





















 Биология
БиологияПохожие презентации:










Потенциал покоя и действия
1.
Потенциал покоя идействия
2.
Потенциалы покоя и действияВозбудимыми тканями являются нервные и мышечные ткани, которые активируют потенциал
действия, который развивается на клеточной мембране в результате активации и инактивации
ионных каналов.
Классификация и характер ионных каналов:
управляемые. По механизму управления: электро-, хемо- и механоуправляемые;
неуправляемые. Не имеют воротного механизма и всегда открыты, ионы идут постоянно,
но медленно.
Потенциал покоя — это разность электрических потенциалов между наружной и внутренней
средой клетки.
Механизм формирования потенциалов покоя. Непосредственная причина потенциала покоя —
это неодинаковая концентрация анионов и катионов внутри и вне клетки. Во-первых, такое
расположение ионов обосновано разницей проницаемости. Во-вторых, ионов калия выходит из
клетки значительно больше, чем натрия.
3.
Потенциал действия — это возбуждение клетки, быстрое колебание мембранного потенциала вследствие диффузииионов в клетку и из клетки.
При действии раздражителя на клетки возбудимой ткани сначала очень быстро активируются и инактивируются
натриевые каналы, затем с некоторым опозданием активируются и инактивируются калиевые каналы.
Вследствие этого ионы быстро диффундируют в клетку или из нее согласно электрохимическому градиенту. Это и есть
возбуждение. По изменению величин и знака заряда клетки выделяют три фазы:
1-я фаза — деполяризация. Уменьшение заряда клетки до нуля. Натрий движется к клетке согласно
концентрационному и электрическому градиенту. Условие движения: открыты ворота натриевого канала;
2-я фаза — инверсия. Изменение знака заряда на противоположный. Инверсия предполагает две части:
восходящую и нисходящую.
Восходящая часть. Натрий продолжает двигаться в клетку согласно концентрационному градиенту, но вопреки
электрическому градиенту (он препятствует).
Нисходящая часть. Калий начинает выходить из клетки согласно концентрационному и электрическому градиенту.
Открыты ворота калиевого канала;
3-я фаза — реполяризация. Калий продолжает выходить из клетки согласно концентрационному, но вопреки
электрическому градиенту.
4.
Критерии возбудимостиПри развитии потенциала действия происходит изменение возбудимости ткани. Это изменение протекает по фазам. Состояние исходной
поляризации мембраны характерно отражает мембранный потенциал покоя, которому соответствует исходное состояние возбудимости а,
следовательно, исходное состояние возбудимой клетки. Это нормальный уровень возбудимости. Период предспайка — период самого начала
потенциала действия. Возбудимость ткани слегка повышена. Эта фаза возбудимости — первичная экзальтация (первичная супернормальная
возбудимость). Во время развития предспайка мембранный потенциал приближается к критическому уровню деполяризации и для достижения
этого уровня сила раздражителя может быть меньше пороговой.
В период развития спайка (пикового потенциала) идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит
перезарядка мембраны, и она утрачивает способность отвечать возбуждением на раздражители сверхпороговой силы. Эта фаза возбудимости
получила название абсолютной рефрактерности, т.е. абсолютной невозбудимости, которая длится до конца перезарядки мембраны. Абсолютная
рефрактерность мембраны возникает в связи с тем, что натриевые каналы полностью открываются, а затем инактивируются.
После окончания фазы перезарядки возбудимость ее постепенно восстанавливается до исходного уровня — это фаза относительной
рефрактерности, т.е. относительной невозбудимости. Она продолжается до восстановления заряда мембраны до величины, соответствующей
критическому уровню деполяризации. Поскольку в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани
понижена, и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя. Снижение возбудимости в фазу
относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых каналов.
Следующему периоду соответствует повышенный уровень возбудимости: фаза вторичной экзальтации или вторичной супернормальной
возбудимости. Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации, по сравнению с состоянием покоя
исходной поляризации, то порог раздражения снижен, т.е. возбудимость клетки повышена. В эту фазу новое возбуждение может возникнуть при
действии раздражителей подпороговой силы. Натриевые каналы в эту фазу инактивированы не полностью. Мембранный потенциал
увеличивается — возникает состояние гиперполяризации мембраны. Удаляясь от критического уровня деполяризации, порог раздражения слегка
повышается, и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины.
5.
Механизм возникновения мембранного потенциала покояКаждая клетка в состоянии покоя характеризуется наличием трансмембранной разности потенциалов
(потенциала покоя). Обычно разность зарядов между внутренней и внешней поверхностями мембран
составляет от -80 до -100 мВ и может быть измерена с помощью наружного и внутриклеточного
микроэлектродов (рис. 1).
Разность потенциалов между наружной и внутренней сторонами мембраны клетки в состоянии ее покоя
называют мембранным потенциалом (потенциалом покоя).
Создание потенциала покоя обеспечивается двумя основными процессами — неравномерным
распределением неорганических ионов между внутри- и внеклеточным пространством и неодинаковой
проницаемостью для них клеточной мембраны. Анализ химического состав вне- и внутриклеточной
жидкости свидетельствует о крайне неравномерном распределении ионов (табл. 1).
В состоянии покоя внутри клетки много анионов органических кислот и ионов К+, концентрация которых
в 30 раз больше, чем снаружи; ионов Na+, наоборот, снаружи клетки в 10 раз больше, чем внутри; СIтакже больше снаружи.
В покое мембрана нервных клеток наиболее проницаема для К+, менее — для СI- и очень мало
проницаема для Na+/ Проницаемость мембраны нервного волокна для Na+ B покое в 100 раз меньше,
чем для K+. Для многих анионов органических кислот мембрана в покое совсем непроницаема.
6.
Рис. 1. Измерение потенциала покоя мышечноговолокна (А) с помощью внутриклеточного
микроэлектрода:
М — микрозлектрод;
И — индифферентный электрод.
Луч на экране осциллографа (В) показывает, что до
прокола мембраны микроэлектродом разность
потенциалов между М и И была равна нулю.
В момент прокола (показан стрелкой) обнаружена
разность потенциалов, указывающая, что внутренняя
сторона мембраны заряжена отрицательно по
отношению к ее наружной поверхности (по Б.И.
Ходорову)
7.
Таблица . Внутри- и внеклеточные концентрации ионов мышечной клетки теплокровногоживотного, ммоль/л (по Дж. Дудел)
Ионы
Внутриклеточная
концентрация
Внеклеточная
концентрация
Na+
12
145
К+
155
4
CI-
4
120
нсо3-
8
27
А- (анионы органических
соединений)
155
-
8.
В силу градиента концентраций К+ выходит на наружную поверхность клетки, вынося свой положительный заряд.Высокомолекулярные анионы не могут следовать за К+ из-за непроницаемости для них мембраны. Ион Na+ также не может
возместить ушедшие ионы калия, ибо проницаемость мембраны для него значительно меньше. СI- по градиенту концентраций
может перемешаться только внутрь клетки, увеличивая тем самым отрицательный заряд внутренней поверхности мембраны.
Вследствие такого перемещения ионов возникает поляризация мембраны, когда наружная ее поверхность заряжается
положительно, а внутренняя — отрицательно.
Электрическое поле, которое создастся на мембране, активно вмешивается в распределение ионов между внутренним и
наружным содержимым клетки. По мере возрастания положительного заряда на наружной поверхности клетки иону К+ как
положительно заряженному становится все труднее перемещаться изнутри наружу. Он движется как бы в гору. Чем больше
величина положительного заряда на наружной поверхности, тем меньшее количество ионов К+ может выходить на
поверхность клетки. При определенной величине потенциала на мембране количество ионов К+, пересекающих мембрану в
том и другом направлении, оказывается равным, т.е. концентрационный градиент калия уравновешивается имеющимся на
мембране потенциалом. Потенциал, при котором диффузионный поток ионов становится равным потоку одноименных ионов,
идущих в обратном направлении, называют потенциалом равновесия для данного иона. Для ионов К+ потенциал равновесия
равен -90 мВ. В миелинизированных нервных волокнах величина потенциала равновесия для ионов СI- близка к значению
мембранного потенциала покоя (-70 мВ). Поэтому, несмотря на то что концентрация ионов СI- снаружи волокна больше, чем
внутри его, не отмечается их одностороннего тока в соответствии с градиентом концентраций. В этом случае разность
концентраций сбалансирована потенциалом, имеющимся на мембране.
Ион Na+ по градиенту концентраций должен был бы входить внутрь клетки (его потенциал равновесия составляет +60 мВ), и
наличие отрицательного заряда внутри клетки не должно было бы препятствовать этому потоку. В этом случае входящий Na+
нейтрализовал бы отрицательные заряды внутри клетки. Однако этого в действительности не происходит, так как мембрана в
покое малопроницаема для Na+.
9.
• Важнейшим механизмом, поддерживающим низкую внутриклеточную концентрацию ионов Na+ и высокую концентрациюионов К+, является натрий-калиевый насос (активный транспорт). Известно, что в клеточной мембране имеется система
переносчиков, каждый из которых связывается стремя находящимися внутри клетки ионами Na+ и выводит их наружу. С
наружной стороны переносчик связывается с двумя находящимися вне клетки ионами К+ которые переносятся в цитоплазму.
Энергообеспечение работы систем переносчиков обеспечивается АТФ. Функционирование насоса по такой системе приводит
к следующим результатам:
поддерживается высокая концентрация ионов К+ внутри клетки, что обеспечивает постоянство величины потенциала покоя.
Вследствие того что за один цикл обмена ионов из клетки выводится на один положительный ион больше, чем вводится,
активный транспорт играет роль в создании потенциала покоя. В этом случае говорят об электрогенном насосе, поскольку
он сам создает небольшой, но постоянный ток положительных зарядов из клетки, а потому вносит прямой вклад в
формирование отрицательного потенциала внутри нее. Однако величина вклада электрогенного насоса в общее значение
потенциала покоя обычно невелика и составляет несколько милливольт;
поддерживается низкая концентрация ионов Na+ внутри клетки, что, с одной стороны, обеспечивает работу механизма
генерации потенциала действия, с другой — обеспечивает сохранение нормальных осмолярности и объема клетки;
поддерживая стабильный концентрационный градиент Na+, натрий-калиевый насос способствует сопряженному К+, Na+ транспорту аминокислот и Сахаров через клеточную мембрану.
• Таким образом, возникновение трансмембранной разности потенциалов (потенциала покоя) обусловлено высокой
проводимостью клеточной мембраны в состоянии покоя для ионов К+, СI-, ионной асимметрией концентраций ионов К+ и
ионов СI-, работой систем активного транспорта (Na+/K+ -АТФаза), которые создают и поддерживают ионную асимметрию.
10.
Потенциал действия нервного волокна, нервный импульсПотенциал действия — это кратковременное колебание разности потенциалов мембраны возбудимой клетки,
сопровождающееся изменением ее знака заряда.
Потенциал действия является основным специфическим признаком возбуждения. Его регистрация свидетельствует о том,
что клетка или ее структуры ответили на воздействие возбуждением. Однако, как уже отмечалось, ПД в некоторых
клетках может возникать спонтанно (самопроизвольно). Такие клетки содержатся в водителях ритма сердца, стенках
сосудов, нервной системе. ПД используется как носитель информации, передающий ее в виде электрических сигналов
(электрическая сигнализации) по афферентным и эфферентным нервным волокнам, проводящей системе сердца, а также
для инициирования сокращения мышечных клеток.
Рассмотрим причины и механизм генерации ПД в афферентных нервных волокнах, образующих первично
воспринимающие сенсорные рецепторы. Непосредственной причиной возникновения (генерации) ПД в них является
рецепторный потенциал.
Если измерять разность потенциалов на мембране ближайшего к нервному окончанию перехвата Ранвье, то в
промежутках между воздействиями на капсулу тельца Пачини она остается неизменной (70 мВ), а во время воздействия
деполяризуется почти одновременно с деполяризацией рецепторной мембраны нервного окончания.
При увеличении силы давления на тельце Пачини, вызывающей возрастание рецепторного потенциала до 10 мВ, в
ближайшем перехвате Ранвье обычно регистрируется быстрое колебание мембранного потенциала, сопровождающееся
перезарядкой мембраны — потенциал действия (ПД), или нервный импульс (рис. 2). Если сила давления на тельце
возрастет еще больше, амплитуда рецепторного потенциала увеличивается и в нервном окончании генерируется уже ряд
потенциалов действия с определенной частотой.
11.
Рис. 2. Схематическое представление механизма преобразования рецепторного потенциала в потенциалдействия (нервный импульс) и распространения импульса по нервному волокну
12.
Суть механизма генерации ПД состоит в том, что рецепторный потенциал вызывает возникновение локальных круговых токовмежду деполяризованной рецепторной мембраной немиелинизированной части нервного окончания и мембраной первого
перехвата Ранвье. Эти токи, носителями которых являются ионы Na+, К+, СI- и другие минеральные ионы, «протекают» не
только вдоль, но и поперек мембраны нервного волокна в области перехвата Ранвье. В мембране перехватов Ранвье в отличие
от рецепторной мембраны самого нервного окончания имеется большая плотность ионных потенциалзависимых натриевых и
калиевых каналов.
При достижении на мембране перехвата Ранвье величины деполяризации около 10 мВ происходит открытие быстрых
потенциалзависимых натриевых каналов и через них в аксоплазму по электрохимическому градиенту устремляется поток
ионов Na+. Он обусловливает быструю деполяризацию и перезарядку мембраны перехвата Ранвье. Однако одновременно с
открытием быстрых потенциалзависимых натриевых каналов в мембране перехвата Ранвье открываются медленные
потенциалзависимые калиевые каналы и из аксоилазмы начинают выходить ионы К+ Их выход запаздывает по отношению ко
входу ионов Na+. Таким образом, входящие с большой скоростью в аксоплазму ионы Na+ быстро деполяризуют и
перезаряжают на короткое время (0,3-0,5 мс) мембрану, а выходящие ионы К+ восстанавливают исходное распределение
зарядов на мембране (реполяризуют мембрану). В результате во время механического воздействия на тельце Пачини силой,
равной или превышающей пороговую, на мембране ближайшего перехвата Ранвье наблюдается кратковременное колебание
потенциала в виде быстрой деполяризации и реполяризации мембраны, т.е. генерируется ПД (нервный импульс).
Поскольку непосредственной причиной генерации ПД является рецепторный потенциал, то его в этом случае еще называют
генераторным потенциалом. Число генерируемых в единицу времени одинаковых по амплитуде и длительности нервных
импульсов пропорционально амплитуде рецепторного потенциала, а следовательно, силе давления на рецептор. Процесс
преобразования информации о силе воздействия, заложенной в амплитуде рецепторного потенциала, в число дискретных
нервных импульсов получил название дискретного кодирования информации.
Более подробно ионные механизмы и временная динамика процессов генерации ПД изучены в экспериментальных условиях
при искусственном воздействии на нервное волокно электрическим током различной силы и длительности.
13.
Природа потенциала действия нервного волокна (нервного импульса)Мембрана нервного волокна в точке локализации раздражающего электрода отвечает на воздействие
очень слабого тока, еще не достигшего порогового значения. Этот ответ получил название локального, а
колебание разности потенциалов на мембране — локального потенциала.
Локальный ответ на мембране возбудимой клетки может предшествовать возникновению потенциала
действия или возникать как самостоятельный процесс. Он представляет собой кратковременное
колебание (деполяризация и реполяризация) потенциала покоя, не сопровождающееся перезарядкой
мембраны. Деполяризация мембраны при развитиии локального потенциала обусловлена
опережающим входом в аксоплазму ионов Na+, а реполяризация — запаздывающим выходом из
аксоплазмы ионов К+.
Если воздействовать на мембрану электрическим током возрастающей силы, то при се величине,
называемой пороговой, деполяризация мембраны может достигнуть критического уровня — Ек, при
котором происходит открытие быстрых потенциалзависимых натриевых каналов. В результате через них
происходит лавинообразно нарастающее поступление в клетку ионов Na+. Вызываемый процесс
деполяризации приобретает самоускоряющийся характер, и локальный потенциал перерастает в
потенциал действия.
Уже упоминалось, что характерным признаком ПД является кратковременная инверсия (перемена) знака
заряда на мембране. Снаружи она на короткое время (0,3-2 мс) становится заряженной отрицательно, а
внутри — положительно. Величина инверсии может составлять до 30 мВ, а величина всего потенциала
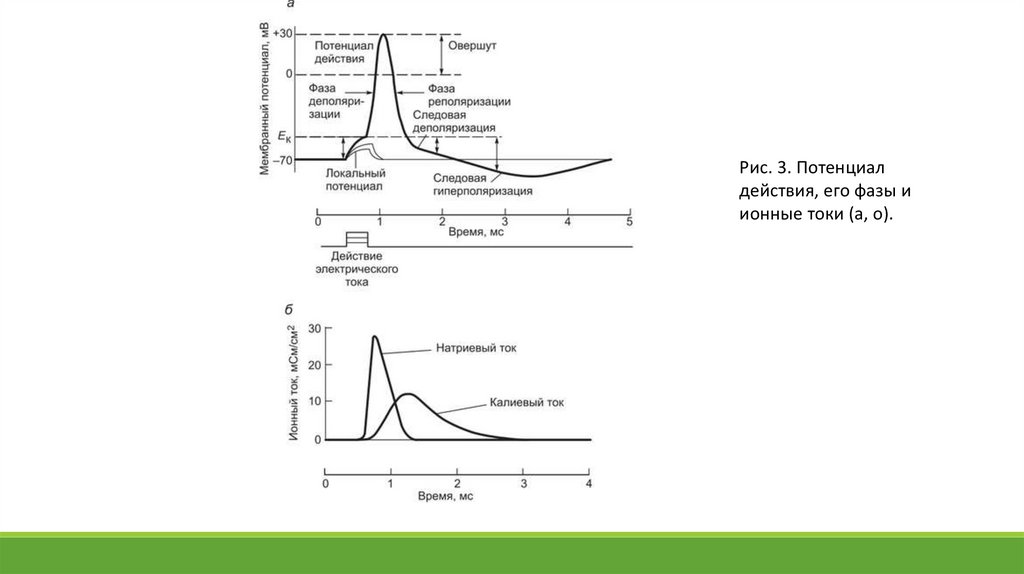
действия — 60-130 мВ (рис. 3).
14.
Таблица. Сравнительная характеристика локального потенциала и потенциала действияХарактеристика
Локальный потенциал
Потенциал действия
Проводимость
Распространяется местно, на 1-2 мм с
затуханием (декрементом)
Распространяется без затухания на
большие расстояния по всей длине
нервного волокна
Закон «силы»
Подчиняется
Не подчиняется
Закон «все или ничего»
Не подчиняется
Подчиняется
Явление суммации
Суммируется, возрастает при
повторных частых подпороговых
раздражениях
Не суммируется
Величина амплитуды
10-40 мВ
80-130 мВ
Способность к возбудимости
Увеличивается
Уменьшается вплоть до полной
невозбудимости (рефрактерность)
Величина раздражителя
Подпороговая
Пороговая и сверхпороговая
15.
Потенциал действия в зависимости от характера изменения зарядов на внутренней поверхности мембраныподразделяют на фазы деполяризации, реполяризации и гиперполяризации мембраны. Деполяризацией называют всю
восходящую часть ПД, на которой выделяют участки, соответствующие локальному потенциалу (от уровня Е0 до Ек),
быстрой деполяризации (от уровня Ек до уровня 0 мВ), инверсии знака заряда (от 0 мВ до пикового значения или начала
реполяризации). Реполяризацией называют нисходящую часть ПД, которая отражает процесс восстановления исходной
поляризации мембраны. Вначале реполяризация осуществляется быстро, но, приближаясь к уровню Е0, скорость се
может замедляться и этот участок называют следовой отрицательностью (или следовым отрицательным потенциалом). У
некоторых клеток вслед за реполяризацией развивается гиперполяризация (возрастание поляризации мембраны). Ее
называют следовым положительным потенциалом.
Начальную высокоамплитудную быстропротекающую часть ПД называют также пик, или спайк. Он включает фазы
деполяризации и быстрой реполяризации.
В механизме развития ПД важнейшая роль принадлежит потенциалзависимым ионным каналам и неодновременному
увеличению проницаемости клеточной мембраны для ионов Na+ и К+. Так, при действии на клетку электрического тока
он вызывает деполяризацию мембраны и, когда заряд мембраны уменьшается до критического уровня (Ек), открываются
потенциалзависимые натриевые каналы. Как уже упоминалось,эти каналы образованы встроенными в мембрану
белковыми молекулами, внутри которых имеются пора и два воротных механизма. Один из воротных механизмов —
активационный обеспечивает (при участии сегмента 4) открытие (активацию) канала при деполяризации мембраны, а
второй (при участии внутриклеточной петли между 3-м и 4-м доменами) — его инактивацию, развивающуюся при
перезарядке мембраны (рис. 4). Поскольку оба этих механизма быстро изменяют положение ворот канала, то
потенциалзависимые натриевые каналы являются быстрыми ионными каналами. Это обстоятельство имеет
определяющее значение для генерации ПД в возбудимых тканях и для его проведения по мембранам нервных и
мышечных волокон.
16.
17.
Рис. 3. Потенциалдействия, его фазы и
ионные токи (а, о).
18.
19.
Рис. 4. Положение вороти состояние активности
потенциалзависимых
натриевого и калиевого
каналов при различных
уровнях поляризации
мембраны
20.
21.
Чтобы потенциалзависимый натриевый канал мог пропускать внутрь клетки ионы Na+, необходимо открыть лишь активационные ворота,поскольку инактивационные в условиях покоя открыты. Это и происходит, когда деполяризация мембраны достигает уровня Ек (рис. 3, 4).
Открытие активационных ворот натриевых каналов приводит к лавинообразному вхождению натрия внутрь клетки, движимому действием сил
его электрохимического градиента. Поскольку ионы Na+ несут положительный заряд, то они нейтрализуют избыток отрицательных зарядов на
внутренней поверхности мембраны, снижают разность потенциалов на мембране и деполяризуют ее. Вскоре ионы Na+ придают внутренней
поверхности мембраны избыток положительных зарядов, что сопровождается инверсией (сменой) знака заряда с отрицательного на
положительный.
Однако натриевые каналы остаются открытыми лишь около 0,5 мс и через этот промежуток времени от момента начала ПД закрываются
инактивационные ворота, натриевые каналы становятся инактивированными и непроницаемыми для ионов Na+, поступление которых внутрь
клетки резко ограничивается.
С момента деполяризации мембраны до уровня Ек наблюдаются также активация калиевых каналов и открытие их ворот для ионов К+. Ионы К+
под действием сил концентрационного градиента выходят из клетки, вынося из нее положительные заряды. Однако воротный механизм
калиевых каналов является медленно функционирующим и скорость выхода положительных зарядов с ионами К+ из клетки наружу запаздывает
по отношению ко входу ионов Na+. Поток ионов К+, удаляя из клетки избыток положительных зарядов, обусловливает восстановление на
мембране исходного распределения зарядов или ее реполяризацию, и на се внутренней стороне через мгновение от момента перезарядки
восстанавливается отрицательный заряд.
Возникновение ПД на возбудимых мембранах и последующее восстановление исходного потенциала покоя на мембране оказываются
возможными потому, что динамика входа в клетку и выхода из клетки положительных зарядов ионов Na+ и К+ различна. Вход иона Na+ по
времени опережает выход иона К+. Если бы эти процессы были равновесными, то разность потенциалов на мембране не изменялась бы.
Развитие способности к возбуждению и генерации ПД возбудимыми мышечными и нервными клетками было обусловлено формированием в их
мембране двух типов разноскоростных ионных каналов — быстрых натриевых и медленных калиевых.
22.
• Для генерации одиночного ПД требуется поступление в клетку относительно небольшого числа ионовNa+, которое не нарушает его распределения вне и внутри клетки. При генерации большого числа ПД
распределение ионов по обе стороны мембраны клетки могло бы нарушиться. Однако в нормальных
условиях это предотвращается работой Na+, К+ -насоса.
• В естественных условиях в нейронах ЦНС потенциал действия первично возникает в области аксонного
холмика, в афферентных нейронах — в ближайшем к сенсорному рецептору перехвате Ранвье нервного
окончания, т.е. в тех участках мембраны, где имеются быстрые селективные потенциалзависимые
натриевые каналы и медленные калиевые каналы. В других типах клеток (например, пейсмекерных,
гладких миоцитах) в возникновении ПД играют роль не только натриевые и калиевые, но и кальциевые
каналы.
• Механизмы восприятия и преобразования в ПД сигналов во вторично чувствующих сенсорных
рецепторах отличаются от механизмов, разобранных для первично чувствствующих рецепторов. В этих
рецепторах восприятие сигналов осуществляется специализированными нейросенсорными
(фоторецепторные, обонятельные) или сенсоэпителиальными (вкусовые, слуховые, вестибулярные)
клетками. В каждой из этих чувствительных клеток имеется свой, особый механизм восприятия
сигналов. Однако во всех клетках энергия воспринимаемого сигнала (раздражителя) преобразуется в
колебание разности потенциалов плазматической мембраны, т.е. в рецепторный потенциал.
23.
• Таким образом, ключевым моментом в механизмах преобразования сенсорными клетками воспринимаемыхсигналов в рецепторный потенциал является изменение проницаемости ионных каналов в ответ на
воздействие. Открытие Na+, Са2+, К+ -ионных каналов при восприятии и преобразовании сигнала достигается в
этих клетках при участии G-белков, вторых внутриклеточных посредников, связывании с лигандами,
фосфорилировании ионных каналов. Как правило, возникший в сенсорных клетках рецепторный потенциал
вызывает высвобождение из них в синаптическую щель нейромедиатора, который обеспечивает передачу
сигнала на постсинаптическую мембрану афферентного нервного окончания и генерацию на его мембране
нервного импульса. Эти процессы подробно описаны в главе, посвященной сенсорным системам.
• Потенциал действия может быть охарактеризован амплитудой и продолжительностью, которые для одного и
того же нервного волокна остаются одинаковыми при распространении ПД по волокну. Поэтому потенциал
действия называют дискретным потенциалом.
• Между характером воздействия на сенсорные рецепторы и числом ПД, возникших в афферентном нервном
волокне в ответ на воздействие, имеется определенная связь. Она заключается в том, что на большие но силе
или продолжительности воздействия в нервном волокне формируется большее число нервных импульсов, т.е.
при усилении воздействия в нервную систему будут посылаться от рецептора импульсы большей частоты.
Процессы преобразования информации о характере воздействия в частоту и другие параметры нервных
импульсов, передаваемых в ЦНС, получили название дискретного кодирования информации.