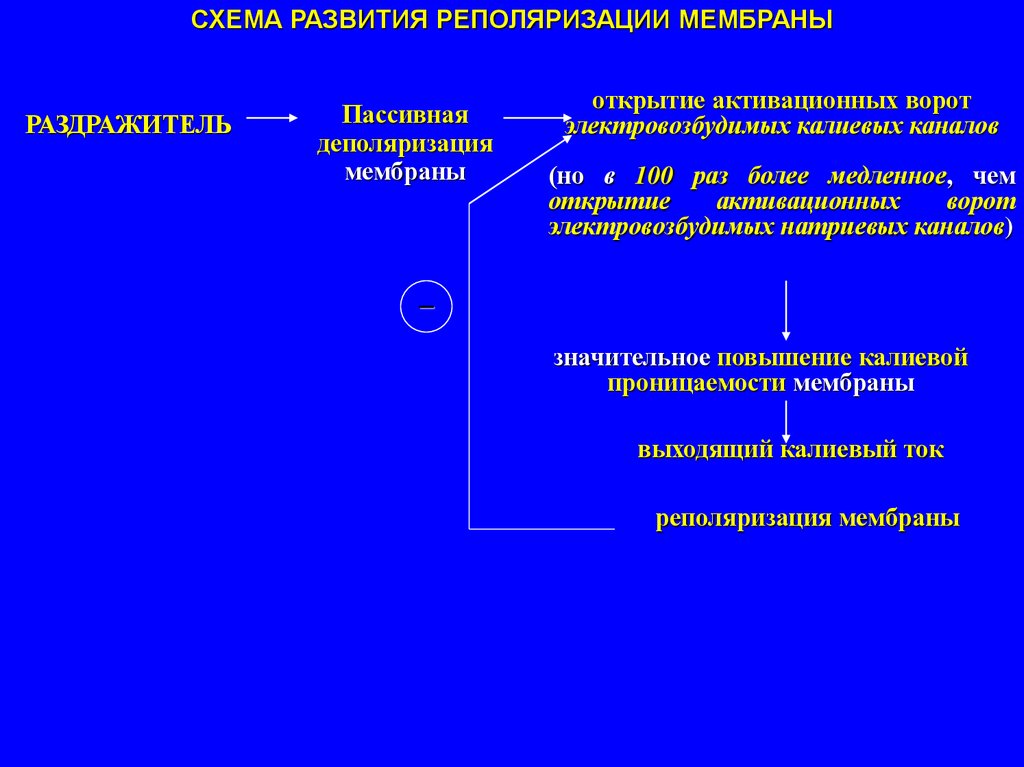
43.
СХЕМА РАЗВИТИЯ РЕПОЛЯРИЗАЦИИ МЕМБРАНЫРАЗДРАЖИТЕЛЬ
Пассивная
деполяризация
мембраны
открытие активационных ворот
электровозбудимых калиевых каналов
(но в 100 раз более медленное, чем
открытие
активационных
ворот
электровозбудимых натриевых каналов)
–
значительное повышение калиевой
проницаемости мембраны
выходящий калиевый ток
реполяризация мембраны
44.
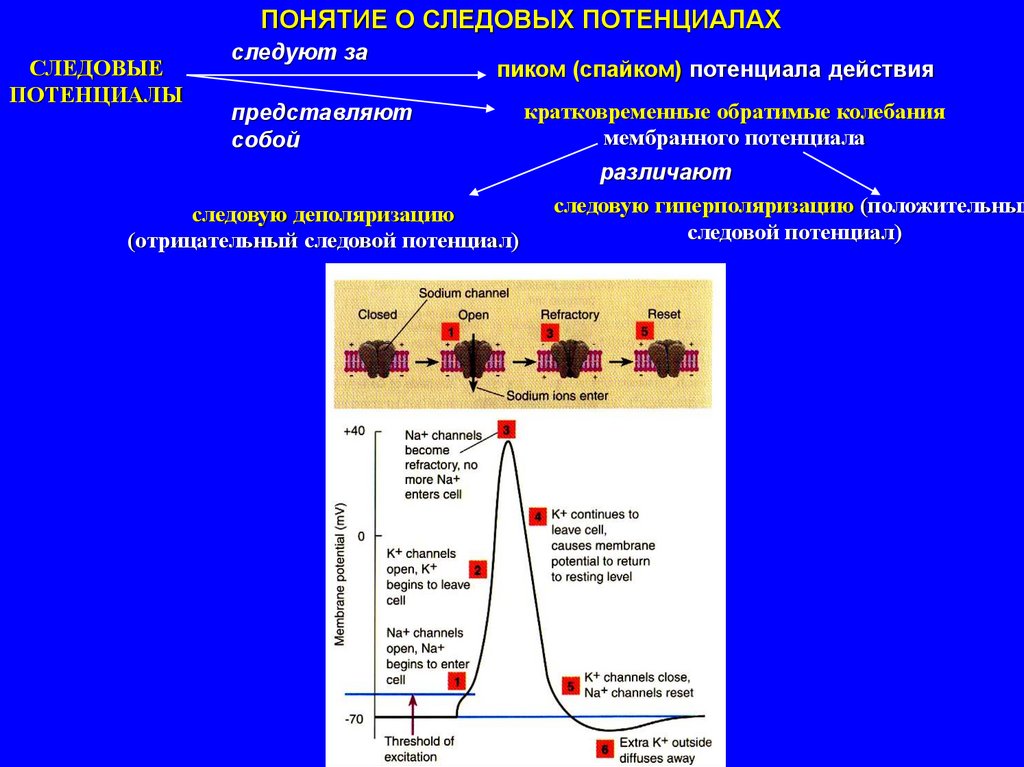
ПОНЯТИЕ О СЛЕДОВЫХ ПОТЕНЦИАЛАХСЛЕДОВЫЕ
ПОТЕНЦИАЛЫ
следуют за
пиком (спайком) потенциала действия
представляют
собой
следовую деполяризацию
(отрицательный следовой потенциал)
кратковременные обратимые колебания
мембранного потенциала
различают
следовую гиперполяризацию (положительный
следовой потенциал)
45.
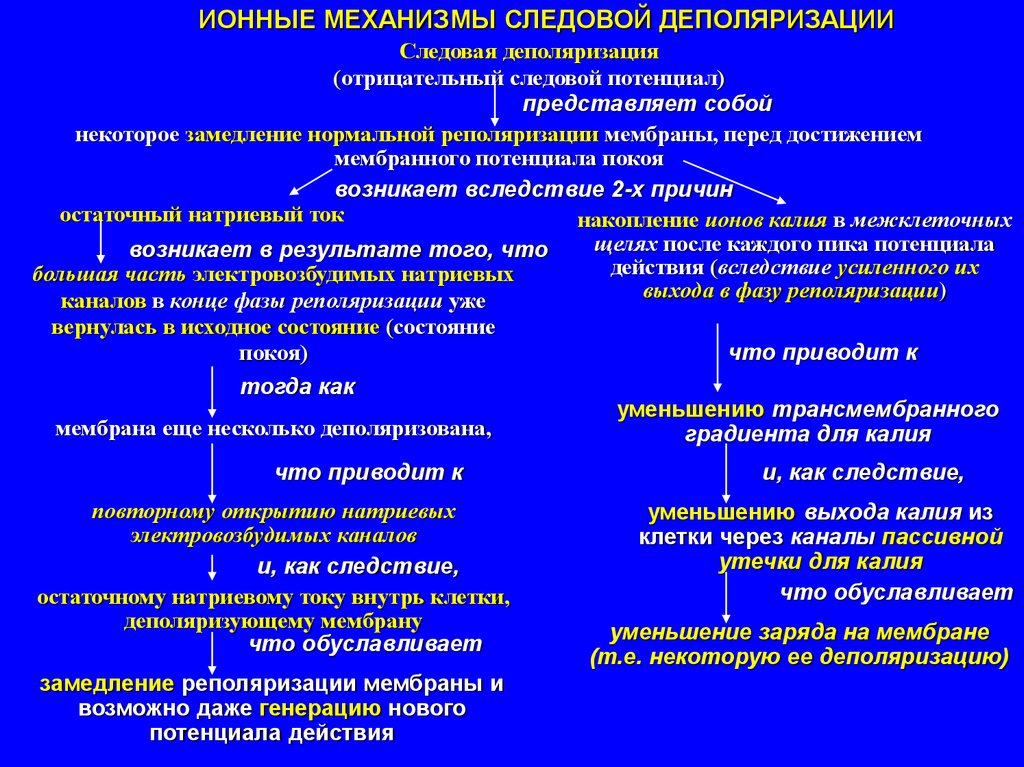
ИОННЫЕ МЕХАНИЗМЫ СЛЕДОВОЙ ДЕПОЛЯРИЗАЦИИСледовая деполяризация
(отрицательный следовой потенциал)
представляет собой
некоторое замедление нормальной реполяризации мембраны, перед достижением
мембранного потенциала покоя
возникает вследствие 2-х причин
остаточный натриевый ток
накопление ионов калия в межклеточных
щелях после каждого пика потенциала
возникает в результате того, что
действия (вследствие усиленного их
большая часть электровозбудимых натриевых
выхода в фазу реполяризации)
каналов в конце фазы реполяризации уже
вернулась в исходное состояние (состояние
что приводит к
покоя)
тогда как
мембрана еще несколько деполяризована,
что приводит к
повторному открытию натриевых
электровозбудимых каналов
и, как следствие,
остаточному натриевому току внутрь клетки,
деполяризующему мембрану
что обуславливает
замедление реполяризации мембраны и
возможно даже генерацию нового
потенциала действия
уменьшению трансмембранного
градиента для калия
и, как следствие,
уменьшению выхода калия из
клетки через каналы пассивной
утечки для калия
что обуславливает
уменьшение заряда на мембране
(т.е. некоторую ее деполяризацию)
46.
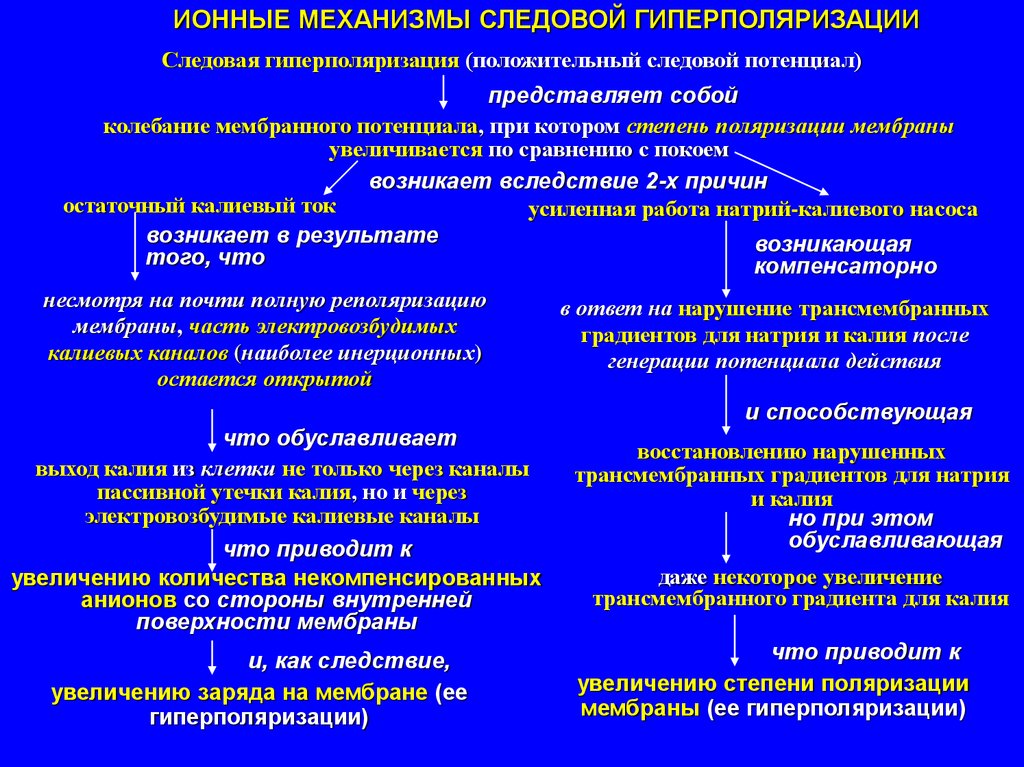
ИОННЫЕ МЕХАНИЗМЫ СЛЕДОВОЙ ГИПЕРПОЛЯРИЗАЦИИСледовая гиперполяризация (положительный следовой потенциал)
представляет собой
колебание мембранного потенциала, при котором степень поляризации мембраны
увеличивается по сравнению с покоем
возникает вследствие 2-х причин
остаточный калиевый ток
усиленная работа натрий-калиевого насоса
возникает в результате
возникающая
того, что
компенсаторно
несмотря на почти полную реполяризацию
в ответ на нарушение трансмембранных
мембраны, часть электровозбудимых
градиентов для натрия и калия после
калиевых каналов (наиболее инерционных)
генерации потенциала действия
остается открытой
и способствующая
что обуславливает
восстановлению нарушенных
выход калия из клетки не только через каналы
трансмембранных градиентов для натрия
пассивной утечки калия, но и через
и калия
электровозбудимые калиевые каналы
но при этом
обуславливающая
что приводит к
даже некоторое увеличение
увеличению количества некомпенсированных
трансмембранного градиента для калия
анионов со стороны внутренней
поверхности мембраны
что приводит к
и, как следствие,
увеличению степени поляризации
увеличению заряда на мембране (ее
мембраны (ее гиперполяризации)
гиперполяризации)
47.
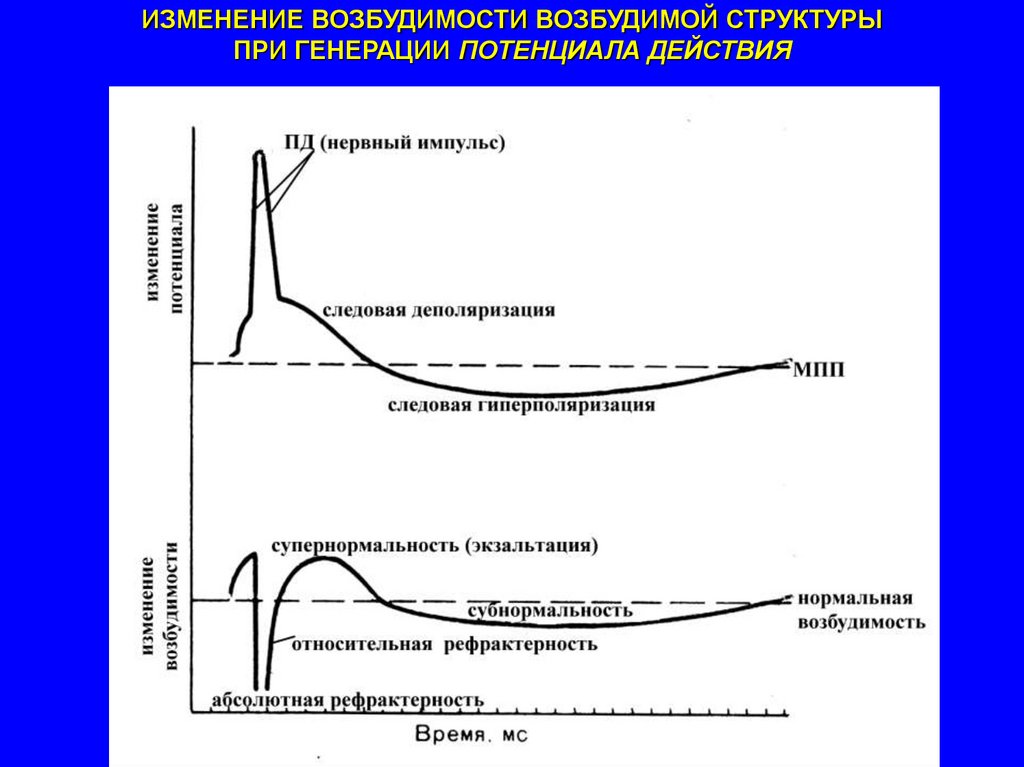
ИЗМЕНЕНИЕ ВОЗБУДИМОСТИ ВОЗБУДИМОЙ СТРУКТУРЫПРИ ГЕНЕРАЦИИ ПОТЕНЦИАЛА ДЕЙСТВИЯ
48.
ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ ПОТЕНЦИАЛА ДЕЙСТВИЯНЕРВНЫЙ ИМПУЛЬС
обладая способностью к
распространению на большие расстояния вдоль возбудимой структуры без затухания
обеспечивает
передачу информации в нервной системе
и в конечном итоге
проведение и передачу регулирующих сигналов к исполнительным органам
(скелетным мышцам, сердечным мышечным волокнам, гладкомышечным клеткам
кровеносных сосудов, стенок полых органов, некоторым железистым клеткам)
что обуславливает
включение их в деятельность или изменение исходной активности
что формирует
ответную реакцию организма на раздражитель
49.
Адекватный раздражитель для возбудимых структурЭЛЕКТРИЧЕСКИЙ ТОК
поскольку в реальных условиях
возбуждение вдоль возбудимой структуры
распространяется за счет
местных ионных токов
возникающих
между возбужденными (деполяризованными) и покоящимися (нормально
поляризованными) участками возбудимой структуры
при этом
местные ионный токи, чтобы вызвать возбуждение соседних покоящихся
участков мембраны возбудимой структуры
должны иметь
определенную силу (не меньшую пороговой)
Схема, отражающая проведение возбуждения по безмякотному нервному волокну
50.
НЕЗАТУХАЮЩЕЕ РАСПРОСТРАНЕНИЕ нервного импульса вдоль аксонанервной клетки
возможно благодаря тому, что
местные ионные токи
возникающие между
возбужденным и покоящимся участками нервного волокна
имеют амплитуду в 5-6 раз большую пороговой (т.е. минимальной и
достаточной для инициации возбуждения)
51.
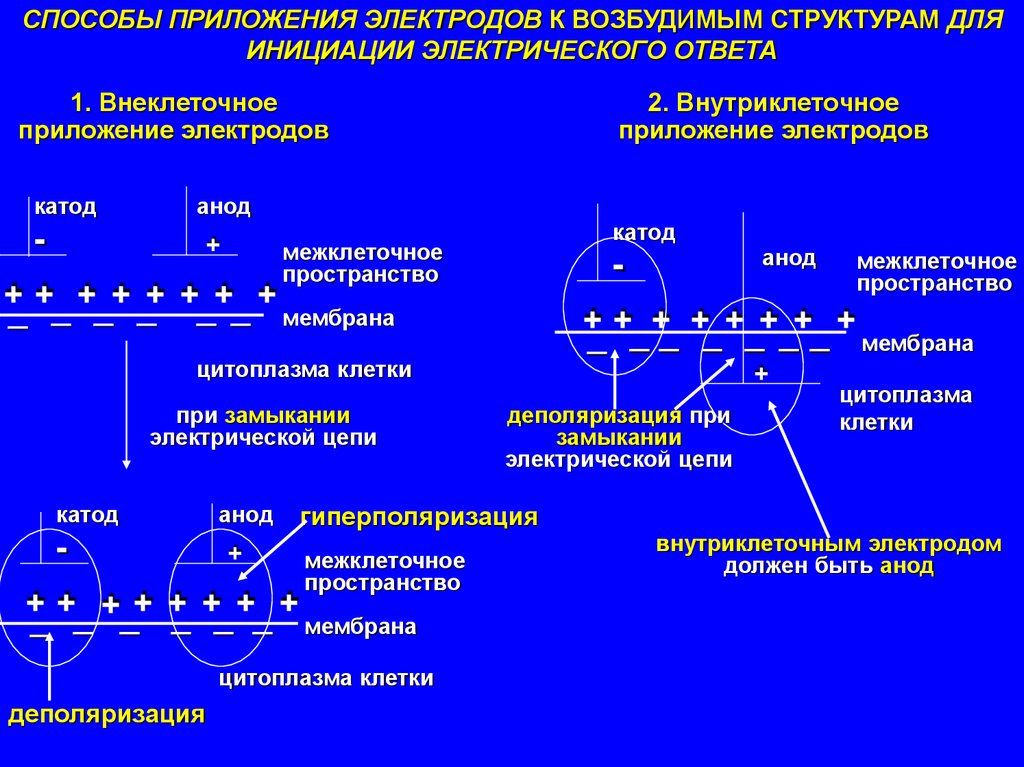
СПОСОБЫ ПРИЛОЖЕНИЯ ЭЛЕКТРОДОВ К ВОЗБУДИМЫМ СТРУКТУРАМ ДЛЯИНИЦИАЦИИ ЭЛЕКТРИЧЕСКОГО ОТВЕТА
1. Внеклеточное
приложение электродов
катод
2. Внутриклеточное
приложение электродов
анод
катод
+
межклеточное
пространство
++ + + + + + +
_ _ _ _ _ _ мембрана
анод
межклеточное
пространство
++ + + + + + +
_ _ _ _ _ _ _ мембрана
цитоплазма клетки
при замыкании
электрической цепи
катод
анод
деполяризация при
замыкании
электрической цепи
цитоплазма
клетки
гиперполяризация
+
межклеточное
пространство
++ ++ + + + +
_ _ _ _ _ _ мембрана
цитоплазма клетки
деполяризация
+
внутриклеточным электродом
должен быть анод
52.
Схема пассивных (электротонических) ответов и локальной деполяризации внервном волокне, возникающих при действии раздражителей подпороговой
силы
Стрелкой отмечен момент начала действия раздражителя
10%, 20%, 50% и 75% – указана сила раздражителя (в % от пороговой).
53.
Схема, отражающая генерацию электротонических ответов, локальныхответов и потенциала действия в аксоне нервной клетки по мере увеличения
силы раздражителя
54.
ПОНЯТИЕ О ЛОКАЛЬНОМ ОТВЕТЕ, ЕГО ПРИРОДАЛОКАЛЬНЫЙ
ОТВЕТ
активная форма ответа возбудимых структур на действие раздражителей, сила
которых ниже пороговой
подобно потенциалу действия
локальный ответ связан с временным изменением проницаемости мембраны для
определенных ионов (преимущественно ионов натрия или кальция)
что обусловлено
кратковременным открытием под действием деполяризующего
раздражителя части электровозбудимых натриевых или кальциевых
каналов
что обуславливает
развитие кратковременной активной деполяризации мембраны
но в отличие от потенциала действия
активная деполяризация при локальном ответе
не приобретает самообновляющийся характер
в связи с чем
амплитуда локального ответа всегда значительно ниже таковой
потенциала действия и зависит от силы раздржителя
55.
СВОЙСТВА ЛОКАЛЬНОГО ОТВЕТАЛОКАЛЬНЫЙ
ОТВЕТ
представляет собой
активную деполяризацию мембрану (т.е. деполяризацию, вызванную временными ионными
токами), которая не приобретает самообновляющийся характер
в связи с чем
амплитуда и длительность локального
ответа
при возникновении локального ответа
зависят от силы и длительности
вызвавшего его раздражителя и всегда
гораздо ниже амплитуды потенциала
действия
задействуются не все имеющиеся
электровозбудимые каналы (т.е. деполяризация не
приобретает самообновляющийся характер)
локальный ответ в связи с гораздо меньшей
амплитудой, по сравнению с таковой
потенциала действия
в связи с чем
локальный ответ способен к СУММАЦИИ
в частности,
как правило, является
при действии нового деполяризующего
раздражителя в момент генерации
подпороговым раздражителем для окружающих локального ответа, он будет себя усиливать
место его возникновения участков мембраны
возбудимой структуры
поскольку
в связи с чем
новый деполяризующий раздражитель
в отличие от потенциала действия, НЕ МОЖЕТ
приведет к открытию дополнительного
РАСПРОСТРАНЯТЬСЯ НА БОЛЬШИЕ
количества незадействованных ранее
РАССТОЯНИЯ, НЕ ЗАТУХАЯ (т.е. носит
электровозбудимых каналов
местный характер)
56.
ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ ЛОКАЛЬНОГО ОТВЕТАЛОКАЛЬНЫЙ ОТВЕТ
благодаря
способности к суммации
делает возможным
интегративную обработку информации, поступающей к возбудимой структуре по
различным "входам"
57.
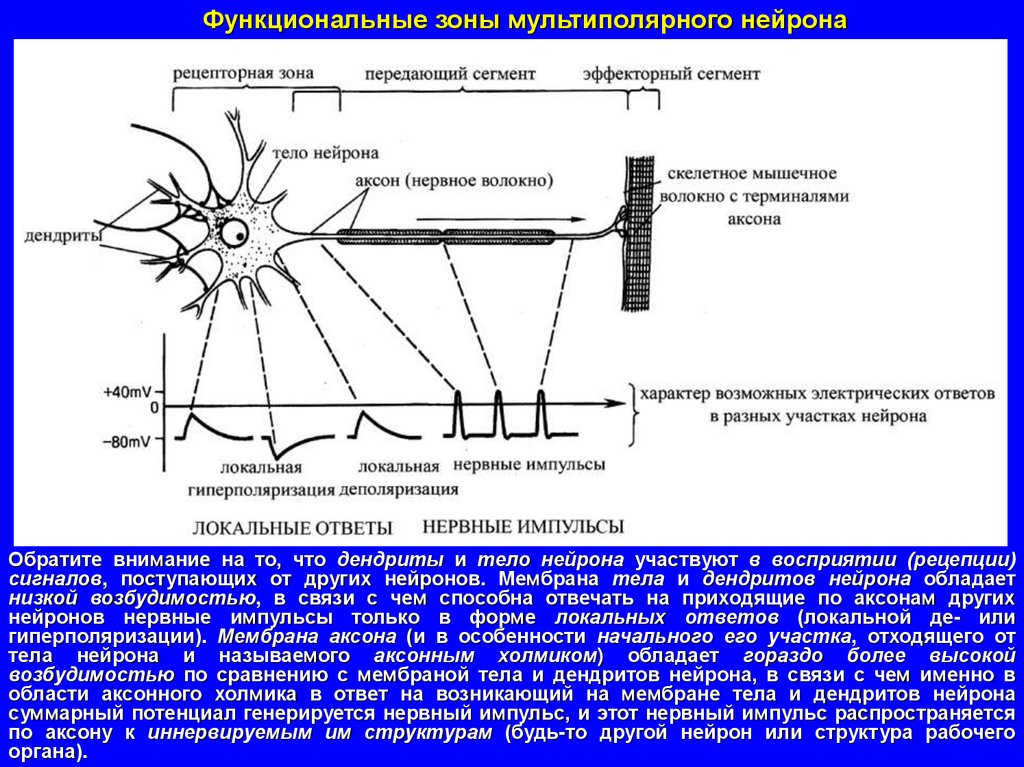
Функциональные зоны мультиполярного нейронаОбратите внимание на то, что дендриты и тело нейрона участвуют в восприятии (рецепции)
сигналов, поступающих от других нейронов. Мембрана тела и дендритов нейрона обладает
низкой возбудимостью, в связи с чем способна отвечать на приходящие по аксонам других
нейронов нервные импульсы только в форме локальных ответов (локальной де- или
гиперполяризации). Мембрана аксона (и в особенности начального его участка, отходящего от
тела нейрона и называемого аксонным холмиком) обладает гораздо более высокой
возбудимостью по сравнению с мембраной тела и дендритов нейрона, в связи с чем именно в
области аксонного холмика в ответ на возникающий на мембране тела и дендритов нейрона
суммарный потенциал генерируется нервный импульс, и этот нервный импульс распространяется
по аксону к иннервируемым им структурам (будь-то другой нейрон или структура рабочего
органа).
58.
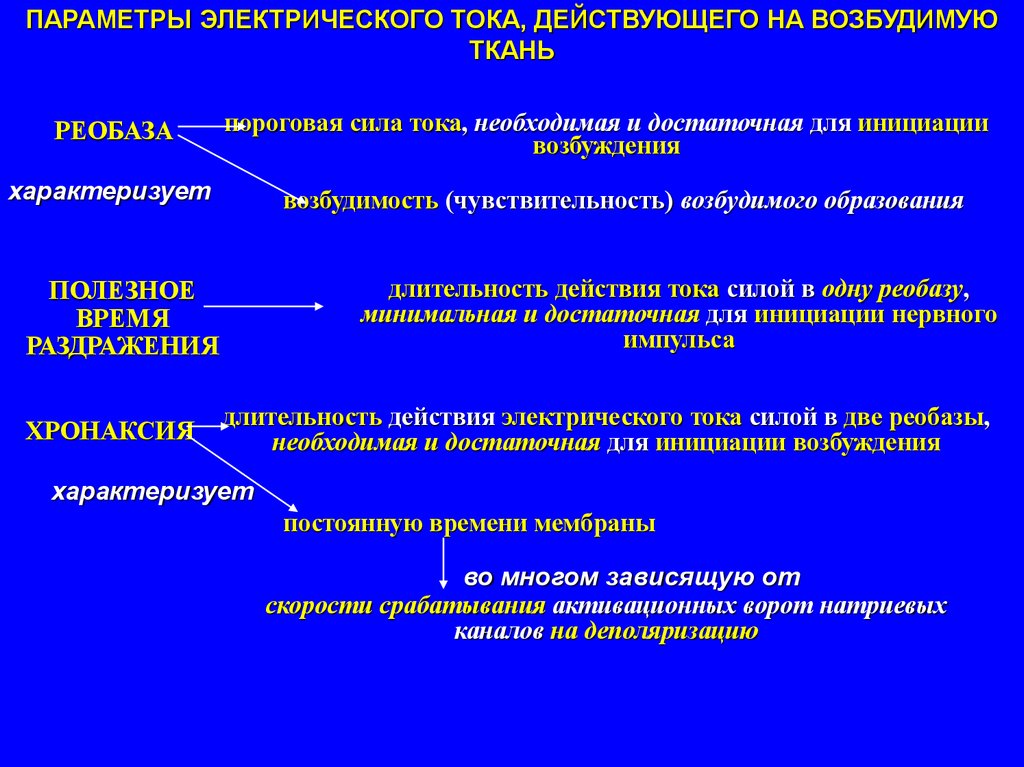
ПАРАМЕТРЫ ЭЛЕКТРИЧЕСКОГО ТОКА, ДЕЙСТВУЮЩЕГО НА ВОЗБУДИМУЮТКАНЬ
РЕОБАЗА
пороговая сила тока, необходимая и достаточная для инициации
возбуждения
характеризует
возбудимость (чувствительность) возбудимого образования
длительность действия тока силой в одну реобазу,
минимальная и достаточная для инициации нервного
импульса
ПОЛЕЗНОЕ
ВРЕМЯ
РАЗДРАЖЕНИЯ
ХРОНАКСИЯ
длительность действия электрического тока силой в две реобазы,
необходимая и достаточная для инициации возбуждения
характеризует
постоянную времени мембраны
во многом зависящую от
скорости срабатывания активационных ворот натриевых
каналов на деполяризацию
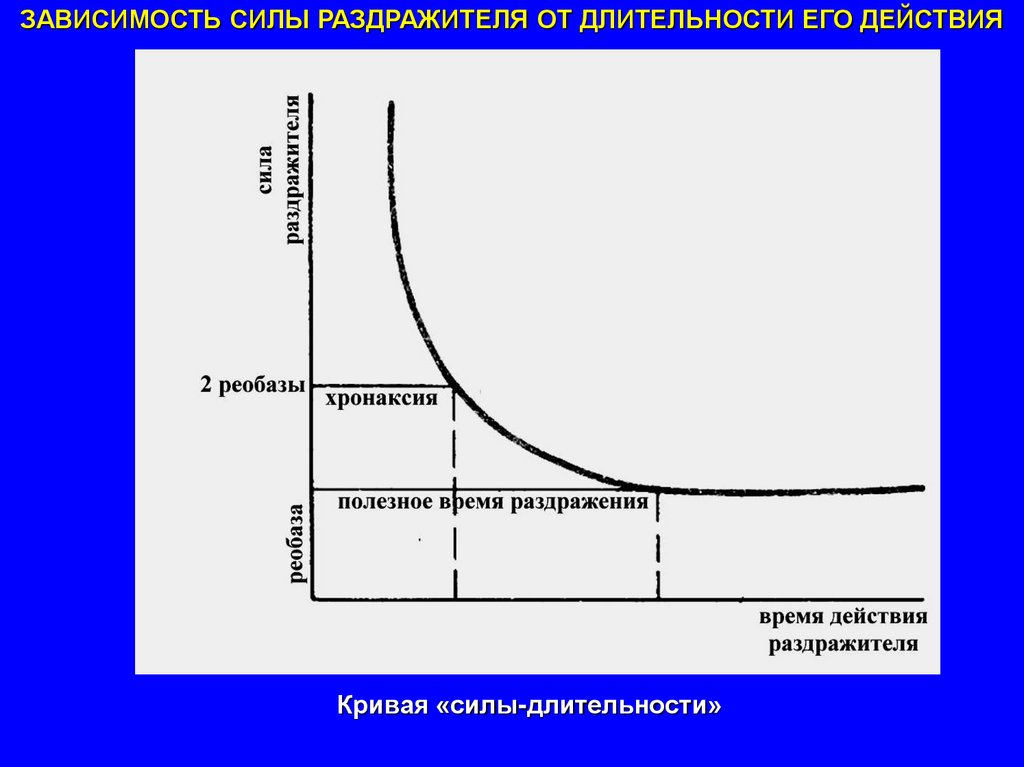
59.
ЗАВИСИМОСТЬ СИЛЫ РАЗДРАЖИТЕЛЯ ОТ ДЛИТЕЛЬНОСТИ ЕГО ДЕЙСТВИЯКривая «силы-длительности»
60.
ОСНОВНЫЕ ЭЛЕКТРОФИЗИОЛОГИЧЕСКИЕ ПАРАМЕТРЫ ВОЗБУДИМЫХСТРУКТУР
ВОЗБУДИМОСТЬ
способность клетки электрически отвечать на действие
раздражителей определенной силы
МЕРА ВОЗБУДИМОСТИ
ПОРОГОВЫЙ ПОТЕНЦИАЛ ( V) возбудимого образования
представляющий собой
разность между мембранным потенциалом покоя (МПП) и
критическим уровнем деполяризации (КУД)
V = МПП – КУД
КРИТИЧЕСКИЙ УРОВЕНЬ
ДЕПОЛЯРИЗАЦИИ (КУД)
мембраны возбудимой структуры
ПОРОГОВАЯ СИЛА
РАЗДРАЖИТЕЛЯ
такая степень деполяризации мембраны
по достижении которой
деполяризация принимает самообновляющийся
характер, и потенциал действия возникает
автоматически
минимальная сила раздражителя, необходимая и достаточная для
того, чтобы он, действуя на возбудимую ткань, разрядил ее
мембрану до критического уровня деполяризации и соответственно
инициировал генерацию потенциала действия
ЛАБИЛЬНОСТЬ
при прочих равных
условиях зависит от
длительности пика потенциала
действия и следовых потенциалов
максимальное количество импульсов, которое возбудимое
образование способно воспроизвести в течение
определенного интервала времени в соответствии с
определенным ритмом раздражения
61.
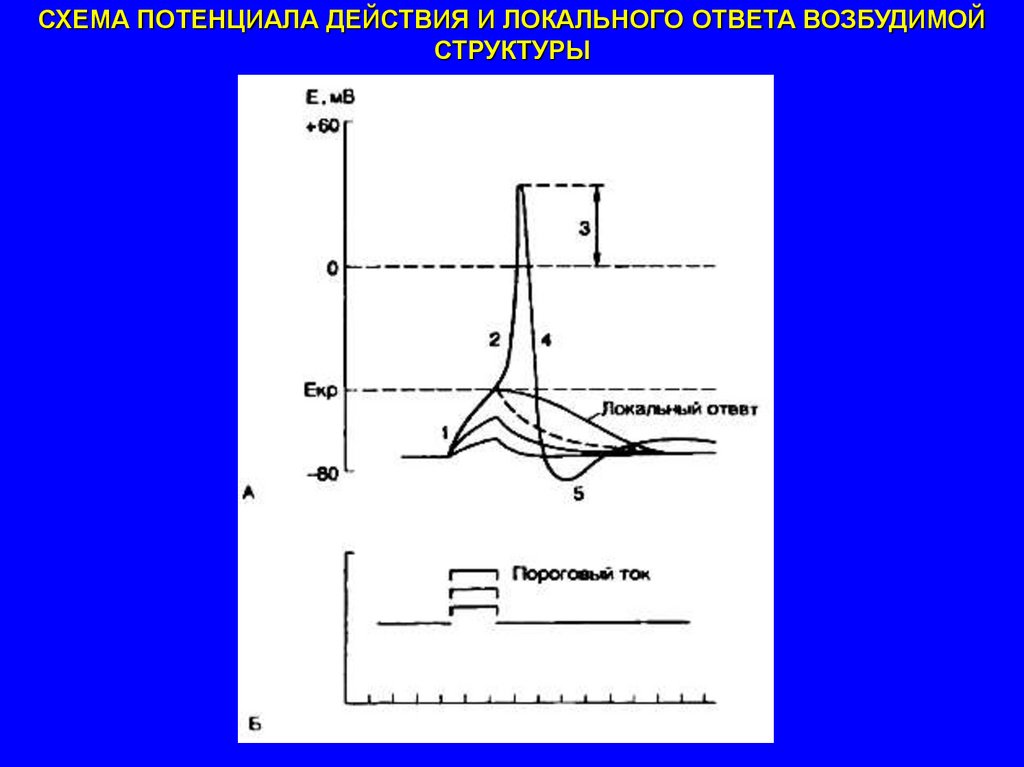
СХЕМА ПОТЕНЦИАЛА ДЕЙСТВИЯ И ЛОКАЛЬНОГО ОТВЕТА ВОЗБУДИМОЙСТРУКТУРЫ
62.
ИЗМЕНЕНИЕ ВОЗБУДИМОСТИ ВОЗБУДИМОЙ СТРУКТУРЫ ПРИКРАТКОВРЕМЕННОЙ ДЕПОЛЯРИЗАЦИИ МЕМБРАНЫ
сопровождается
Кратковременная
уменьшением порогового потенциала
деполяризация
и, как следствие,
мембраны возбудимой
структуры
повышением возбудимости
63.
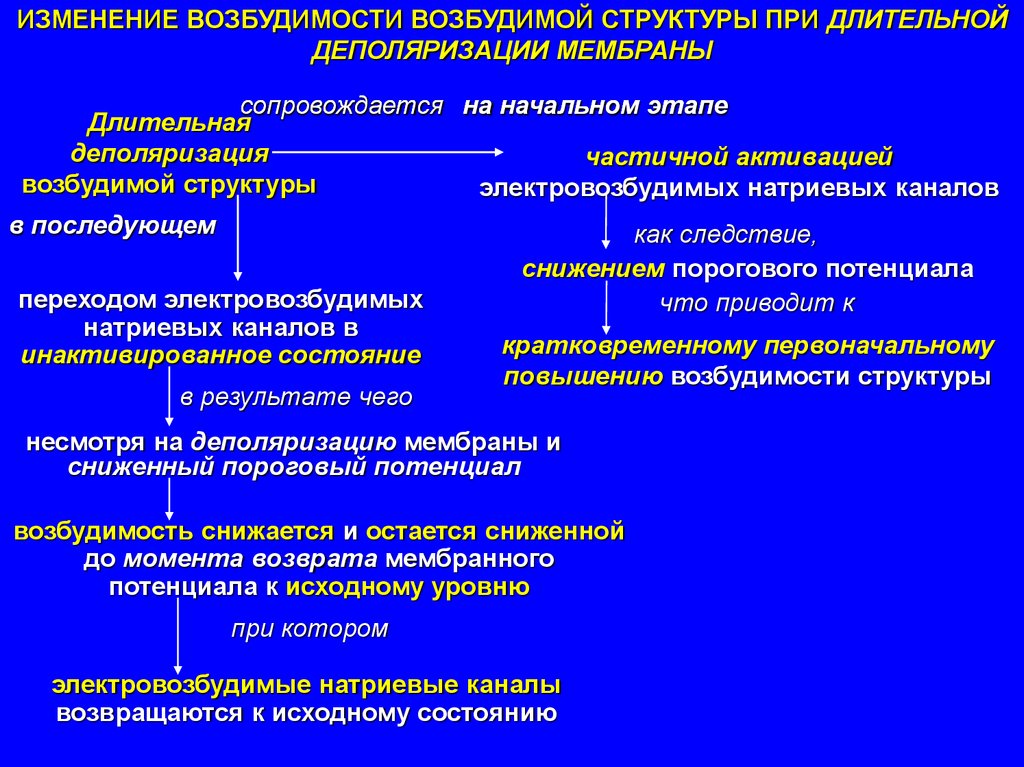
ИЗМЕНЕНИЕ ВОЗБУДИМОСТИ ВОЗБУДИМОЙ СТРУКТУРЫ ПРИ ДЛИТЕЛЬНОЙДЕПОЛЯРИЗАЦИИ МЕМБРАНЫ
сопровождается на начальном этапе
Длительная
деполяризация
частичной активацией
возбудимой структуры
электровозбудимых натриевых каналов
в последующем
как следствие,
переходом электровозбудимых
натриевых каналов в
инактивированное состояние
в результате чего
снижением порогового потенциала
что приводит к
кратковременному первоначальному
повышению возбудимости структуры
несмотря на деполяризацию мембраны и
сниженный пороговый потенциал
возбудимость снижается и остается сниженной
до момента возврата мембранного
потенциала к исходному уровню
при котором
электровозбудимые натриевые каналы
возвращаются к исходному состоянию
64.
ИЗМЕНЕНИЕ ВОЗБУДИМОСТИ ВОЗБУДИМОЙ СТРУКТУРЫ ПРИКРАТКОВРЕМЕННОЙ ГИПЕРПОЛЯРИЗАЦИИ МЕМБРАНЫ
Кратковременная
гиперполяризация
приводит к
первоначальному увеличению порогового
потенциала
и, как следствие,
снижению возбудимости возбудимой
структуры
65.
1. Мембраны всех живых клеток (пока они живы) заряжены (изнутри –отрицательно, снаружи – положительно)
2. Преимущественно этот заряд создается за счет диффузии ионов калия из
клетки наружу по концентрационному градиенту
3. Небольшой вклад в создание заряда на мембране вносит Na+/K+-насос
(благодаря своей электрогенности)
4. Определенная величина мембранного потенциала важна для клетки,
поскольку предопределяет определенную конформацию белков мембраны
и, следовательно, их функциональную активность
5. Для возбудимых тканей определенная величина мембранного потенциала
покоя предопределяет «покой» клетки и ее готовность к возбуждению
6. Способностью активно изменять заряд на своей мембране под действием
раздражителя (электрически отвечать на раздражитель) обладают только
клетки возбудимых тканей (нервной, мышечной и железистой), поскольку в
их мембране имеются электровозбудимые (потенциалзависимые) каналы
7. Электрические ответы необходимы для «коммуницирования» нервных
клеток друг с другом, а также с клетками исполнительных органов, для
нормальной работы некоторых органов (в мышечной ткани они инициируют
сокращение, в железистой – секрецию веществ)
66.
8. Различают 2 типа активных электрических ответов возбудимых структур:потенциал действия и локальные ответы
Потенциал действия
Локальные ответы
Быстрое (от нескольких десятых мс
до нескольких мс, в кардиомиоцитах
– до 300 мс) высокоамплитудное
колебание мембранного потенциала,
имеющее всегда стандартную
(максимально возможную для данной
структуры) амплитуду (от 50 мВ до
140 мВ)
проявляется в начальной быстрой
деполяризации мембраны и
последующей более медленной ее
реполяризации
способен распространяться вдоль
возбудимой структуры на большие
расстояния, не заткхая
обеспечивает возможность проведения
и передачи информации в нервной
системе, инициирует сокращение
мышечной ткани
Локальная
деполяризация
Локальная
гиперполяризация
относительно низкоамплитудные
колебания мембранного
потенциала, в связи с чем не
способны к распространению на
большие расстояния от места
своего возникновения
их амплитуда и длительность
зависят от силы и длительности
вызвавшего их раздражителя
способны к суммации
возникают, как правило, на теле и
дендритах нейрона, делают
возможным интегральную
обработку информации,
поступающей к нейрону через
различные входы