

 – липофильные молекулы с Red-Ox свойствами: перенос 2ē + 2Н+")
 и флавинмононуклеотид (ФМН, FMN) – компоненты многих Red-Ox ферментов: перенос 2ē + 2Н+")










































")



")
: спорофитный и гаметофитный контроль")
: спорофитный и гаметофитный контроль")



 Биология
БиологияПохожие презентации:


 наследование")







Электрон - транспортная цепь митохондрий
1.
2.
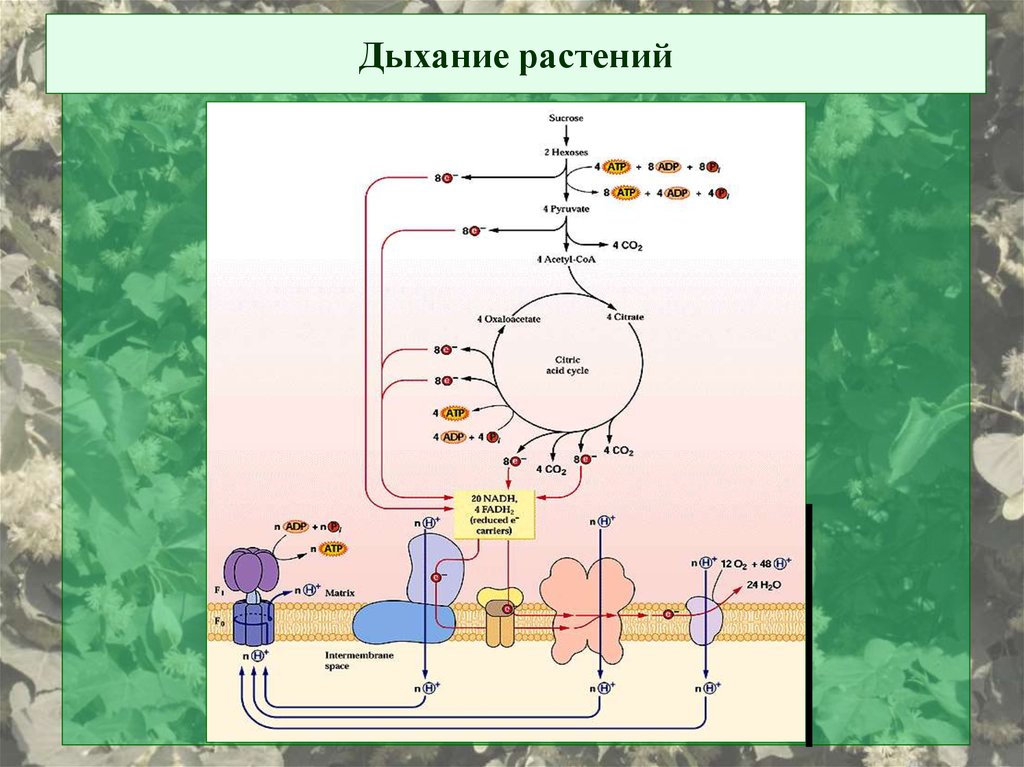
Дыхание растений3.
Общая схема дыхательной ЭТЦ: четыре белковых комплекса,объединяемые подвижными переносчиками е-
Переносчики е- в ЭТЦ
выстаиваются
согласно своим
Red/Ox потенциалам
– «под горку»
НАДН → ФП (ФМН) → FeS (min 5шт) → UQ(пул) →
-0,32v
-(0,3 – 0,1)v
-(0,37-0,02)v
0v
→ цит b → FeS → цит с1 → цит с → цит а-а3 → О2
0.1v
0,28v 0,22v
0,25v 0,29-0,55v 0,82v
4. Хиноны (убихиноны и пластохиноны) – липофильные молекулы с Red-Ox свойствами: перенос 2ē + 2Н+
E0’: от 0,0 до +0,10 V(в связанном виде – до -0,3 V)
в среднем – около 0 V
5. Флавинадениндинуклеотид (ФАД, FAD) и флавинмононуклеотид (ФМН, FMN) – компоненты многих Red-Ox ферментов: перенос 2ē + 2Н+
Eo : от -0,5 до +0,2 Vв зависимости от белка,
стабилизации кольца, резонанса
6. Железо-серные белки - 2Fe-2S и 4Fe-4S: перенос только одного ē
Eо’:от – 0,42V (Fd)
до + 0,35V
(Fe-S бел. Риске)
7. Гемы – коферменты цитохромов: перенос только одного ē
Cyt bCyt с
Cyt а
Eо’:
от -0,18 V (cyt b6)
до +0,55 V (cyt a3)
8.
Общая схема дыхательной ЭТЦ: четыре белковых комплекса,объединяемые подвижными переносчиками е-
Подвижные переносчики:
1. Убихиноны – их много («пул убихинонов»)
2. Цитохром с – 104 ак. Локализован с наружной стороны внутренней
мембраны. Гем ковалентно связан с белком (через cys14 и cys17)
Примерная стехиометрия:
1 комплекс I : 3 bc1 : 7 aa3 : 9 cyt c : 50 UQ
9.
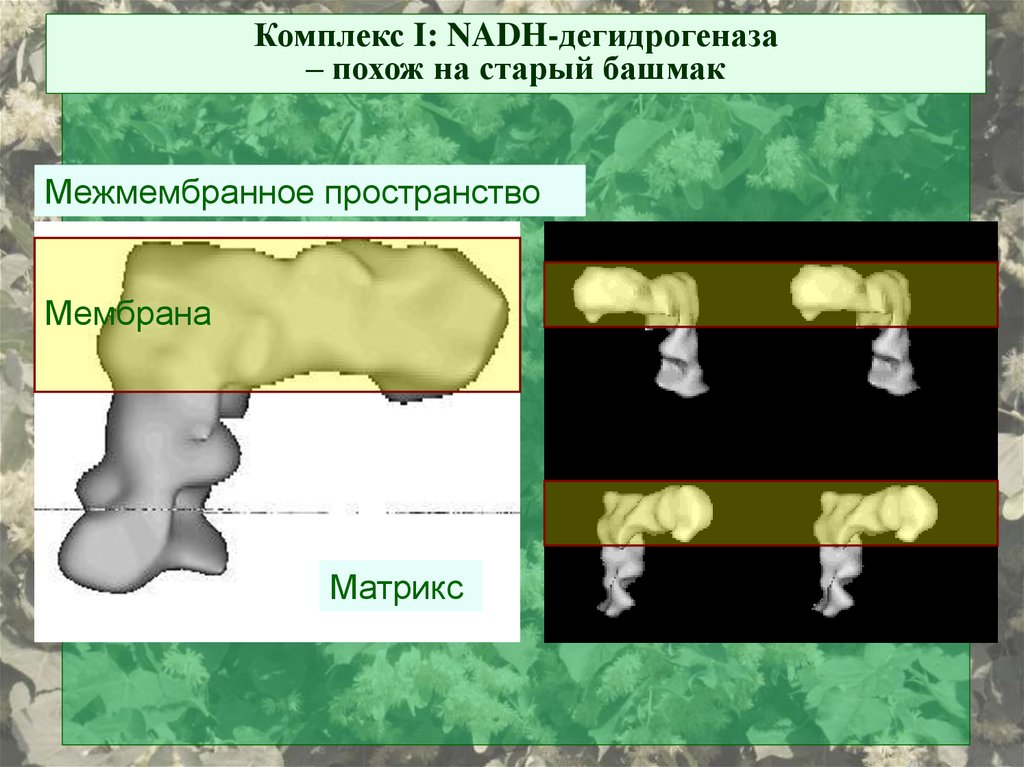
Комплекс I: NADH-дегидрогеназа– похож на старый башмак
Межмембранное пространство
Мембрана
Матрикс
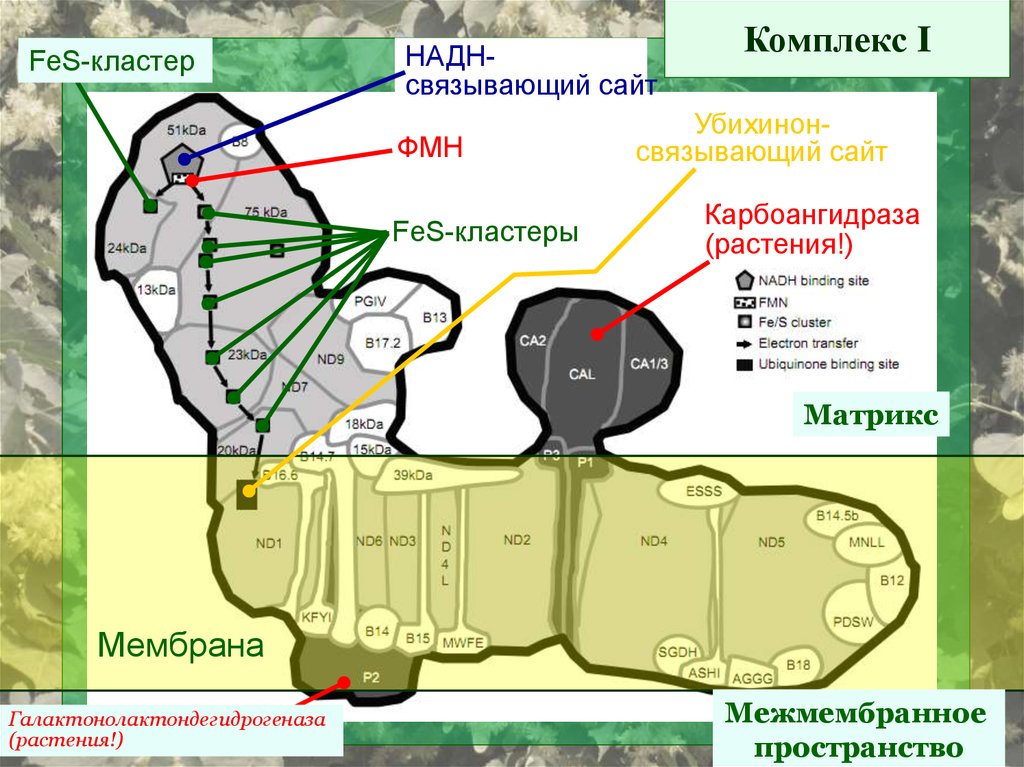
10.
FeS-кластерНАДНсвязывающий сайт
ФМН
FeS-кластеры
Комплекс I
Убихинонсвязывающий сайт
Карбоангидраза
(растения!)
Матрикс
Мембрана
Галактонолактондегидрогеназа
(растения!)
Межмембранное
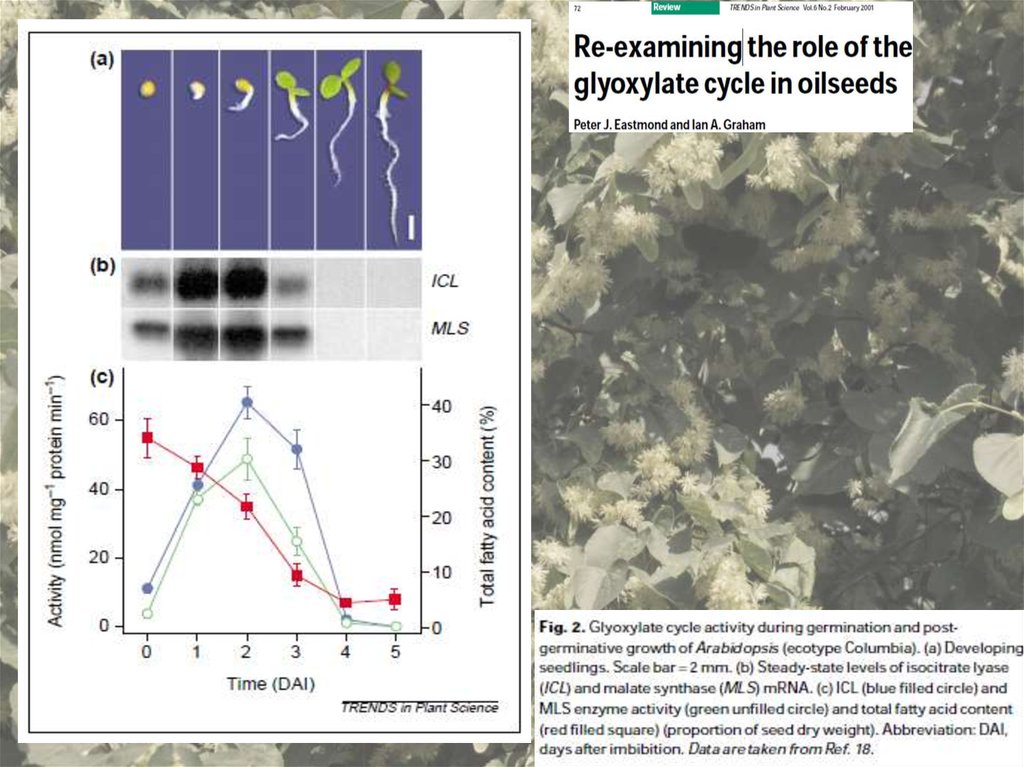
пространство
11.
Комплекс I: NADH-дегидрогеназа12.
Особенность ЭТЦ дыхания растений – дополнительныеNAD(P)H-дегидрогеназы
13.
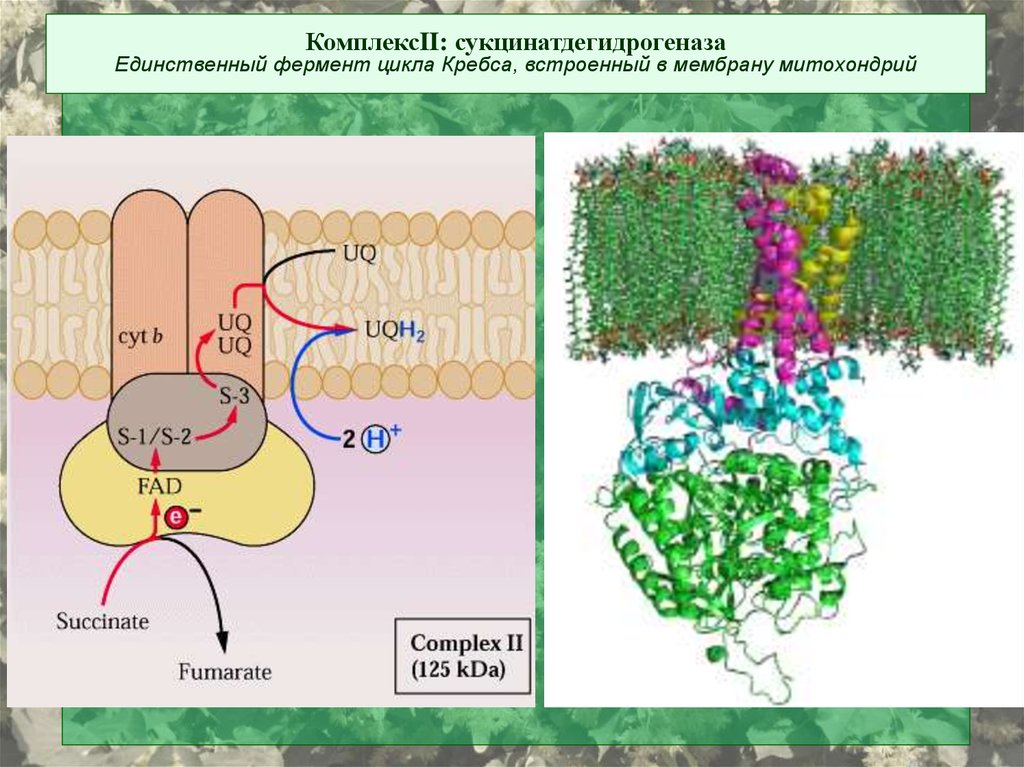
КомплексII: сукцинатдегидрогеназаЕдинственный фермент цикла Кребса, встроенный в мембрану митохондрий
14.
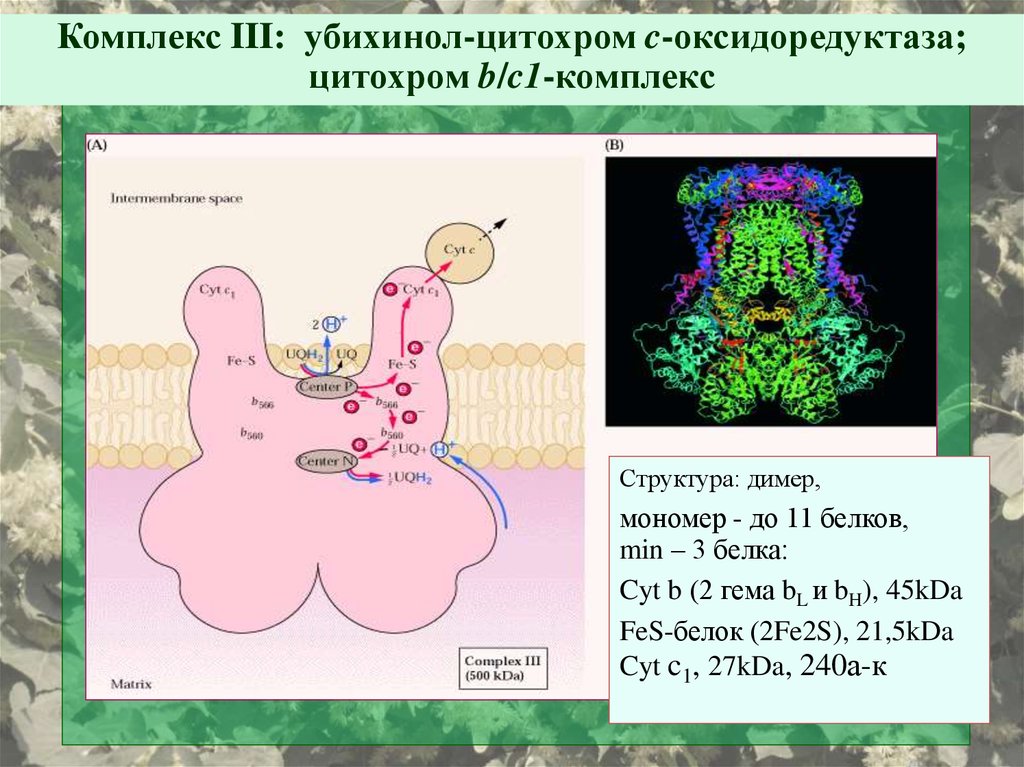
Комплекс III: убихинол-цитохром c-оксидоредуктаза;цитохром b/c1-комплекс
Структура: димер,
мономер - до 11 белков,
min – 3 белка:
Cyt b (2 гема bL и bH), 45kDa
FeS-белок (2Fe2S), 21,5kDa
Cyt c1, 27kDa, 240а-к
15.
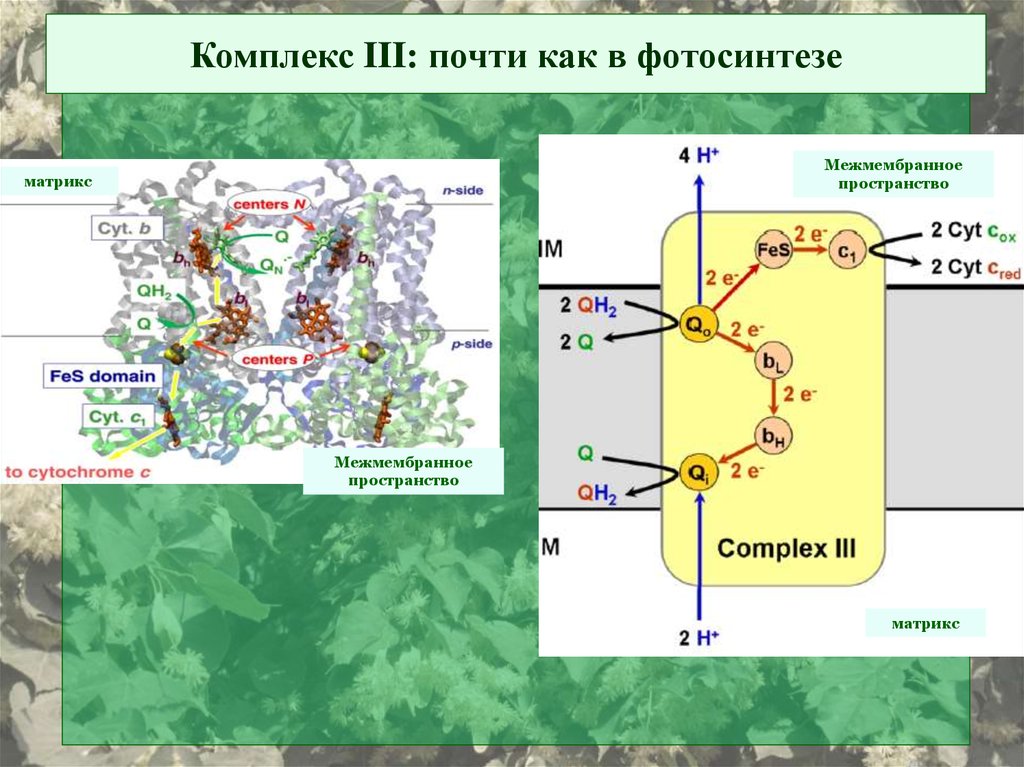
Комплекс III: почти как в фотосинтезеМежмембранное
пространство
матрикс
Межмембранное
пространство
матрикс
16.
Цитохром с: белок, ассоциированный с мембранойL-Аскорбат
D-Лактат
Пируват
L-Галактонолактон
D-L DH
GL DH
12 кДа
Пул
UQ
cyt c
III
Комплекс I
17.
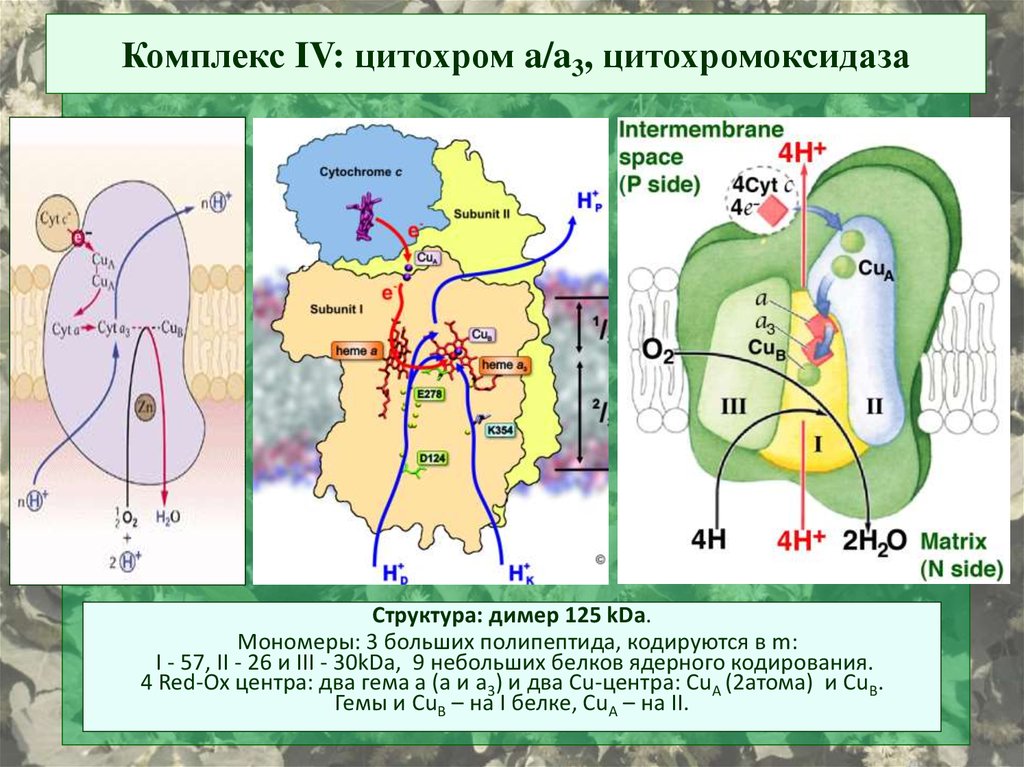
Комплекс IV: цитохром a/a3, цитохромоксидазаСтруктура: димер 125 kDa.
Мономеры: 3 больших полипептида, кодируются в m:
I - 57, II - 26 и III - 30kDa, 9 небольших белков ядерного кодирования.
4 Red-Ox центра: два гема а (a и a3) и два Cu-центра: СuА (2атома) и CuВ.
Гемы и CuB – на I белке, CuA – на II.
18.
Комплекс IV: цитохром a/a3, цитохромоксидазамежмембранное пространство
цит с
H+
ē
H+
CuA
СuA
гем а
II
гем а3 -СuB
I
O2
H+
H+
H+
D
K H
2H2O
«двуядерный» центр гем а3–СuВ,
19. Предполагаемая схема работы цитохрома а-а3
AFe2+ O Cu+
О
P
ē
2H+
D-канал
Fe2+–O Cu+
О
O2
Fe2+
R
Cu+
ē
Fe2+ =O Cu+
Fe3+
Cu+
O
F
H
H
H
O
ē
2H+
Fe3+
Cu2+
ē
2H2O
К-канал
D-канал
20.
АТФ-синтаза: вальсирующий комплексСтруктура:
Две субъединицы: F0 и F1
F1: α3β3γδε
α - 59, β - 56, γ - 36
δ - 17.5, ε - 13,5kDa
F0: а (I), 15kDa,
b (II), 12.5kDa
c (III), 8kDa
а:b:с – 1:2:(6-15)
α , β, ε субъединицы F1
и субъединица а (I) F0
кодируются в
митохондриальном
геноме.
21.
22.
Альтернативная оксидаза2QH2 +O2 → 2Q + 2H2O
Термогенез ароидных
растений
Arum maculatum,
Symplocarpus foetidus,
Sauromatum guttatum,
Philodendron selloum.
Альтернативная оксидаза
Один полипептид
Один ядерный ген АОХ1
В мембране Мх:
мономерная
или димерная форма
Впервые белок АО
был получен из мтх
Sauromatum guttatum
23. Sauromatum guttatum
24.
Symplocarpus foetidus25.
Helicodeceros muscivorus26.
Особенность ЭТЦ дыхания растений –альтернативная оксидаза
Гомодимер, 37 kDa, ~ 350 а-к. Передает ē от убихинона на кислород.
Устойчива к цианиду!
Есть изозимы (минимум 2): конститутивная и индуцибельная АО
Две формы: окисленная и восстановленная. Окисленная неактивна.
27.
Альтернативная оксидаза28.
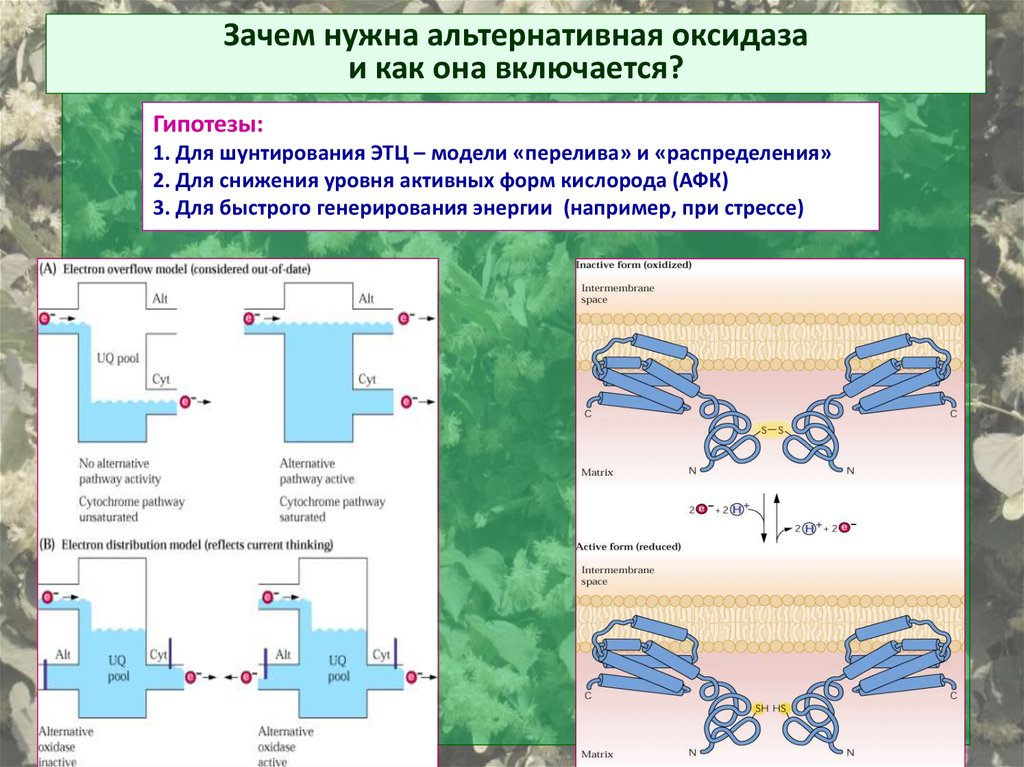
Зачем нужна альтернативная оксидазаи как она включается?
Гипотезы:
1. Для шунтирования ЭТЦ – модели «перелива» и «распределения»
2. Для снижения уровня активных форм кислорода (АФК)
3. Для быстрого генерирования энергии (например, при стрессе)
29.
Регуляция экспрессии и активности альтернативнойоксидазы
30. Лирическое отступление
31. Откуда берется цианид?
Цианогенные гликозидыC≡N
глюкоза – О – C – СН3
R
R – CH3
(ЛИНАМАРИН) Trifolium sp.,
Maniholt sp.
R – фенил (ПРУНАЗИН) Prunus sp (Rosaceae)
Vicia sp., Sorghum sp.,
Linum sp.
32. Цианогенные гликозиды
33.
• Цианогенныегликозиды
продуцируют
2500-3000 видов
растений
сем. бобовых,
розоцветных,
мимозовых
• Клевер, кассава,
лен, сорго, лотос,
гевея,
пассифлора,
ячмень, абрикос,
акации
34.
Цианогенные гликозидыСинильная кислота – одно из самых
быстродействующих веществ. Нарушает
клеточное дыхание. Симптомы: жжение
в горле, шум в ушах, чувство страха,
тошнота, рвота.
Padus racemosa
Padus serotina
Черемуха обыкновенная
Черемуха виргинская
Amygdalus nana
Миндаль низкий (бобовник)
Trifolium repens
Клевер ползучий
Lupinus polyphyllus
Syringa pannonica
Люпин многолистный Сирень венгерская
35.
36.
Родина: Бразилия(культивируется
6–7 тыс. лет)
Синонимы:
Маниок, тапиока,
кассава, «юкка»
Корнеклубни:
Длина – до 1 м
Вес – до 15 кг
Урожайность – в среднем 120 ц/га
Рекорд – 348 ц/га
37.
38.
HCNцианид,
синильная кислота
Линамарин: 100–500 мг/кг
39.
C≡Nглюкоза – О – C – СН3
линамараза
R
H2O
глюкоза
CN
НО – C – СН3
СН3
ЦИАНОГИДРИН АЦЕТОНА
О
лиаза
C – СН3 + НCN
СН3
ацетон
40.
41.
42.
43.
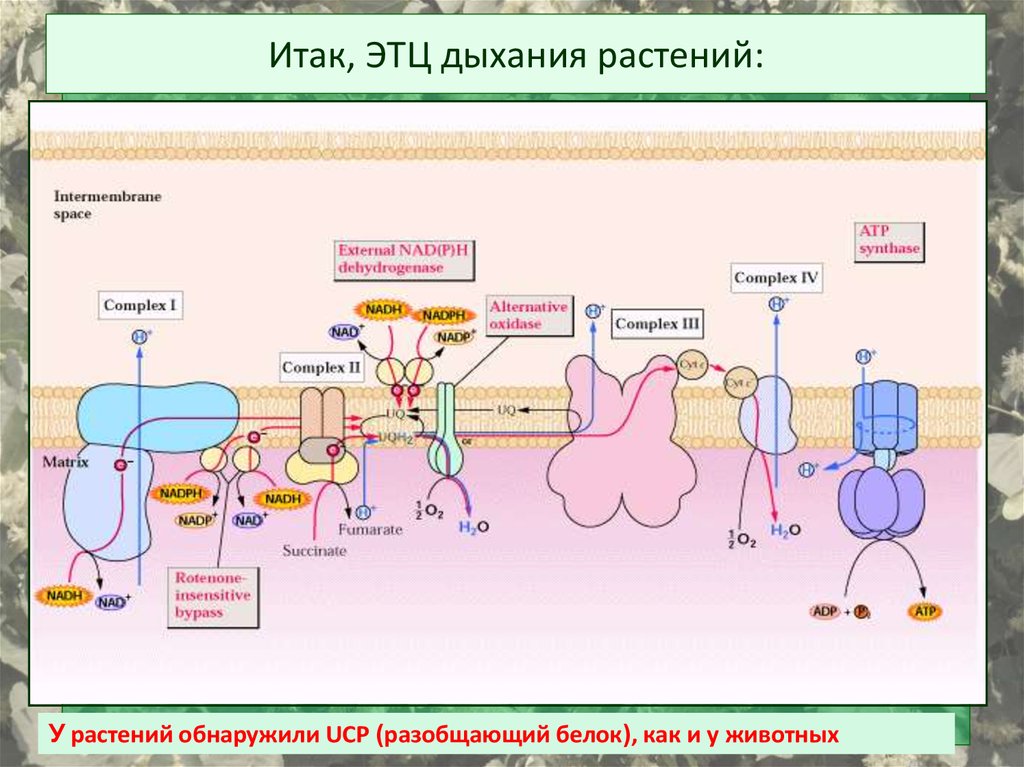
Итак, ЭТЦ дыхания растений:У растений обнаружили UCP (разобщающий белок), как и у животных
44.
ēН2О
ФC II
Цит b/f
ФC I
АДФ
АТФ
Пластида
Гликолиз,
3-ФГК (С3)
Цикл Кальвина
Ф
1,3-ДФГК (С3)
3-ФГК (С3)
Гликолиз
АТФ
3-ФГА(С3)
Ф
Ф
Порины
Порины
NADP+
NADPH
Ф
3-ФГА (С3)
1,3-ДФГК (С3)
АДФ
NADH
NAD+
ЭТЦ
Н2О
О2
Митохондрия
Цитозоль
(симпласт)
45.
ēНО
2
ФC II
Пластида
ФC I
Цит b/f
МАЛИК-ЭНЗИМ:
NADP+зависимая
МДГ декарб.
Порины
Гликолиз
ФЕП-карбоксилаза
NADPH
Оксалоацетат (С4)
Оксалоацетат (С4)
Порины
ЭТЦ
Н2О
О2
Митохондрия АТФ
NADP+зависимая
МДГ декарб.
Малат (С4)
СО2
Гликолиз Пируват (С )
3
Малат
NAD+зависимая
МДГ декарб.
Гликолиз
NADH
АДФ+Ф
NADP+
Пируват- Оксалоацетат
NADH
карбоксилаза
NAD+
Цитозоль
(симпласт)
NAD+зависимая
МДГ декарб.
Малат (С4)
СО2
NAD+
Пируват (С3)
46.
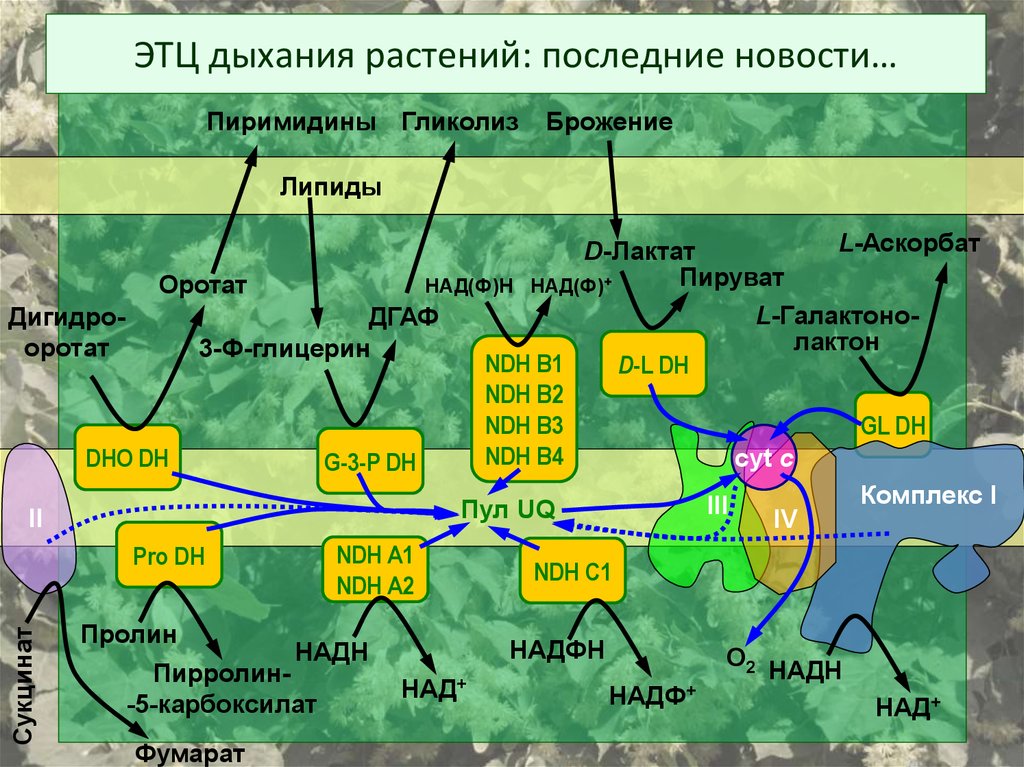
ЭТЦ дыхания растений: последние новости…Пиримидины Гликолиз
Брожение
Липиды
Сукцинат
L-Аскорбат
D-Лактат
Пируват
НАД(Ф)Н НАД(Ф)+
Оротат
L-ГалактоноДигидроДГАФ
лактон
оротат
3-Ф-глицерин
NDH В1
D-L DH
NDH B2
NDH B3
GL DH
NDH B4
cyt c
DHO DH
G-3-P DH
Комплекс I
III
Пул
UQ
II
IV
NDH A1
Pro DH
NDH C1
NDH A2
Пролин
Пирролин-5-карбоксилат
Фумарат
НАДФН
НАДН
НАД+
НАДФ+
О2 НАДН
НАД+
47.
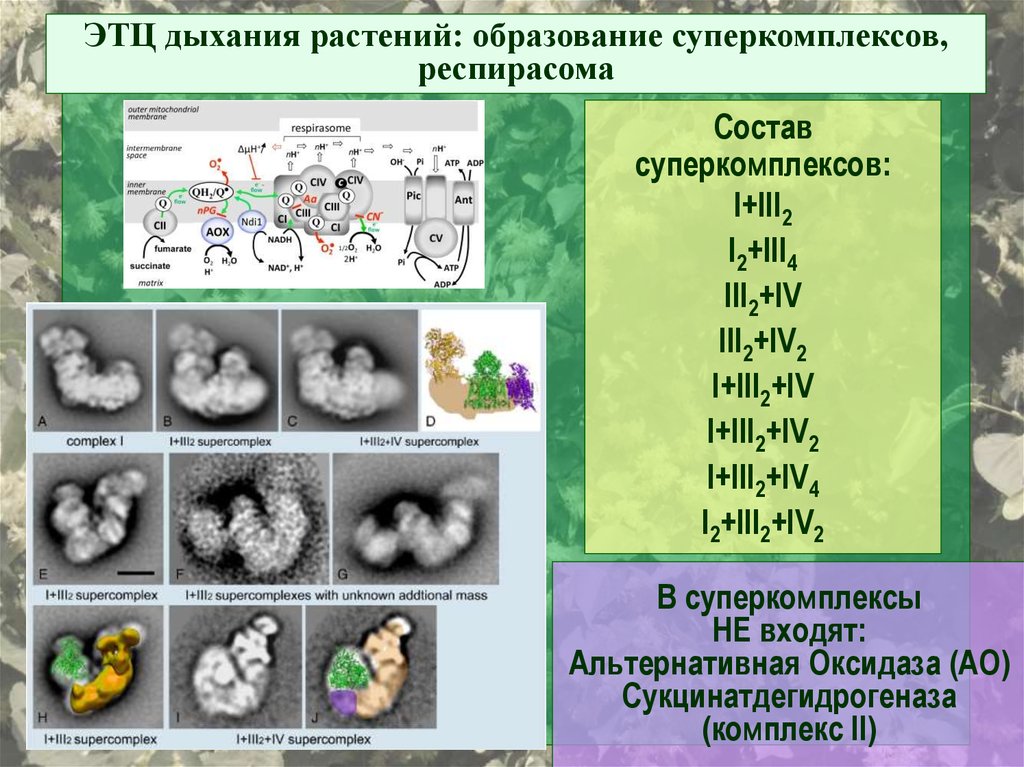
ЭТЦ дыхания растений: образование суперкомплексов,респирасома
Состав
суперкомплексов:
I+III2
I2+III4
III2+IV
III2+IV2
I+III2+IV
I+III2+IV2
I+III2+IV4
I2+III2+IV2
В суперкомплексы
НЕ входят:
Альтернативная Оксидаза (АО)
Сукцинатдегидрогеназа
(комплекс II)
48.
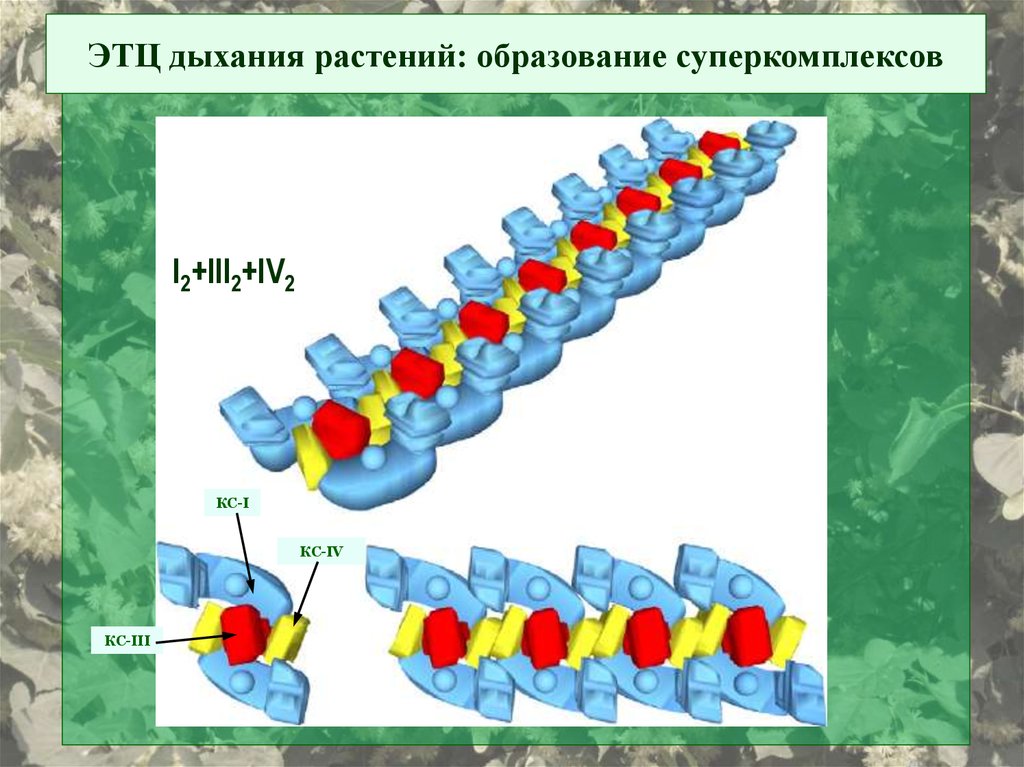
ЭТЦ дыхания растений: образование суперкомплексовI2+III2+IV2
КС-I
КС-IV
КС-III
49.
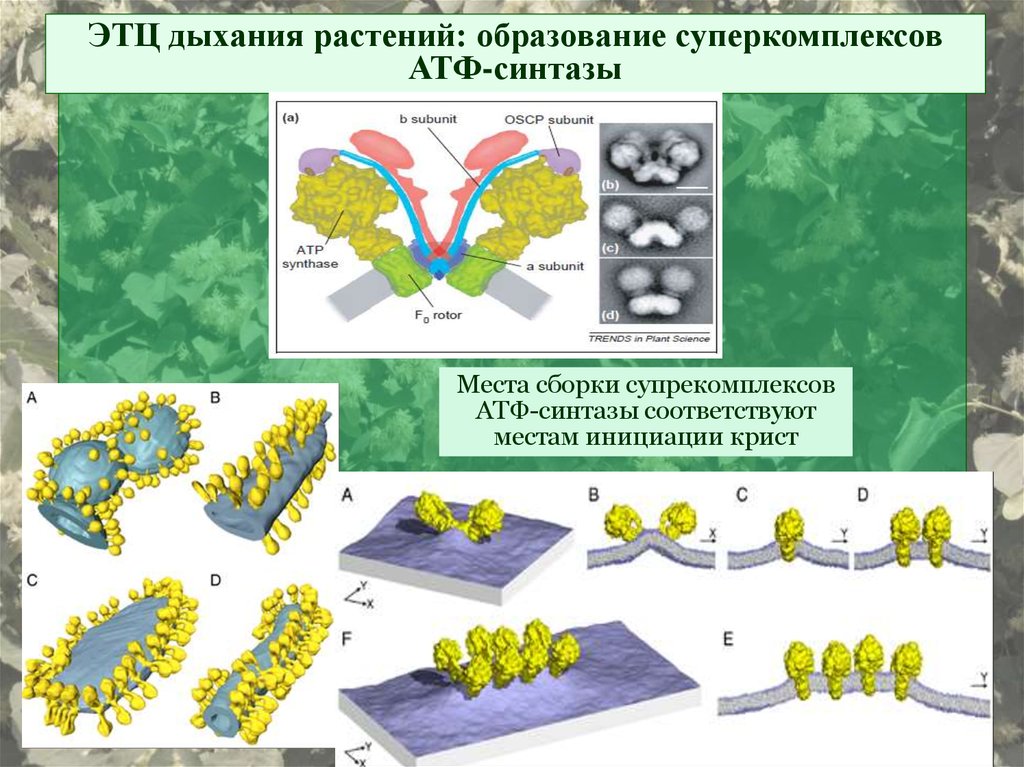
ЭТЦ дыхания растений: образование суперкомплексовАТФ-синтазы
Места сборки супрекомплексов
АТФ-синтазы соответствуют
местам инициации крист
50.
ЭТЦ дыхания растений: образование суперкомплексовАТФ-синтазы
Кристы
работают
как
протонные
«ловушки»
51.
52. Множество кольцевых молекул митохондриальной ДНК растений – результат гомологичных рекомбинаций по повторам.
53. Предполагаемая структура «мастер-хромосомы» митохондрий кукурузы
54. Линейная ДНК митохондрий хламидомонады
55. Сопоставление хлоропластного и митохондриального геномов риса
56. Варианты редактирования хлоропластных и митохондриальных РНК растений
57. Гены митохондрий
1. Синтез белка. - 3 гена рРНК (оперон rrn)- 10 генов белков пластидных рибосом (rpl/rps)
- 16 генов тРНК (trn) – не хватает! – импорт!
2. Дыхание
- 9 генов белков НАД Н дегидрогеназы (nad)
- ген апоцитохрома b (cob);
- 5 генов белков биосинтеза цитохрома с (ccb)
- 3 гена субъединиц цитохромоксидазы (гены сох).
- 3 гена субъединиц сукцинатдегидрогеназы (sdh)
у печеночников
- 4 гена АТФ-синтазы (atp)
Всего: около 50 генов (у печеночных мхов – более 100) ,
из них около 20 - «рабочих» и около 30 - «домашнего хозяйства».
58.
• Ядерный генов контролирует :некоторые т РНК
ДНК – полимеразу
РНК – полимеразу
Некоторые рибосомальные белки
Синтез белков, входящие в состав комплексов
электрон – транспортной цепи дыхания
• Синтез ферментов ЦТК
59.
60.
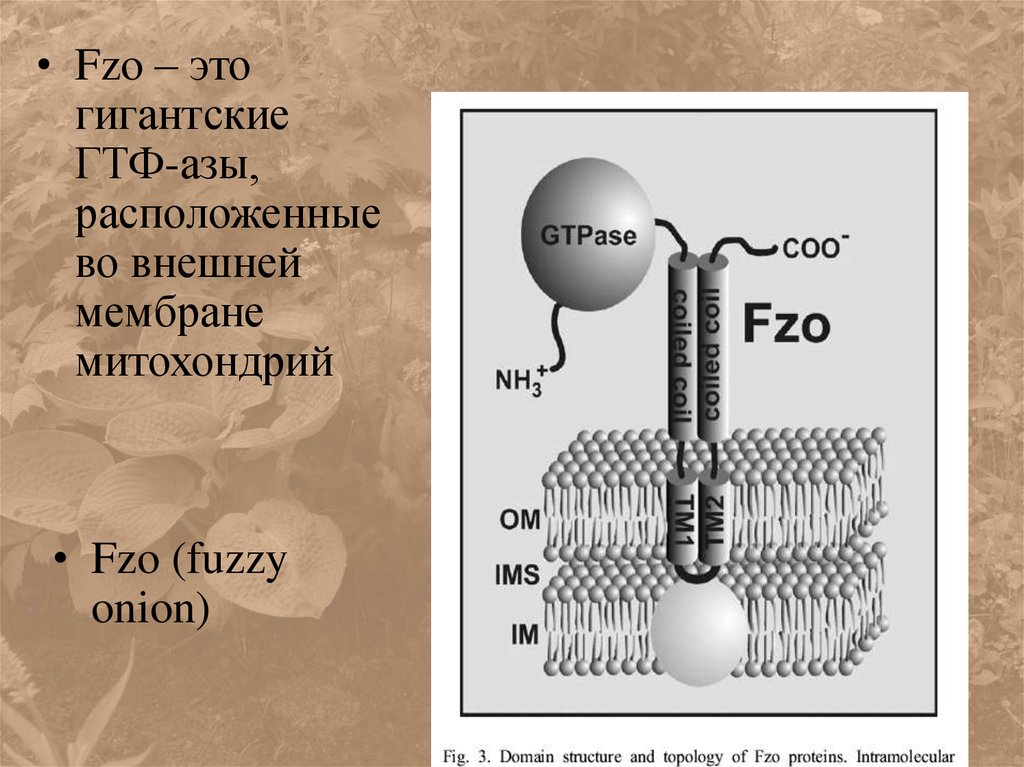
• Fzo – этогигантские
ГТФ-азы,
расположенные
во внешней
мембране
митохондрий
• Fzo (fuzzy
onion)
61. Цитоплазматическая мужская стерильность (ЦМС)
Ядерный ген«Техасская» - Т-ЦМС
Restorer of Fertility =
rf1rf1
= Rf
rf2rf2
Т-мх
стерильность
Т-мх
Rf1Rf1
Rf2Rf2
восстановление
фертильности
«USDA»-тип - S-ЦМС
Ядерный ген Rf3
«Молдавская» - M-ЦМС
«Парагвайская»
(Charrua-тип) - С-ЦМС Ядерный ген Rf4
62. АТФ-синтаза: ключевой фермент «Техасской» ЦМС
13 kDa-белокпоявляется
в митохондриях
как результат
незаконной
рекомбинации
c геном atp6
и встраивается
в АТФ-синтазу,
вызывая ЦМС
63.
АТФ-синтаза: ключевой фермент «Техасской» ЦМС13 kDa-белок
появляется
в митохондриях
как результат
незаконной
рекомбинации
c геном atp6
и встраивается
в АТФ-синтазу,
вызывая ЦМС
Гельминтоспориоз!
64. АТФ-синтаза: ключевой фермент ЦМС у петунии
У петуниинезаконная
рекомбинация
происходит
с участием
гена atp9.
Изменения
в АТФ-синтазном
комплексе
вызывают ЦМС
65. Цитоплазматическая мужская стерильность (ЦМС)
«USDA»-тип - S-ЦМСЯдерный ген Rf3
«Молдавская» - Т-ЦМС
При S-ЦМС(=М-ЦМС)
появляются
линейные фрагменты
в митохондриальном геноме:
S1 и S2 – эписомы.
Возможно, более крупные
кольцевые молекулы мт-ДНК
за счет рекомбинации
могут линеаризоваться.
66. Цитоплазматическая мужская стерильность (ЦМС): спорофитный и гаметофитный контроль
«USDA»-тип - S-ЦМСЯдерный ген Rf3
«Молдавская» - M-ЦМС
S-мх
S-мх
S-мх
rf3rf3
Стерильность
Rf3rf3
Полное (!)
восстановление
фертильности
Rf3Rf3
Восстановление
фертильности
67. Цитоплазматическая мужская стерильность (ЦМС): спорофитный и гаметофитный контроль
«Техасская» - Т-ЦМС Ядерные гены Rf1 и Rf2T-мх
Т-мх
Т-мх
rf1rf1
Rf1rf1
Rf1Rf1
Стерильность
Половина (!)
пыльцевых зерен
Rf2-ген –
фертильна
доминантная
гомозигота
Восстановление
фертильности
68.
69.
70.
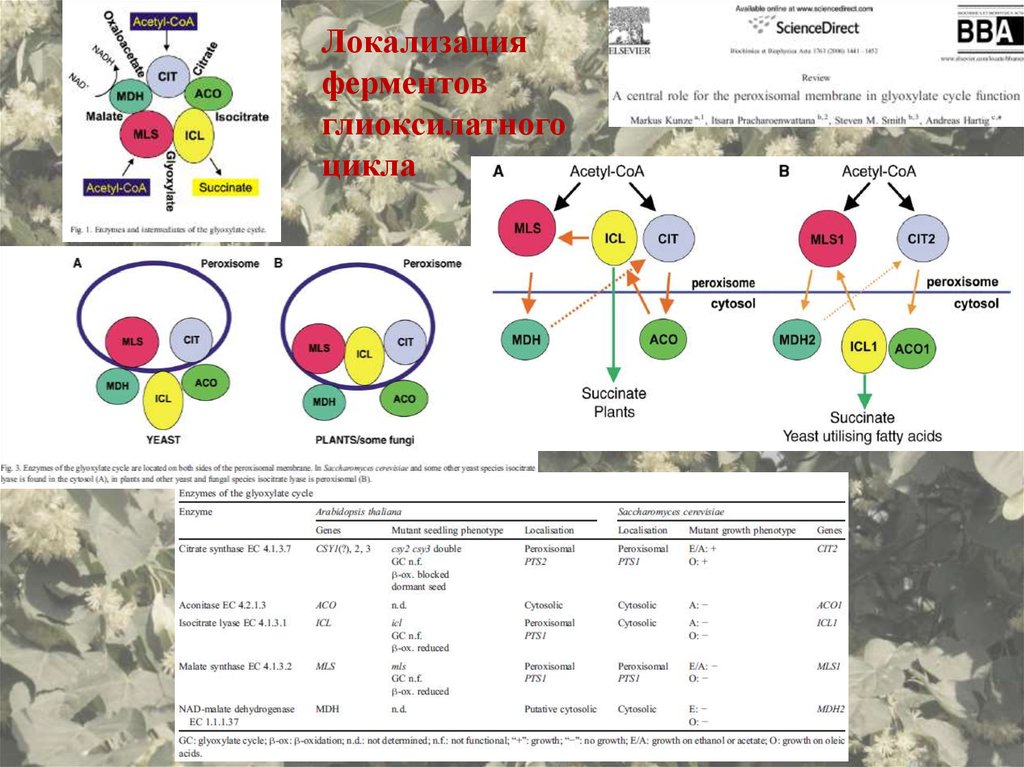
Локализацияферментов
глиоксилатного
цикла
71. Растительные пероксисомы – источник сигнальных молекул
72.
Метаболический переход отгетеротрофного к фототрофному
питанию