



медиаторов")

")
")
")

 медиаторы")

:")
")
")
")
")





























 Биология
БиологияПохожие презентации:





")




Типы межклеточной сигнализации
1. Типы межклеточной сигнализации
Миллиарды нейронов и еще большее число глиальных клеток
нервной системы различным образом взаимодействуют между
собой, передавая друг другу сигналы различной природы и
обеспечивая тем самым сложно организованную регуляцию
мозговой деятельности.
В зависимости от механизма выделяют четыре типа
межклеточной сигнализации:
1.
2.
3.
4.
Неспецифическая сигнализация
Электрическая несинаптическая передача
Электрический синапс
Химический синапс
2. Типы межклеточной сигнализации
1.Неспецифическая
сигнализация
2.
Электрическая
несинаптическая
передача
3.
Электрический синапс
4.
Химический синапс
3. Медиаторы других организмов
4. Медиаторы других организмов
5. Общая характеристика (нейро)медиаторов
Медиаторы, как вещества, обеспечивающие передачу сигнала между нейронамидолжны удовлетворять ряду критериев:
1)
соединение (или его метаболические предшественники) должно содержаться в
соответствующем нейроне и в более высокой концентрации в нервном окончании;
2)
нейрон должен содержать ферменты для синтеза этого соединения, а в нервном
окончании должен действовать механизм его высвобождения;
3)
данное соединение должно высвобождаться при стимуляции нейрона; должно
быть доказано его присутствие во внеклеточном пространстве;
4)
должна существовать система инактивации этого соединения: фермент для его
деградации или механизм поглощения;
5)
должно быть показано наличие специфических рецепторов в субсинаптической
области;
6)
соединения, поступающие извне, должны оказывать такое же действие, как
стимуляция нерва.
6. Критерии медиаторов
Анатомический:присутствие вещества в необходимых количествах в
пресинаптических окончаниях нейрона.
Биохимический:
наличие в пресинаптическом нейроне
ферментативных систем, синтезирующих вещество, а
также систем, удаляющих или инактивирующих это
вещество в синаптической щели.
Физиологический:
выделение вещества из пресинаптического
нейрона при его адекватной стимуляции;
идентичность эффектов, вызываемых прямым
воздействием вещества на постсинаптическую
клетку и адекватной стимуляцией пресинаптического
нейрона.
Фармакологический:
фармакологические препараты, специфика действия
которых известна, вызывают ожидаемые эффекты
(изменение синтеза, накопления, высвобождения,
инактивации, обратного захвата вещества).
7. Классификация медиаторов (малые молекулы)
АминыАцетилхолин
Дофамин
Адреналин
Норадреналин
Октопамин
Гистамин
Серотонин
Acetylcholine, ACh
Dopamine, DA
Epinephrine
Norepinephrine, NE
Octopamine
Histamine
Serotinin, 5-HT
Аминокислоты
Глютамат
Glutamate, Glu
Аспартат
Aspartate, Asp
Гамма-амино масляная
кислота (ГАМК)
GABA
Глицин
Glycine, Gly
Таурин
Taurine
Пуриновые нуклеотиды
АТФ
Аденозин
ATP
Adenosine
катехоламины
моноамины
8. Классификация медиаторов (малые молекулы)
9. Классификация медиаторов (большие молекулы)
Известно более 100 нейропептидов (обычно из 3-36 аминокислот),выполняющих медиаторную функцию:
Пример: метионин-энкефалин
10. Медиаторы пептиды
Пептиды мозга и пищеварительного трактаОпиоидные пептиды
Пептиды нейрогипофиза
Гипоталамические релизинг-гормоны
Разные пептиды
Неполярные
Полярные незаряженные
Заряженные отрицательно
Заряженные положительно
11. Необычные (нетипичные) медиаторы
Эндоканнабиноиды:анандамид (anandamide)
2-арахидонил-глицерол (2-arachidonylglycerol, 2-AG)
Эндо-, потому что есть экзо∆9-тетрагидроканнабинол (∆9-tetrahydrocannabinol), психоактивный
компонент марихуаны (Cannabis sativa)
12. NO - вторичный посредник нежели необычный медиатор
NO (окись азота) выполняет роль в межклеточного коммуникатора и иногдасчитается необычным медиатором.
13. Принцип Г. Дейла (H. Dale):
«каждый нейрон выделяет из своих аксонных терминалей только одинмедиатор».
Почему «принцип Дейла» «не работает»:
- в одной везикуле вместе с медиатором может присутствовать несколько
нейропептидов;
- во многих нейронах присутствуют везикулы с «малыми» медиаторами и
секреторные гранулы (везикулы с нейропептидами).
В то же время «принцип Дейла» не противоречит тому, что
по крайней мере, не известно случаев, когда из одной аксонной терминали
нейрона выделял один медиатор, а из другой аксонной терминали - какойлибо другой медиатор.
Другими словами - из отдельного нейрона выделяется одно и то же.
14. Ацетилхолин (Acetylcholine, AСh)
Некоторые синапсы:- нервно-мышечные (в скелетной
мускулатуре);
- -мотонейроны на клетках Реншоу
в спинном мозге;
- преганглионарные нейроны
симпатического и
парасимпатического отделов на
постганглионарных нейронах в
вегетативной НС;
- постганглионарные
парасимпатические нейроны.
Инактивация:
деградация ферментом (эстеразой)
и обратный захват холина
15. Глютамат (L-Глутаминовая кислота, Glutamate, Glu)
Главный возбуждающий медиатор в мозге животных (более, чем в половине синапсовЦНС), а также в нервно-мышечных синапсах ракообразных и насекомых.
Некоторые синапсы:
- кора больших полушарий,
- обонятельная луковица,
- гиппокамп,
- мозжечок,
- сетчатка.
Не проходит через ГЭБ и
синтезируется из глютамина,
который переносится
транспортерами
из глиальных клеток.
Инактивация:
обратный захват транспортерами
16. -Аминомасляная кислота (ГАМК, GABA)
-Аминомасляная кислота (ГАМК, GABA)Главный тормозный медиатор в ЦНС (~ в трети синапсов).
Инактивация:
обратный захват транспортерами
17. Глицин (Glycine, Gly)
Тормозный медиатор примерно в половине тормозных синапсовспинного мозга (остальные содержат ГАМК)
Некоторые синапсы:
- клетки Реншоу спинного мозга;
- тормозные интернейроны
промежуточного мозга и
ретикулярной формации;
- сетчатка.
Инактивация:
обратный захват транспортерами
18. Биогенные амины
Регулируют множество функций в мозге ипериферической НС – от регуляции
гомеостаза до высших психических
функций.
Включают
три катехоламина - дофамин,
норадреналин
и адреналин,
-серотонин
- и гистамин.
19. Катехоламины
Имеют в своем составе общую группу катехола(выделено рамкой).
Синтезируются из тирозина через предшественник
ДОФА (DOPA), последовательно превращаясь друг в
друга в результате модификации радикала.
20. Катехоламины: дофамин
Главные источники:- нейроны полосатого тела (черной субстанции, substantia
nigra), включены в регуляцию произвольных
(целенаправленных) движений (экстрапирамидная система);
- нейроны вентральной покрышки, обеспечивают регуляцию
мотивационных состояний, механизмы вознаграждения и
подкрепления.
Некоторые синапсы:
- средний мозг: черная субстанция
(Substantia nigra),
- вентральная покрышка,
- обонятельная луковица,
- гипоталамус,
- перивентрикулярная область продолговатого
мозга,
- амакриновые клетки сетчатки.
Инактивация:
обратный захват транспортерами
Катаболизм:
деградация моноаминоксидазой (МАО)
и катехол-О-метилтрансферазой (COMT)
21. Инактивация катехоламинов
22. Катехоламины: норадреналин
Главные источники:- нейроны голубого пятна (Locus coeruleus) диффузно
иннервируют обширные области ЦНС (спинной мозг,
мозжечок, таламус и кору мозга), обеспечивают регуляцию сна
и бодрствования
- постганглионарные симпатические нейроны.
Некоторые синапсы:
- латеральная ретикулярная формация моста,
- продолговатый мозг,
- ядро одиночного тракта,
- постганглионарные симпатические нейроны.
Инактивация:
обратный захват транспортерами,
Катаболизм:
деградация в терминали МАО и COMT
23. Катехоламины: адреналин
Главные источники:в ЦНС нейроны
- латеральной покрышки
- и продолговатого мозга.
Некоторые синапсы:
- нейроны латеральной покрышки
- и продолговатого мозга (роль гормона-модулятора).
Инактивация:
обратный захват транспортерами
Катаболизм:
деградация в терминали МАО и COMT
24. Индоламин: серотонин
Синтезируется из триптофаначерез гидрокситриптофан
25. Индоламин: серотонин
Главные источники:- в ЦНС нейроны ядер шва
ростральной части моста.
Некоторые синапсы:
от нейронов ядер шва в
- лимбической системе,
- базальных ганглиях,
- коре больших полушарий.
Инактивация:
обратный захват транспортерами
Катаболизм:
деградация в терминали МАО
26. Имидазоламин: гистамин
Синтезируется из гистидина27. Имидазоламин: гистамин
Главные источники:- нейроны туберомамиллярных тел заднего гипоталамуса
Некоторые синапсы:
от нейронов гипоталамуса практически во всех отделах мозга
Инактивация:
обратный захват транспортерами
Катаболизм:
деградация в терминали МАО
и гистамин-метилтрансферазой
28. Синтез и инактивация гистамина
29. Пуриновые нуклеотиды: АТФ и аденозин
Все синаптические везикулы содержат АТФ, которая выделяется с одним илинесколькими «классическими» медиаторами.
Некоторые синапсы:
- вегетативные нервы на гладких мышцах
- широко распространены в ЦНС
и многих других тканях
Аденозин (на рис. без фосфатных групп, в рамке) не пакуется в везикулы, образуется
из АТФ с участием внеклеточных АТФаз и Са2+.
Инактивация:
обратный захват транспортерами
Катаболизм:
деградация в синаптической щели
апиразой (apyrase) и экто-5’-нуклеотидазой
30. Нейропептиды
Образуются из больших пре-пропептидов (син., пре-пробелков) в шероховатомЭПР.
Часто соседствуют в одних везикулах с «малыми» медиаторам.
Инактивация:
внеклеточные ферменты пептидазы разделяют их на аминокислотные
фрагменты.
31. Примеры нейропептидов
мозга и желудочно-кишечного трактаопиоидные пептиды
пептиды нейрогипофиза
Неполярные
Полярные незаряженные
Заряженные отрицательно
Заряженные положительно
выделяемые в гипоталамусе пептиды
разные пептиды
32. Семейства нейроактивных пептидов «химическая» классификация
33. Семейства нейроактивных пептидов «функциональная» классификация
всего 4534. Эндоканнабиноиды
Анандамид (anandamide)Синтезируется из мембранного
липида фосфатидил-этанол-амина в
две стадии в результате повышения
внутриклеточной концентрации Са2+.
Механизм высвобождения из клеток
неизвестен.
Предполагается, что он
диффундирует через клеточную
мембрану и достигает соседних
клеток.
Через свои рецепторы уменьшает
выделение ГАМК из тормозных
терминалей, предположительно
действуя на потенциал-зависимые
Са2+- и/или К+-каналы
пресинаптической мембраны.
Инактивация:
гидролизуется гидролазой жирных
кислот
35. Эндоканнабиноиды
2-арахидонил-глицерол (2-arachidonylglycerol, 2-AG)36. Механизм действия эндоканнабиноидов
37. NO - вторичный посредник нежели необычный медиатор
NO синтезируется NO-синтазой, которая в нейронах регулируетсяСа2+/калмодулином.
NO напрямую активирует ассоциированную с мембраной гуанилатциклазу.
В отличие от типичных медиаторов NO (газ)
- не накапливается в везикулах;
- не имеет специфических мембранных рецепторов;
- не имеет активного механизма инактивации и быстро окисляется до NO2.
38. Нейромодуляторы
В отличие от нейромедиаторов имеют следующие характеристики:- не обладают самостоятельным физиологическим действием, а
модифицируют эффект нейромедиаторов;
- действие имеет тонический характер – медленное развитие и большая
продолжительность (через метаботропные рецепторы);
- в месте воздействия не имеют специфического механизма инактивации
(деградации, обратного поглощения в пресинаптическую терминаль)
- не обязательно синаптическое (и даже нейронное) происхождение – могут
выделяются железами или синтезироваться во внутриклеточных каскадах, а
также поступают из внеклеточного пространства в результате растекания
(spill over);
- действие не сопряжено с эффектом нейромедиатора (самостоятельный
эффект);
- действуют на постсинаптические и пресинаптические мембранные
рецепторы, а также на различные участки нейрона, в том числе и
внутриклеточно.
39. Пресинаптическая модуляция: ауторецепторы
Высвобождаемый медиатор воздействует на собственные пресинаптическиерецепторы, уменьшая или увеличивая свое последующее высвобождение.
Пример:
НА воздействует на пресинаптические 2–рецепторы и тормозит
последующую секрецию НА.
По фармакологическим
характеристикам пресинаптические
ауторецепторы отличаются от
постсинаптических того же
медиатора.
40.
Пресинаптическая модуляцияCharacteristics of a norepinephrine (NE)-containing catecholamine neuron. Tyrosine (Tyr) is accumulated by
the neuron and is then metabolized sequentially by tyrosine hydroxylase (TH) and L-aromatic amino acid decarboxylase (L-AADC) to dopamine (DA). The DA is then taken up through the vesicular monoamine transporter into vesicles. In DA neurons, this is the final step. However, in this NE-containing cell, DA is metabolized to NE by dopamine-b-hydroxylase (DBH), which is found in the vesicle. Once NE is released, it can interact with postsynaptic noradrenergic receptors or presynaptic noradrenergic autoreceptors. The accumulation of NE by the high-affinity membrane NE transporter (NET) terminates the actions of NE. Once taken
back up by the neuron, NE can be metabolized to inactive compounds (DHPG) by degradative enzymes such
as monoamine oxidase (MAO) or taken back up by the vesicle.
41.
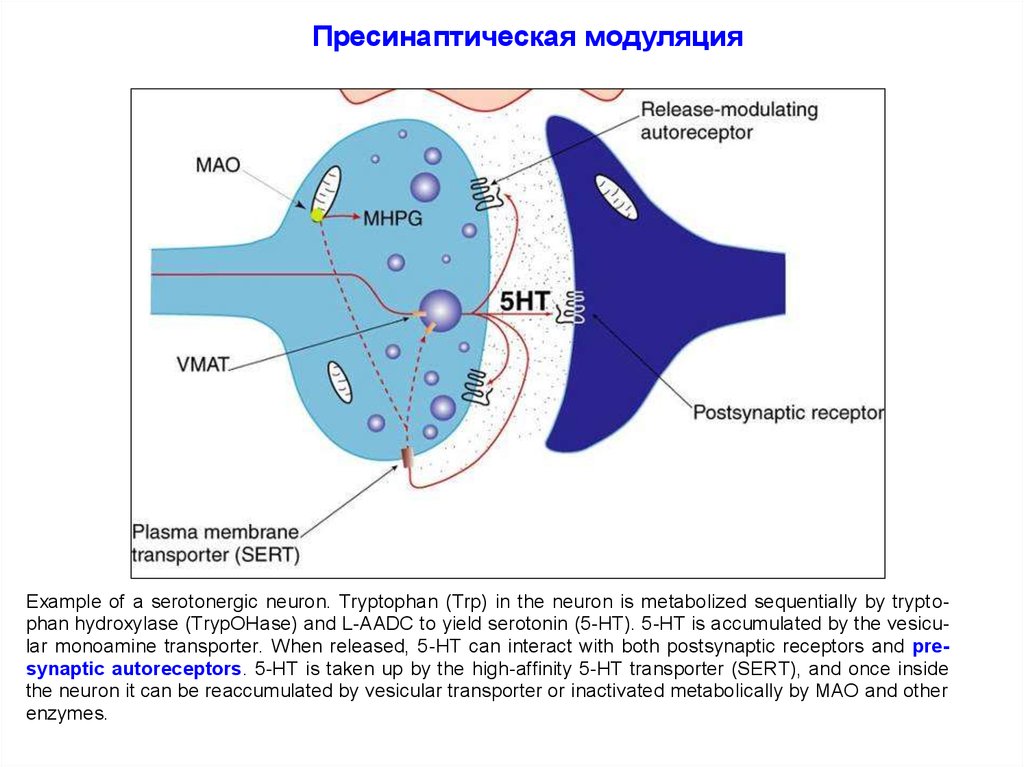
Пресинаптическая модуляцияExample of a serotonergic neuron. Tryptophan (Trp) in the neuron is metabolized sequentially by tryptophan hydroxylase (TrypOHase) and L-AADC to yield serotonin (5-HT). 5-HT is accumulated by the vesicular monoamine transporter. When released, 5-HT can interact with both postsynaptic receptors and presynaptic autoreceptors. 5-HT is taken up by the high-affinity 5-HT transporter (SERT), and once inside
the neuron it can be reaccumulated by vesicular transporter or inactivated metabolically by MAO and other
enzymes.
42.
Пресинаптическая модуляцияDepiction of an excitatory amino acid (glutamate) synapse. Glutamate, synthesized via metabolic pathways, is concentrated through a vesicular transporter into secretory granules. After release from the presynaptic terminal, glutamate can interact with postsynaptic and/or release-modulating receptors. Glutamate is then cleared from the synaptic region by high-affinity plasma membrane transporters or by recycling through adjacent glia.
43.
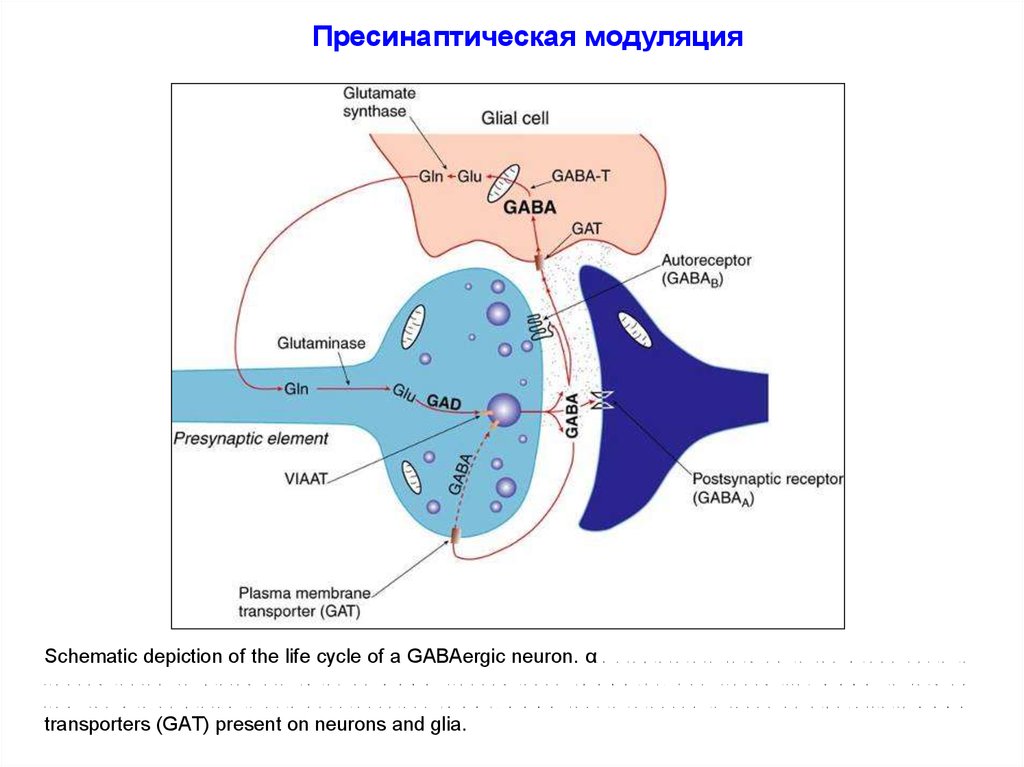
Пресинаптическая модуляцияSchematic depiction of the life cycle of a GABAergic neuron. α
t
r
a
n
f
r
o
m
s
a
m
t
h
i
e
n
a
G
t
l
u
e
d
b
t
y
o
g
g
l
u
l
t
u
a
t
m
a
m
i
a
c
t
a
e
c
(
i
d
G
d
l
e
u
)
c
b
a
r
y
b
G
o
x
A
y
l
B
a
A
s
e
t
(
G
r
a
n
A
transporters (GAT) present on neurons and glia.
s
D
a
)
m
.
i
G
n
A
a
s
B
e
A
(
t
h
-
K
G
e
A
a
t
B
i
s
t
o
g
A
-
r
e
l
T
l
u
t
)
e
a
.
a
r
T
s
a
h
e
t
e
f
e
d
t
i
s
o
r
r
a
t
m
n
a
e
s
k
m
e
d
i
i
n
t
b
n
t
e
y
t
h
r
e
K
G
h
i
A
g
h
r
B
-
e
b
A
a
f
s
i
f
i
n
c
s
i
f
t
y
y
c
o
r
G
l
e
i
m
A
s
e
B
d
A
44. Пресинаптическая модуляция: гетерорецепторы
Модулируют эффект медиаторов, высвобождающихся из других нейронов.или модуляторов несинаптического происхождения.
Пример:
Пресинаптические мАцХР на НАокончаниях симпатических нервов
взаимодействуют с АцХ,
секретируемым из парасимпатических холинергических аксонов.
Модуляция состоит в изменениях:
- возбудимости нервных окончаний,
- биосинтеза медиаторов,
- входа Са2+,
- стадий экзоцитоза.
мАцХР
- распространение АцХ из терминалей
постганглионарных парасимпатических нейронов
45. Пресинаптическая модуляция: гетерорецепторы
Пресинаптические СВ1 снижают высвобождение глютамата, АцХ и ГАМКчерез активацию G / -димера, который блокирует Са2+-каналы и активирует
K+-каналы.
Пресинаптические СВ1 снижают
высвобождение глютамата, АцХ и
ГАМК через активацию G / -димера,
который блокирует Са2+-каналы и
активирует K+-каналы.
Синтез эндоканнабиноидов
запускается при увеличении
внутриклеточного Са2+ и (или)
активации липидных каскадов.
Эндоканнабиноиды транспортируются
(механизм неизвестен) из
постсинаптической клетки и
связываются с пресинаптическими
рецепторами – т.н.
эндоканнабиноидная ретроградная
регуляция выделения медиатора.
46. Постсинаптическая модуляция: ауторегуляция
Отрицательная ауторегуляцияДесенситизация (десенсибилизация) рецепторов при длительном воздействии
медиатора.
Пример:
Если постсинаптическая мембрана подвергается действию увеличенных
концентраций АцХ, наблюдается медленное снижение постсинаптического
ответа. При этом не меняется сродство рецептора к АцХ, снижение ответа
происходит из-за того, что ионные каналы не открываются.
Механизмы десенситизации:
- через фосфорилирование
- физическое удаление рецептора из мембраны (интернализация и
деградация).
47. Постсинаптическая модуляция: ауторегуляция
Положительная ауторегуляцияГиперсенситизация (гиперсенсибилизация) при недостаточности воздействия
медиатора.
Пример:
Если денервировать мышцу, то вся мышечная поверхность в течение
нескольких дней приобретает способность активироваться АцХ.
Плотность АцХР за пределами синаптической области значительно
увеличивается.
Это обусловлено не перемещением субсинаптических АцХР, а появлением
новых внесинаптических АцХР. Кинетические свойства новых
внесинаптических каналов отличаются от свойств субсинаптических каналов:
время открытия новых каналов в 3 раза больше, а проводимость составляет
только 1/3 от проводимости нормальных каналов.
При этом сродство к лигандам и имуннологические свойства одинаковы.
Гиперсенсибилизация может быть устранена путем реинервации: когда
восстанавливается нормальная инервация мышцы - внесинаптические АцХР
исчезают. Это наблюдается также и при длительной электростимуляции
мышечного волокна.
48. Постсинаптическая модуляция: гетерорегуляция
постсинаптическое межрецепторное взаимодействие между сопутствующимимедиаторами.
Сопутствующие (сосуществующие) медиаторы характеризуются
- совместной локализацией,
- совместным высвобождением
- и общей клеткой-мишенью.
Совместная локализация - синтез и депонирование в одних и тех же
пресинаптических окончаниях (возможно, в разных синаптических пузырьках).
Совместное высвобождение – экзоцитоз двух (и более) медиаторов в
результате одного процесса активации пресинаптического окончания.
Примеры сочетания медиаторов:
1) Несколько «классических» медиаторов (АцХ и серотонин).
2) «Классический(ие)» медиатор(ы) и нейропептид(ы) (нейропептид Y и НА).
3) Несколько нейропептидов.
4) Пурины и «классические» медиаторы (НА и АТФ).