


")














































































 Биология
БиологияПохожие презентации:










Репродукция и дифференцировка клеток
1. Министерство образования и науки Российской Федерации Дальневосточный федеральный университет Кафедра клеточной биологии и
генетикиА.П. Анисимов
РЕПРОДУКЦИЯ И
ДИФФЕРЕНЦИРОВКА КЛЕТОК
Часть I. Базовый уровень
Конспект-презентация лекционного курса
Владивосток
2017
2.
3.
Настоящее учебное пособие подготовлено в форме конспекта-презентациилекционного спецкурса, читаемого автором на протяжении многих лет для студентовбиологов, специализирующихся по профилю «Клеточная биология». Не считая
исторического введения, в нем есть три большие темы. Первая тема посвящена
структуре и общим процессам митотического (клеточного) цикла; вторая – динамике
пролиферативных процессов в онтогенезе, от первых делений зиготы до зрелых
тканей с особенностями их кинетики и дифференцировки; в третьей теме
рассматриваются молекулярно-генетические механизмы регуляции клеточного
размножения и дифференцировки, представляющие сложную иерархию
взаимодействий различных факторов – от средовых до генетических.
К настоящему времени по этим проблемам накоплен и продолжает расти
огромный научный материал. Отразить его с достаточной полнотой в одном учебном
(семестровом) пособии не представляется возможным, поэтому предлагаем
настоящий спецкурс «Репродукция и дифференцировка клеток» лишь как «Часть I –
Базовый уровень». Это означает, что без этих начальных знаний невозможно
осваивать самые актуальные материалы по структуре и генетической мобильности
хроматина, биологии и технологии стволовых клеток, регенерации тканей и органов,
злокачественной трансформации клеток и другим проблемам, которые, как хотелось
бы автору, должны быть отражены в Части II под условным названием «Актуальные
проблемы». К сотрудничеству в этой сложной работе, а также к полезной критике
данного пособия приглашаю специалистов.
В нашем пособии минимум текста, но достаточно иллюстраций, что обычно
любят студенты, поскольку позволяет удовлетворительно усвоить на базовом уровне
учебный материал. Учебное пособие могут использовать, кроме биологических
цитологов, гистологов и эмбриологов, генетики, физиологи, биохимики, а также
студенты биомедицинского профиля (лечебники, биофизики, биохимики) и другие
специалисты, которым потребны современные знания в области клеточной биологии.
4. ВВЕДЕНИЕ 1. Проблема репродукции и дифференциации клеток в клеточной теории (История проблемы, до 1953 г.)
17 в. 1665 г. – Роберт Гук = открытие клеток у растений.
1671 г. – Грю, 1675 г. – Мальпиги = у животных.
До 19 в. Новых принципиальных положений не высказано.
Начало 19 в. = формируется клеточная теория:
1802 – Бриссо-Мирбе, 1809 – Ламарк, 1824 – Дютроше
= все организмы состоят из клеток.
1830 – Ян Пуркинье = описание клеточного ядра (зародышевый пузырек
яйца курицы).
1831 – итальянец Роберт Броун = обнаружил ядро в растительной клетке.
Ввел термин nucleus. Идея о постоянном наличии ядра во всех клетках.
Роль ядра – неизвестна. (=Комочек слизи, питательные вещества ...).
1838-39 – Теодор Шванн (немецкий зоолог) = окончательная и наиболее
полная формулировка клеточной теории.
5.
Одним из постулатов в клеточной теории Т.Шванна принята идея М. Шлейдена об
эндогенном цитогенезисе в форме
«свободного клеткообразования» из
недифференцированной бластемы:
= В слизистой массе зародыша (бластеме)
образуются зернышки, вокруг которых
конденсируется слизь и появляется оболочка –
формируется цитобласт (=ядро).
Конденсация продолжается, образуется вторая
оболочка – получается молодая клетка (=цит).
По Шванну, у животных
цитобластема находится
в межклеточном веществе.
6.
• В то же время, уже в 1824 г. наблюдали деление клеток (делениебластомеров раннего зародыша).
1832 – деление перегородками у нитчатых водорослей.
• Но эти факты не принимались как универсальный способ
клеткообразования. Поведение и роль ядра не понимали.
• 1847 – Горянинов (акад. Петербургской АН) допускал Шлейденовский
цитогенезис для возникновения первых в эволюции клеток. Но в
индивидуальном развитии признавал деление (перегородками,
почкованием).
• 1841-1852 – Ремак. Его первые наблюдения –
• деление клеток крови цыпленка.
Дал описание и обзор фактов по делению клеток и ядер перегородками
и почкованием (позже Флемминг назвал такое деление амитозом).
Утверждал, что это универсальное явление.
= Большой толчок к исследованию деления клеток.
• 1855-1859 – Рудольф Вирхов. «Целлюлярная патология».
Полностью развенчал теорию цитогенезиса Шлейдена, которая
господствовала благодаря авторитету Шванна. Вирхов (1858): все клетки
образуются путем деления! (Omnis cellula e cellulae!).
7.
• Однако Вирхов митоза не знал. Его видели,видели перестройки ядра, но полной картины и
объяснения не было. Накапливались отдельные
сведения о поведении ядра, рисовали мета- и
анафазы, не понимая сути.
• 1867 – Гофмейстер = наблюдал митоз у растений.
1873 – Шнейдер (зоолог) = первое описание последовательных картин митоза
на дробящихся яйцах плоского червя. Это наблюдение осталось неизвестным
для биологов.
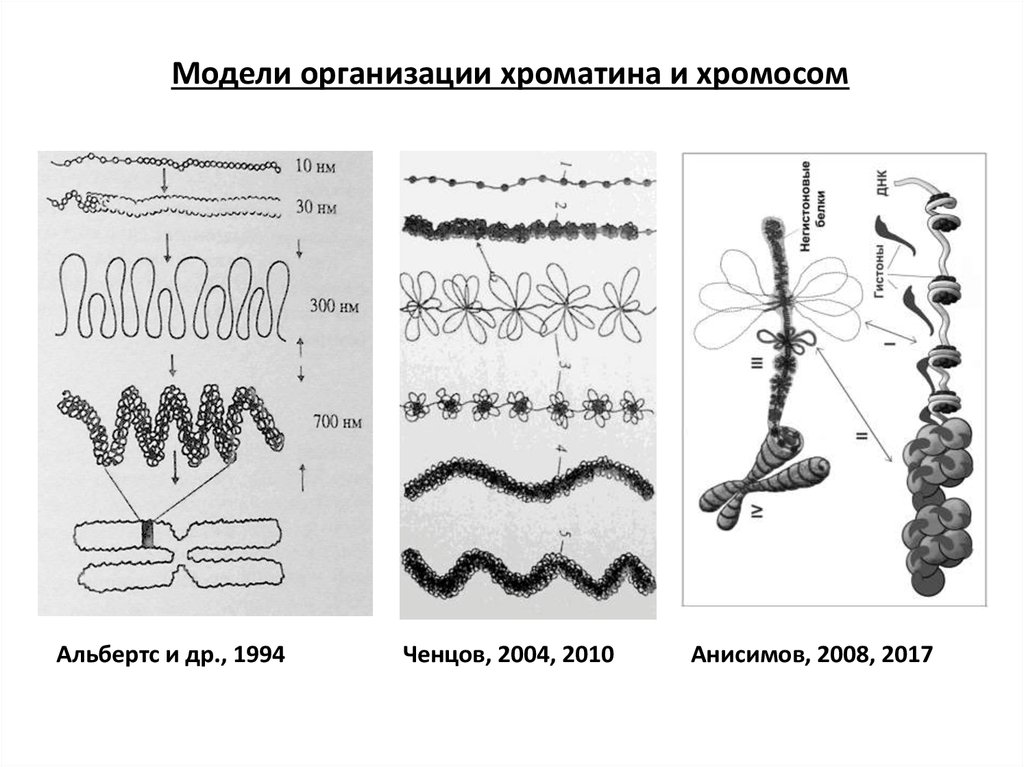
1874 – Бючли = редукционное деление у нематод и улиток. Но по его
заключению, ядро выталкивается из клетки, а новообразование ядер и клеток
происходит из вакуолек и зернышек цитоплазмы, все от Шлейдена).
1876 – Ван Бенеден = открыл клеточный центр.
8.
1879 – Вальтер Флемминг = первое верное описаниепоследовательных фаз митоза
Клетки кожи саламандры, крупные хромосомы. Термины
Флеминга: «митоз», «амитоз» (то, что рисовал Ремак),
«хроматин» и др.
По Флеммингу:
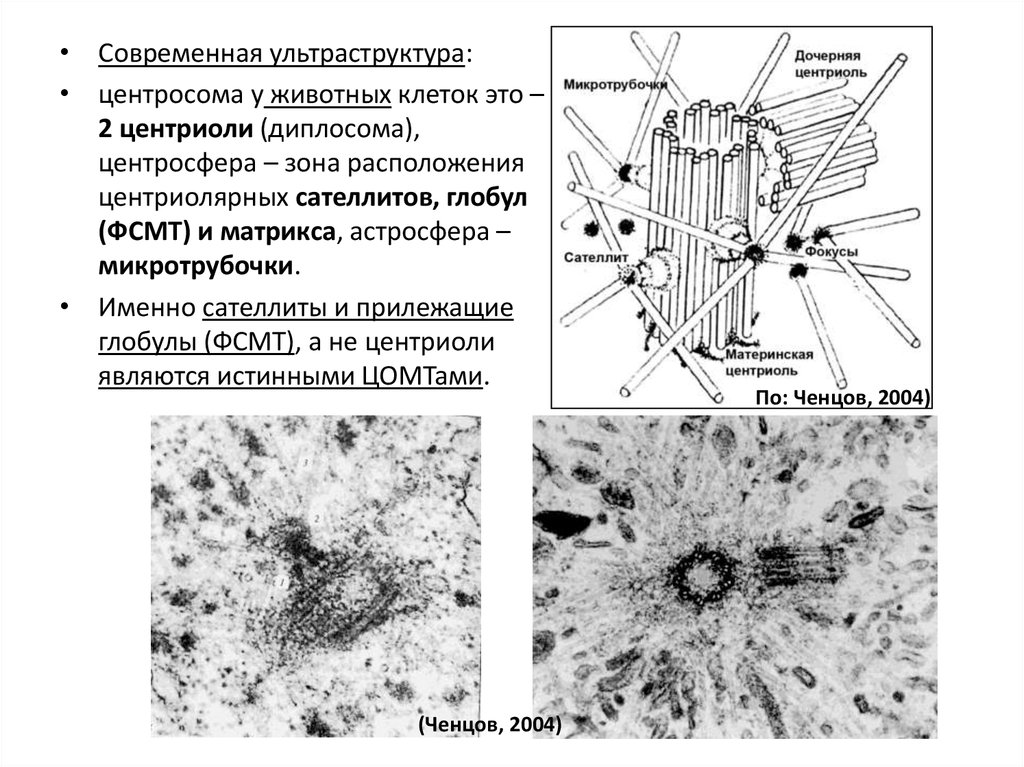
хроматин
превращается в
нити, идет
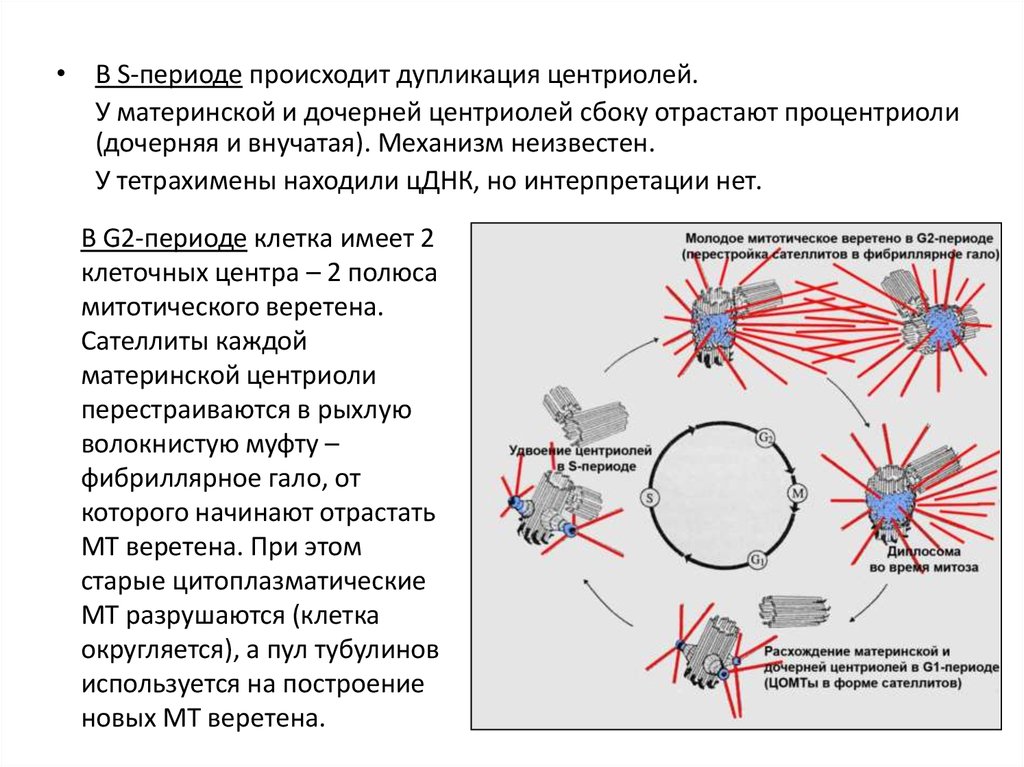
продольное
расщепление
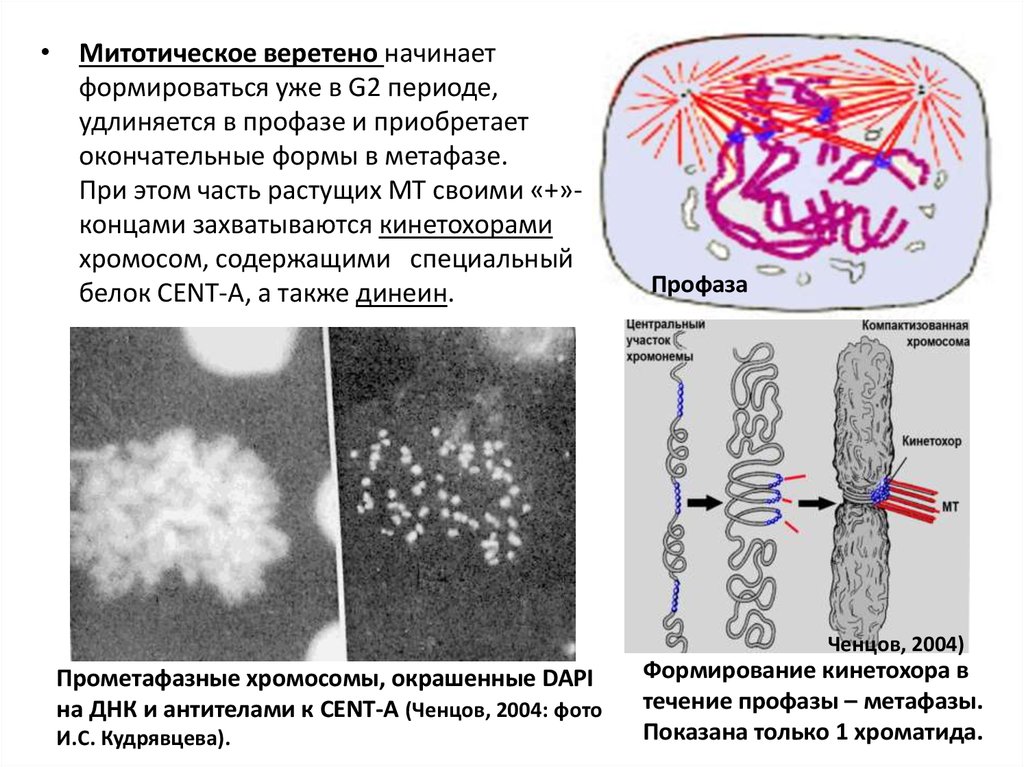
нитей и
расхождение к
полюсам. (термин
«хромосома»
позднее дал
Вальдейер, 1883).
9.
Лавина работ. «Ядро затмило цитоплазму».
Особо отметим цикл работ Ван Бенедена – дробление у аскариды.
Но объяснения смысла продольного расщепления хромосом не было.
1883 – Вильгельм Ру, крупнейший биолог-теоретик: зернышки ядра
соединяются в нити, они (зернышки) разнокачественные. Только
продольное расщепление нитей обеспечивает равноценное разделение
содержимого ядра по двум клеткам.
Это величайшая догадка и, по сути, теория митоза.
1884-85 – О. Гертвиг, Страсбургер, Рабль. Сформулирована гипотеза: ядро –
носитель наследственности.
Открыт закон постоянства числа хромосом в соматических клетках одного
биологического вида.
Эта идея наиболее развита Августом Вейсманом в его учении о
зародышевой и соматической плазме.
Дальнейшее развитие этой концепции – вся теория гена Томаса
Моргана. Оформление цитогенетики с хромосомной теорией
наследственности (Сеттон, нач. 20 в.).
1896 – Э. Вильсон (американский цитолог). Первая капитальная сводка
по цитологии и цитогенетике: «Клетка и ее роль в развитии и
наследственности» (русский перевод 1936, 1940).
10.
• Параллельно с морфологическими наблюдениями развиваются ибиохимические.
1868 – Мишер = обнаружил новый класс органических соединений –
нуклеопротеиды (из ядер лимфоцитов, гноя).
1872 – в ядрах спермы лосося обнаружены протамины и НК.
1889 – получены чистые НК, свободные от белка.
• Однако в начале 20 в. наследственный материал связывали с белками.
Н. Кольцов = идея самоудвоения белков хромосом.
• Лишь в 40-х годах выявлены различия ДНК (ТНК) и РНК и установлена их
локализация (Фельген, Розенбек, Белозерский, Девидсон).
• Возникло предположение о связи ДНК с хромосомами и ее роли в хранении
и передаче наследственной информации.
• 1953 – Уотсон и Крик = расшифровка структуры ДНК и механизма
репликации. Позже Крик = расшифровка генетического кода (ЦДМБ).
• 1953 – Говард и Пелк = сформулированы понятие митотического цикла.
Сошлись биохимические и цитологические работы. Начало современного
этапа в изучении механизмов репродукции и дифференцировки клеток.
• 1953 – умер Сталин = начало освобождение генетики и цитологии в СССР от
засилия лысенковщины.
11. 2. Актуальность новых исследований
• Кто и для чего изучает репродукцию и дифференцировку клеток?Проблемы репродукции и дифференцировки клеток имеют
междисциплинарное значение. Трудно найти другое направление, в
котором так тесно переплетались бы интересы разных биологических
и медицинских наук. При этом вопросы репродукции (размножения) и
дифференцировки (специализации) клеток тесно взаимосвязаны
между собой. Дифференцировка начинается «из-под» репродукции, в
ходе митотических циклов. Рассматривая дифференцировку, исходим
из клеточного цикла; изучая циклы, неизбежно выходим на
дифференцировку.
• Чьи же это проблемы?
• Цитология (в узком смысле). Морфология митоза, цикл хромосом и
др. клеточных структур, связь митоза и дифференцировки с
клеточным метаболизмом.
• Биология развития. Гаметогенез, в т.ч. специфический цикл мейоза.
Репродукция и дифференцировка клеток как механизм
эмбрионального морфогенеза (дробление, гаструляция, органогенез).
• Гистология. Нормальный и репаративный гистогенез.
Физиологическая регенерация (обновление) тканей и органов.
12.
• Частные гистофизиологические дисциплины. Особенно гематология ииммунология. Без знания динамики и механизмов регуляции клеточной
репродукции и дифференцировки такие дисциплины не состоятельны.
• Онкология и радиобиология. Малигнизация (опухолевая трансформация)
клеток. Именно задачи онкологии и радиобиологической защиты стали
главным стимулом развития этих проблем в 50-60-е годы прошлого века и
сохраняются по сей день. Задача: понять механизмы контроля, регуляции
клеточного цикла, дифференцировки и смерти (апоптоза) клеток, чтобы
ими управлять в патологических ситуациях.
• Молекулярная биология и генетика. Регуляторы клеточного цикла и
дифференцировки – молекулы, главным образом белки, кодируемые
соответствующими генами (гены клеточного цикла и дифференцировки).
Белковые регуляторы специфичны к тканевым клеткам-мишеням. Тут
задействованы разнообразные гормоны, олигопептидные факторы роста,
стимуляторы и ингибиторы, разнообразные молекулы-рецепторы,
действующие в мембранах, цитоплазме и хромосомах на уровне
сигналинга, активации транскрипции и трансляции. Ряд химических
канцерогенов имеют сродство с регуляторами пролиферации
(размножения) клеток. Открытие и изучение онкогенов и антионкогенов –
молекулярно-генетическая проблема. Внутриклеточные сети
молекулярного сигналинга – пути к управлению этими процессами.
13.
• Аналитическая и препаративная химия, фармакология. Получениечистых препаратов – стимуляторов и ингибиторов репродукции и
дифференцировки клеток.
• Биотехнология. Клеточно-тканевая инженерия, заместительная
терапия. Проблема стволовых клеток. Культивирование стволовых
клеток, их искусственная детерминация и пересадка в организм, к
больным органам. Выращивание тканей и органов in vitro.
• Таким образом, проблемы репродукции и дифференцировки клеток –
междисциплинарная область биологии, медицины, биотехнологии.
Это область интересов синтетической науки – клеточной биологии.
14. ТЕМА 1. КЛЕТОЧНЫЙ ЦИКЛ
15.
1. Общая характеристика клеточного(митотического) цикла
• Опыты Говарда и Пелка (1953). Открытие митотического цикла.
Корешки конского боба Vicia faba, меристема. Авторадиография с 35РО4.
• РНК из препаратов экстрагировали, так что метка
получалась только от ДНК.
• Показано, что синтез ДНК (редупликация хромосом)
происходит посреди интерфазы.
Рассчитаны параметры цикла: Т = 27 ч (реально 24 ч)
- время генерации, продолжительность цикла.
Выделены G1, S, G2, M – периоды (фазы) митотического цикла:
S – synthesis – синтетический; G1 – gap – пресинтетический, или
постмитотический; G2 – постсинтетический, или премитотический; M –
mitosis – митоз).
• Определение митотического цикла:
Митотический цикл – циклическая последовательность
взаимосвязанных и упорядоченных процессов, обеспечивающих
подготовку к митотическому делению клетки и сам митоз.
16.
• Данные авторадиографии синтеза ДНК вскоре были подтвержденыметодом цитофотометрии содержания ДНК (в реакции Фёльгена) по ходу
митотического цикла. Использованы синхронизированные по циклу
клеточные культуры.
• Показано удвоение массы ДНК (от 2с до 4с) в середине интерфазы.
• Т.о., графическое выражение МЦ возможно как кольцевое, так и
линейное.
17.
• 1960-е годы. Большая работа по определению пролиферативнойактивности и изменчивости митотических циклов в различных тканях
животных и растений, в эмбриогенезах, при патологиях (онкогенез и др.).
• Формируется новое направление – кинетика клеточных популяций.
Важнейшее новое открытие – обнаружение G0 –
периода покоя – обратимого выхода из цикла.
Посреди G1-периода выявлена точка r (restrict
point) – точка рестрикции (ограничения)
возможностей (= check point – контрольная точка).
В это время решается судьба клетки:
1) в новый цикл на S-фазу;
2) в дифференцировку – работу – смерть (некроз,
апоптоз);
3) в период покоя G0 до «особого распоряжения»;
4) в апоптоз – программированная смерть клетки.
Т.о., открылась новая идея для изучения регуляции клеточной репродукции
и дифференцировки через r-точки (check points). Позднее подобные точки
обнаружены по всему циклу.
Но – обо всем по порядку.
18. 2. Методы изучения клеточного цикла и пролиферативной активности клеток
• 2.1. АвторадиографияВключение в ДНК 14С- или 3Н-нуклеозидов (аденозин, тимидин и др.).
Тейлор (1957) впервые применил 3Н-тимидин. Простота и безопасность в
работе, высокое разрешение, точность локализации метки
(мелкозернистые ядерные фотоэмульсии с Ag+).
- Импульсная метка = оценка индекса меченых
S-фазных ядер (ИМЯ), 0-10-20-...-50…%.
ИМЯ=30%
- Насыщение меткой = оценка пролиферативного пула (Р), 0-100%.
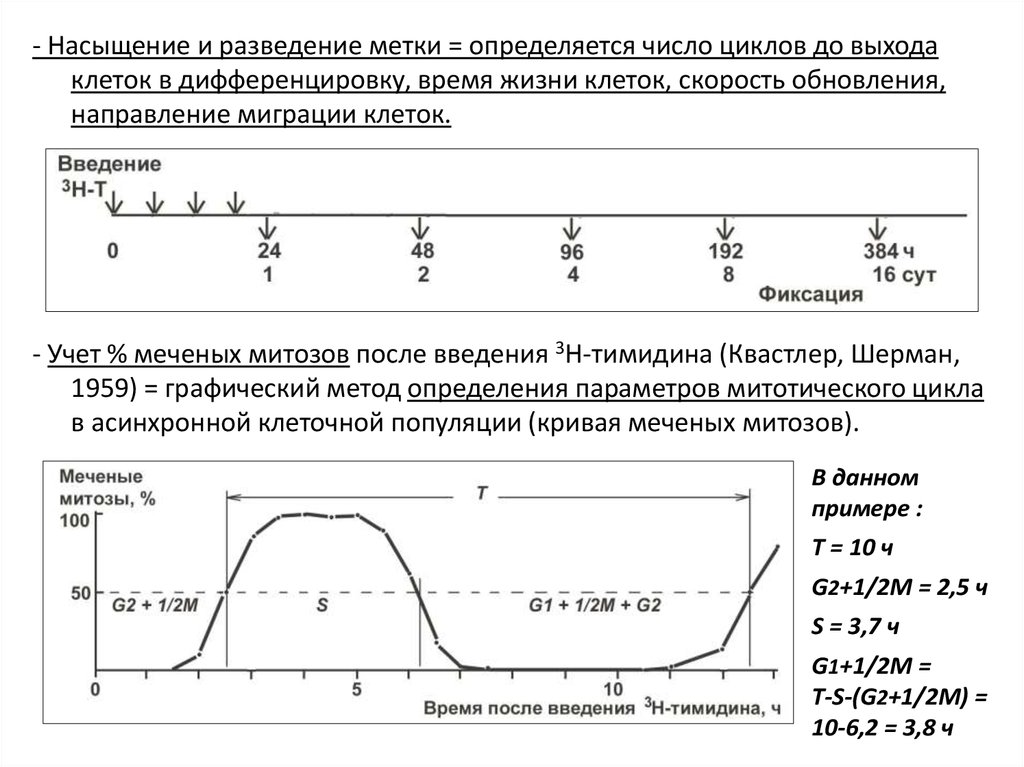
19.
- Насыщение и разведение метки = определяется число циклов до выходаклеток в дифференцировку, время жизни клеток, скорость обновления,
направление миграции клеток.
- Учет % меченых митозов после введения 3Н-тимидина (Квастлер, Шерман,
1959) = графический метод определения параметров митотического цикла
в асинхронной клеточной популяции (кривая меченых митозов).
В данном
примере :
T = 10 ч
G2+1/2M = 2,5 ч
S = 3,7 ч
G1+1/2M =
T-S-(G2+1/2M) =
10-6,2 = 3,8 ч
20.
• 2.2. Цитофотометрия или проточная цитофлюорометрия ДНК (2-4с)- Окраска ДНК по Фельгену на мазках, суспензиях или давленых препаратах.
Цитофотометрический имидж-анализ содержания ДНК (от 2с до 4с) в
отдельных ядрах (необходимо не менее 200-300 клеток на 1 пробу).
- Флюоресцентная микроскопия при окраске ДНК пропидиум-йодидом (PI)
или ди-амино-фенил-индолом (DAPI). Проточная цитофлюорометрия
суспензии клеток (оценивает до 10 тыс. клеток на 1 пробу).
= Оценка пропорции G1-, S-, G2-фазных клеток как критерий пролиферативной
активности, быстрый анализ циклов в диагностике опухолей.
21.
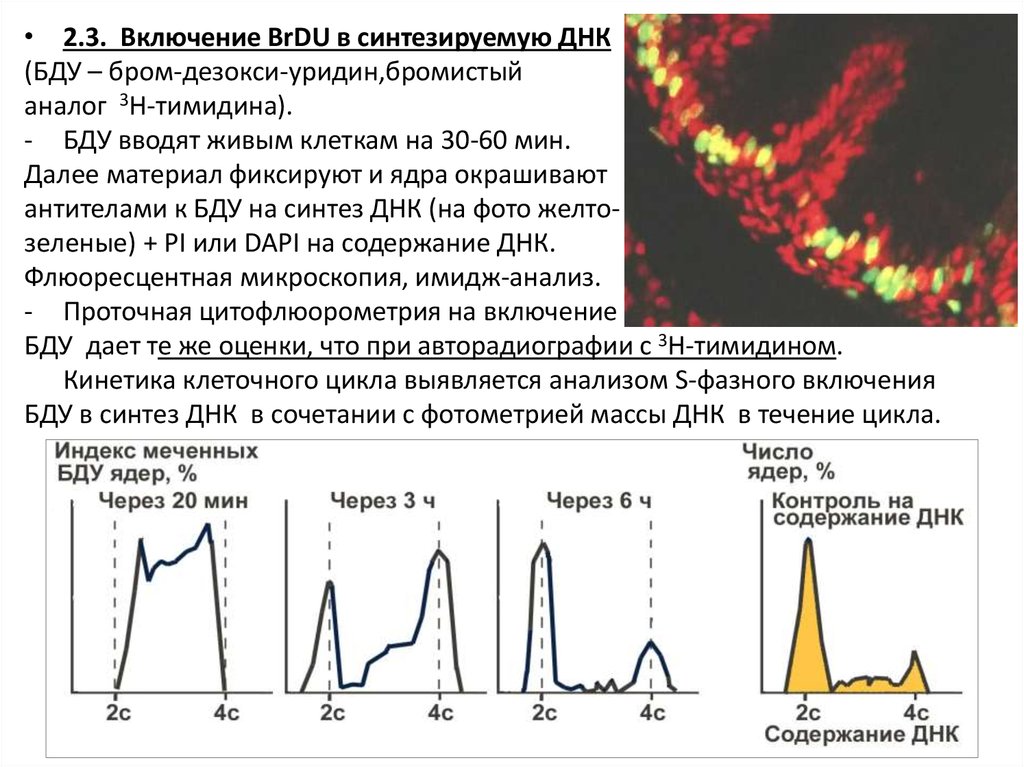
• 2.3. Включение BrDU в синтезируемую ДНК(БДУ – бром-дезокси-уридин,бромистый
аналог 3Н-тимидина).
- БДУ вводят живым клеткам на 30-60 мин.
Далее материал фиксируют и ядра окрашивают
антителами к БДУ на синтез ДНК (на фото желтозеленые) + PI или DAPI на содержание ДНК.
Флюоресцентная микроскопия, имидж-анализ.
- Проточная цитофлюорометрия на включение
БДУ дает те же оценки, что при авторадиографии с 3Н-тимидином.
Кинетика клеточного цикла выявляется анализом S-фазного включения
БДУ в синтез ДНК в сочетании с фотометрией массы ДНК в течение цикла.
22.
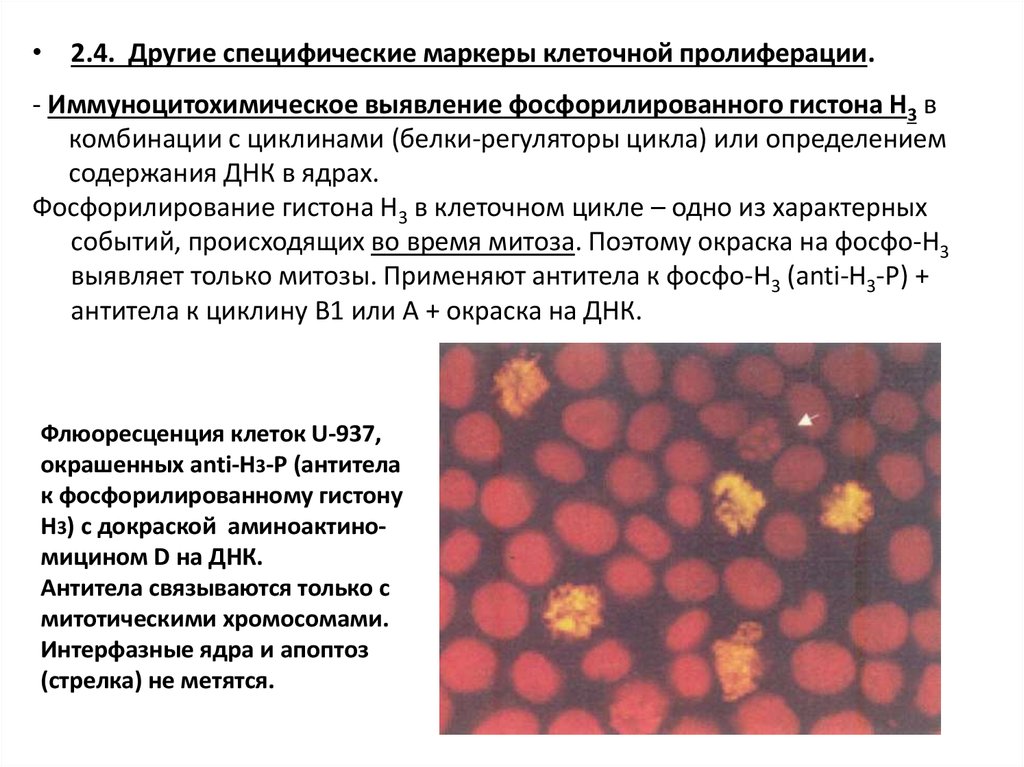
• 2.4. Другие специфические маркеры клеточной пролиферации.- Иммуноцитохимическое выявление фосфорилированного гистона Н3 в
комбинации с циклинами (белки-регуляторы цикла) или определением
содержания ДНК в ядрах.
Фосфорилирование гистона Н3 в клеточном цикле – одно из характерных
событий, происходящих во время митоза. Поэтому окраска на фосфо-Н3
выявляет только митозы. Применяют антитела к фосфо-Н3 (anti-H3-P) +
антитела к циклину B1 или А + окраска на ДНК.
Флюоресценция клеток U-937,
окрашенных anti-H3-P (антитела
к фосфорилированному гистону
Н3) с докраской аминоактиномицином D на ДНК.
Антитела связываются только с
митотическими хромосомами.
Интерфазные ядра и апоптоз
(стрелка) не метятся.
23.
= Проточная цитофлюорометрия на фосфо-Н3 выявляет митотическийиндекс – % клеток в М-фазе по отношению ко всем клеткам или
другим фазам цикла (если сочетать с оценкой содержания ДНК):
24.
- Иммуноцитохимическое выявление белка Ki-67 в циклирующихклетках.
Моноклональные антитела Ki-67 были получены в Кильском университете
(Univ. of Kiel), Германия. 67 – номер удачного клона.
Сам белок Ki-67 – это антиген неясной локализации, связанный с
пролиферацией; выявляется во всех активных фазах митотического
цикла: G1, S, C2, M, но только не в G0.
= Проточная
цитофлюорометрия
оценивает общий
пролиферативный пул и
соотношение клеток в разных
периодах цикла.
В нормальных (не опухолевых)
клетках преобладает G0-период
(по сути дифференцировка), хотя
есть и циклирующие клетки.
В опухоли G0-клетки практически
отсутствуют, всё пролиферирует).
25.
-Иммуноцитохимическое выявление циклоспецифического антигена
PCNA (Proliferating Cell Nuclear Antigen) с помощью соответствующих
антител.
PCNA – одна из субъединиц ДНК-полимеразного комплекса, поэтому antiPCNA связывается только в S-фазных ядрах. Это – филогенетически
древние (консервативные) молекулы. Имеют значительную гомологию у
животных, дрожжей, растений.
= Проточная цитофлюорометрия
выявляет в ядрах уровень
экспрессии PCNA в сочетании с
оценкой содержания ДНК (для
определения фаз цикла) –
фактически это S-фазный индекс
(ИМЯ в авторадиографии с 3Нтимидином).
26. 3. Репликация ДНК
• 3.1. Подготовка к синтезу ДНК.• Подготовка к синтезу ДНК запускается во 2-й
половине G1-периода, после точки check point
(точка r).
• Для синтеза ДНК необходимо наличие 4
видов дезоксирибонуклеотидов (ДНтидов):
тимидиловый ТТФ, адениловый АТФ,
гуаниловый ГТФ и цитидиловый ЦТФ.
• Метаболический фонд нуклеотидов невелик.
В основном он состоит из рибонуклеозидов,
которые и являются источником образования
дезоксирибонуклеотидов.
Для этого происходит:
энзиматическое восстановление рибозы до дезоксирибозы: РНзиды
преобразуются в ДНзиды;
и далее фосфорилирование: нуклеозиды переходят в нуклеотиды.
27.
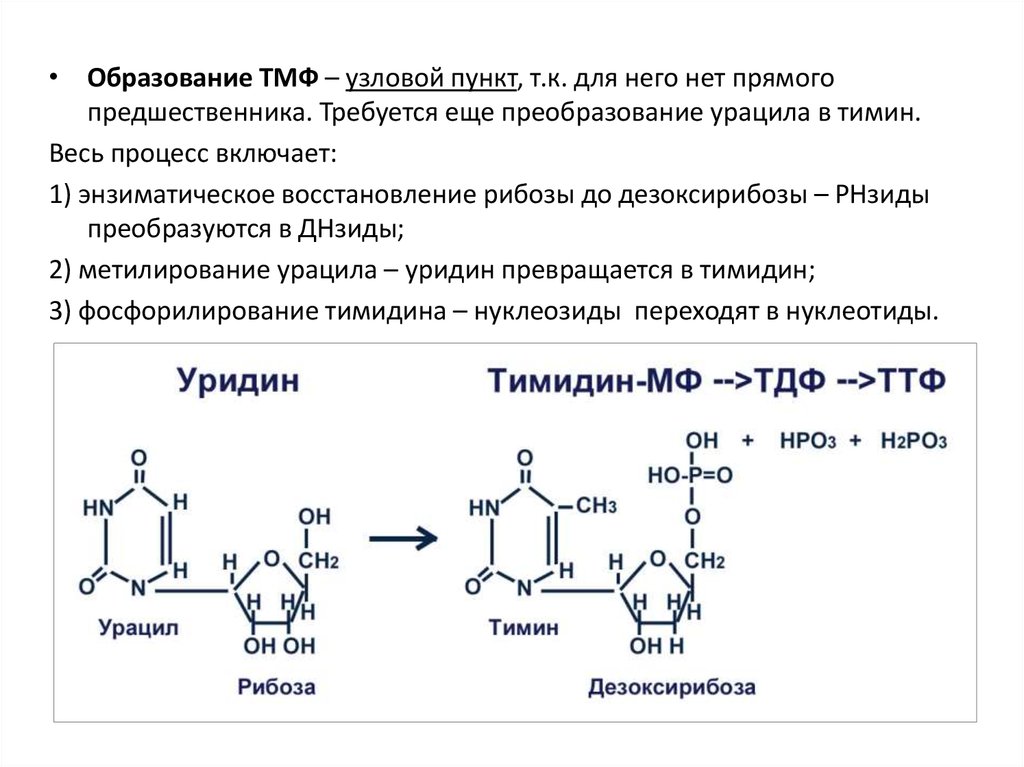
• Образование ТМФ – узловой пункт, т.к. для него нет прямогопредшественника. Требуется еще преобразование урацила в тимин.
Весь процесс включает:
1) энзиматическое восстановление рибозы до дезоксирибозы – РНзиды
преобразуются в ДНзиды;
2) метилирование урацила – уридин превращается в тимидин;
3) фосфорилирование тимидина – нуклеозиды переходят в нуклеотиды.
28.
• Фосфорилирование нуклеозидов происходит с помощью специфическихкиназ. Но не все киназы имеются всегда в одинаковом количестве.
Адениловая, гуаниловая и цитидиловая киназы имеются всегда, а
тимидиловая киназа практически отсутствует в покоящихся клетках и
появляется только в цикле, в короткий срок перед S-фазой.
• С другой стороны, происходит катаболизм предшественников. Кислая
фосфатаза способна дефосфорилировать ТМФ, выступая антагонистом
киназы.
[Опыты с 3Н-тимидином. У земляной белки (низкая физиологическая
активность) включение тимидина слабое, у золотистого хомячка
(высокая активность) – сильное. Оказалось, что у белки резко увеличена
активность ферментов катаболизма тимидина и др. нуклеотидов.
Возможно, те же различия имеются у мидии и гребешка?].
• Т.о., активность киназ тимидиловой системы и их антагонистов фосфатаз
управляет пулом тимидинфосфатов и является лимитирующим
фактором в подготовке к репликации ДНК. Синтез и распад нуклеотидов
перед S-фазой – один из механизмов регуляции цикла (с помощью
специальных киназ и циклинов, Глава 3).
29.
• 3.2. Полуконсервативный механизм репликации ДНК.• Молекула ДНК – двухцепочечная. Теоретически возможны 3 способа ее
репликации: консервативный, полуконсервативный, дисперсный.
• При разработке модели строения и удвоения ДНК Уотсон и Крик
исходили гипотетически из полуконсервативного принципа: разрыв
водородных связей, расхождение цепей, достройка на них
комплементарных цепей (в соответствии с правилом Чаргафа: А-Т, Г-Ц).
В результате каждая новая молекула ДНК должна иметь одну нить от
исходной молекулы, а вторую вновь синтезированную.
• Но метода доказательства не было. А уже появились наблюдения
фрагментации ДНК, сестринских хроматидных обменов (СХО). Позднее
СХО были определены как мутации, но сначала это могло
свидетельствовать в пользу дисперсного механизма репликации ДНК.
• Первые исследования выполнены изотопными методами.
30.
• 1) Опыты с 3Н-тимидином (Тейлор, 1957-1963).Корешки Vicia faba. Импульсное (0,5—1 час) мечение 3Н-тимидином S-фазы
1-го цикла. Отмывка и дальнейший рост (2-й цикл) без меченого
тимидина.
Фиксация на митозы 1-го цикла (через 3-5 час.) и 2-го цикла (через сутки):
Первая волна митозов: во всех хромосомах мечены обе хроматиды.
Вторая волна митозов: в каждой хромосоме мечена только одна хроматида.
• Эти данные объясняются только исходя из полуконсервативного принципа
репликации:
31.
• 2) Аналогичные опыты с БДУ (фон Латт, 1973).Обычная ДНК (с тимидином) хорошо красится по Гимза и флюоресцирует с
Хёст-33258. Участки с включенным БДУ (вместо тимидина) плохо красятся
и не флюоресцируют.
• Клетки китайского хомячка культивировали in vitro в течение 2 поколений
в среде с БДУ.
• Хроматиды красились (и светились) по-разному:
- сильно крашенные – исходные, без метки БДУ,
- средне крашенные – с 1 БДУ-замещенной нитью ДНК,
- слабо крашенные – с 2 БДУ-замещенными нитями ДНК.
32.
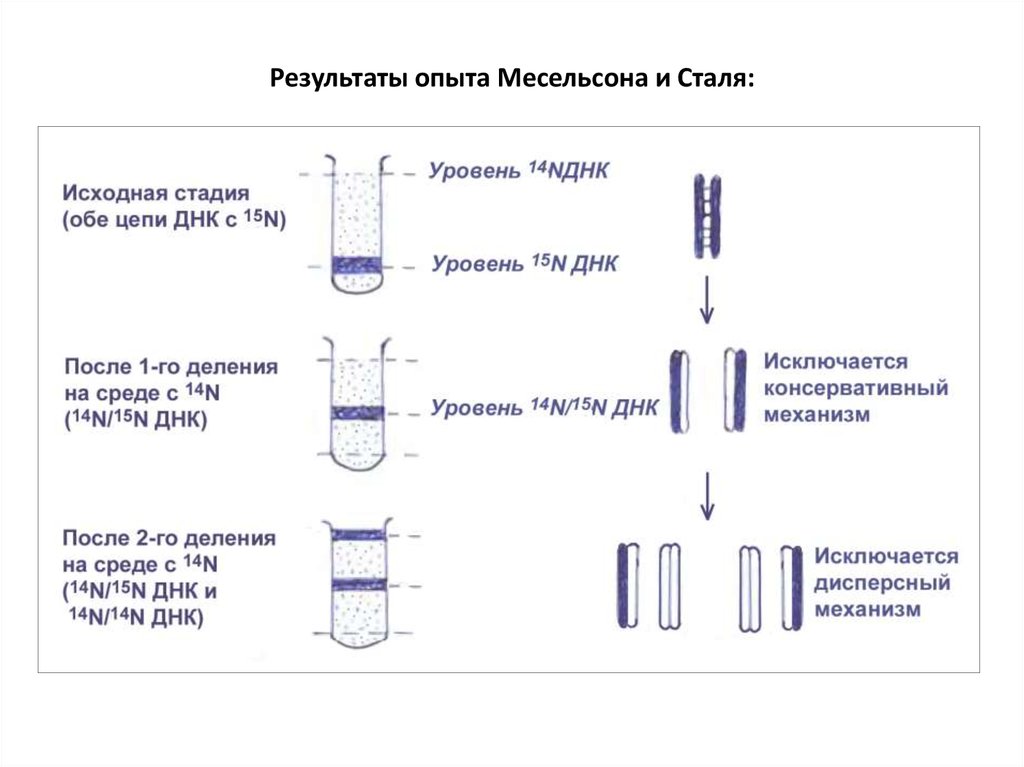
• 3) Опыты c 14N и 15N на E. coli (Месельсон, Сталь, 1957, 1958).У бактерий одна кольцевая хромосома, упакованная в нуклеоид.
Удвоение ДНК происходит за 20-30 мин, и сразу наступает деление
клетки.
• Биохимический подход. Разработан метод равновесного
центрифугирования в солевом растворе для разделения ДНК разной
удельной плотности: легкая с 14N и тяжелая 15N.
• E. coli выращивали 10 поколений на среде с 15NH4Cl. Практически
весь азот заместился на тяжелый 15N.
Далее культура перенесена на среду с обычным 14NH4Cl. Начался синтез
легкой ДНК.
В первом и втором циклах ДНК выделяли и центрифугировали.
33.
Результаты опыта Месельсона и Сталя:34.
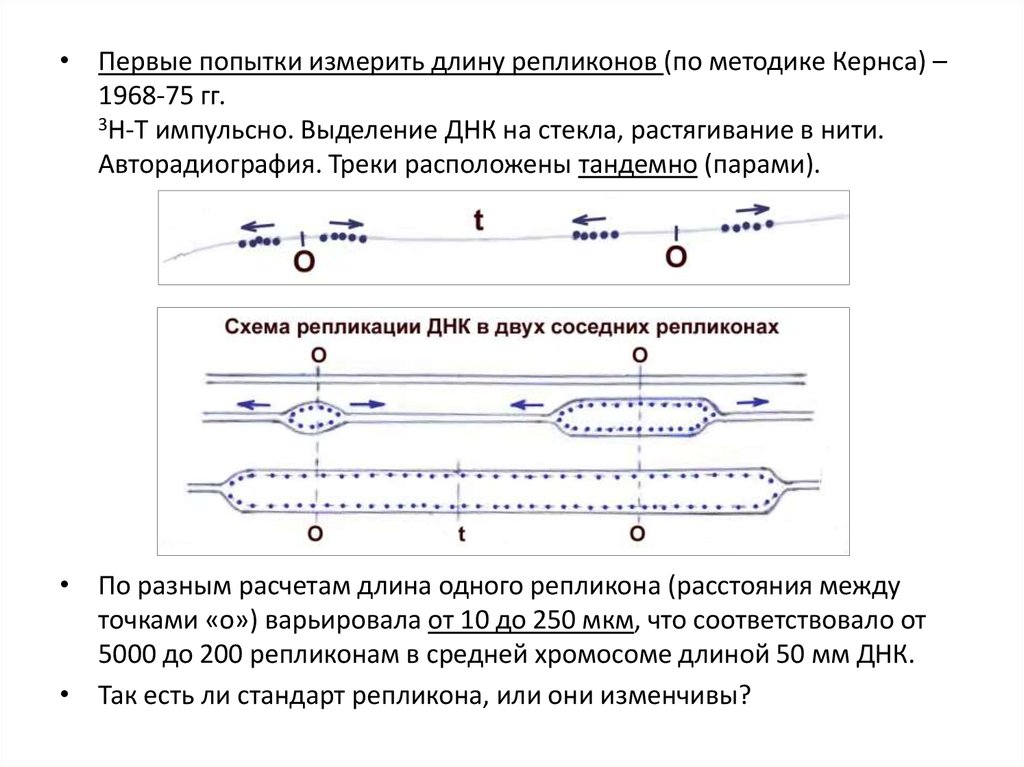
• Опыты с 3Н-тимидином на E. сoli (Кернс, 1963).Включение 3Н-Т, разрушение клеток с развертыванием нуклеоида и
высвобождением ДНК на предметном стекле, фиксация и
авторадиография через 5, 10, 20 ... мин от начала S-периода.
• Наблюдали образование 2-го кольца ДНК по полуконсервативному
механизму.
35.
3.3. Репликоны. Моно- и полирепликонные хромосомы.• У прокариот (уже в опытах Кернса на E. сoli)
выявлены закономерности репликации ДНК:
• - репликация начинается в определенной
стартовой точке «О» (origin),
• - репликация идет сразу в 2 направлениях
(2 репликативные вилки) и заканчивается в
точке «T» (terminal),
• - вся хромосома – одна единица репликации,
один репликон (то же самое показано у фага
λ, вируса SV40).
• Т.о., у прокариот и вирусов –
монорепликонные хромосомы.
• У эукариот все оказалось много сложнее.
36.
• Информация к размышлению:- В 2n наборе хромосом человека содержится 6 пкг (6х10-12 г) ДНК (6х109 пн).
то около 2 м, по 20-80 мм в отдельных хромосомах (сравни с 2 мм у E. сoli).
- Скорость репликации у эукариот (50 пн/сек) на порядок ниже, чем у
прокариот (500 пн/сек). С такой скоростью при монорепликонной
организации хромосом период синтеза ДНК длился бы сотни часов; на
самом деле он занимает до 5-10 ч.
- Уже в ранних работах (Тейлор, 1960) показано, что импульсная метка 3Н-Т
обнаруживается сразу во многих участках хромосомы. Особенно хорошо
это видно на политенных хромосомах дрозофилы. Каждый диск
(хромомер) – отдельный репликон?
• Т.о., по длине хромосомы имеется множество точек инициации синтеза
ДНК. Каждый автономно реплицирующийся участок– репликон.
Хромосомы эукариот полирепликонные.
37.
• Первые попытки измерить длину репликонов (по методике Кернса) –1968-75 гг.
3Н-Т импульсно. Выделение ДНК на стекла, растягивание в нити.
Авторадиография. Треки расположены тандемно (парами).
• По разным расчетам длина одного репликона (расстояния между
точками «о») варьировала от 10 до 250 мкм, что соответствовало от
5000 до 200 репликонам в средней хромосоме длиной 50 мм ДНК.
• Так есть ли стандарт репликона, или они изменчивы?
38.
• Дополнительная информация к размышлению.У одного и того же организма длительность S-фазы (tS) на разных стадиях
развития и в разных зачатках (органах) сильно меняется.
У саламандры: эмбриональные клетки ранней бластулы
tS = 1 ч;
дифференцированные взрослые клетки
tS = 48 ч;
премейотический синтез в сперматоцитах tS = 200 ч.
У морского ежа: первые деления дробления - Т = 1,5-2 ч, tS = 0,8 ч;
поздняя гаструла
- Т = 50-60 ч, tS = 25-30 ч.
У дрозофилы: первые циклы дробления
- Т = 10 мин, tS = 3,5 мин !
(расчетная длина репликона = 3-4 мкм).
клеточная культура от взрослых мух - все на порядок больше.
Т.о., время S-фазы удлиняется от нескольких минут до нескольких часов.
При этом во всех случаях скорость движения репликативных вилок
примерно одинаковая!
Следовательно, в ходе развития уменьшается число точек инициации и,
соответственно, увеличивается длина репликонов. Большинство точек,
активных на ранних стадиях, не используются (отключаются) на более
поздних стадиях.
39.
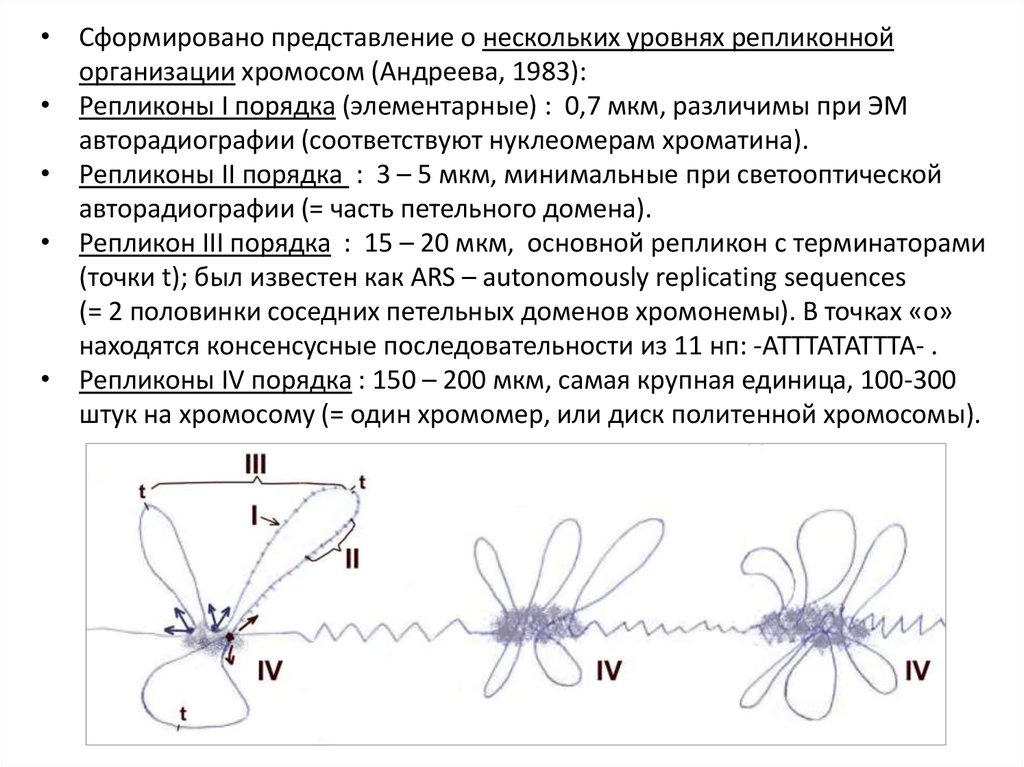
• Сформировано представление о нескольких уровнях репликоннойорганизации хромосом (Андреева, 1983):
• Репликоны I порядка (элементарные) : 0,7 мкм, различимы при ЭМ
авторадиографии (соответствуют нуклеомерам хроматина).
• Репликоны II порядка : 3 – 5 мкм, минимальные при светооптической
авторадиографии (= часть петельного домена).
• Репликон III порядка : 15 – 20 мкм, основной репликон с терминаторами
(точки t); был известен как ARS – autonomously replicating sequences
(= 2 половинки соседних петельных доменов хромонемы). В точках «о»
находятся консенсусные последовательности из 11 нп: -АТТТАТАТТТА- .
• Репликоны IV порядка : 150 – 200 мкм, самая крупная единица, 100-300
штук на хромосому (= один хромомер, или диск политенной хромосомы).
40.
• Т.о., в онтогенезе большинство точек «о» постепенно теряютинициирующую активность, длина регистрируемых репликонов,
соответственно, увеличивается.
• Сначала, в раннем эмбриогенезе, включены все элементарные
репликоны, потом по 1 точке в репликонах II порядка, далее III, IV.
• Имеем сложный многоуровневый (иерархичный) механизм регуляции
репликации ДНК. Иерархические уровни репликации ДНК отражают
дискретность структурной организации хроматина.
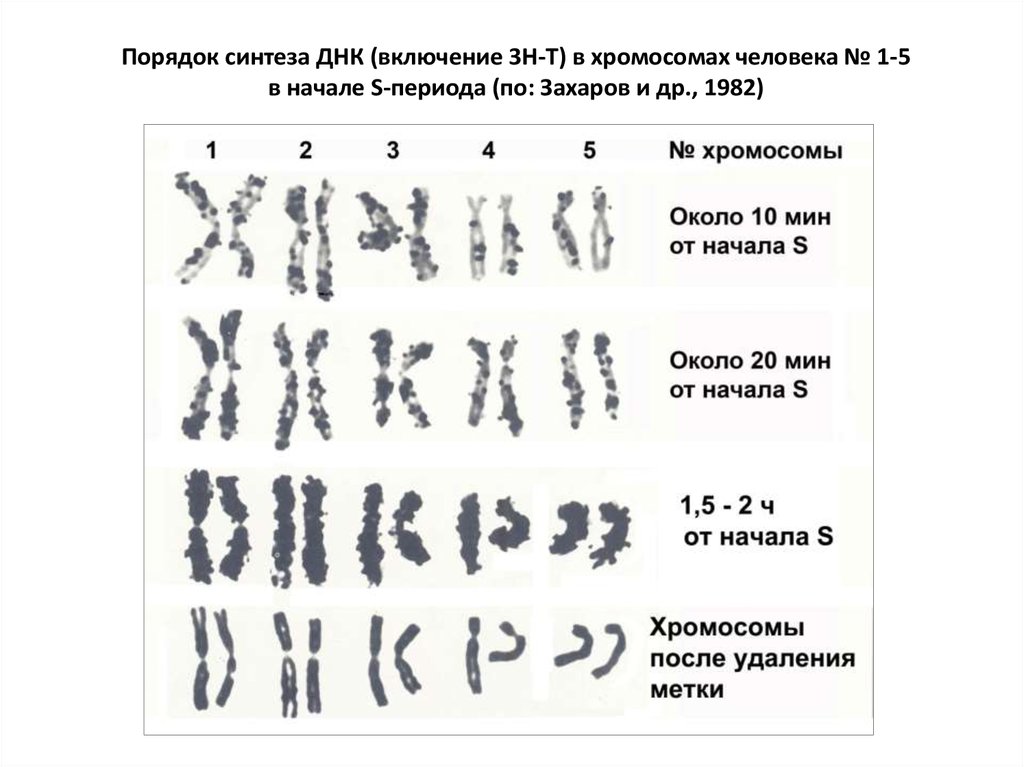
41. 3.4. Хронология синтеза ДНК. Рано и поздно реплицирующиеся участки
• В первых циклах дробления зиготы репликоны инициируются почтиодновременно. В делящихся (стволовых) клетках взрослого организма
соседние малые репликоны (I-II порядков) тоже синхронизированы
(последовательное расположение тандемов).
Но репликоны III-IV порядков инициируются асинхронно. Т.о., по длине
хромосомы и при сравнении разных хромосом наблюдается
асинхронность репликации ДНК.
• Однако это не означает беспорядочности репликации. Время начала и
окончания репликации в тех или иных хромосомах (график репликации)
сохраняется неизменным в ряду клеточных делений. Каждая хромосома
характеризуется высокой стабильностью порядка репликации, имеет
специфический рисунок репликации.
42.
Порядок синтеза ДНК (включение 3Н-Т) в хромосомах человека № 1-5в начале S-периода (по: Захаров и др., 1982)
43.
• При этом обнаружена универсальная закономерность:- в первой половине S-фазы реплицируется эухроматин, во второй –
гетерохроматин (правило поздней репликации гетерохроматина).
Lima-de-Faria (1959) впервые обнаружил это на сперматоцитах
кузнечика: в гетерохроматической Х-хромосоме ДНК синтезировалась
только в конце S-периода. То же выявлено в клетках растений,
млекопитающих, моллюсков и др.
Распределение метки в хромосомах человека (46, ХХ) при включении 3Н-Т
в начале S-периода
и
в конце S-периода
(стрелкой показана Х-хромосома) (Захаров и др., 1982)
Так возникла проблема рано и поздно реплицирующейся ДНК.
44.
Справка. В интерфазе клеточного цикла, при переходе хромосом в хроматин
определенные участки остаются в конденсированном состоянии; различимы
эухроматин и гетерохроматин (Heitz,1928). Гетерохроматин локализован в
основном по периферии ядра. Brown (1966): гетерохроматин есть
факультативный и структурный (конститутивный).
Факультативный гетерохроматин – не работающие конденсированные гены.
Яркий пример – Х-хромосома (тельце Бара, половой хроматин) у
млекопитающих и человека.
Структурный гетерохроматин – постоянно конденсированный в одних и тех же
районах обеих гомологичных хромосом (теломерный, центромерный,
интеркалярный, околоядрышковый). Образован разными видами
сателлитной ДНК, многоповторными последовательностями нуклеотидов.
• Оказалось, что правило Лима-де-Фария справедливо только для
факультативного гетерохроматина, который действительно реплицируется
в конце S-фазы.
СатДНК реплицируется на протяжении всего S-периода, причем каждая
фракция имеет свой пик синтеза в определенной части S.
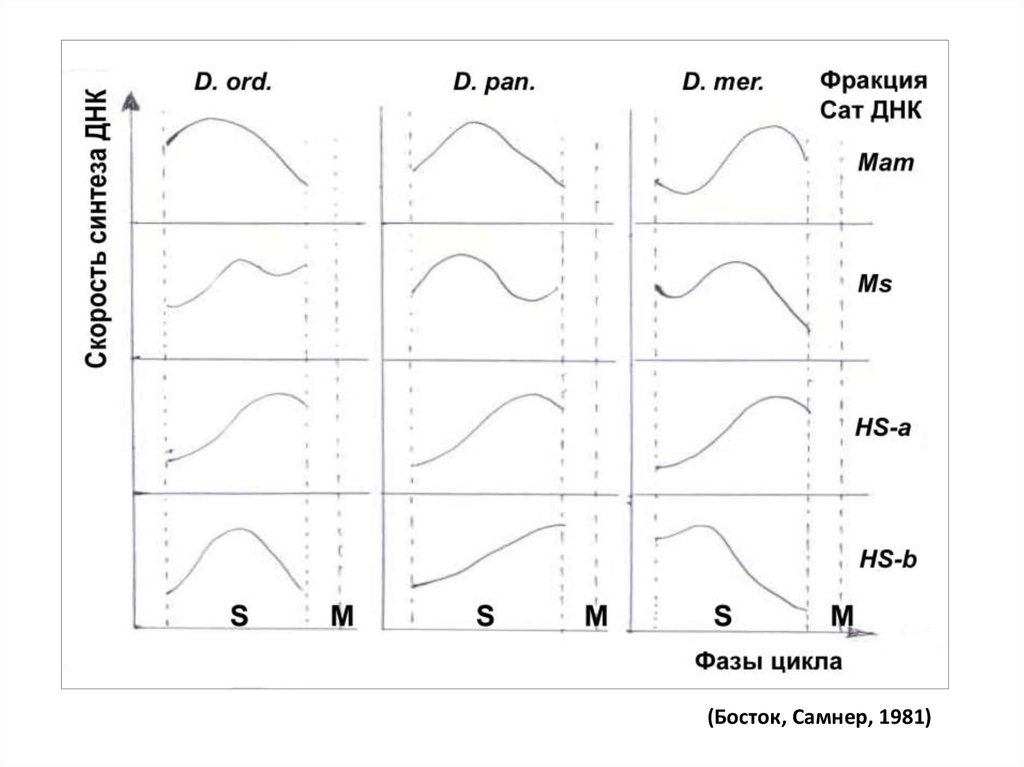
Пример: на 3 видах кенгуровых крыс выявляли время репликации разных
фракций сатДНК:
45.
(Босток, Самнер, 1981)46.
• Выводы: по репликации гетерохроматиновой ДНК:• - Синтез разных фракций сатДНК может идти в начале, середине или
конце S-фазы;
• - Хронология репликации видоспецифична;
• - Распространенное представление о поздней репликации
гетерохроматина справедливо только в отношении факультативного
гетерохроматина.
• - Эухроматиновая ДНК реплицируется преимущественно в первой
половине S-фазы, но отдельные локусы реплицируются в середине и
даже в конце S-фазы. Например, репликация ядрышковых
организаторов:
NB: У хомячка и крысы
гены рРНК (ЯОР)
локализованы в 1 паре
хромосом, у человека –
в 5 парах. Каждая пара
хромосом имеет свой
график репликации.
(Босток, Самнер, 1981)
47.
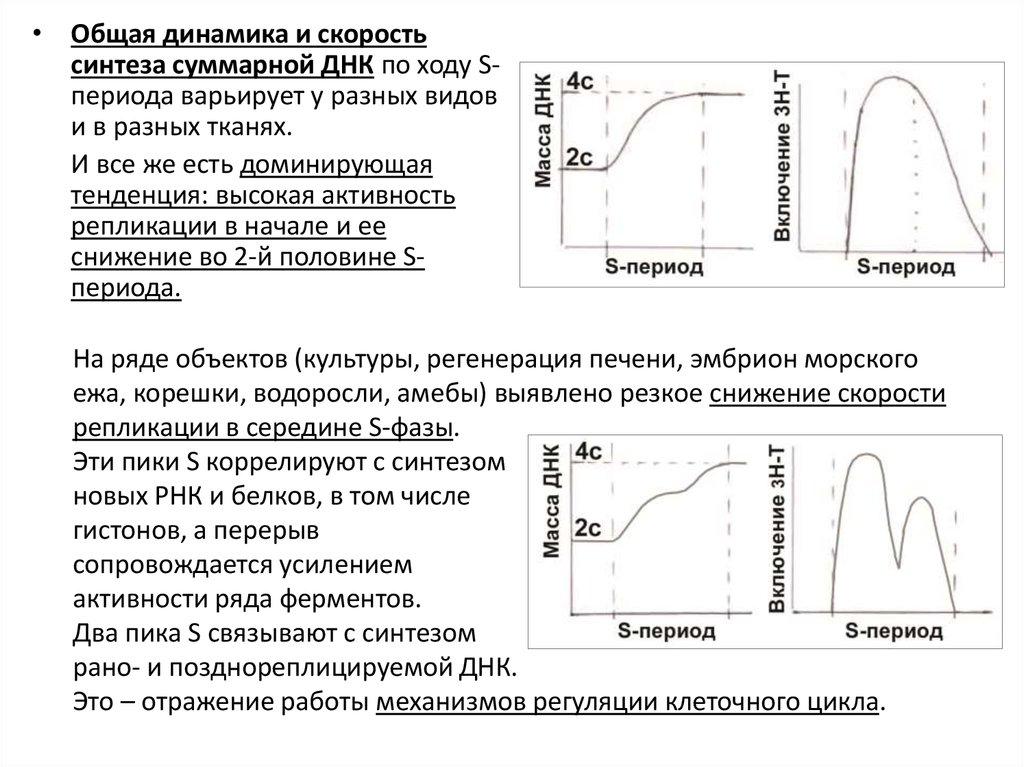
• Общая динамика и скоростьсинтеза суммарной ДНК по ходу Sпериода варьирует у разных видов
и в разных тканях.
И все же есть доминирующая
тенденция: высокая активность
репликации в начале и ее
снижение во 2-й половине Sпериода.
На ряде объектов (культуры, регенерация печени, эмбрион морского
ежа, корешки, водоросли, амебы) выявлено резкое снижение скорости
репликации в середине S-фазы.
Эти пики S коррелируют с синтезом
новых РНК и белков, в том числе
гистонов, а перерыв
сопровождается усилением
активности ряда ферментов.
Два пика S связывают с синтезом
рано- и позднореплицируемой ДНК.
Это – отражение работы механизмов регуляции клеточного цикла.
48.
• 3.5. Особенности репликации теломерной ДНК.• Теломерная ДНК – консервативная сателлитная
высокоповторная ДНК.
• Выделена, клонирована и расшифрована у
многих организмов:
- человек и другие позвоночные (более 100
видов): ……………………………….. (TTAGGG)n;
- беспозвоночные: та же или близкая
последовательность;
- тетрахимена: …………………….. (TTGGGG)n;
- растения: …………………………… (TTTAGGG)n.
Число повторов (n) варьирует у разных
видов: максимальное n у мыши – 100 тыс. нп, у
обезьяны – 25 тыс. нп, у человека – 12 тыс. нп.
Но число повторов не одинаково и в
разных типах клеток одного организма,
например у человека в разных зрелых клетках
n= 250-1500 нп.
49.
Роль теломерной сатДНК :
Обеспечивает индивидуальность и интактность хромосом. При утрате
теломеров образуются индуцированные концы хромосом, которые
деградируют или связываются с
другими хромосомами
(в митозе образуются мосты).
Предотвращают разрушение эухроматической (=генетической) части
хромосомы при множественной репликации. Концы хромосом сокращаются
в каждом цикле из-за недорепликации по последнему РНК-праймеру (по
модели Оказаки). Сокращается теломерная (сателлитная!) ДНК, а геномная
ДНК до поры до времени защищена.
За теломером имеется субтеломерный участок (другая сатДНК, разная у
разных видов). Это дополнительный буфер между теломером и генами.
• На этом построена теломерная гипотеза старения клеток (основана А.М.
Оловниковым, 1970-е годы):
Соматические клетки при каждом делении теряют часть теломерных
повторов, которые изначально защищают генетическую зону от укорочения
(разрушения). Когда теломеры и субтеломеры израсходуются, разрушаются
концевые гены, разрушаются хромосомы. Клетка погибает.
50.
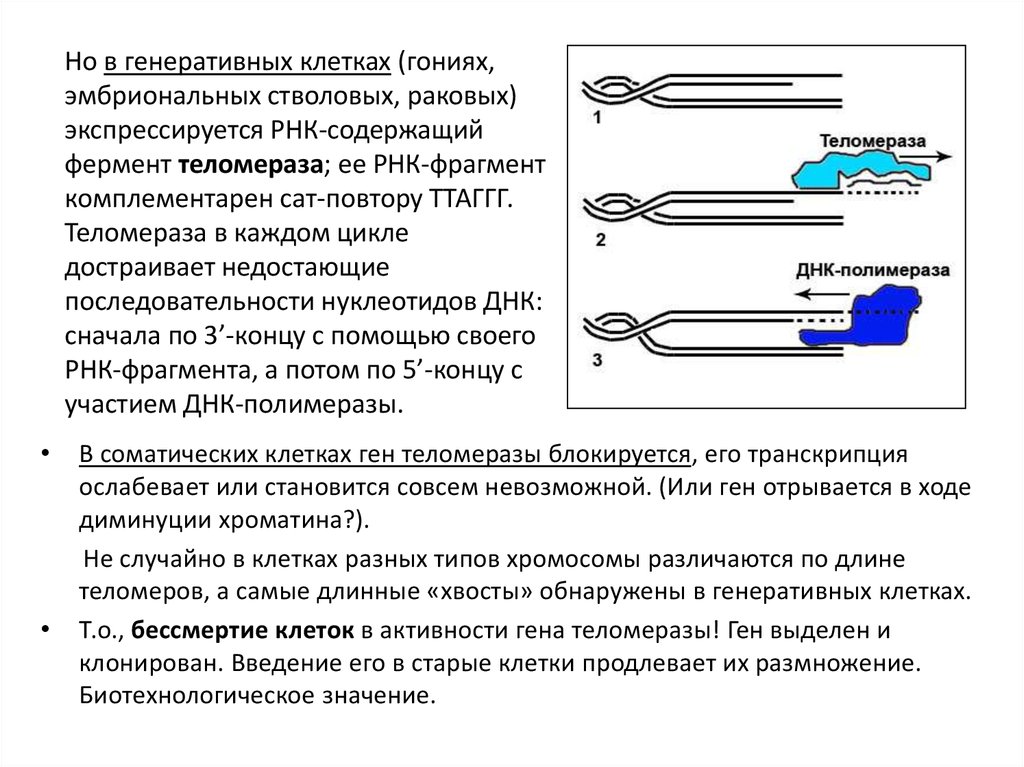
Но в генеративных клетках (гониях,эмбриональных стволовых, раковых)
экспрессируется РНК-содержащий
фермент теломераза; ее РНК-фрагмент
комплементарен сат-повтору ТТАГГГ.
Теломераза в каждом цикле
достраивает недостающие
последовательности нуклеотидов ДНК:
сначала по 3’-концу с помощью своего
РНК-фрагмента, а потом по 5’-концу с
участием ДНК-полимеразы.
В соматических клетках ген теломеразы блокируется, его транскрипция
ослабевает или становится совсем невозможной. (Или ген отрывается в ходе
диминуции хроматина?).
Не случайно в клетках разных типов хромосомы различаются по длине
теломеров, а самые длинные «хвосты» обнаружены в генеративных клетках.
Т.о., бессмертие клеток в активности гена теломеразы! Ген выделен и
клонирован. Введение его в старые клетки продлевает их размножение.
Биотехнологическое значение.
51.
• 3.6. Репаративный синтез ДНК.• В ДНК постоянно возникают ошибки и нарушения по азотистым
основаниям или сахаро-фосфатному остову.
Ошибки – спонтанные (происходят при репликации или
рекомбинации ДНК).
Нарушения – индуцированные (происходят под действием УФ
света, R-лучей, химических веществ).
• Но в клетке есть гены, обеспечивающие синтез не только
реплицирующих, но и репарирующих ферментов. Обычно эти
ферменты совмещены в ДНК-полимеразных комплексах.
52.
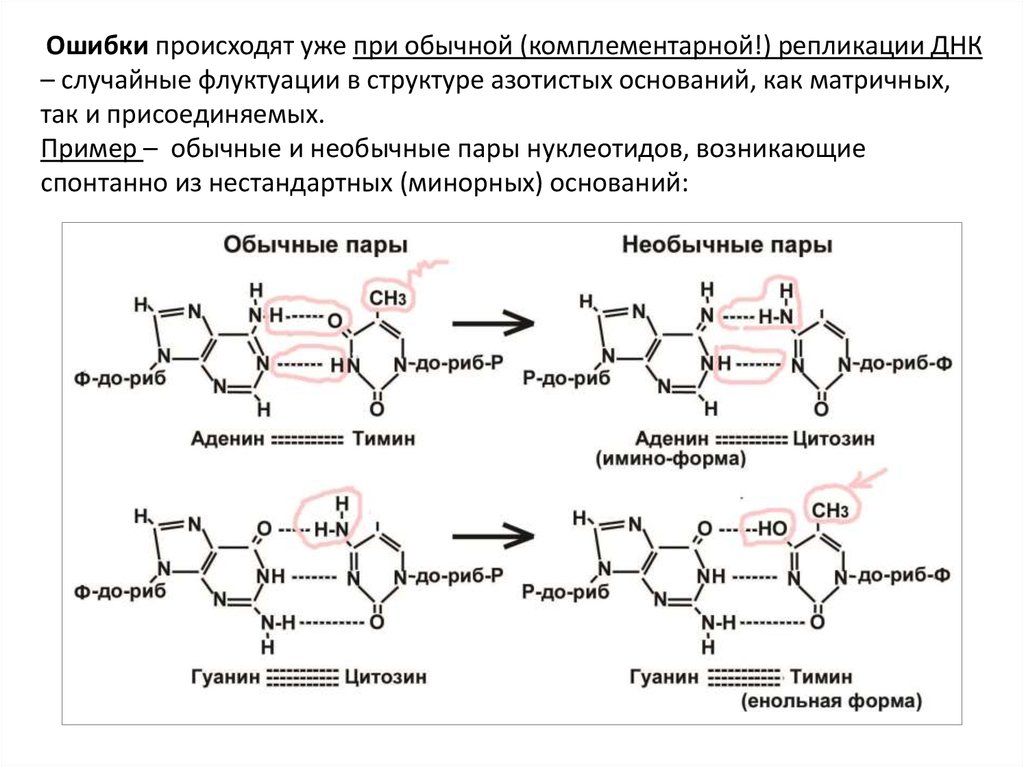
Ошибки происходят уже при обычной (комплементарной!) репликации ДНК– случайные флуктуации в структуре азотистых оснований, как матричных,
так и присоединяемых.
Пример – обычные и необычные пары нуклеотидов, возникающие
спонтанно из нестандартных (минорных) оснований:
53.
В этих случаях рост цепи сразу останавливается, и экзонуклеаза (домен в
составе ДНК-полимеразы Pol I) удаляет необычный нуклеотид. Т.о., уже в
репликативной вилке, контролируется каждый новый нуклеотид. В случае
мутации гена экзонуклеазы Pol I неправильные нуклеотиды не устраняются
и закрепляются в форме новых мутаций.
В репарации и репликации участвуют также:
Pol II (заполняет пробелы между фрагментами Оказаки),
Pol III (ключевая полимераза по хромосомной ДНК E. coli),
ДНК-лигазы (соединяют цепи ДНК при репликации, репарации и
рекомбинации),
топоизомеразы I и II (разрывы и сшивки на «острие» репликативной вилки
– шарнир).
Нарушения (повреждения) – нарушения остова ДНК, модификации
оснований или ошибочные спаривания оснований при рекомбинации (в
ходе кроссинговера).
2-цепочечные повреждения в одном месте, в том числе 2-цепочечные
разрывы, они всегда летальны. (Уникальное исключение – Т4-лигаза E. coli:
при внедрении фага Т4 соединяет 2-цепочечные фрагменты на концах
разрыва.)
54.
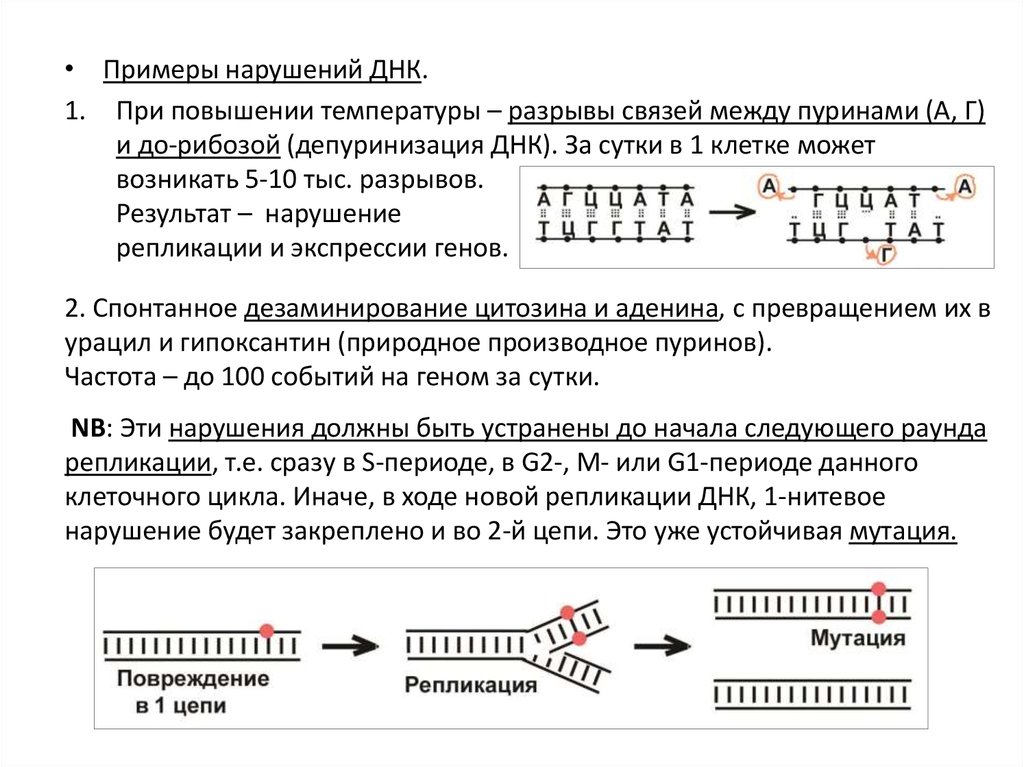
• Примеры нарушений ДНК.1. При повышении температуры – разрывы связей между пуринами (А, Г)
и до-рибозой (депуринизация ДНК). За сутки в 1 клетке может
возникать 5-10 тыс. разрывов.
Результат – нарушение
репликации и экспрессии генов.
2. Спонтанное дезаминирование цитозина и аденина, с превращением их в
урацил и гипоксантин (природное производное пуринов).
Частота – до 100 событий на геном за сутки.
NB: Эти нарушения должны быть устранены до начала следующего раунда
репликации, т.е. сразу в S-периоде, в G2-, M- или G1-периоде данного
клеточного цикла. Иначе, в ходе новой репликации ДНК, 1-нитевое
нарушение будет закреплено и во 2-й цепи. Это уже устойчивая мутация.
55.
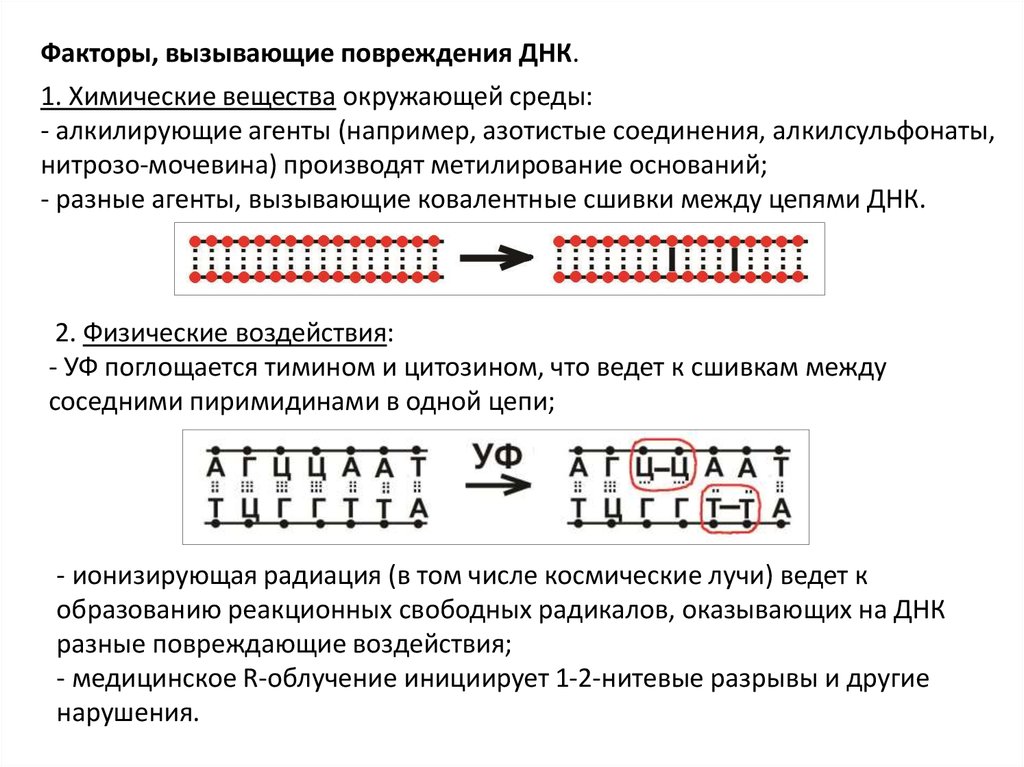
Факторы, вызывающие повреждения ДНК.1. Химические вещества окружающей среды:
- алкилирующие агенты (например, азотистые соединения, алкилсульфонаты,
нитрозо-мочевина) производят метилирование оснований;
- разные агенты, вызывающие ковалентные сшивки между цепями ДНК.
2. Физические воздействия:
- УФ поглощается тимином и цитозином, что ведет к сшивкам между
соседними пиримидинами в одной цепи;
- ионизирующая радиация (в том числе космические лучи) ведет к
образованию реакционных свободных радикалов, оказывающих на ДНК
разные повреждающие воздействия;
- медицинское R-облучение инициирует 1-2-нитевые разрывы и другие
нарушения.
56.
• Как это восстанавливается до начала следующей репликации?1. Прямое восстановление исходной структуры:
- специальные ферменты могут просто снимать метильные группы,
полученные под действием алкилирующих агентов;
- ферменты фотолиазы (фотореактивирующие ферменты бактерий и
низших эукариот) под действием видимого света превращают
пиримидиновые димеры, полученные при УФ облучении, в нормальные
мономеры.
57.
• 2. Замена модифицированных оснований:(1) фермент узнает повреждение ДНК и надрезает цепь вблизи или
отрывает основание от до-рибозы;
(2) экзонуклеаза удаляет нуклеотид или целую группу, оставляя брешь;
(3) удаленный участок синтезируется заново с 3’-конца по целой цепи;
(4) ДНК-лигаза сшивает концы разрыва.
• Могут использоваться и эндонуклеазы, например разнообразные APэндонуклеазы (апуриновые, апиримидиновые), где произошла
депуринизация и депиримидинизация. Разрушается участок из
нескольких нуклеотидов, потом он ресинтезируется.
58.
• Пиримидиновые димеры вырезаются специальными нуклеазамивлево и вправо, всего на 12 нуклеотидов. Далее пробел заполняется с
помощью полимеразы и лигазы:
59.
Значение репарации ДНК.Повреждения ДНК блокируют репликацию (передачу информации в
ряду поколений) или сохраняются в геномах потомков в форме мутаций.
Поврежденная ДНК еще более уязвима. Поэтому повреждения надо
устранять.
Мутации по генам ферментов репарации – прямой путь к мутациям
структурных генов, т.е. к разнообразным генетическим заболеваниям,
включая онкологические .
В частности, у людей с пигментной ксеродермой нарушена способность
к выщеплению пиримидиновых димеров из УФ-облученной ДНК
(мутация в 1 из 9 генов). Даже при слабом воздействии солнечного
света развивается рак кожи.
60. 4. Синтез РНК и белков в клеточном цикле
• Размножение клеток, как правило, сопровождается их ростом,примерно в 2 раза на цикл.
• (Старое представление о «критической массе», необходимой для
начала деления. Опыты с амебой: периодическое отрезание части
цитоплазмы ведет к бесконечной задержке митоза.)
• Исключения: дробление зиготы и некоторые специальные случаи.
• В основе роста клетки лежат синтезы РНК и белков (транскрипция и
трансляция генетической программы).
61.
• 4.1. Синтез РНК.• Необходим синтез и процессинг 3 форм РНК – иРНК, тРНК, рРНК (не
считая регуляторных мяРНК).
• Гены пре-иРНК (10-80s РНК) локализованы диффузно по эухроматину
всех хромосом. Это уникальные или умеренно (10-20) повторенные
последовательности; в геноме человека около 25 тыс. таких генов.
Пре-иРНК – подвергается процессингу и сплайсингу в перихроматиновых
гранулах – информоферах (разрушается до 80-90 % интронной РНК).
Готовая иРНК (сшитые экзонные фрагменты) выходит через ядерные
поры в цитоплазму, связывается с новыми белками и образует
информосомы.
• Гены пре-тРНК (4s РНК) – локализованы так же диффузно; это умеренные
повторы. В геноме 61 вид генов тРНК (по числу кодонов). Процессинг
идет без сплайсинга.
• Гены пре-рРНК (45s РНК) – локализованы в ядрышках (ЯОР, NOR), они
присутствуют в фибриллярных центрах и фибриллярном компоненте
ядрышка. Это 1 вид гена, он образует умеренные повторы. У человека до
500 повторов в 5 парах хромосом.
45s РНК процессирует без сплайсинга до 18s, 28s и 5,8s рРНК.
(5s рРНК синтезируется на других хромосомах).
62.
63.
• Количество РНК в клетке вне цикла остается постоянным, т.к. синтез ираспад уравновешены. Но при вступлении клетки в митотический
цикл синтезы начинают преобладать, и общее содержание РНК
увеличивается.
• - Данные цитофотометрии: общая масса РНК или только рРНК в ходе
интерфазы примерно удваивается.
• - Морфологические данные: увеличение объема ядрышек в
циклирующих клетках (регенерирующая печень, крипты кишечного
эпителия и др.).
• Динамика синтеза и процессинга РНК исследуется на
синхронизированных клеточных культурах (или в асинхронных
клеточных популяциях) при одновременном учете ДНК (фаза цикла) и
РНК.
Оценивают включение в РНК 3Н-уридина или флуоресцентно
меченного урацила (в форме УТФ), активность РНК-полимеразы I
(ядрышковая), РНК-полимеразы-II (хроматиновая).
64.
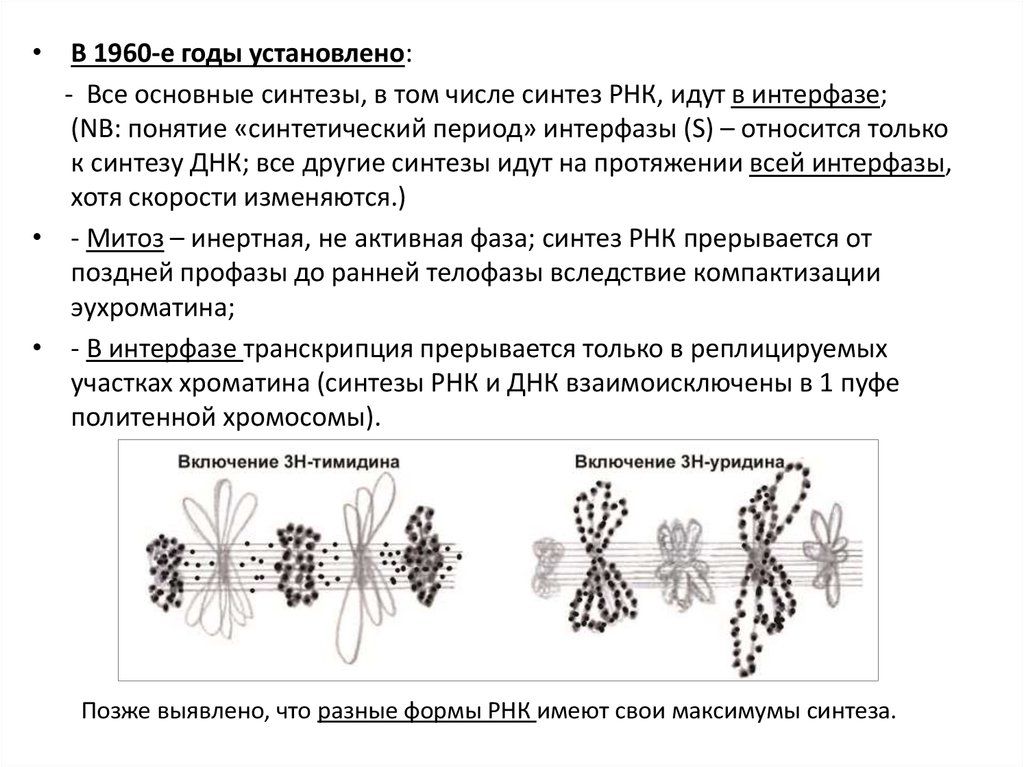
• В 1960-е годы установлено:- Все основные синтезы, в том числе синтез РНК, идут в интерфазе;
(NB: понятие «синтетический период» интерфазы (S) – относится только
к синтезу ДНК; все другие синтезы идут на протяжении всей интерфазы,
хотя скорости изменяются.)
• - Митоз – инертная, не активная фаза; синтез РНК прерывается от
поздней профазы до ранней телофазы вследствие компактизации
эухроматина;
• - В интерфазе транскрипция прерывается только в реплицируемых
участках хроматина (синтезы РНК и ДНК взаимоисключены в 1 пуфе
политенной хромосомы).
Позже выявлено, что разные формы РНК имеют свои максимумы синтеза.
65.
• Синтез рРНК (ядрышковая, рибосомная) – показано разобщениетранскрипции и процессинга.
• Volpe et al. (1976). Культура клеток человека HeLa,
синхронизированная по фазам цикла.
Импульсная метка 3Н-уридина, интервал проб – 1 час.
• Суммарную РНК выделяли и фракционировали центрифугированием
по скорости седиментации в градиенте сахарозы. Выделяли
ядрышковые фракции РНК: 45s (первичный транскрипт), 32s
(промежуточный процессинг), 28s и 18s (конечные рРНК формы).
• Радиоактивность фракций РНК, отражающую скорость синтеза (45s)
или процессинга (32s, 28s, 18s), оценивали сценциляционным
счетчиком.
• Результат:
66.
• Синтез 45s РНК (пре-рРНК) резко возрастает в G1-периоде (быстрыйрост клетки после митоза), снижается в S и G2, прекращается в M.
• Процессинг идет в 2 волны; на самом деле это 1 волна, прерванная
митозом:
- ранний процессинг (в S-G2) дает первую партию рибосом, поздний
процессинг (продолжение волны в G1) дает последнюю партию.
• Синтез 45s РНК и ее процессинг (появление 32s, 18s, 28s РНК)
разобщены во времени, каждый имеет свою динамику.
67.
• Синтез иРНК – показан эффект дозы генов.• Многочисленные исследования по включению 3Н-уридина в
суммарную РНК ядра или только в РНК хроматина (без ядрышка).
Отсюда разнобой результатов.
• Детальные исследования группы Pfeiffer (1968) на
синхронизированной культуре HeLa, подтвержденные позже на
других объектах (др. культуры, растения, инфузории, плесень).
• Применяли 3Н-уридин импульсно (0,5-1 час) в разных точках цикла.
• Результат:
68.
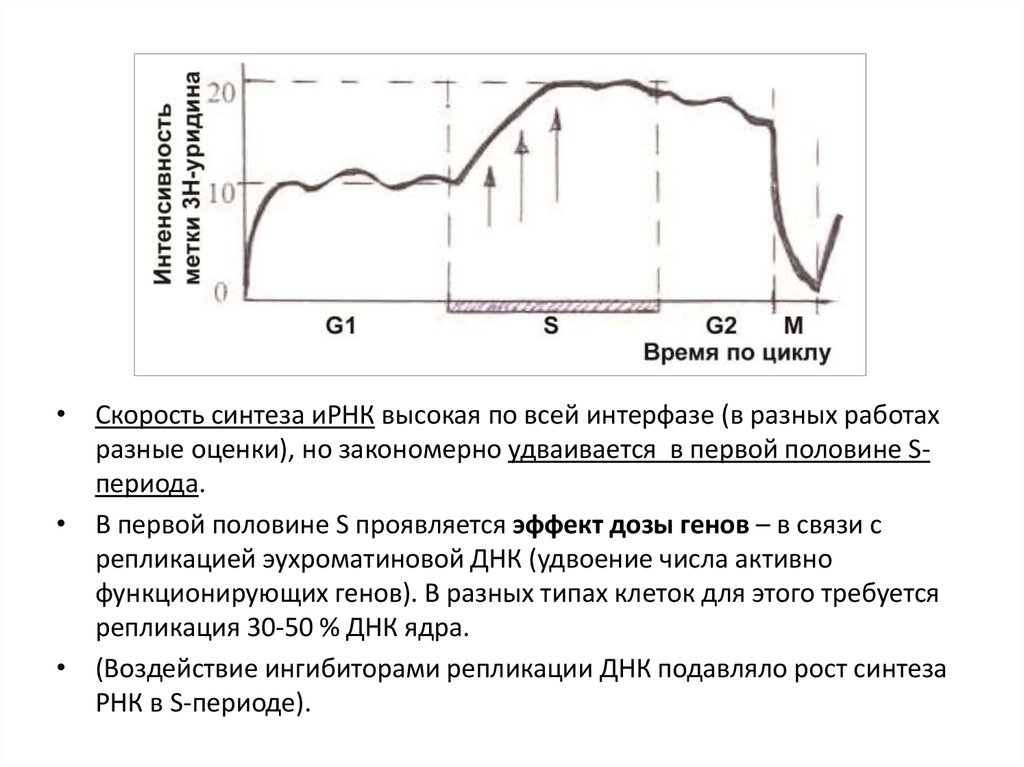
• Скорость синтеза иРНК высокая по всей интерфазе (в разных работахразные оценки), но закономерно удваивается в первой половине Sпериода.
• В первой половине S проявляется эффект дозы генов – в связи с
репликацией эухроматиновой ДНК (удвоение числа активно
функционирующих генов). В разных типах клеток для этого требуется
репликация 30-50 % ДНК ядра.
• (Воздействие ингибиторами репликации ДНК подавляло рост синтеза
РНК в S-периоде).
69.
• NB: Четкая зависимость синтеза РНК от дозы генов выявляется невсегда.
• Надо учитывать:
• Артефакты: несовершенство методик, плохая синхронизация цикла.
• Различия клеток: степень их митотической потентности (стволовости),
степень дифференцировки, т.е. разное соотношение ауто- и
гетеросинтезов РНК.
• Изменения активности РНК-полимераз или их поступления в ядро.
• Колебания пула нуклеотидов – общих предшественников РНК и ДНК.
• Возможность изменения проницаемости мембран и т.д. и т.п.
70.
• 4.2. Синтез белков.Синтез белков выявляют по включению в живые клетки меченых
аминокислот: 3Н- или 14С- лейцин, глицин, пролин, метионин и др.
• Радиоактивную метку регистрируют на микроскопических препаратах
(авторадиография) или в выделенных белках (счетчиком).
• Используют клеточные культуры, синхронизированные по фазам
цикла.
• Результаты по синтезу общих (суммарных) белков очень различаются
в зависимости от типа клеток, их возраста, используемого
предшественника и др. факторов.
• Но есть и общие тенденции.
71.
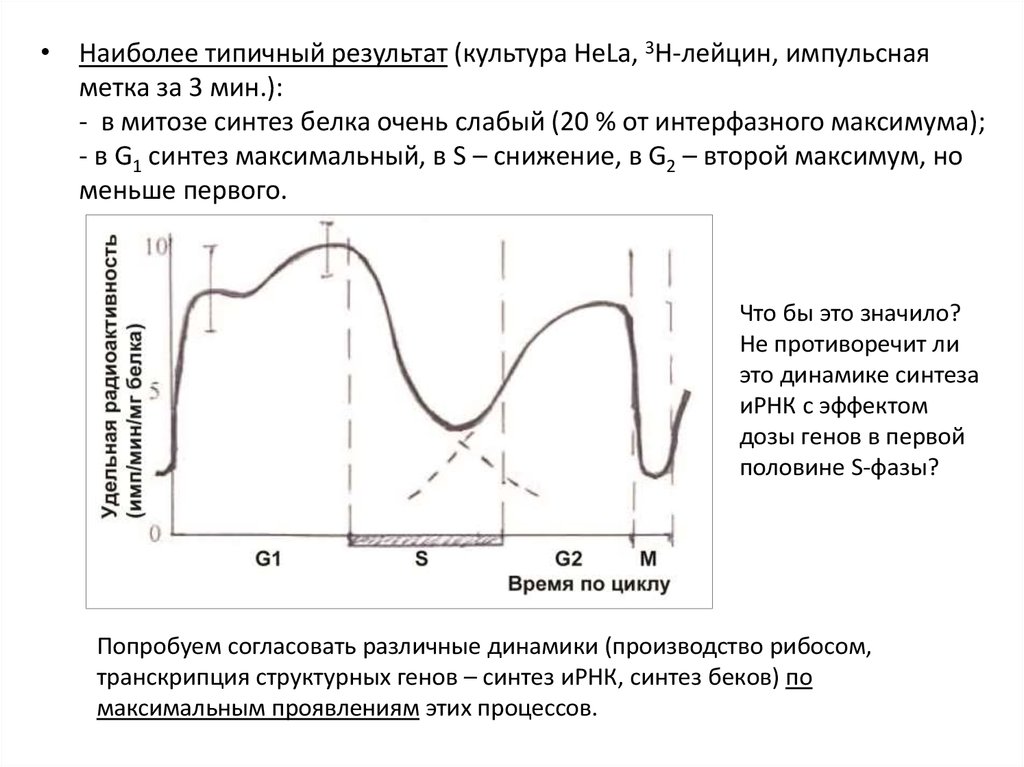
• Наиболее типичный результат (культура HeLa, 3Н-лейцин, импульснаяметка за 3 мин.):
- в митозе синтез белка очень слабый (20 % от интерфазного максимума);
- в G1 синтез максимальный, в S – снижение, в G2 – второй максимум, но
меньше первого.
Что бы это значило?
Не противоречит ли
это динамике синтеза
иРНК с эффектом
дозы генов в первой
половине S-фазы?
Попробуем согласовать различные динамики (производство рибосом,
транскрипция структурных генов – синтез иРНК, синтез беков) по
максимальным проявлениям этих процессов.
72.
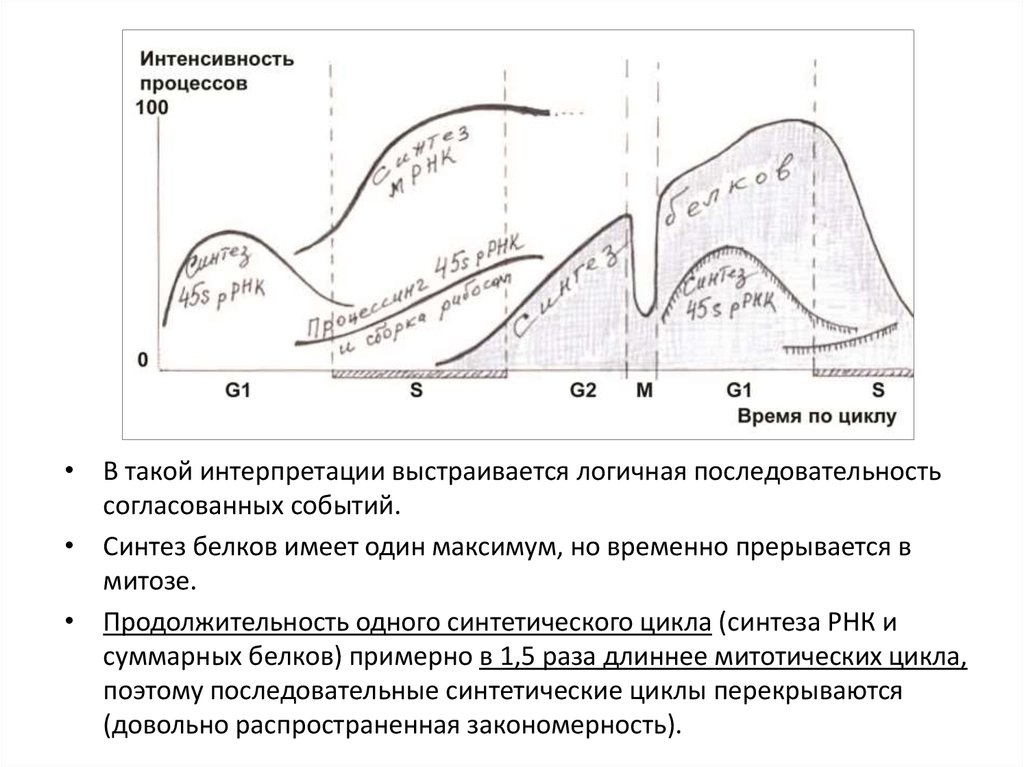
• В такой интерпретации выстраивается логичная последовательностьсогласованных событий.
• Синтез белков имеет один максимум, но временно прерывается в
митозе.
• Продолжительность одного синтетического цикла (синтеза РНК и
суммарных белков) примерно в 1,5 раза длиннее митотических цикла,
поэтому последовательные синтетические циклы перекрываются
(довольно распространенная закономерность).
73.
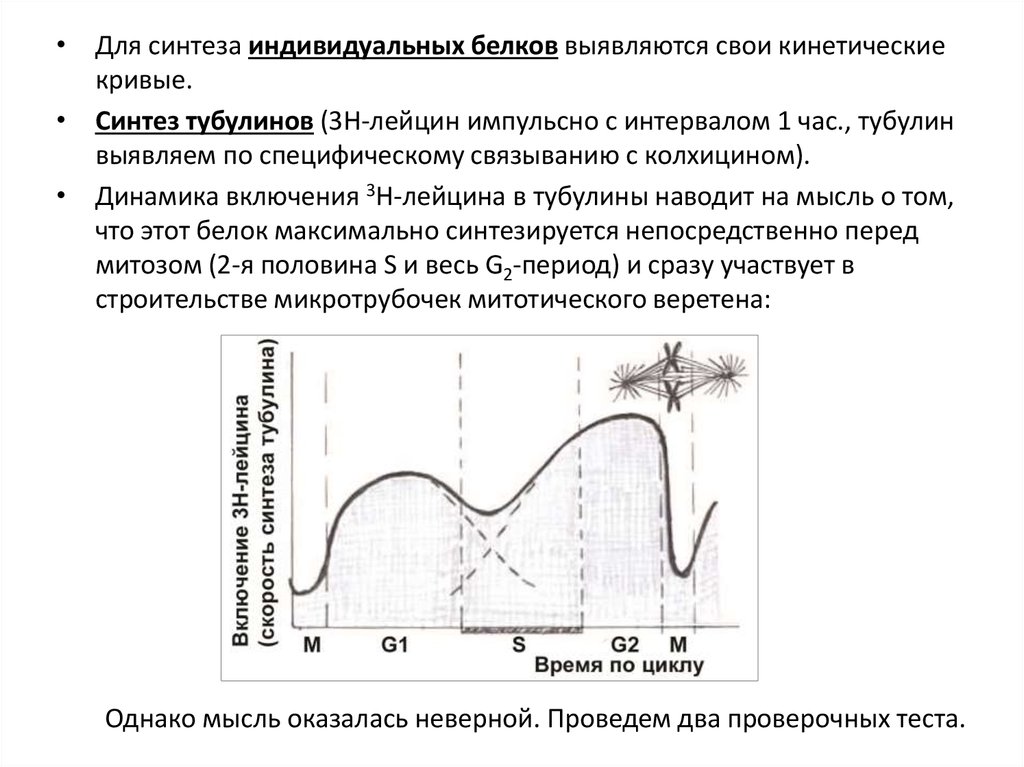
• Для синтеза индивидуальных белков выявляются свои кинетическиекривые.
• Синтез тубулинов (3Н-лейцин импульсно с интервалом 1 час., тубулин
выявляем по специфическому связыванию с колхицином).
• Динамика включения 3Н-лейцина в тубулины наводит на мысль о том,
что этот белок максимально синтезируется непосредственно перед
митозом (2-я половина S и весь G2-период) и сразу участвует в
строительстве микротрубочек митотического веретена:
Однако мысль оказалась неверной. Проведем два проверочных теста.
74.
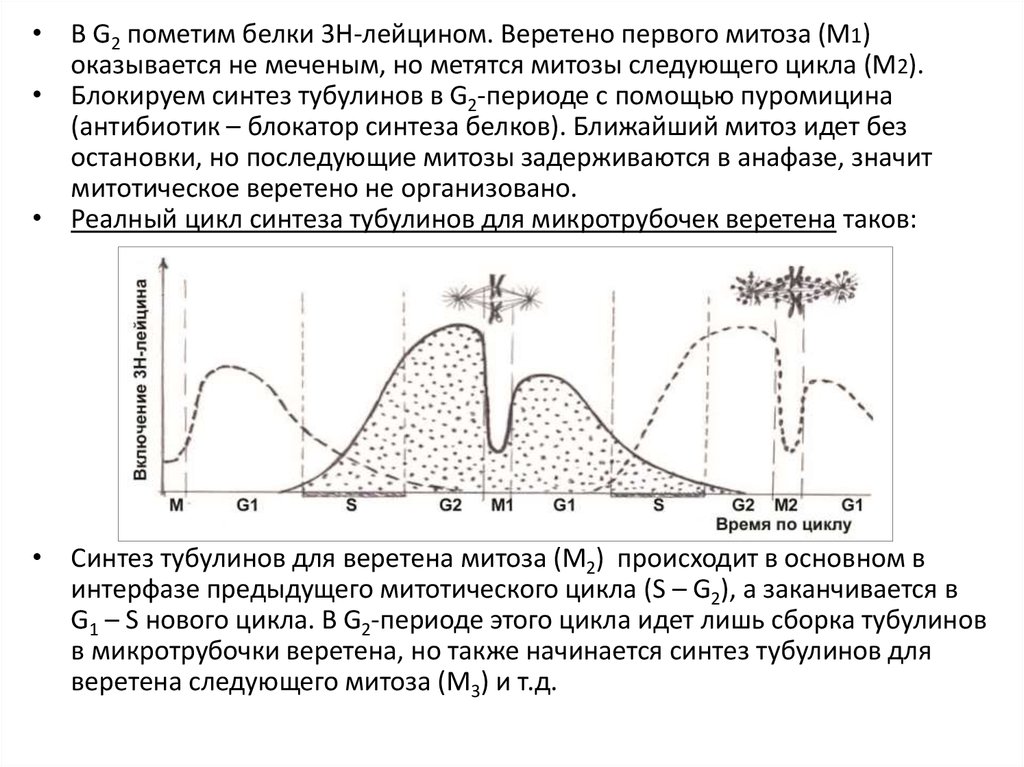
• В G2 пометим белки 3Н-лейцином. Веретено первого митоза (М1)оказывается не меченым, но метятся митозы следующего цикла (М2).
• Блокируем синтез тубулинов в G2-периоде с помощью пуромицина
(антибиотик – блокатор синтеза белков). Ближайший митоз идет без
остановки, но последующие митозы задерживаются в анафазе, значит
митотическое веретено не организовано.
• Реалный цикл синтеза тубулинов для микротрубочек веретена таков:
• Синтез тубулинов для веретена митоза (М2) происходит в основном в
интерфазе предыдущего митотического цикла (S – G2), а заканчивается в
G1 – S нового цикла. В G2-периоде этого цикла идет лишь сборка тубулинов
в микротрубочки веретена, но также начинается синтез тубулинов для
веретена следующего митоза (М3) и т.д.
75.
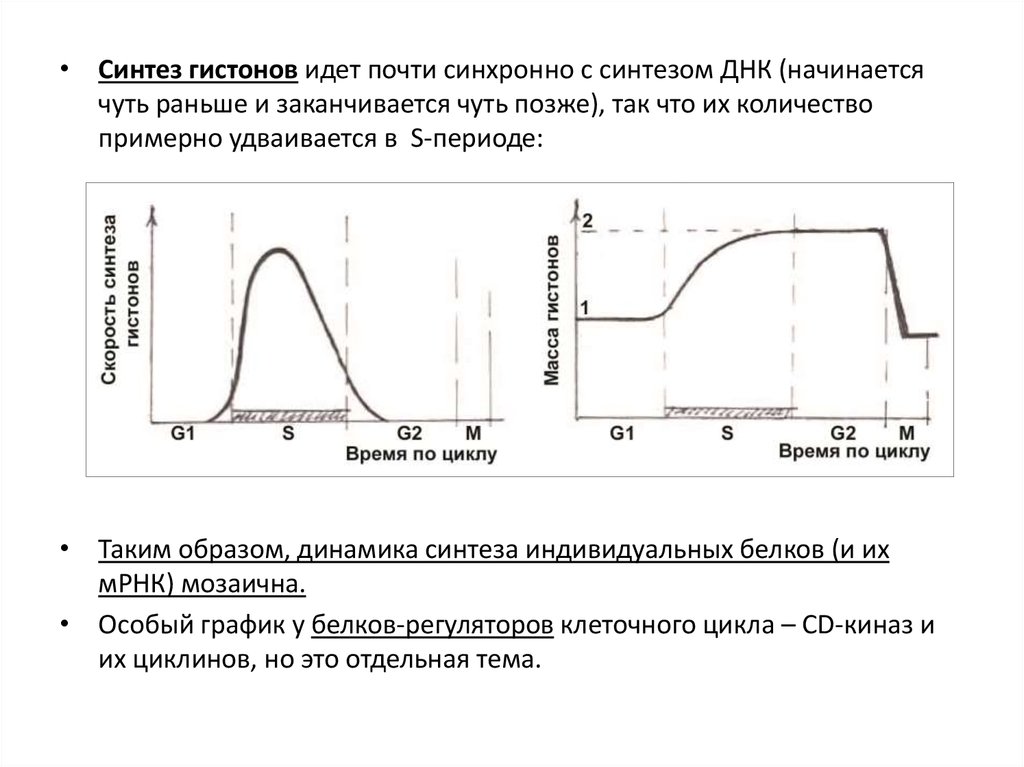
• Синтез гистонов идет почти синхронно с синтезом ДНК (начинаетсячуть раньше и заканчивается чуть позже), так что их количество
примерно удваивается в S-периоде:
• Таким образом, динамика синтеза индивидуальных белков (и их
мРНК) мозаична.
• Особый график у белков-регуляторов клеточного цикла – CD-киназ и
их циклинов, но это отдельная тема.
76. 5. Митоз. Хромосомный цикл
В митозе связаны воедино преобразования хромосом – хромосомный цикл,формирование и работа веретена – центросомный цикл и разделение
клеточного тела – цитокинетический цикл. Начнем с хромосомного цикла.
5.1. Концепция структурной непрерывности хромосом
В интерфазе клеточного цикла морфологически обособленные хромосомы
не видны. Однако они здесь присутствуют в форме хроматина – в
деконденсированном (декомпактизованном, деспирализованном)
состоянии.
- В конденсированном состоянии (метаболически не активном), в процессе
митоза, выполняется функция разделения генетического материала.
- В деконденсированном состоянии, в интерфазе, - функции репликации и
транскрипции.
При этом каждая хромосома в цикле остается цельной непрерывной нитью,
деконденсация происходит без разборки, без разрывов, а
пространственная организация хроматина обеспечена связями с ядерным
матриксом и ламиной ядерной оболочки.
77.
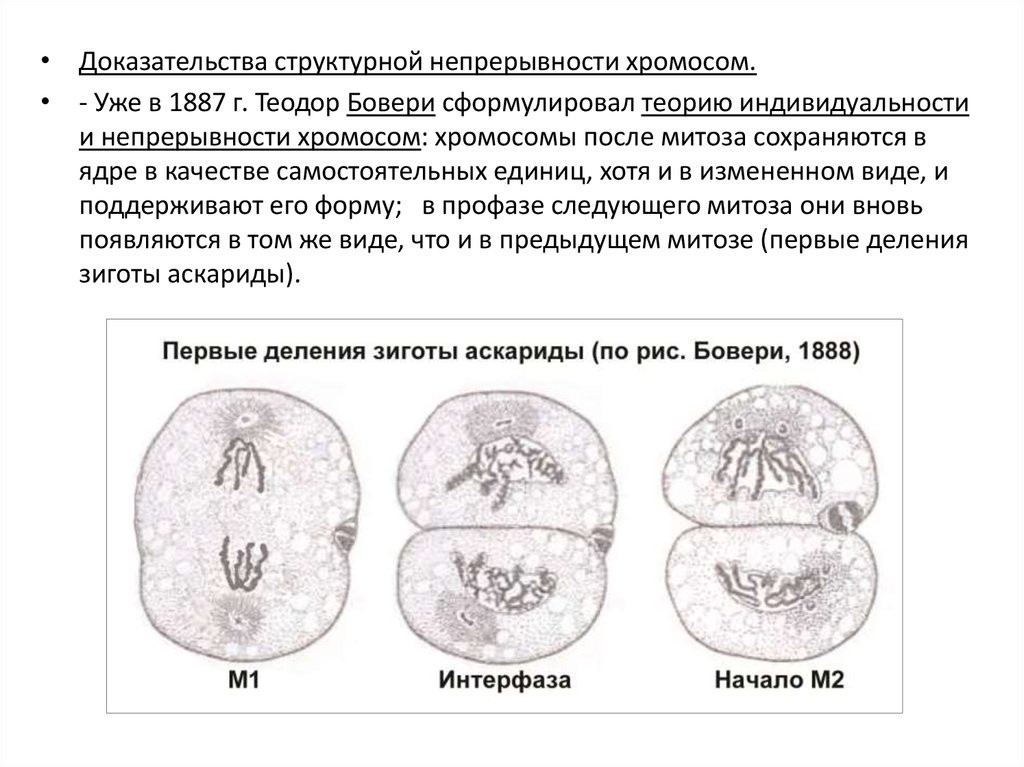
• Доказательства структурной непрерывности хромосом.• - Уже в 1887 г. Теодор Бовери сформулировал теорию индивидуальности
и непрерывности хромосом: хромосомы после митоза сохраняются в
ядре в качестве самостоятельных единиц, хотя и в измененном виде, и
поддерживают его форму; в профазе следующего митоза они вновь
появляются в том же виде, что и в предыдущем митозе (первые деления
зиготы аскариды).
78.
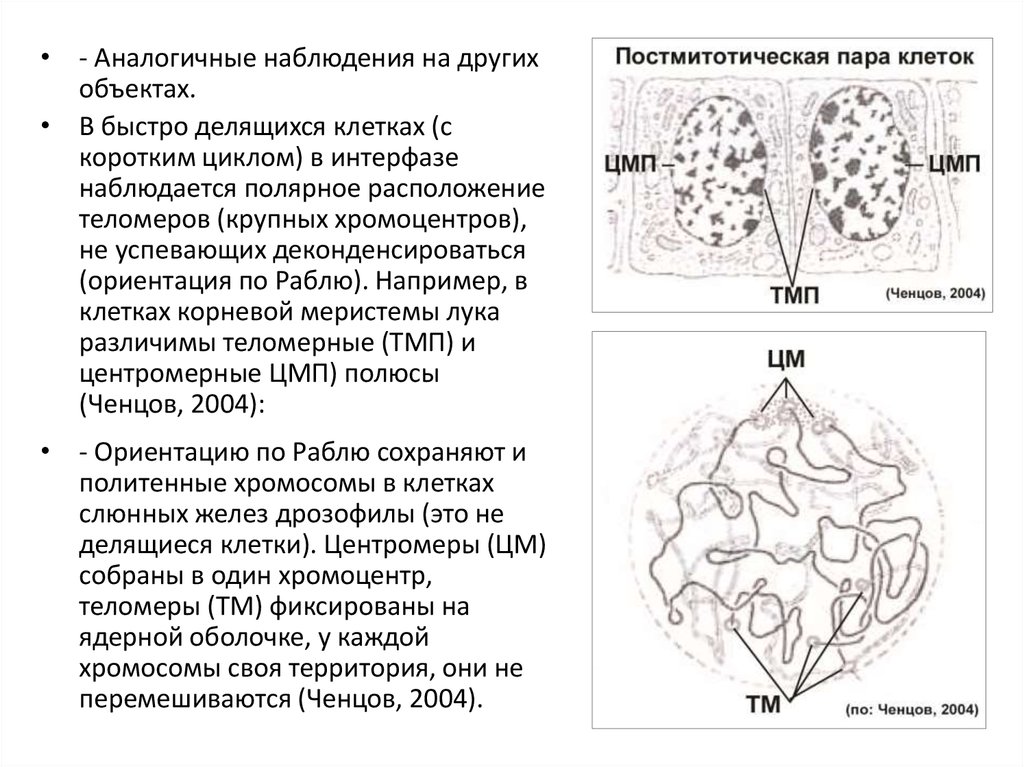
• - Аналогичные наблюдения на другихобъектах.
• В быстро делящихся клетках (с
коротким циклом) в интерфазе
наблюдается полярное расположение
теломеров (крупных хромоцентров),
не успевающих деконденсироваться
(ориентация по Раблю). Например, в
клетках корневой меристемы лука
различимы теломерные (ТМП) и
центромерные ЦМП) полюсы
(Ченцов, 2004):
• - Ориентацию по Раблю сохраняют и
политенные хромосомы в клетках
слюнных желез дрозофилы (это не
делящиеся клетки). Центромеры (ЦМ)
собраны в один хромоцентр,
теломеры (ТМ) фиксированы на
ядерной оболочке, у каждой
хромосомы своя территория, они не
перемешиваются (Ченцов, 2004).
79.
- Современные картины. Гибридизация in situ (на цитологическом препарате)между общим хроматином и ДНК отдельных хромосом выявляет в
интерфазных ядрах, уже далеких от митоза, индивидуальные хромосомные
территории (некоторые из них еще сохраняют ориентацию по Раблю). Наличие
индивидуальных хромосомных территорий свидетельствует об их целостности
и связи с ядерной оболочкой
(Podgornaya
et al., 2003)
80.
• - Постоянство хромосомной территории для гетерохроматической Ххромосомы в ядрах у особей женского пола млекопитающих (половойхроматин, тельце Барра).
• - Закономерное расположение определенных хромосом сохраняется и в
метафазной пластинке. Это означает, что в интерфазном ядре они как-то
фиксированы и сохраняют целостность, не рассыпаются на фрагменты.
• - Работы по доказательству полуконсервативного механизма репликации
ДНК. Во 2-м делении в хромосомах одна хроматида мечена, другая нет.
Значит, ДНК хроматиды в интерфазе не распадается, сохраняет
индивидуальную целостность.
• - По данным формальной генетики, в процессах кроссинговера,
скрещивания, при хромосомных разрывах функциональной единицей
выступает хроматида. Сцепленное наследование признаков (Морган) –
тоже следствие неизменного линейного расположения генов в
хроматиде.
• - У дрозофилы и др. объектов биохимическое определение
молекулярной массы ДНК (центрифугирование, электрофорез) дало
полное совпадение с расчетной молекулярной массой (исходя из
размера митотических хромосом). Т.о., хромосома – одна гигантская
молекула ДНК, ее длина не изменяется в клеточном цикле.
81.
• 5.2. Структурный цикл хромосомСтруктурные преобразования хромосом – основная характеристика митоза.
82.
Интерфаза. Хромосомы не видны, они деконденсированы до хроматина. Бывают
различимы крупные блоки гетерохроматина, обычно прицентромерного и
теломерного. Удвоение ДНК (хроматид) визуально определяется только по объему
и плотности ядра. Генетическая формула клетки: 2n2c → 2n4c.
Профаза. Появляются нитевидные хромосомы (плотный клубок, рыхлый клубок –
процесс компактизации). Они сначала длинные и тонкие. Иногда видно, что они
уже двойные – это сестринские хроматиды. Начинает разрушаться ядерная
оболочка. Генетическая формула клетки: 2n4c.
Прометафаза. Компактизация продолжается. Центромерные участки хромосом
под действием нитей веретена выталкиваются к экватору клетки. Ядерная
оболочка распалась. Генетическая формула клетки: 2n4c.
Метафаза. Максимальная компактизация хромосом. Все они бихроматидные.
Плечи разделены, но центромеры пока еще связаны. Т.к. все центромеры собраны
в экваторе (под действие веретена), хромосомы организованы в виде метафазной
пластинки. Генетическая формула клетки: 2n4c.
Анафаза. Сохраняется максимальная компактизация. Хроматиды разделяются и
под действием веретена расходятся к его полюсам. Генетическая формула клетки:
4n4c.
Телофаза. Формируются дочерние ядра. Начинается деконденсация хромосом,
они представляют плотный клубок. Происходит цитокинез. Клетки входят в новую
интерфазу. Генетическая формула клетки: 2n2c х 2.
83.
• У разных видов животных ирастений метафазная пластинка
выглядит по-разному, так как
различны параметры кариотипа:
число хромосом, их размеры и
форма.
Диплоидные (2n) хромосомные наборы:
1 – 22 хромосомы жабы Bufo arenarum;
2 – 140 хромосом ящерицыTupinambis
teguixin;
3 – 12 хромосом чернушки Nigella
orientalis (сем. лютиковые);
4 – 208 хромосом камчатского краба
Paralithodes camtschatica;
5 – 46 хромосом человека Homo sapiens;
6 – 23 хромосомы прямокрылого
насекомого Chromacris miles;
7 – 8 хромосом сложноцветного
растения Hypochoeris tweedie.
NB: Все рисунки выполнены при одном
увеличении (Мюнтцинг, 1967).
84.
• Каким же образом в митотической хромосоме длиной 4 мкмупаковано 4 см ДНК?
Как в ядре диаметром 10 мкм вмещается 1,8 метра ДНК?
Как упакована / распакована ДНК в составе интерфазного хроматина?
Насколько стабильна или лабильна эта упаковка?
Как она влияет на функциональную активность генов?
Чем и как контролируется?
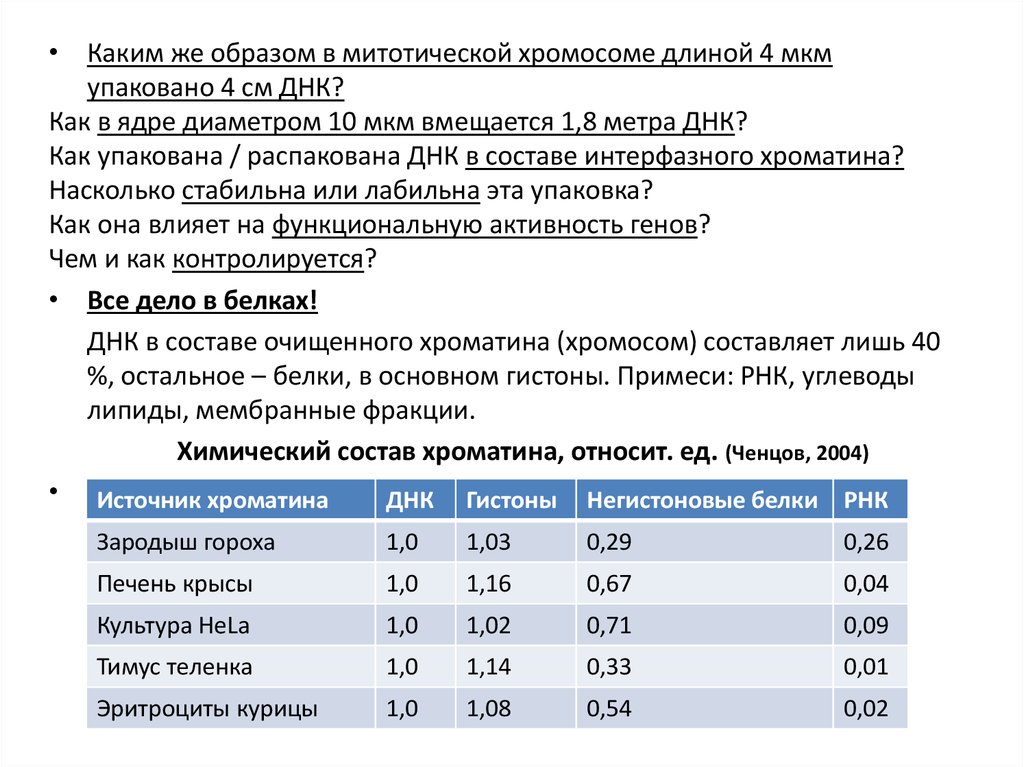
• Все дело в белках!
ДНК в составе очищенного хроматина (хромосом) составляет лишь 40
%, остальное – белки, в основном гистоны. Примеси: РНК, углеводы
липиды, мембранные фракции.
Химический состав хроматина, относит. ед. (Ченцов, 2004)
• Источник хроматина
ДНК Гистоны Негистоновые белки РНК
Зародыш гороха
1,0
1,03
0,29
0,26
Печень крысы
1,0
1,16
0,67
0,04
Культура HeLa
1,0
1,02
0,71
0,09
Тимус теленка
1,0
1,14
0,33
0,01
Эритроциты курицы
1,0
1,08
0,54
0,02
85.
• Гистоны отвечают за низшие уровни компактизации. Это оснóвные(щелочные) белки, богатые лизином и аргинином. Весьма
однообразны и консервативны. У эукариот всего 5-7 типов гистонов
(Н1, Н2А, Н2В, Н3, Н4, Н5...).
• Н3 и Н4 – аргининбогатые гистоны, самые консервативные (у коровы
и гороха различия всего по 2 аминокислотам).
• Н2А и Н2В – умеренно лизинбогатые. Имеется межвидовая
вариабельность.
• Н1 – большое семейство лизинбогатых гистонов, самые щелочные
(легко отделяются от хроматина в 0,5М солевых растворах). Наименее
консервативные, имеются межвидовые и даже межтканевые
вариации.
• Н5 – содержит много аргинина и серина. Замещает гистон Н1 в
эритроцитах низших позвоночных.
86.
• NB: С помощью положительных зарядов лизина и аргинина гистоныобразуют нековалентные (солевые, ионные, электростатические)
связи с фосфатными группами ДНК. (В растворах с высокой ионной
силой: 1-2М NaCl – все гистоны полностью отделяются от ДНК.)
Поэтому хроматиновый ДНП-комплекс имеет лабильную
организацию.
Для всех гистонов (особенно Н1)
характерно кластерное
распределение лизина и аргинина
на N- и С-концах молекулы. Именно
концы молекулы (щелочные,
положительно заряженные)
взаимодействуют с ДНК и
соседними гистонами. Средние
участки нейтральны и образуют 3-4
α-спиральных домена.
(Ченцов, 2004)
87.
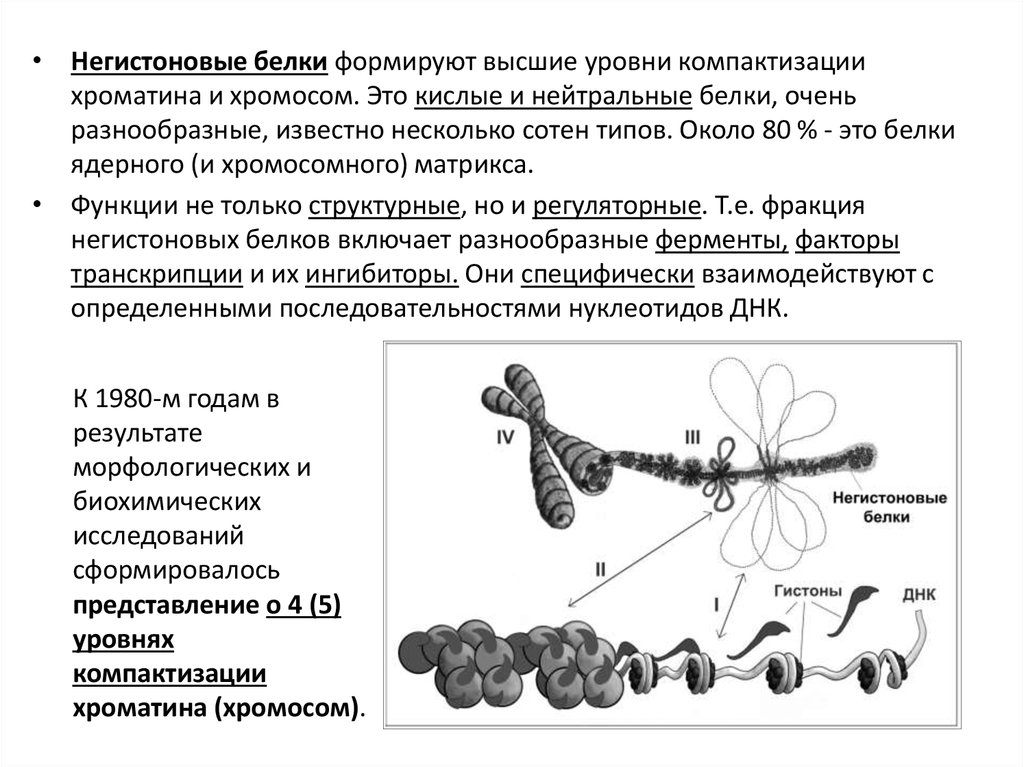
• Негистоновые белки формируют высшие уровни компактизациихроматина и хромосом. Это кислые и нейтральные белки, очень
разнообразные, известно несколько сотен типов. Около 80 % - это белки
ядерного (и хромосомного) матрикса.
• Функции не только структурные, но и регуляторные. Т.е. фракция
негистоновых белков включает разнообразные ферменты, факторы
транскрипции и их ингибиторы. Они специфически взаимодействуют с
определенными последовательностями нуклеотидов ДНК.
К 1980-м годам в
результате
морфологических и
биохимических
исследований
сформировалось
представление о 4 (5)
уровнях
компактизации
хроматина (хромосом).
88.
• (I) Нуклеосомная фибрилла диаметром 10 нм. (1нм = 0,001 мкм).Хроматин в щелочной среде с низкой ионной силой распадается на тонкие
фибриллы типа «бусы на нитке». «Бусы» – ДНК-гистоновые глобулы
диаметром 10-11 нм, скорость седиментации – 11S. «Нитка» – отрезки ДНК
(толщина 2нм) длиной около 20 нм (разрушается ДНКазой).
Микрококковая нуклеаза режет хроматин на отрезки, кратные 200 нп (200,
400, 600... нп) и 11S (11S, 22S, 33S…). Т.о., выявляются мономеры
(элементарные частицы хроматина), их димеры, тримеры ... Элементарные
частицы получили название нуклеосомы.
(Ченцов, 2004)
89.
• Максимальная нуклеосома (11S) включает:фрагмент ДНК около 200 нп, сердцевину (кор) из
8 гистонов – октамер (2хН2А, 2хН2В, 2хН3, 2хН4) и
одну молекулу гистона Н1. Вокруг кора 146 нп
образуют 1,75 оборота, остальные 54+ нп (8-114)
образуют свободный линкер.
Гистоны Н1 расположены поверх нуклеосом,
(Альбертс и др., 1994)
взаимодействуя с кором и линкером.
• Минимальная нуклеосома – то же без линкера и Н1.
• Коэффициент компактизации ДНК в нуклеосомной фибрилле = 6-7х.
NB: гистон Н1 для образования самих нуклеосом не требуется, но
участвует в стягивании соседних нуклеосом при формировании
следующего уровня компактизации.
Нуклеосомная структура хроматина устанавливается сразу в момент
репликации ДНК, так как синтез гистонов немного опережает синтез ДНК.
Старые и новые октамеры гистонов (коровые частицы) распределяются
между сестринскими цепями ДНК случайным образом. В репликативной
вилке, а также под РНК-полимеразой при синтезе РНК нуклеосомы,
вероятно, расщепляются в полунуклеосомы, не сползая с ДНК.
90.
(2) Основная хроматиновая фибрилла диаметром 30 нм.
Фибрилла 30 нм формируется под действием гистона Н1. При его
удалении фибрилла разворачивается в нуклеосомную (10нм) нить.
Существуют 2 модели упаковки 30 нм фибриллы: соленоидная и
нуклеомерная (см. рис.).
Нуклеомеры – сверхбусины, «супербиды». При обработке нуклеазой они
могут разделяться и образовывать 30 нм глобулы с константой
седиментации 45S. В ЭДТА и при удалении магния 30 нм глобулы
разворачиваются в нуклеосомные цепочки (по 6-8 нуклеосом).
Гистоны Н1 также организованы в кооперативные группы по 6-8 молекул.
Коэффициент компактизации ДНК в 30 нм фибрилле = 40-42х.
NB: Второй уровень упаковки не траскрибируется! Взаимодействие с
ферментами и многими регуляторными белками невозможно. Гены
блокированы.
(Ченцов, 2004)
91.
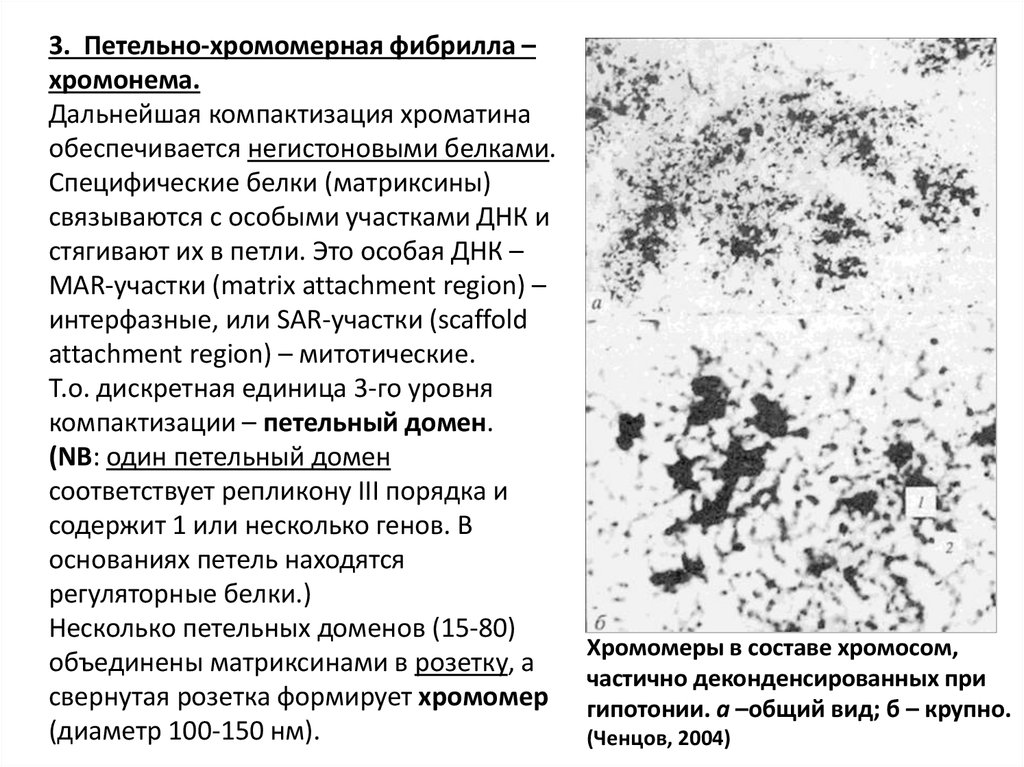
3. Петельно-хромомерная фибрилла –хромонема.
Дальнейшая компактизация хроматина
обеспечивается негистоновыми белками.
Специфические белки (матриксины)
связываются с особыми участками ДНК и
стягивают их в петли. Это особая ДНК –
MAR-участки (matrix attachment region) –
интерфазные, или SAR-участки (scaffold
attachment region) – митотические.
Т.о. дискретная единица 3-го уровня
компактизации – петельный домен.
(NB: один петельный домен
соответствует репликону III порядка и
содержит 1 или несколько генов. В
основаниях петель находятся
регуляторные белки.)
Несколько петельных доменов (15-80)
объединены матриксинами в розетку, а
свернутая розетка формирует хромомер
(диаметр 100-150 нм).
Хромомеры в составе хромосом,
частично деконденсированных при
гипотонии. а –общий вид; б – крупно.
(Ченцов, 2004)
92.
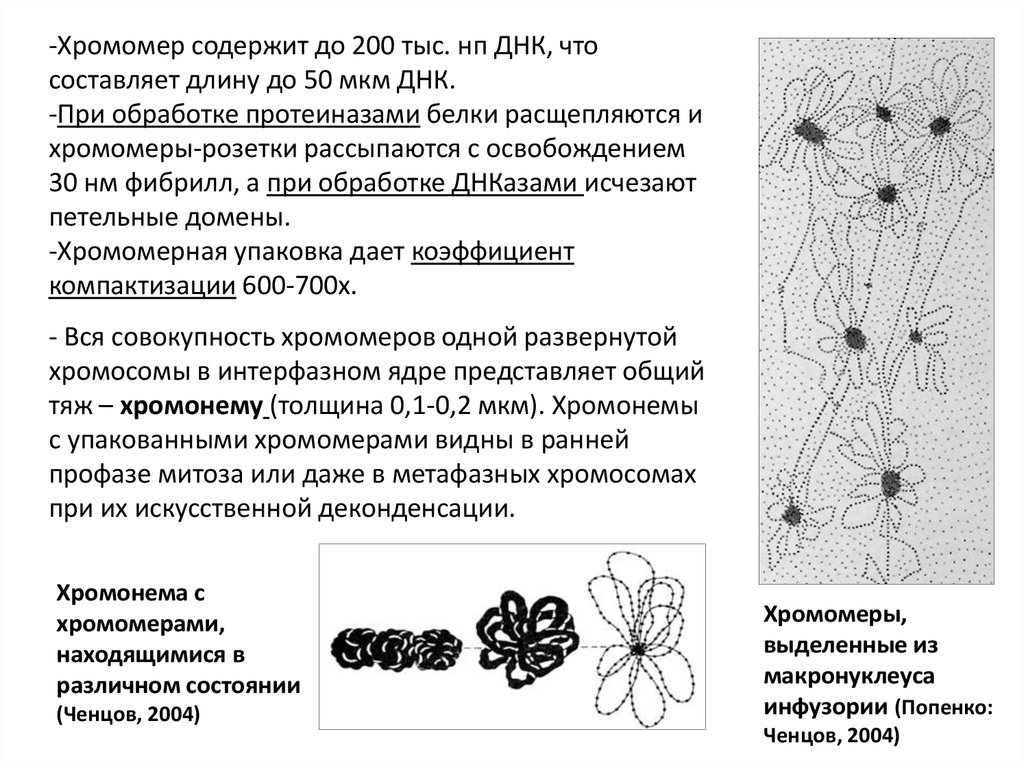
-Хромомер содержит до 200 тыс. нп ДНК, чтосоставляет длину до 50 мкм ДНК.
-При обработке протеиназами белки расщепляются и
хромомеры-розетки рассыпаются с освобождением
30 нм фибрилл, а при обработке ДНКазами исчезают
петельные домены.
-Хромомерная упаковка дает коэффициент
компактизации 600-700х.
- Вся совокупность хромомеров одной развернутой
хромосомы в интерфазном ядре представляет общий
тяж – хромонему (толщина 0,1-0,2 мкм). Хромонемы
с упакованными хромомерами видны в ранней
профазе митоза или даже в метафазных хромосомах
при их искусственной деконденсации.
Хромонема с
хромомерами,
находящимися в
различном состоянии
(Ченцов, 2004)
Хромомеры,
выделенные из
макронуклеуса
инфузории (Попенко:
Ченцов, 2004)
93.
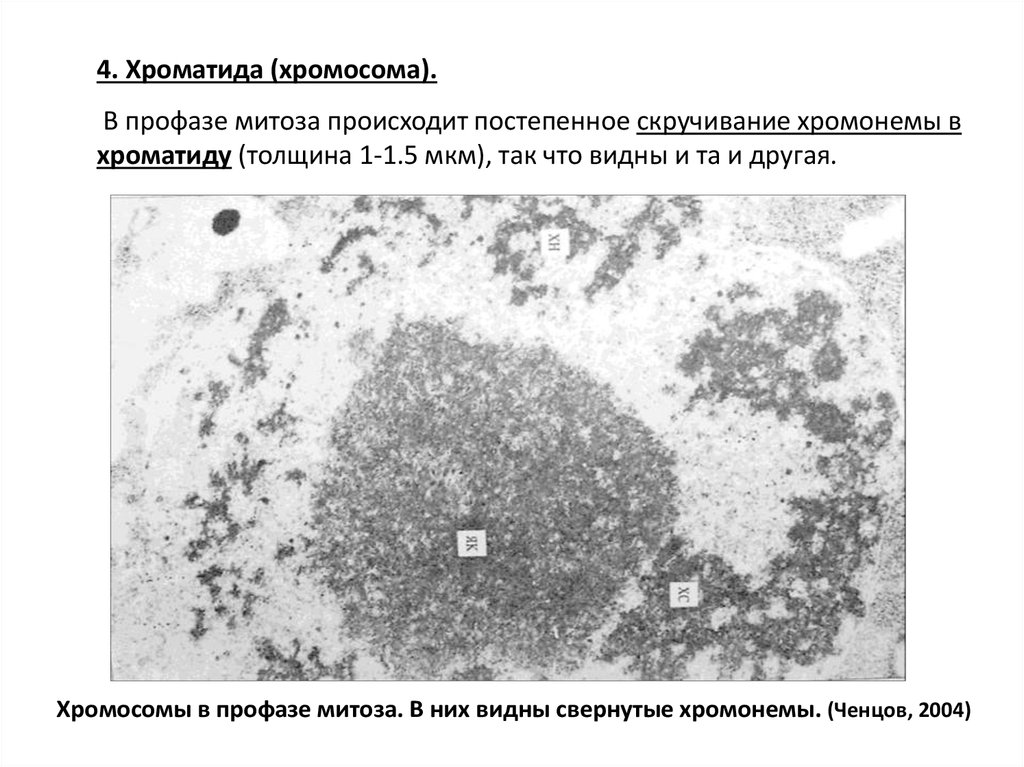
4. Хроматида (хромосома).В профазе митоза происходит постепенное скручивание хромонемы в
хроматиду (толщина 1-1.5 мкм), так что видны и та и другая.
Хромосомы в профазе митоза. В них видны свернутые хромонемы. (Ченцов, 2004)
94.
Максимальная компактизация хромосом достигается в метафазе.
Коэффициент = 10000х.
Метафазные
хромосомы на
срезе в
трансмиссионном
ЭМ (а, б)
и с поверхности в
сканирующем ЭМ
(в). (Ченцов, 2004)
В телофазе и раннем G1-периоде происходят обратные процессы –
раскручивание хроматид в хромонемы, развертывание хромомеров в
розетки, петли 30 нм фибрилл локально разворачиваются в 10 нм нити.
Становится возможной транскрипция генов и репликация.
95.
Модели организации хроматина и хромосомАльбертс и др., 1994
Ченцов, 2004, 2010
Анисимов, 2008, 2017
96. 6. Митоз. Цикл митотического веретена
Веретено обеспечивает расхождение хромосом в митозе. Это совокупностьмикротрубочек и моторных белков-транслокаторов. Цикл веретена в целом
согласован с циклом хромосом, что обеспечивает синхронность процессов.
6.1. Микротрубочки (25 нм) – элементы цитоскелета, состоят из белков
тубулинов и микротрубочко-ассоциированных протеинов (MAPs).
Локализация МТ различна : бывают одиночные МТ, рыхлые пучки, центриоли
и базальные тельца (3х9), аксонема ресничек и жгутиков (2х9 + 2). Во время
митоза большая часть МТ входит в состав митотического веретена.
Полимеризация МТ.
Субъединицы α- и β-тубулина (5 нм) образуют гетеродимеры,
потом кольца (нуклеация), потом трубочки в 13 продольных
протофиламентов. Полимеризация сопровождается
гидролизом ГТФ до ГДФ.
Димеры изначально поляризованы, поэтому и каждая МТ
оказывается полярной. Быстро растущий плюс-конец и
медленно растущий или разбираемый минус-конец (разная
скорость гидролиза ГТФ, до его полной остановки).
(по:Campbell, 1996)
97.
• Предполагаютсяразличные модели
полимеризации
тубулинов и
самосборки
микротрубочек
(по: Ченцов, 2004)
98.
• NB: Рост на +конце преобладает (быстрый гидролиз ГТФ) = удлинениеМТ. Но при недостатке тубулинов и ГТФ МТ могут разбираться с двух
концов = полная разборка. Разборка усиливается при понижении
температуры и в присутствии Са2+.
• Колхицин (алкалоид) связывается с тубулином и препятствует
полимеризации МТ = разборка цитоплазматических МТ (потеря формы
клеток) и МТ митотического веретена. То же с колцемидом и
нокодозолом, но после отмывки МТ восстанавливаются.
• Таксол, напротив, способствует полимеризации тубулина, даже при
низких концентрациях = избыточный рост МТ.
• MAP-белки ускоряют полимеризацию = стабилизируют МТ в
дифференцированных клетках.
• ВЫВОД: МТ являются очень динамичными структурами, способны
быстро возникать и разрушаться. Время полужизни МТ = около 5 мин!
За 15 мин. обновляется около 80% МТ.
• В дифференцированных клетках до 20 % МТ стабилизированы, «живут»
несколько часов (дополнительные MAPs, ацетилирование по лизину,
удаление терминального тирозина).
99.
• Функции микротрубочек:• - Скелетная (каркасная)
функция – стабилизация
формы клетки,
поддержание отростков
(нейроны, фибробласты,
эритроциты, радиолярии
и т.п.).
Клетка без МТ – шар.
(по: Ченцов, 2004)
• - Растяжение клеток, быстрый рост в длину, совместно с клеточной
стенкой (типично для растений).
• NB: Двигательная функция ! – внутриклеточный направленный
транспорт органелл и включений, экзоцитоз (мерокриновая секреция),
движение ресничек и жгутиков, работа митотического веретена.
100.
• Механизм движения органелл по микротрубочкам.1960-е годы. В аксонах нервных клеток открыт быстрый
транспорт веществ – намного более быстрый, чем общий ток
цитоплазмы. Скорость = 20-40 см/сут.
• Это оказалось перемещение вакуолей (везикул) и гранул по
микротрубочкам.
• Т.о., микротрубочки – не просто направляющие пути для
перемещения органелл. Это «рельсы», по которым движутся
«локомотивы», без них это движение невозможно.
• NB: Транспорт по микротрубочкам идет в двух противоположных
направлениях.
(Ченцов, 2004)
101.
• Антероградный транспорт – вперед, от тела нейрона к окончаниюаксона, т.е. к «+»-концам микротрубочек.
Моторный белок – кинезин с АТФазной активностью.
Связующий белок – кинектин – специфичен для разных «грузов»:
митохондрий, везикул Гольджи, лизосом, синаптических пузырьков,
антипараллельных МТ в митотическом веретене.
• Ретроградный транспорт – назад, от нервных окончаний к телу
нейрона, т.е. к «-»-концам микротрубочек.
Моторный белок – динеин, тоже АТФазно-активный.
(по: Ченцов, 2004)
Различают цитозольный
динеин – работает в
цитоплазме на
транспорт везикул и
расхождение хромосом
в митозе,
а также аксонемный
динеин – в киноцилиях
(ресничках и жгутиках).
102.
• NB: В клетке одновременно сочетаются различные направлениятранспорта по МТ – на периферию и к центру.
• Это же самое мы увидим в работе митотического веретена.
(по: Ченцов, 2004)
• (NB: имеется принципиальное сходство тубулин-кинезиновой и
тубулин-динеиновой двигательных систем с актин-миозиновой
мышечной системой: скелетные элементы + двигательные АТФазные
белки. Возможна их гомология.)
103.
6.2. Центры организациимикротрубочек - ЦОМТы
Рост МТ полярный, нуклеация и
полимеризация идут из центров
организации микротрубочек – ЦОМТов.
На ЦОМТах фиксированы минус-концы,
блокированные с помощью МАРs,
к периферии обращены плюс-концы.
(по: Ченцов, 2004)
Происходит постоянная закладка новых МТ у ЦОМТов и их рост к
периферии. (Доказано в опытах с разрушением МТ колцемидом или
холодом с последующим их восстановлением из центра (Ченцов и сотр.)
NB: Отрастающие МТ могут терять связь с ЦОМТами, после чего
перемещаются (тредмиллинг), стабилизируются или разбираются.
104.
• Что такое ЦОМТ?Старые работы:
в клетках имеется клеточный
центр (как органоид, а не
геометрический центр клетки),
состоящий из плотного тельца
центросомы, светлого поля –
центросферы и астральной
лучистости – астросферы.
105.
• Современная ультраструктура:• центросома у животных клеток это –
2 центриоли (диплосома),
центросфера – зона расположения
центриолярных сателлитов, глобул
(ФСМТ) и матрикса, астросфера –
микротрубочки.
• Именно сателлиты и прилежащие
глобулы (ФСМТ), а не центриоли
являются истинными ЦОМТами.
(Ченцов, 2004)
По: Ченцов, 2004)
106.
• NB: Пара центриолей – материнская и дочерняя. Воспроизводятся вклеточном цикле по консервативному принципу: отрастанием сбоку.
Механизм не известен. Центриоли имеют разный возраст.
Сателлиты и ФСМТ имеются только на материнской центриоли.
• NB: У высших растений, низших грибов, некоторых простейших
ЦОМТы не имеют четкой локализации, так как в них нет центриолей.
Диффузный ЦОМТ-материал локализован на периферии клеточного
ядра (снаружи или внутри), откуда МТ расходятся радиально.
Это подтверждает, что истинные ЦОМТы – не центриоли, а
прилежащий материал.
Сами центриоли – лишь «пассажиры», расходящиеся с гарантией в
дочерние клетки во время митоза (аналогия Ю.С. Ченцова).
• А функция центриолей – это потенциальные базальные тельца
киноцилий (ресничек и жгутиков).
107.
• 6.3. Центросомный цикл, сборка и работа митотического веретена• Центросомы (центриоли) несут ЦОМТы, а ЦОМТы организуют
митотическое веретено и расхождение хроматид в митозе.
Т.о., существует центросомный цикл.
В центросомном цикле происходит 2 процесса:
- расхождение и удвоение
центриолей,
- перестройка ЦОМТов и
микротрубочек.
В G1-периоде (после
митоза) – 1 диплосома.
Веретено разрушено.
Материнская центриоль
имеет сателлиты.
Начинается рост
микротрубочек, их отрыв и
движение на периферию.
(по: Ченцов, 2004)
108.
• В S-периоде происходит дупликация центриолей.У материнской и дочерней центриолей сбоку отрастают процентриоли
(дочерняя и внучатая). Механизм неизвестен.
У тетрахимены находили цДНК, но интерпретации нет.
В G2-периоде клетка имеет 2
клеточных центра – 2 полюса
митотического веретена.
Сателлиты каждой
материнской центриоли
перестраиваются в рыхлую
волокнистую муфту –
фибриллярное гало, от
которого начинают отрастать
МТ веретена. При этом
старые цитоплазматические
МТ разрушаются (клетка
округляется), а пул тубулинов
используется на построение
новых МТ веретена.
109.
• В митозе у материнских центриолей сохраняется фибриллярное гало(вместо сателлитов).
От гало центриолей обоих полюсов исходит поток обновляемых МТ
(показано в опытах).
В телофазе веретено
разбирается. Гало исчезает.
Восстанавливаются сателлиты.
NB: Такая динамика
характерна для тканевых и
культуральных клеточных
линий с умеренной скоростью
пролиферации.
В эмбриональных клеточных
циклах, с коротким циклом,
удвоение центриолей
происходит много раньше –
еще в предыдущем цикле.
110.
• Митотическое веретено начинаетформироваться уже в G2 периоде,
удлиняется в профазе и приобретает
окончательные формы в метафазе.
При этом часть растущих МТ своими «+»концами захватываются кинетохорами
хромосом, содержащими специальный
белок CENT-A, а также динеин.
Профаза
Ченцов, 2004)
Прометафазные хромосомы, окрашенные DAPI
на ДНК и антителами к CENT-A (Ченцов, 2004: фото
И.С. Кудрявцева).
Формирование кинетохора в
течение профазы – метафазы.
Показана только 1 хроматида.
111.
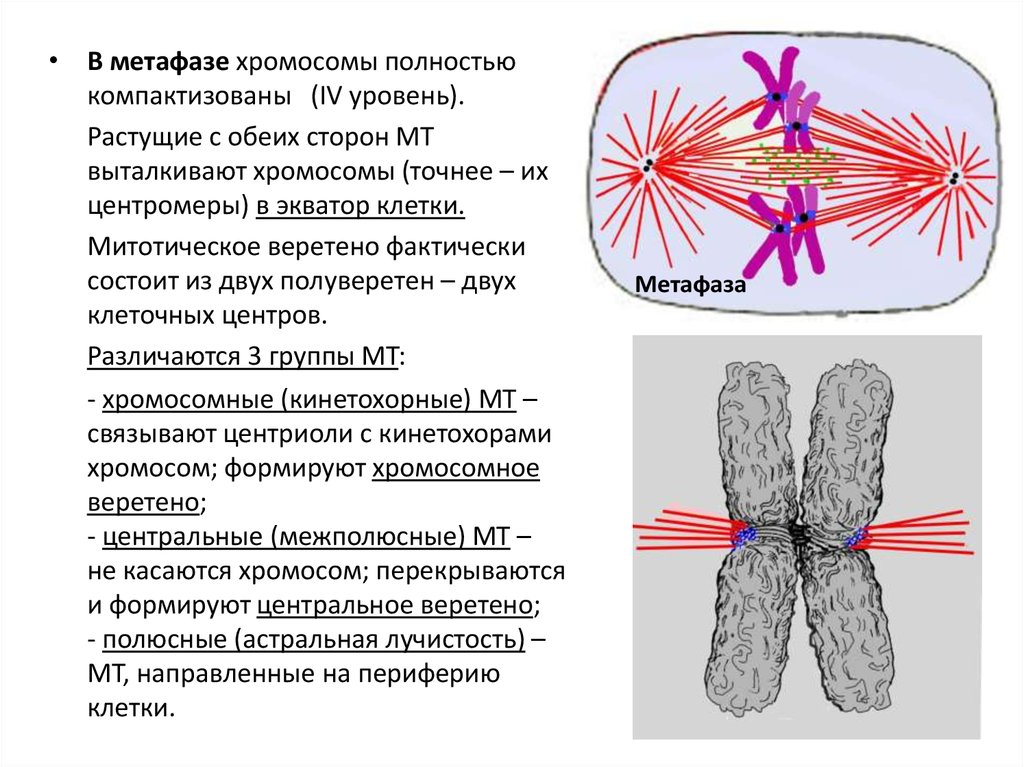
• В метафазе хромосомы полностьюкомпактизованы (IV уровень).
Растущие с обеих сторон МТ
выталкивают хромосомы (точнее – их
центромеры) в экватор клетки.
Митотическое веретено фактически
состоит из двух полуверетен – двух
клеточных центров.
Различаются 3 группы МТ:
- хромосомные (кинетохорные) МТ –
связывают центриоли с кинетохорами
хромосом; формируют хромосомное
веретено;
- центральные (межполюсные) МТ –
не касаются хромосом; перекрываются
и формируют центральное веретено;
- полюсные (астральная лучистость) –
МТ, направленные на периферию
клетки.
Метафаза
112.
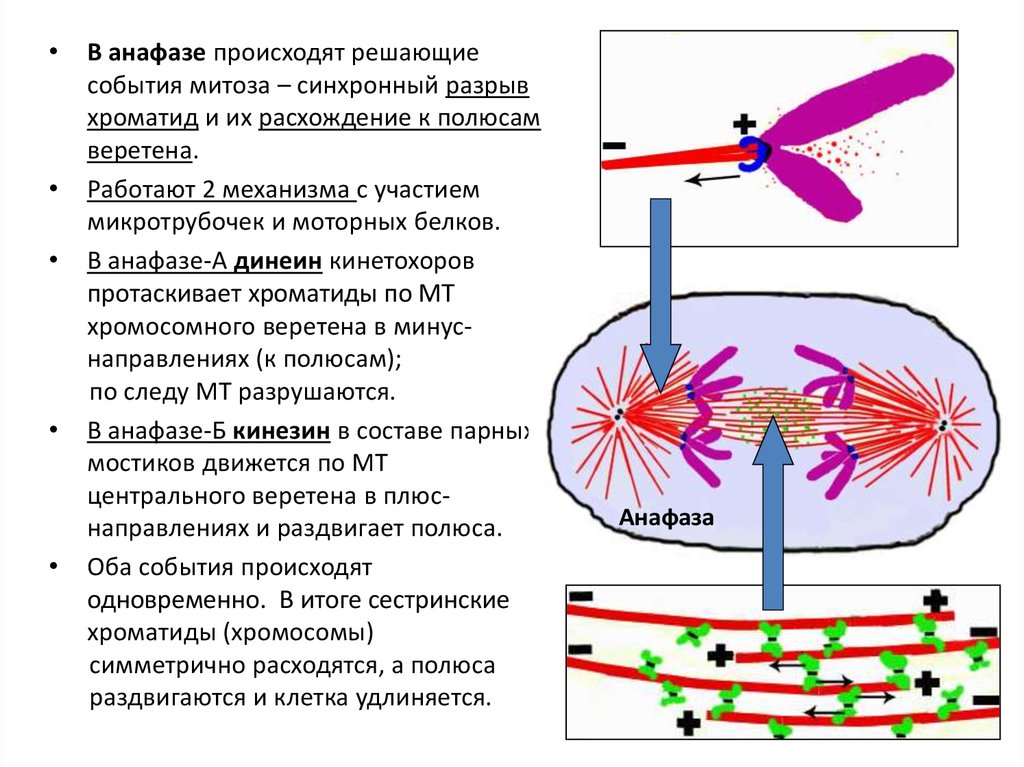
В анафазе происходят решающие
события митоза – синхронный разрыв
хроматид и их расхождение к полюсам
веретена.
Работают 2 механизма с участием
микротрубочек и моторных белков.
В анафазе-А динеин кинетохоров
протаскивает хроматиды по МТ
хромосомного веретена в минуснаправлениях (к полюсам);
по следу МТ разрушаются.
В анафазе-Б кинезин в составе парных
мостиков движется по МТ
центрального веретена в плюснаправлениях и раздвигает полюса.
Оба события происходят
одновременно. В итоге сестринские
хроматиды (хромосомы)
симметрично расходятся, а полюса
раздвигаются и клетка удлиняется.
Анафаза
113.
В телофазе завершается кариокинез (деление ядерных структур –
хромосом) и осуществляется цитокинез (цитотомия) – деление самой
клетки.
- Разошедшиеся хроматиды
собираются у полюсов и формируют
новые ядра (2n2c);
- хроматиды деконденсируются и
превращаются в хроматин, работа
генов (транскрипция) возобновляется;
- в том числе восстанавливаются
ядрышки – начинается производства
Телофаза
рРНК и сборка рибосом;
- восстанавливаются ядерные
оболочки путем сборки везикул ЭПР.
Микротрубочки веретена, кроме
астральных, разрушаются. Каждая
клетка получает один клеточный центр
– пару центриолей в окружении МТ.
Параллельно идет цитокинез.
114.
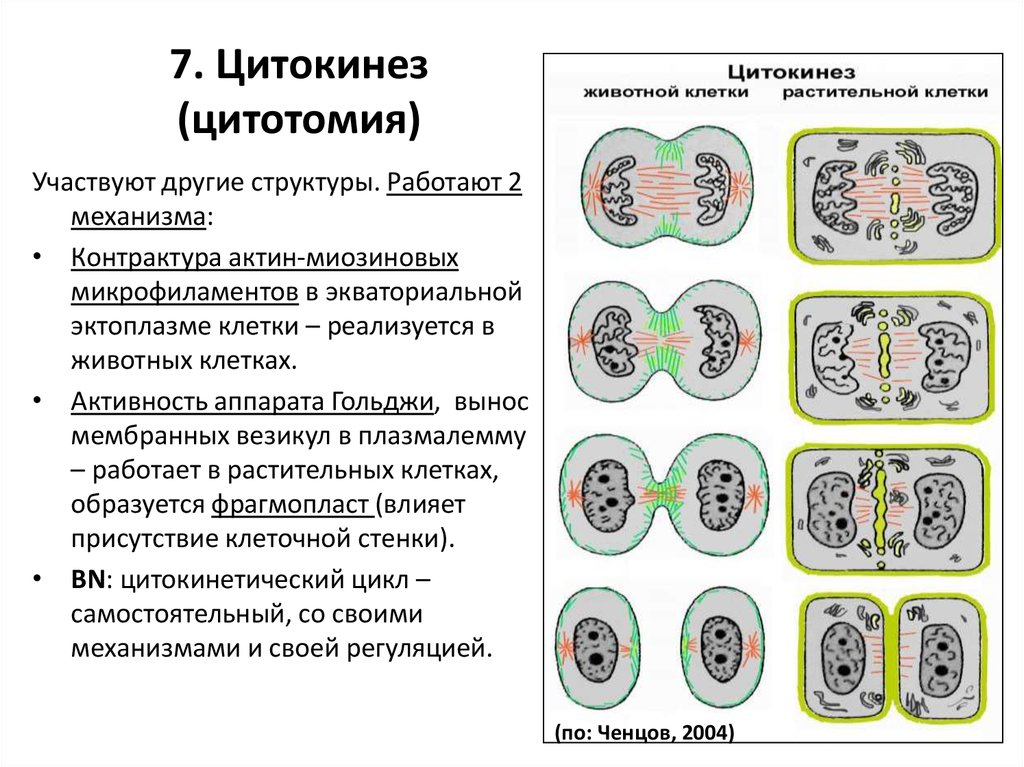
7. Цитокинез(цитотомия)
Участвуют другие структуры. Работают 2
механизма:
• Контрактура актин-миозиновых
микрофиламентов в экваториальной
эктоплазме клетки – реализуется в
животных клетках.
• Активность аппарата Гольджи, вынос
мембранных везикул в плазмалемму
– работает в растительных клетках,
образуется фрагмопласт (влияет
присутствие клеточной стенки).
• BN: цитокинетический цикл –
самостоятельный, со своими
механизмами и своей регуляцией.
(по: Ченцов, 2004)
115.
ПРИЛОЖЕНИЕМитотический цикл в клеточной культуре
(Ченцов, 2004; фото А.В. Буракова и А.Л. Ходякова)
G1-период; в клеточном центре пара
S-G2; расхождение дочерней
центриолей – диплосома.
и материнской центриолей.
Микротрубочки окрашены антителами к тубулину. Центриоли желтые.
116.
МетафазаАнафаза
Микротрубочки веретена и клеточные центры – желтые; хромосомы –голубые;
промежуточные филаменты цитоскелета – красные.
117.
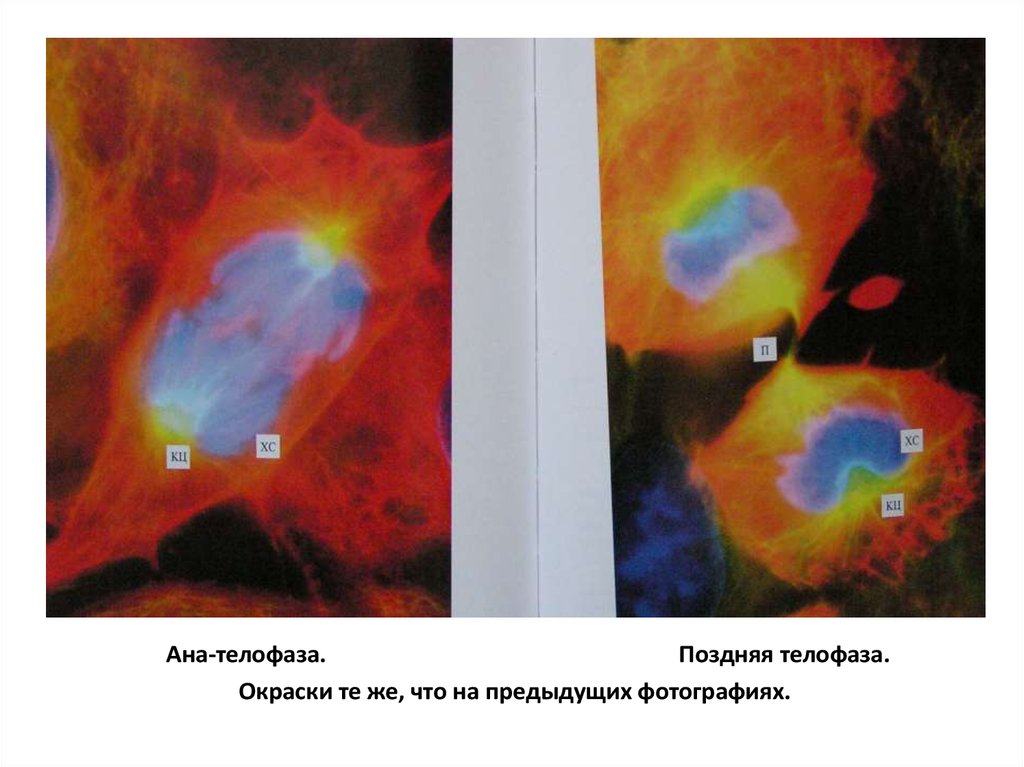
Ана-телофаза.Поздняя телофаза.
Окраски те же, что на предыдущих фотографиях.
118.
• Т.о., митотический цикл – сочетание трех относительнонезависимых циклов:
- хромосомный цикл;
- центросомный цикл;
- цитокинетический цикл.
• Каждый цикл имеет свою регуляцию. В норме эти циклы
согласованы, но возможны рассогласования, приводящие
к нарушениям митоза с различными последствиями.