



 Биология
БиологияПохожие презентации:

")








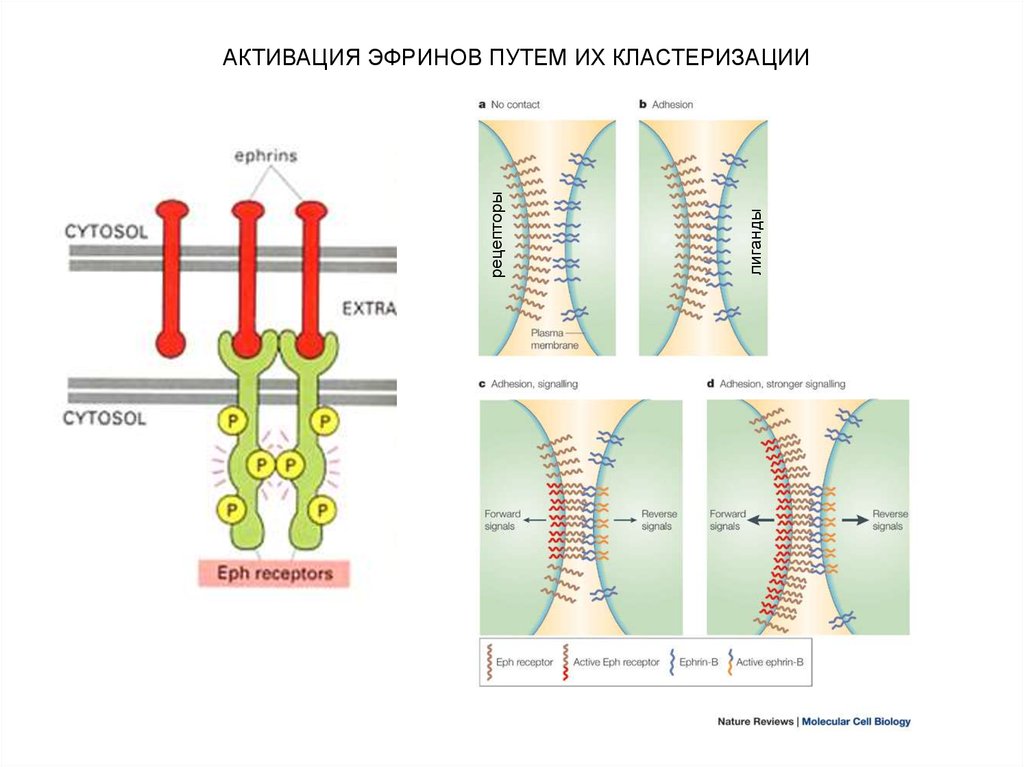
Активизация эфринов путем их кластеризации
1.
лигандырецепторы
АКТИВАЦИЯ ЭФРИНОВ ПУТЕМ ИХ КЛАСТЕРИЗАЦИИ
2.
ДВУНАПРАВЛЕННАЯ СИГНАЛИЗАЦИЯ ЧЕРЕЗ ЭФРИНЫОбратная сигнализация
Прямая сигнализация
3.
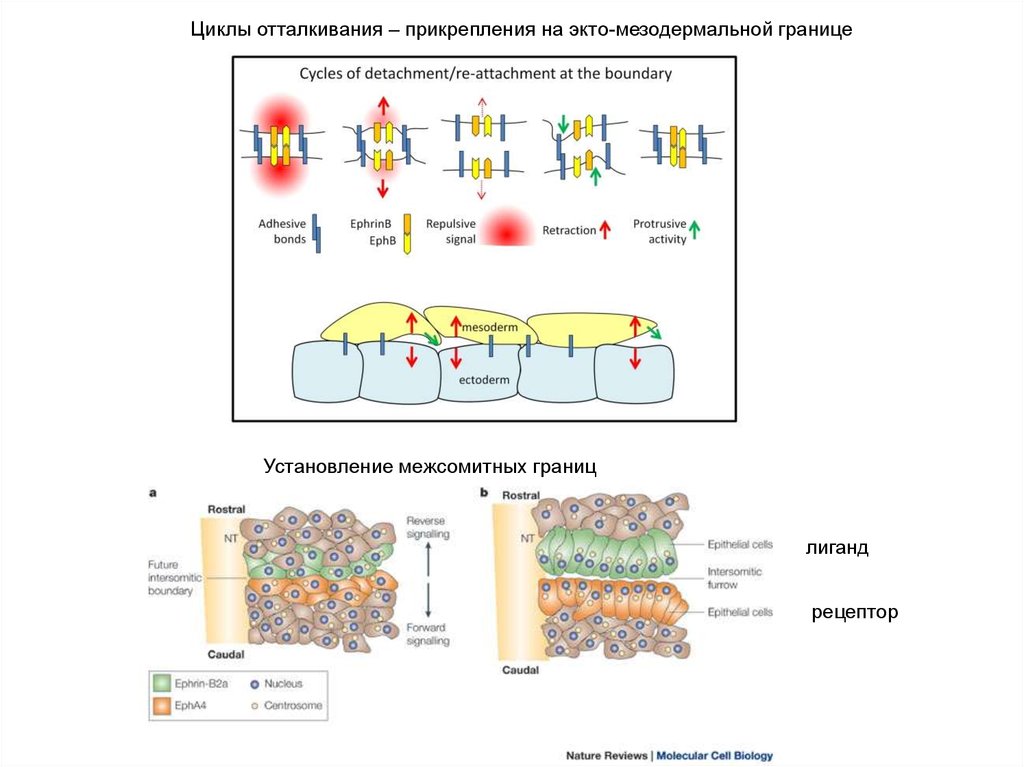
Циклы отталкивания – прикрепления на экто-мезодермальной границеУстановление межсомитных границ
лиганд
рецептор
4.
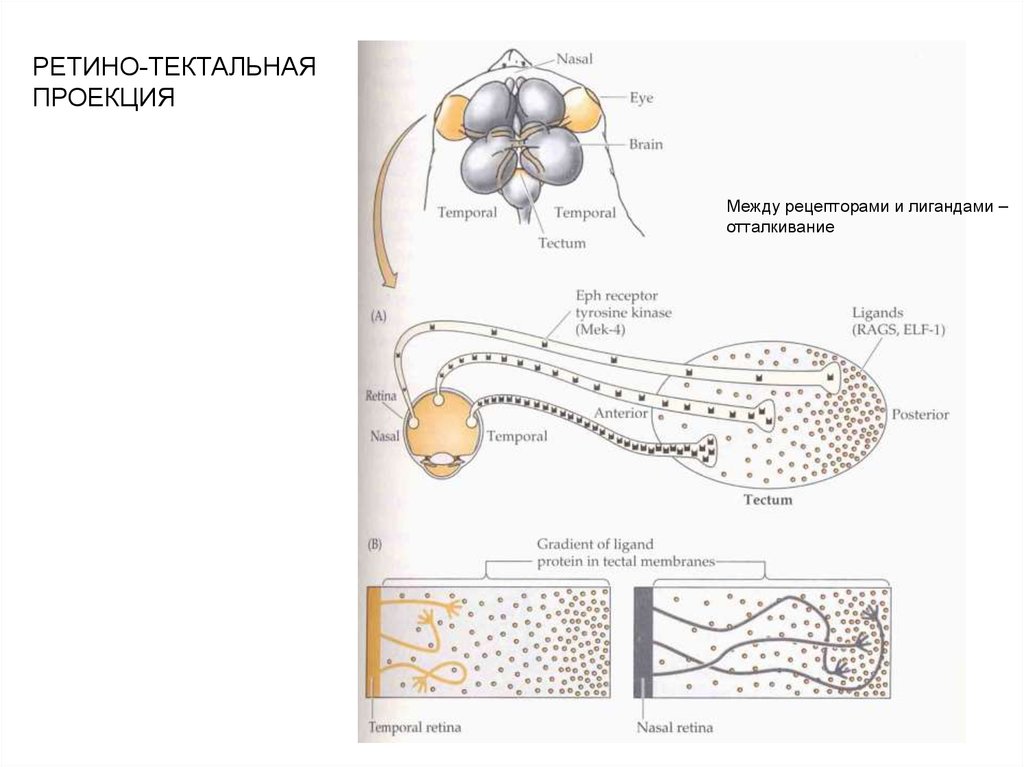
РЕТИНО-ТЕКТАЛЬНАЯПРОЕКЦИЯ
Между рецепторами и лигандами –
отталкивание
5.
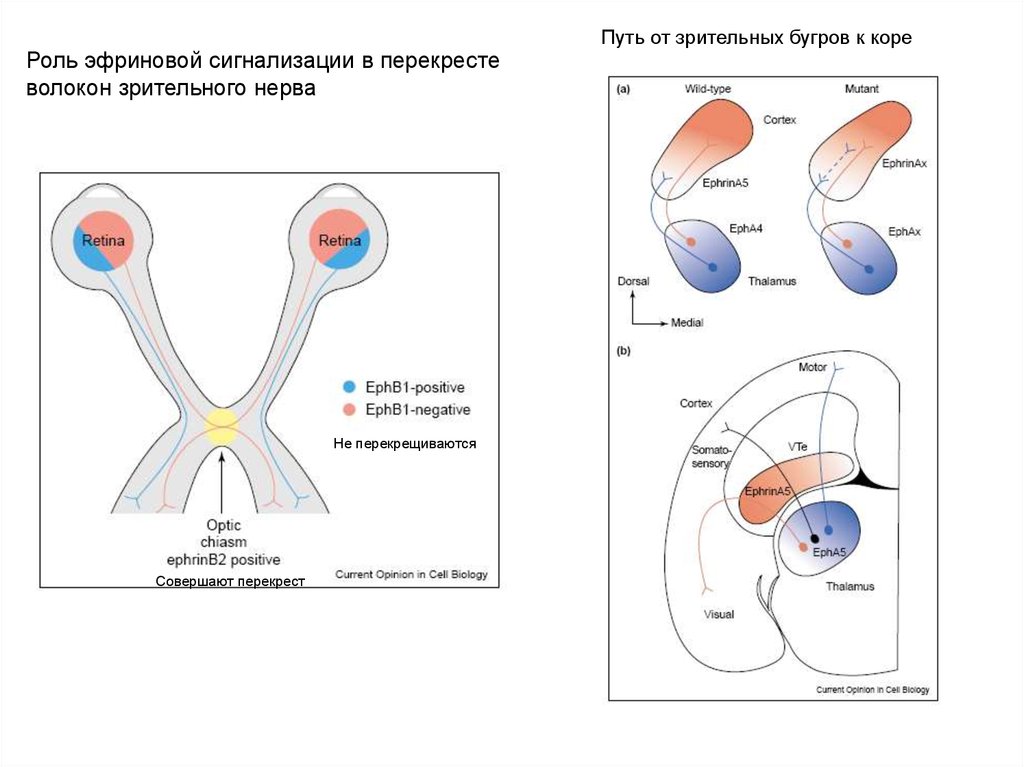
Роль эфриновой сигнализации в перекрестеволокон зрительного нерва
Не перекрещиваются
Совершают перекрест
Путь от зрительных бугров к коре
6.
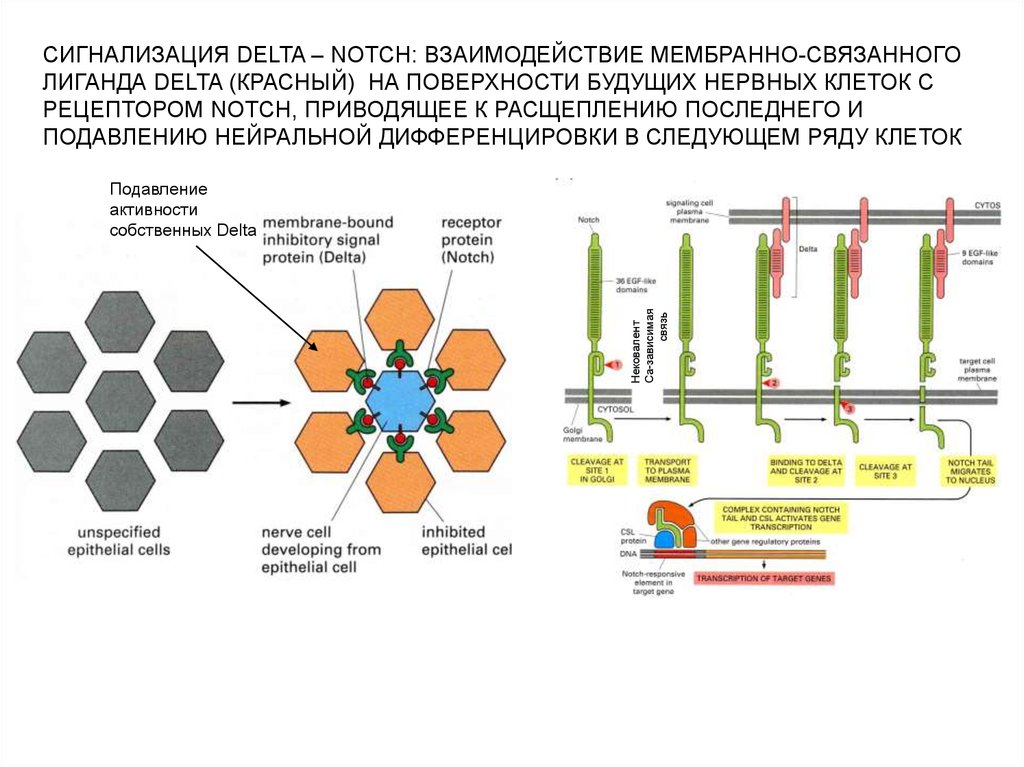
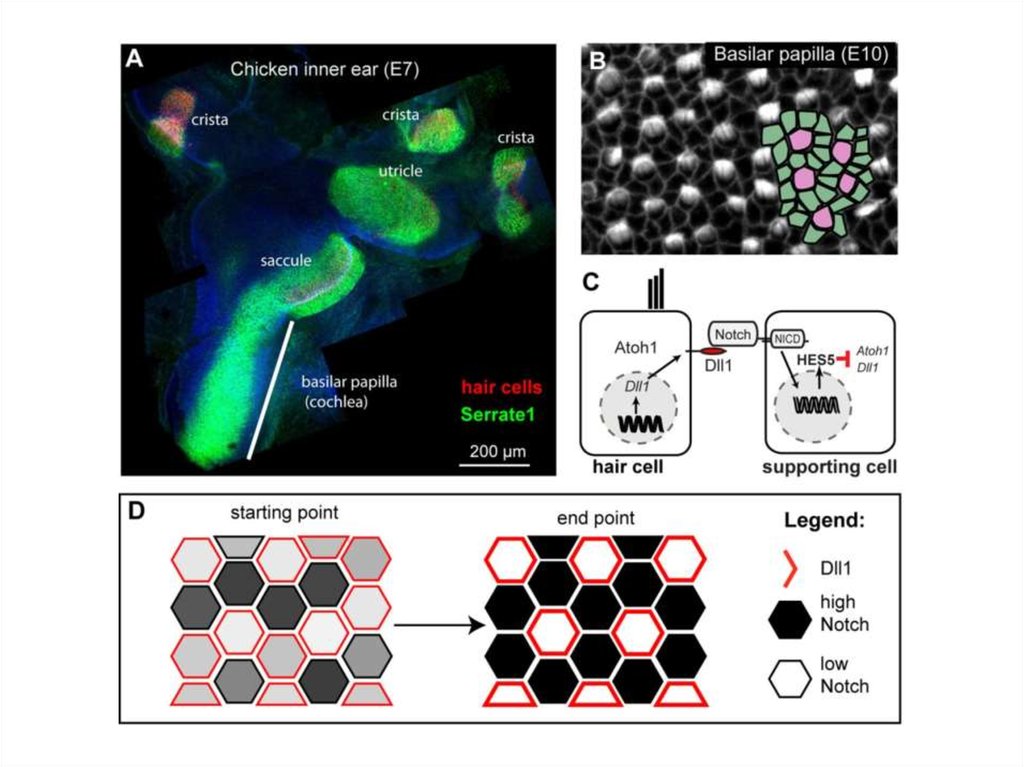
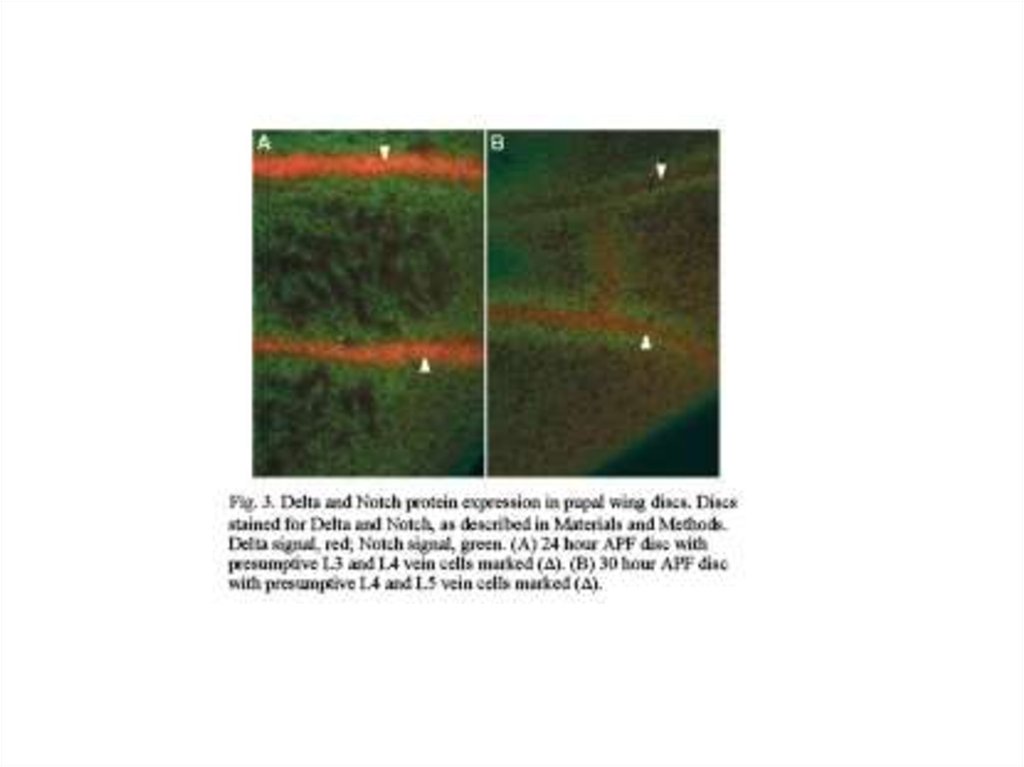
СИГНАЛИЗАЦИЯ DELTA – NOTCH: ВЗАИМОДЕЙСТВИЕ МЕМБРАННО-СВЯЗАННОГОЛИГАНДА DELTA (КРАСНЫЙ) НА ПОВЕРХНОСТИ БУДУЩИХ НЕРВНЫХ КЛЕТОК С
РЕЦЕПТОРОМ NOTCH, ПРИВОДЯЩЕЕ К РАСЩЕПЛЕНИЮ ПОСЛЕДНЕГО И
ПОДАВЛЕНИЮ НЕЙРАЛЬНОЙ ДИФФЕРЕНЦИРОВКИ В СЛЕДУЮЩЕМ РЯДУ КЛЕТОК
Нековалент
Са-зависимая
связь
Подавление
активности
собственных Delta
7.
8.
9.
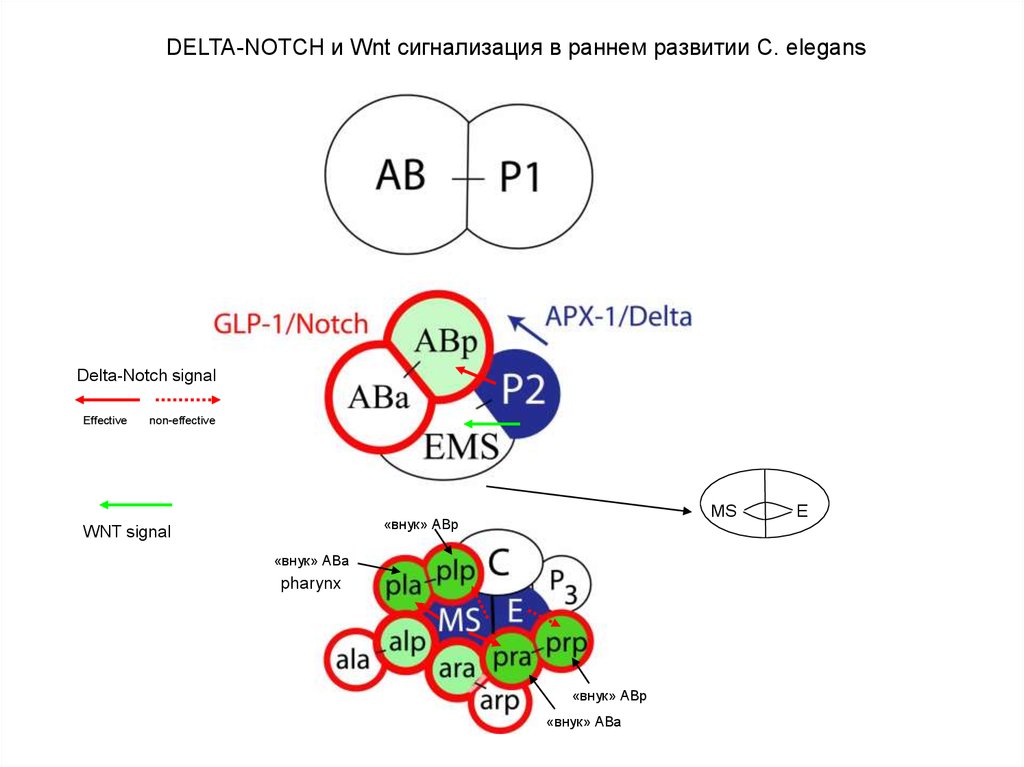
DELTA-NOTCH и Wnt сигнализация в раннем развитии C. elegansDelta-Notch signal
Effective
non-effective
MS
«внук» АВp
WNT signal
«внук» АВа
pharynx
«внук» АВp
«внук» АВа
E
10.
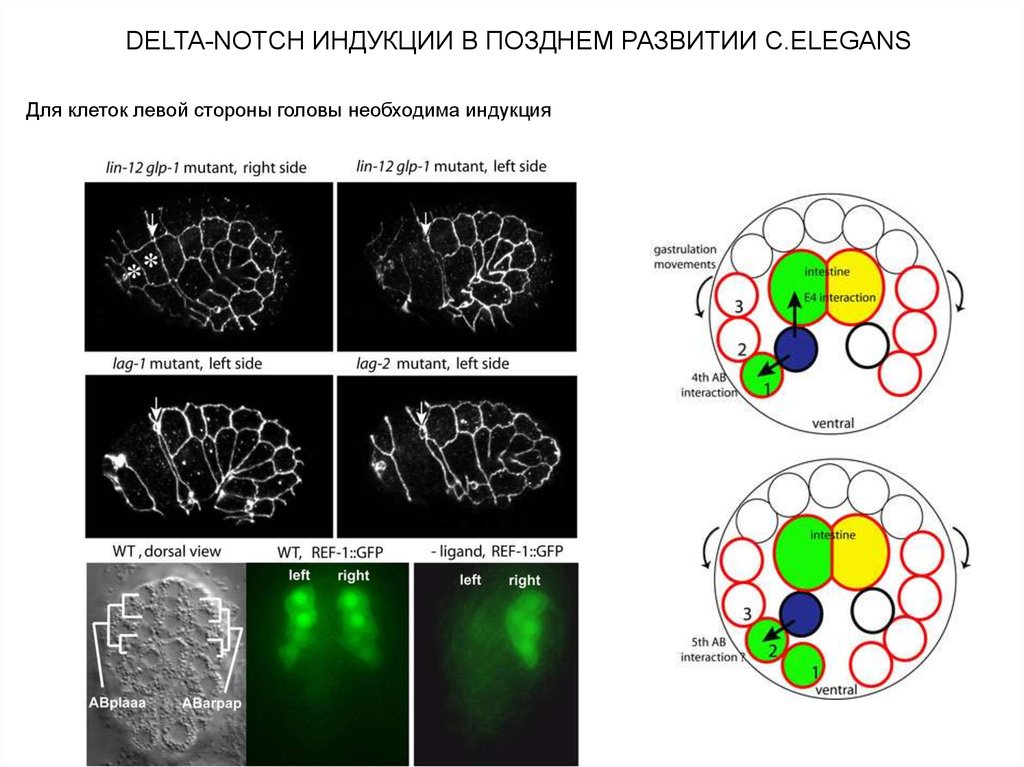
DELTA-NOTCH ИНДУКЦИИ В ПОЗДНЕМ РАЗВИТИИ C.ELEGANSДля клеток левой стороны головы необходима индукция
11.
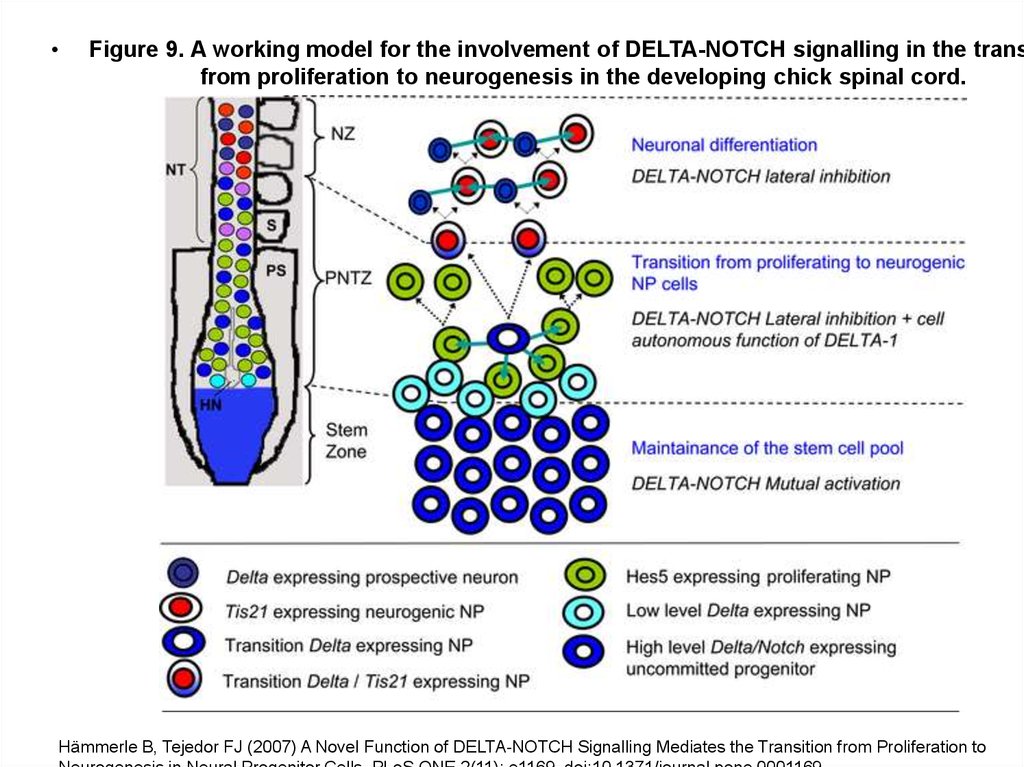
Figure 9. A working model for the involvement of DELTA-NOTCH signalling in the trans
from proliferation to neurogenesis in the developing chick spinal cord.
Hämmerle B, Tejedor FJ (2007) A Novel Function of DELTA-NOTCH Signalling Mediates the Transition from Proliferation to
12.
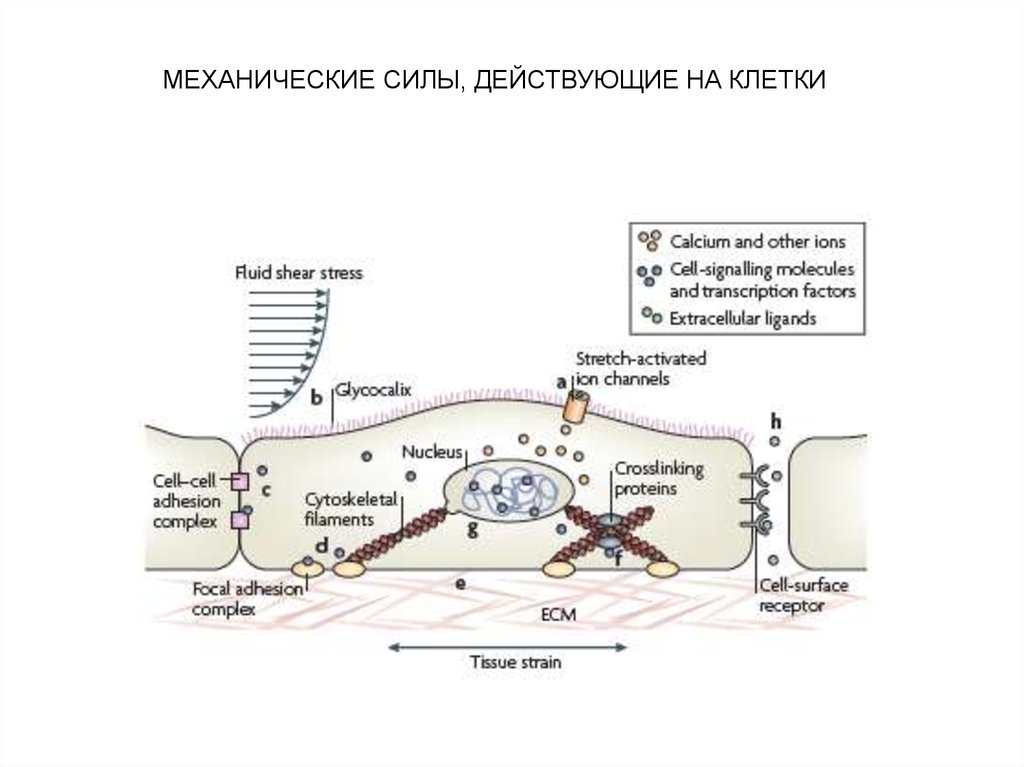
МЕХАНИЧЕСКИЕ СИЛЫ, ДЕЙСТВУЮЩИЕ НА КЛЕТКИ13.
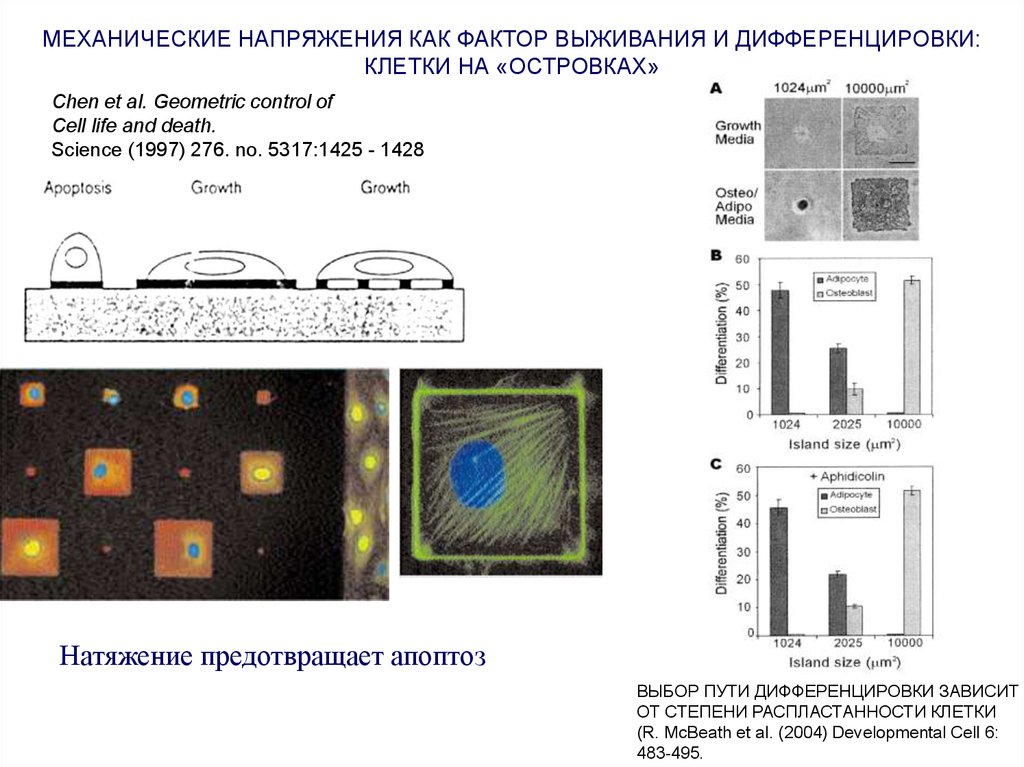
МЕХАНИЧЕСКИЕ НАПРЯЖЕНИЯ КАК ФАКТОР ВЫЖИВАНИЯ И ДИФФЕРЕНЦИРОВКИ:КЛЕТКИ НА «ОСТРОВКАХ»
Chen et al. Geometric control of
Cell life and death.
Science (1997) 276. no. 5317:1425 - 1428
Натяжение предотвращает апоптоз
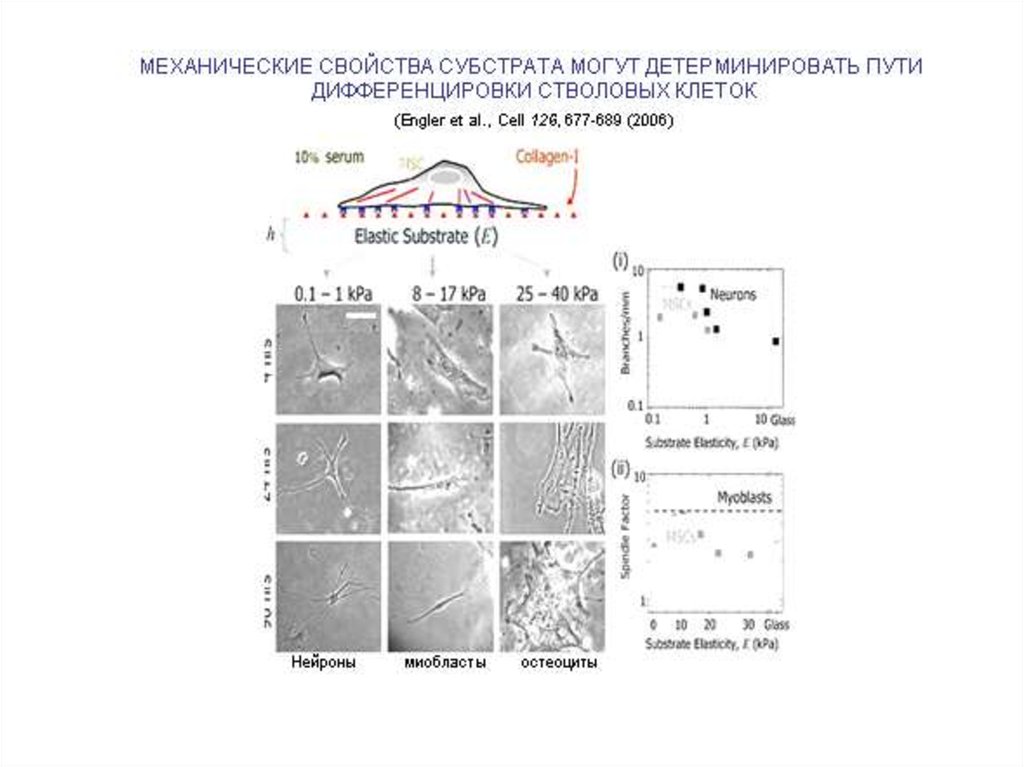
ВЫБОР ПУТИ ДИФФЕРЕНЦИРОВКИ ЗАВИСИТ
ОТ СТЕПЕНИ РАСПЛАСТАННОСТИ КЛЕТКИ
(R. McBeath et al. (2004) Developmental Cell 6:
483-495.
14.
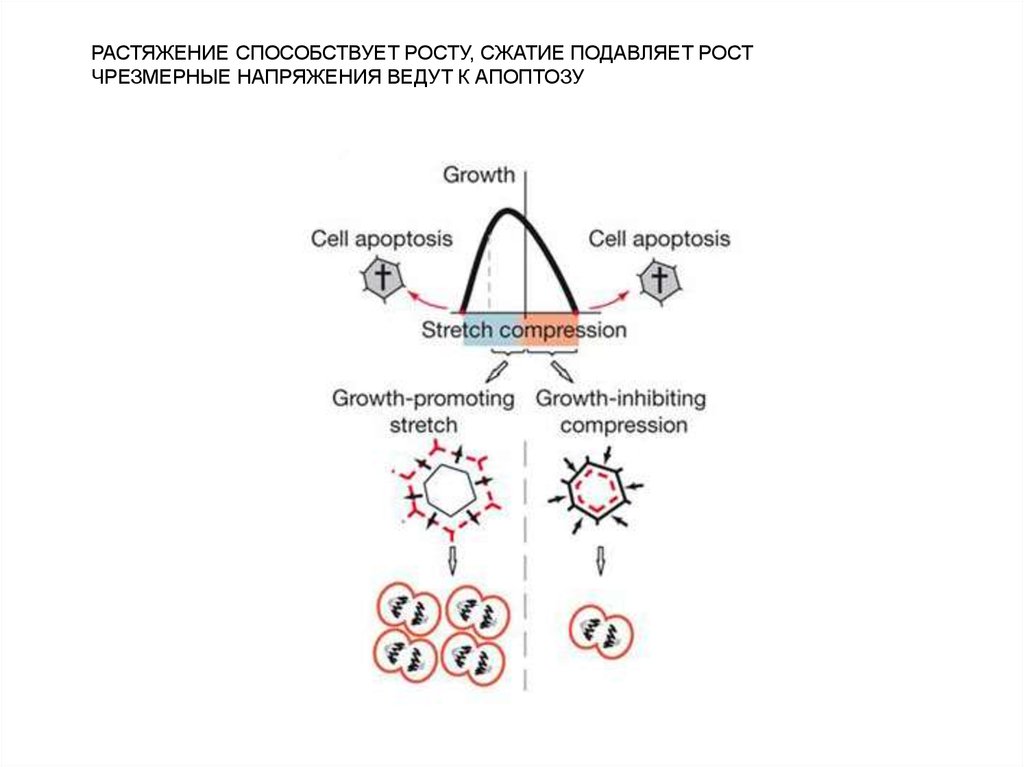
РАСТЯЖЕНИЕ СПОСОБСТВУЕТ РОСТУ, СЖАТИЕ ПОДАВЛЯЕТ РОСТЧРЕЗМЕРНЫЕ НАПРЯЖЕНИЯ ВЕДУТ К АПОПТОЗУ
15.
16.
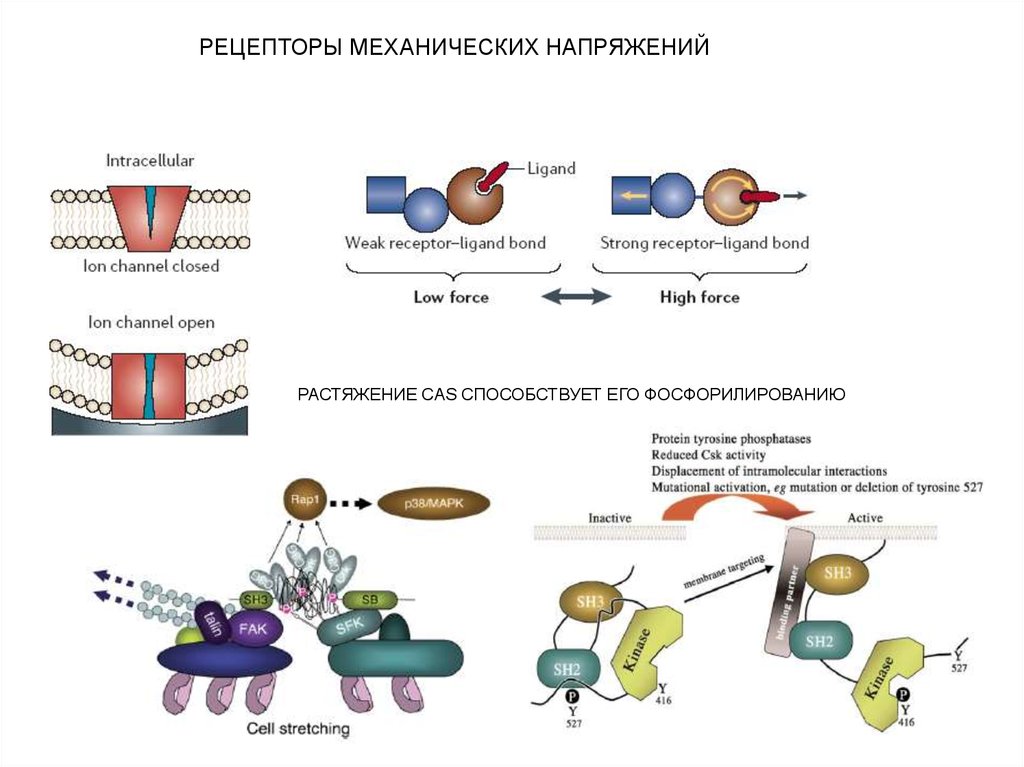
РЕЦЕПТОРЫ МЕХАНИЧЕСКИХ НАПРЯЖЕНИЙРАСТЯЖЕНИЕ CAS СПОСОБСТВУЕТ ЕГО ФОСФОРИЛИРОВАНИЮ
17.
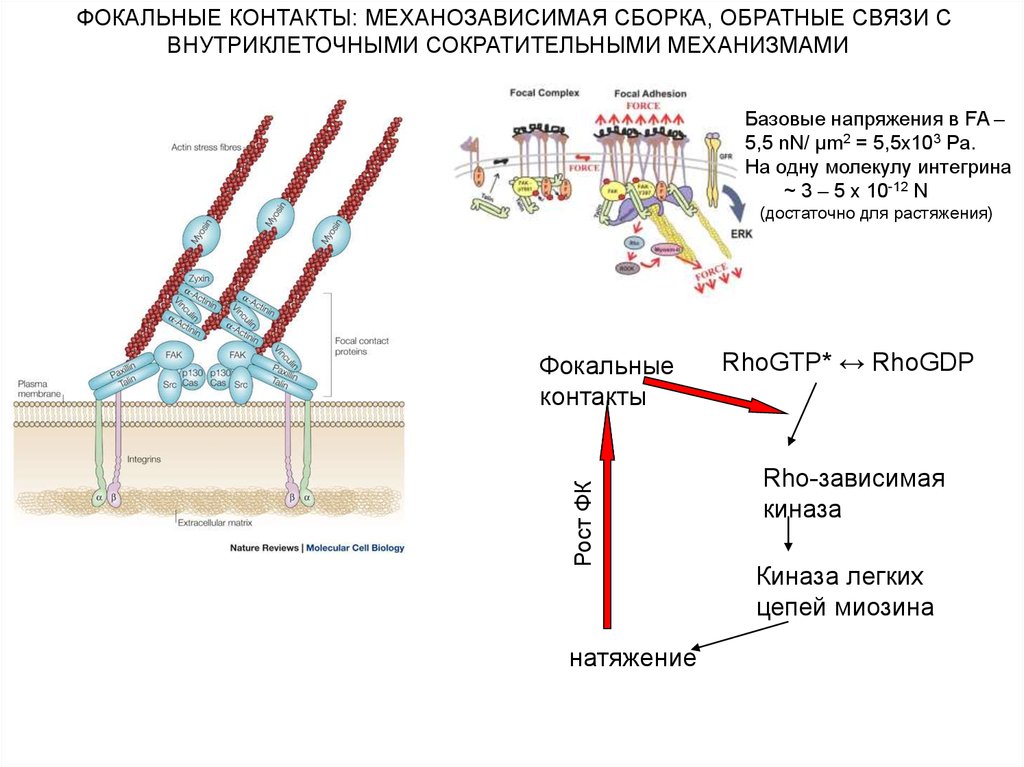
ФОКАЛЬНЫЕ КОНТАКТЫ: МЕХАНОЗАВИСИМАЯ СБОРКА, ОБРАТНЫЕ СВЯЗИ СВНУТРИКЛЕТОЧНЫМИ СОКРАТИТЕЛЬНЫМИ МЕХАНИЗМАМИ
Базовые напряжения в FA –
5,5 nN/ µm2 = 5,5x103 Pa.
На одну молекулу интегрина
~ 3 – 5 х 10-12 N
(достаточно для растяжения)
Рост ФК
Фокальные
контакты
натяжение
RhoGTP* ↔ RhoGDP
Rho-зависимая
киназа
Киназа легких
цепей миозина
18.
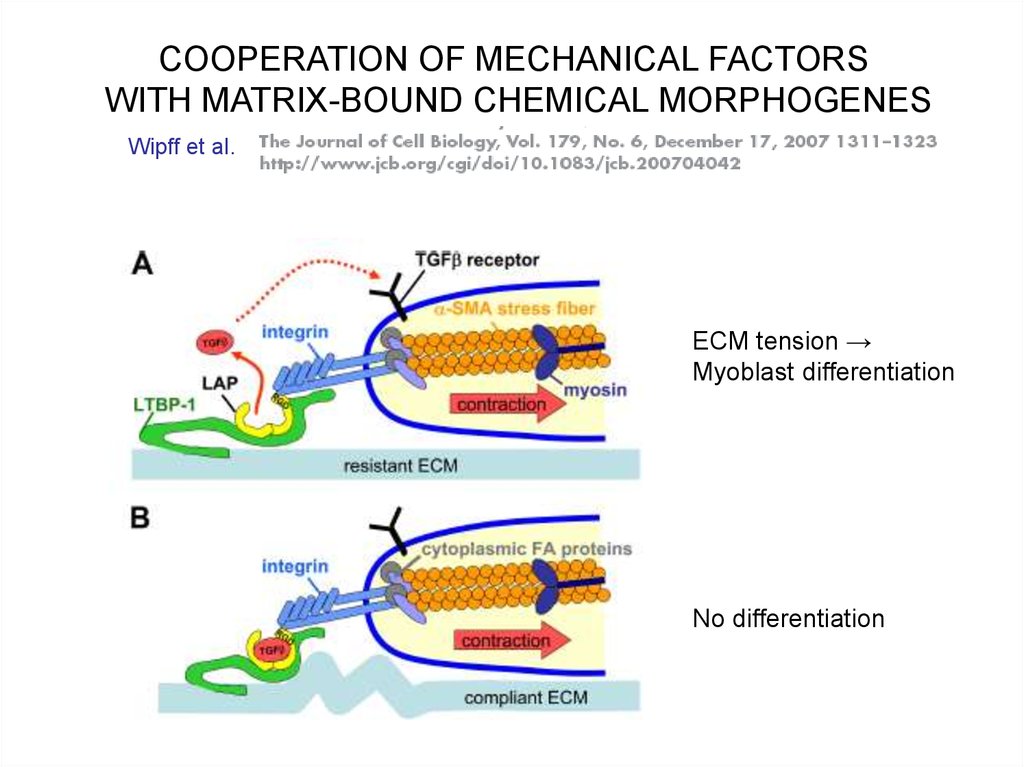
COOPERATION OF MECHANICAL FACTORSWITH MATRIX-BOUND CHEMICAL MORPHOGENES
Wipff et al.
ECM tension →
Myoblast differentiation
No differentiation
19.
ДИНАМИЧЕСКИЙ БАЛАНСМЕХАНИЧЕСКИХ СИЛ В
МЕЖКЛЕТОЧНЫХ КОНТАКТАХ
ТИПА ADHERENT JUNCTIONS
20.
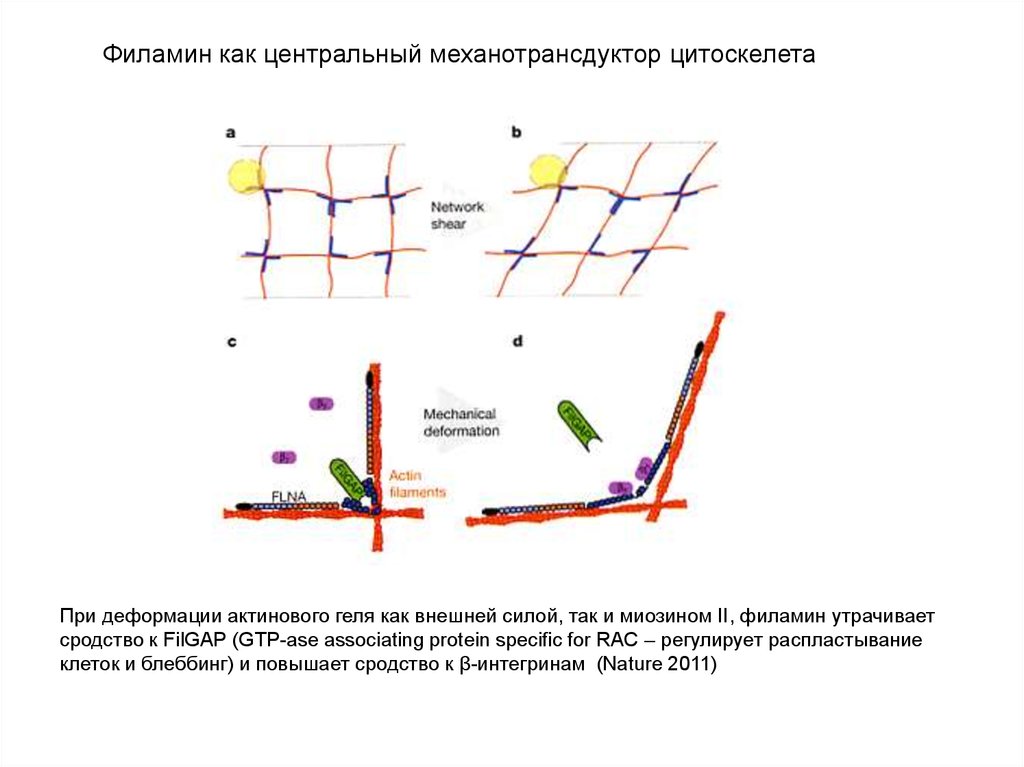
Филамин как центральный механотрансдуктор цитоскелетаПри деформации актинового геля как внешней силой, так и миозином II, филамин утрачивает
сродство к FilGAP (GTP-ase associating protein specific for RAC – регулирует распластывание
клеток и блеббинг) и повышает сродство к β-интегринам (Nature 2011)
21.
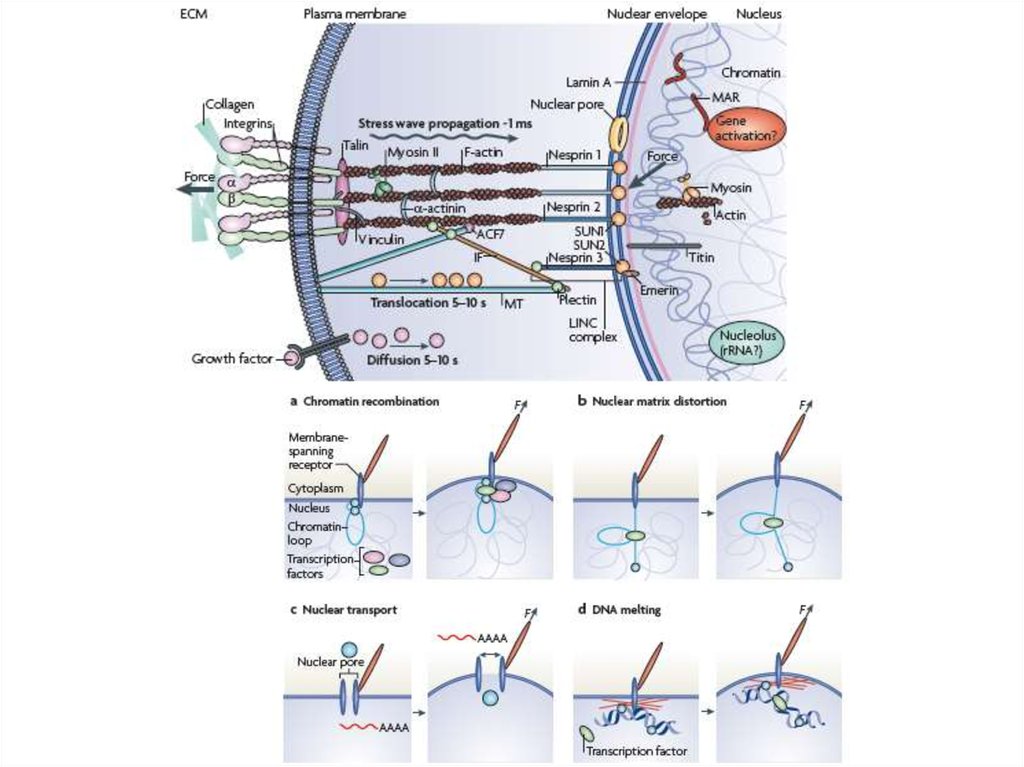
Механозависимые реснички22.
23.
24.
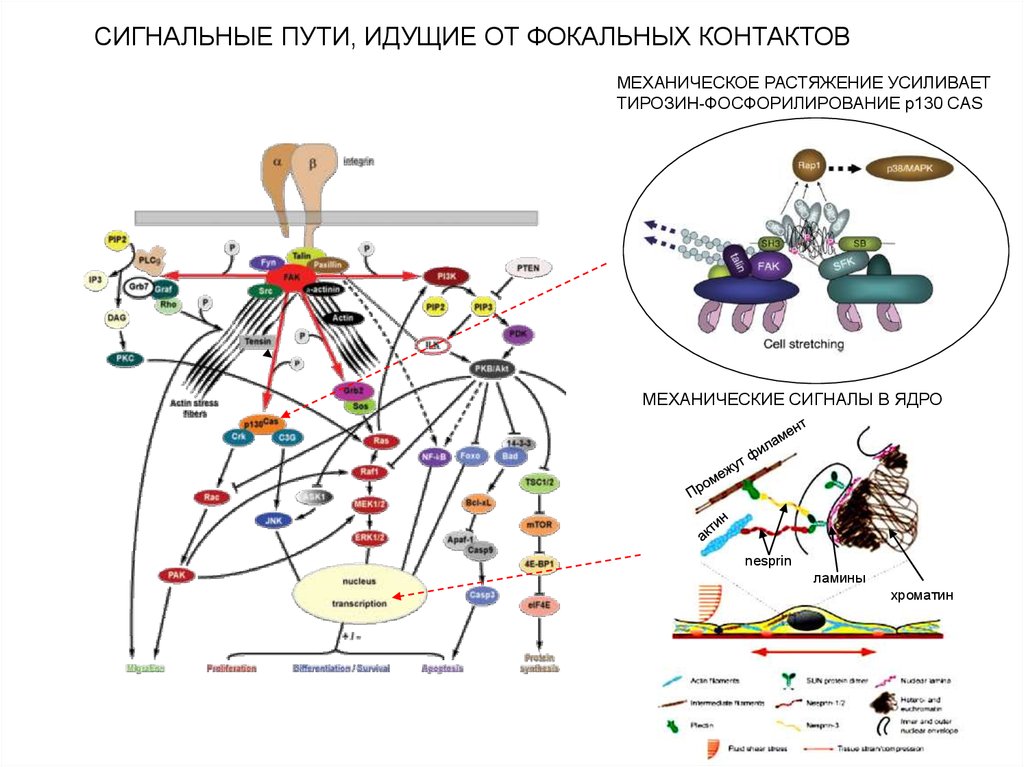
СИГНАЛЬНЫЕ ПУТИ, ИДУЩИЕ ОТ ФОКАЛЬНЫХ КОНТАКТОВМЕХАНИЧЕСКОЕ РАСТЯЖЕНИЕ УСИЛИВАЕТ
ТИРОЗИН-ФОСФОРИЛИРОВАНИЕ p130 CAS
МЕХАНИЧЕСКИЕ СИГНАЛЫ В ЯДРО
nesprin
ламины
хроматин
25.
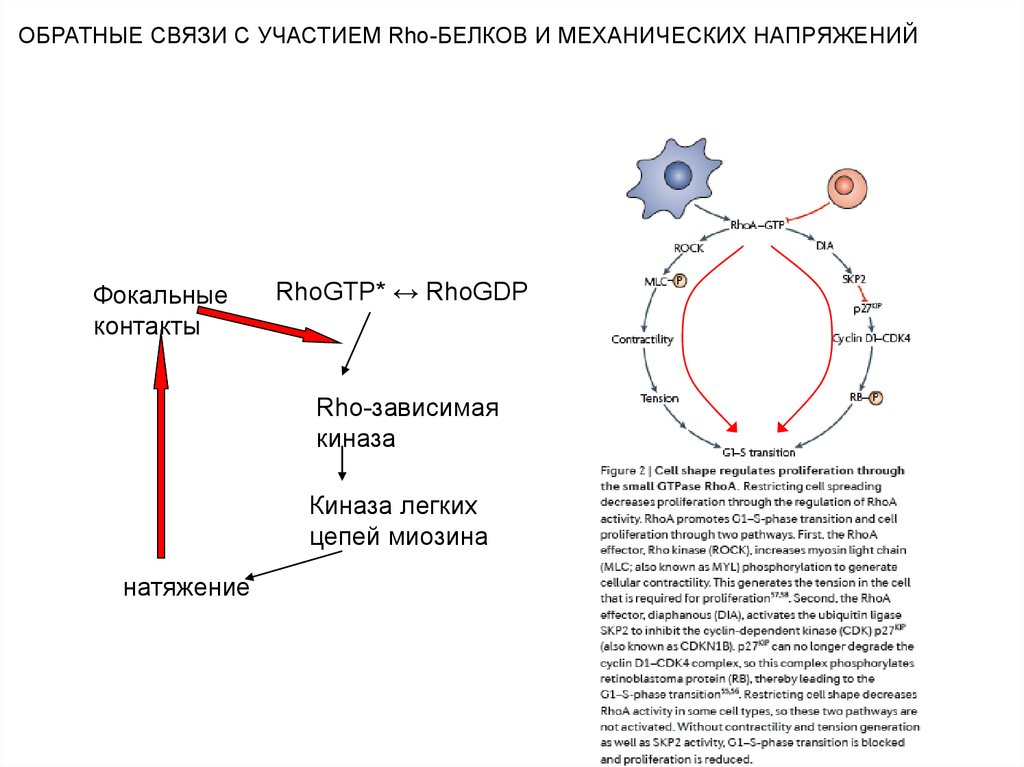
ОБРАТНЫЕ СВЯЗИ С УЧАСТИЕМ Rho-БЕЛКОВ И МЕХАНИЧЕСКИХ НАПРЯЖЕНИЙФокальные
контакты
RhoGTP* ↔ RhoGDP
Rho-зависимая
киназа
Киназа легких
цепей миозина
натяжение