 наследование")



")







")



 Биология
БиологияПохожие презентации:










Нехромосомное (цитоплазматическое) наследование
1. Нехромосомное (цитоплазматическое) наследование
2. Пластидная наследственность
Не все эукариотические
гены локализованы в
хромосомах клеточных
ядер. После переоткрытия
менделеевских законов
наследование стало ясно,
что некоторые типы
изменчивости не
подчиняются этим
законам. В 1909 г. Карл
Корренс опубликовал
работу по наследованию
пестролистности у Mirabilis
galapa, в которой был
описан не менделевский
тип наследования.
Зелёные листья ночной
красавицы (Mirabilis jalapa)
3.
Для многих видов декоративных растений
характерно пёстролистность – появление
белых и жёлтых пятен и полос на листьях
зелёных растений. Жёлтые участки могут быть
небольших размеров, но иногда жёлтыми
становятся целые побеги, тогда как другие
остаются зелёными или пёстрыми. Корренс
брал пыльцу с цветков, растущих на жёлтых,
пёстрых и зелёных побегов, и нанося её
(удалив тычинки) на пестики цветов, растущих
на побегах всех трёх типов.
Свойства, проросших из таких семян растений
определяются характером материнского цветка
и не зависят от свойств цветка, с которого
была взята пыльца. Эти результаты были
первым примером цитоплазматической
наследственности.
Корренс предположил наличие «смешанных
клеток» в пестрых участках ткани, в которых
содержатся как зеленые так и бесцветные
пластиды. Позднее Реннер и Баур выдвинули
гипотезу, согласно которой пластиды
(хлоропласты) являются носителями
наследственности, и тип окраски передается
дочерним пластидам при их делении.
Наследование пестролистности у ночной
красавицы — пример материнского типа
наследования.
4.
В отличие от ночной красавицыу герани небольшое число
пластид передается по мужской
линии (двуродительское
наследование). Как показали
более поздние исследования
Тилни—Бассет (1970),
наследование окраски
листьев находится под
ядерным контролем и под
контролем пластид
женского родителя (мужской
родитель оказывает
незначительное влияние изза малого количества
пластид в спермиях и
большого - в яйцеклетке).
•В дальнейших исследованиях было выяснено, что биосинтез хлорофилла, от
которого зависит окраска листа, контролируется главным образом ядерными
генами. Однако некоторые гены локализованы в пластидной ДНК. Окраска
хлоропластов зависит от фотосинтезируемого пигмента - хлорофилла. При
действии мутаций, нарушающих биосинтез хлорофилла, в хлоропластах не
образуется пигмент. В случае пестролистности материнского растения в
яйцеклетке присутствуют пластиды двух типов: нормальные и мутантные, у
которых зеленого хлорофилла нет. При делении зиготы пластиды случайно
распределяются по дочерним клеткам, в результате листья у нового
поколения растений могут быть: зелеными, белыми, либо пестрыми с
чередующимися зелеными и белыми участками.
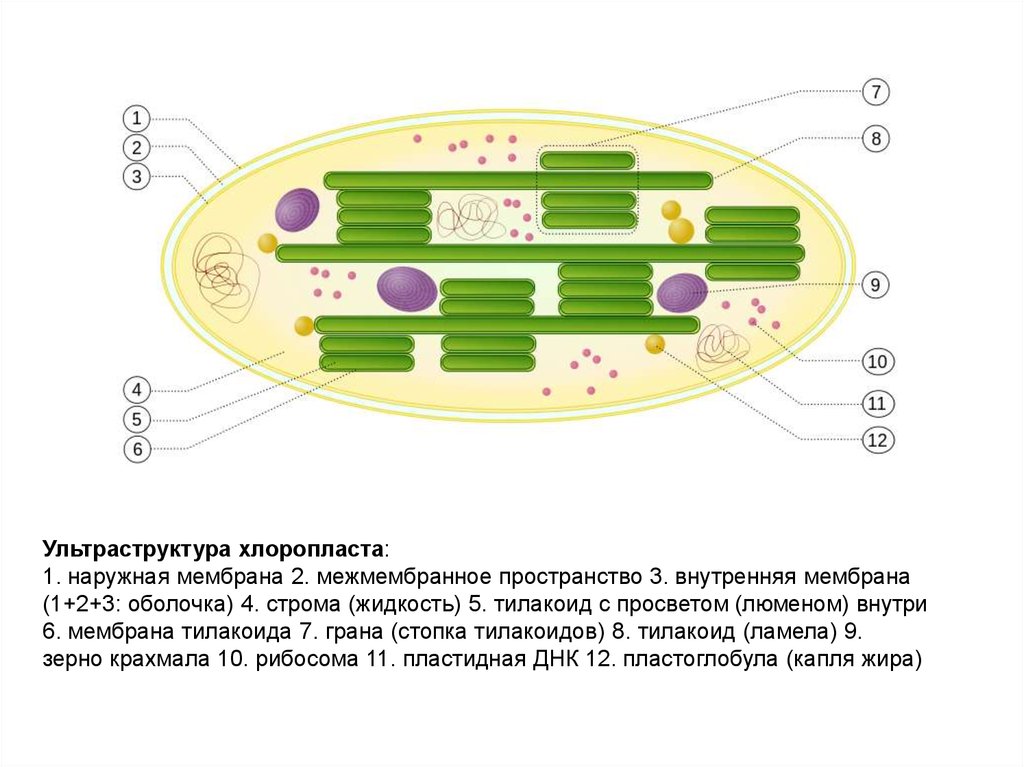
5.
Ультраструктура хлоропласта:1. наружная мембрана 2. межмембранное пространство 3. внутренняя мембрана
(1+2+3: оболочка) 4. строма (жидкость) 5. тилакоид с просветом (люменом) внутри
6. мембрана тилакоида 7. грана (стопка тилакоидов) 8. тилакоид (ламела) 9.
зерно крахмала 10. рибосома 11. пластидная ДНК 12. пластоглобула (капля жира)
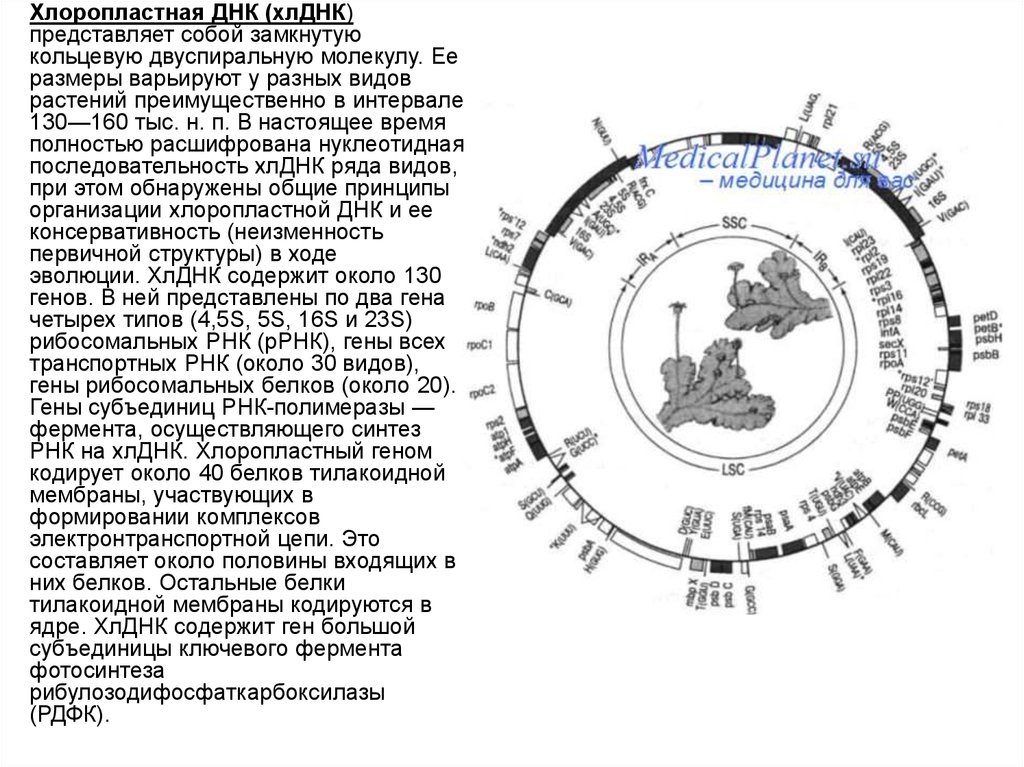
6.
Хлоропластная ДНК (хлДНК)
представляет собой замкнутую
кольцевую двуспиральную молекулу. Ее
размеры варьируют у разных видов
растений преимущественно в интервале
130—160 тыс. н. п. В настоящее время
полностью расшифрована нуклеотидная
последовательность хлДНК ряда видов,
при этом обнаружены общие принципы
организации хлоропластной ДНК и ее
консервативность (неизменность
первичной структуры) в ходе
эволюции. ХлДНК содержит около 130
генов. В ней представлены по два гена
четырех типов (4,5S, 5S, 16S и 23S)
рибосомальных РНК (рРНК), гены всех
транспортных РНК (около 30 видов),
гены рибосомальных белков (около 20).
Гены субъединиц РНК-полимеразы —
фермента, осуществляющего синтез
РНК на хлДНК. Хлоропластный геном
кодирует около 40 белков тилакоидной
мембраны, участвующих в
формировании комплексов
электронтранспортной цепи. Это
составляет около половины входящих в
них белков. Остальные белки
тилакоидной мембраны кодируются в
ядре. ХлДНК содержит ген большой
субъединицы ключевого фермента
фотосинтеза
рибулозодифосфаткарбоксилазы
(РДФК).
7. Митохондриальная наследственность (митохондрион)
Митохондриальная наследственность
(митохондрион)
Митохондриальная
ДНК (мтДНК) — ДНК,
находящаяся (в отличие
от ядерной ДНК) в
митохондриях,
органоидах эукариотическ
их клеток.
Гены, кодированные
мтДНК, относятся к
группе плазмагенов,
расположенных вне ядра
(вне хромосомы).
Совокупность этих
факторов
наследственности,
сосредоточенных в
цитоплазме клетки,
составляет плазмон данн
ого вида организмов (в
отличие от генома)
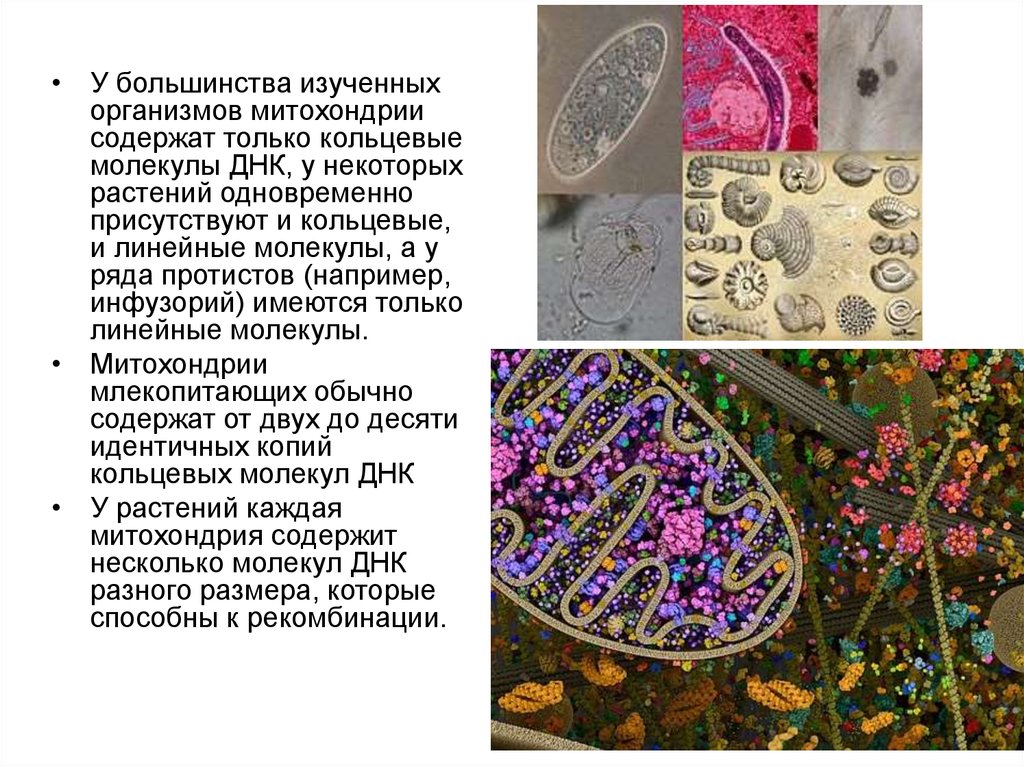
8.
• У большинства изученныхорганизмов митохондрии
содержат только кольцевые
молекулы ДНК, у некоторых
растений одновременно
присутствуют и кольцевые,
и линейные молекулы, а у
ряда протистов (например,
инфузорий) имеются только
линейные молекулы.
• Митохондрии
млекопитающих обычно
содержат от двух до десяти
идентичных копий
кольцевых молекул ДНК
• У растений каждая
митохондрия содержит
несколько молекул ДНК
разного размера, которые
способны к рекомбинации.
9. Геном митохондрий
У млекопитающих каждая молекула
мтДНК содержит 15000-17000 пар
оснований (у человека 16565 пар
нуклеотидов, по другому источнику
16569 пар) и содержит 37 генов — 13
кодируют белки, 22 — гены тРНК, 2 —
рРНК (по одному гену для 12S и 16S
рРНК). Другие многоклеточные животные
имеют схожий набор митохондриальных
генов, хотя некоторые гены могут иногда
отсутствовать.
Генный состав мтДНК разных видов
растений, грибов и особенно
протистов различается более
значительно. Так, у жгутиконосцаякобиды Reclinomonas americana найден
наиболее полный из известных
митохондриальных геномов: он
содержит 97 генов, в том числе 62 гена,
кодирующих белки
(27 рибосомальных белков, 23 белка,
участвующих в работе электронтранспортной цепи и в окислительном
фосфорилировании, а также
субъединицы РНК-полимеразы).
Один из наиболее маленьких
митохондриальных геномов
имеет малярийный плазмодий (около
6.000 п.о., содержит два гена рРНК и три
гена, кодирующих белки).
Схема митохондриального генома человека
10. Геном митохондрий человека кодирует следующие белки и РНК:
Белки или РНКГены
NADH-дегидрогеназа
MT-ND1, MT-ND2, MT-ND3, MT-ND4, MTND4L, MT-ND5, MT-ND6
(комплекс I)
Кофермент Q - цитохром c
редуктаза/Цитохром b
(комплекс III)
MT-CYB
цитохром c оксидаза
(комплекс IV)
MT-CO1, MT-CO2, MT-CO3
АТФ-синтаза
MT-ATP6, MT-ATP8
рРНК
MT-RNR1 (12S), MT-RNR2 (16S)
тРНК
MT-TA, MT-TC, MT-TD, MT-TE, MT-TF, MTTG, MT-TH, MT-TI, MT-TK, MT-TL1, MTTL2, MT-TM, MT-TN, MT-TP, MT-TQ, MTTR, MT-TS1, MT-TS2, MT-TT, MT-TV, MTTW, MT-TY, MT1X
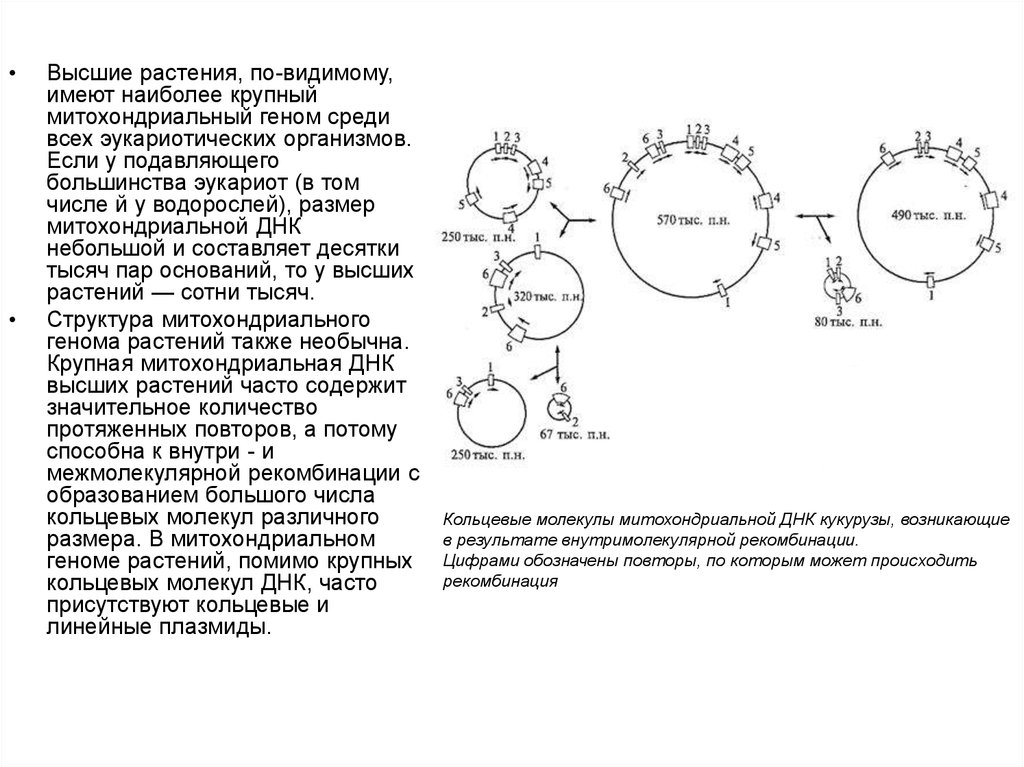
11.
Высшие растения, по-видимому,
имеют наиболее крупный
митохондриальный геном среди
всех эукариотических организмов.
Если у подавляющего
большинства эукариот (в том
числе й у водорослей), размер
митохондриальной ДНК
небольшой и составляет десятки
тысяч пар оснований, то у высших
растений — сотни тысяч.
Структура митохондриального
генома растений также необычна.
Крупная митохондриальная ДНК
высших растений часто содержит
значительное количество
протяженных повторов, а потому
способна к внутри - и
межмолекулярной рекомбинации с
образованием большого числа
кольцевых молекул различного
размера. В митохондриальном
геноме растений, помимо крупных
кольцевых молекул ДНК, часто
присутствуют кольцевые и
линейные плазмиды.
Кольцевые молекулы митохондриальной ДНК кукурузы, возникающие
в результате внутримолекулярной рекомбинации.
Цифрами обозначены повторы, по которым может происходить
рекомбинация
12.
•К настоящему временипроведено полное
секвенирование
митохондриальной ДНК
некоторых водорослей, а
также высших растений
(печеночника, арабидопсиса,
риса). Показано, что у разных
объектов в составе мтДНК
содержится от 25 (у
хламидомонады) до 104 (у
печеночника) генов. У высших
растений мтДНК содержит,
как правило, чуть больше 50
генов Подавляющая часть
генома митохондрий растений
состоит из некодирующих
последовательностей. Гены
отделены друг от друга по
меньшей мере несколькими
тысячами нуклеотидных пар,
поэтому каждый ген обычно
считывается автономно.
Схема строения кольцевой ДНК («мастер»-хромосомы) митохондрий
Кукурузы
13. Митохондриальная наследственность - наследование по материнской линии
Митохондриальная наследственность наследование по материнской линии• Поскольку в сперматозоиде, который вносит половину хромосом
будущего организма, содержится мало митохондрий, которые
обычно разрушаются после оплодотворения, митохондриальная
наследственность определяется, в основном,
митохондриями яйцеклетки.
14. Митохондриа́льные заболева́ния — группа наследственных заболеваний, связанных с дефектами в функционировании митохондрий,
Митохондриа́льные заболева́ния — группа наследственныхзаболеваний, связанных с дефектами в функционировании митохондрий,
приводящими к нарушениям энергетических функций в
клетках эукариотов, в частности — человека
Митохондриальные заболевания обусловлены генетическими,
структурными, биохимическими дефектами митохондрий, приводящими
к нарушениям тканевого дыхания. Они передаются только по женской
линии к детям обоих полов. Патологические нарушения клеточного
энергетического обмена могут проявляться в виде дефектов различных
звеньев в цикле Кребса, в дыхательной цепи, процессах бетаокисления и т. д.
У человека можно выделить 2 группы митохондриальных заболеваний:
ярко выраженные наследственные синдромы,
обусловленные мутациями генов, ответственных за
митохондриальные белки (синдром Барта, синдром КернсаСейра, синдром Пирсона, синдром MELAS, синдром MERRF и другие).
«Вторичные митохондриальные заболевания», включающие
нарушение клеточного энергообмена как важное звено
формирования патогенеза (болезни соединительной ткани, синдром
хронической
усталости, гликогеноз, кардиомиопатия, мигрень, печеночная
недостаточность, панцитопения, а
также гипопаратиреоз, диабет, рахит и другие).
15. Применение исследований митохондриальной ДНК
• Так как митохондриальная ДНК не является высококонсервативной иимеет высокую скорость мутирования, она является хорошим
объектом для изучения филогении (эволюционного родства) живых
организмов. Для этого определяют последовательности
митохондриальной ДНК у разных видов и сравнивают их при помощи
специальных компьютерных программ и получают эволюционное
древо для изученных видов. Исследование митохондриальных ДНК
собак позволило проследить происхождение собак от диких
волков. Исследование митохондриальной ДНК в популяциях человека
позволило вычислить «митохондриальную Еву», гипотетическую
прародительницу всех живущих в настоящее время людей.
Работы по определению гаплотипов
(специфических нуклеотидных
последовательностей) человека на основе
исследований митохондриальной ДНК и
Y-хромосомы позволяют прослеживать
пути миграции, восстанавливать
историю народов.
16. Происхождение ДНК органелл
Широко распространено мнение,
что митохондрии и хлоропласты
произошли от прокариотических
эндосимбионтов, которые обитали в
цитоплазме предшественников эукариот.
Как полагают, митохондриям дали начало
пурпурные бактерии, а хлоропластам
(позднее) — цианобактерии (синезеленые
водоросли) или близкие к ним организмы.
Симбионты проникли в эукариотические
клетки и в ходе эволюции потеряли свою
автономность, передав большое число
важнейших генов в ядерный геном. В
результате независимая бактериальная
клетка превратилась в полуавтономную
органеллу, сохранившую главную
исходную функцию — способность к
фотосинтезу (у хлоропластов) и систему
окислительного фосфорилирования (у
митохондрий).
Хотя многие гены этих древних бактерий
все еще используются для синтеза белков
органеллы, большая их часть по неясным
причинам включилась в ядерный геном,
где они кодируют ферменты, которые
сходны с бактериальными и
синтезируются на рибосомах в
цитоплазме, а затем переходят в
органеллу.
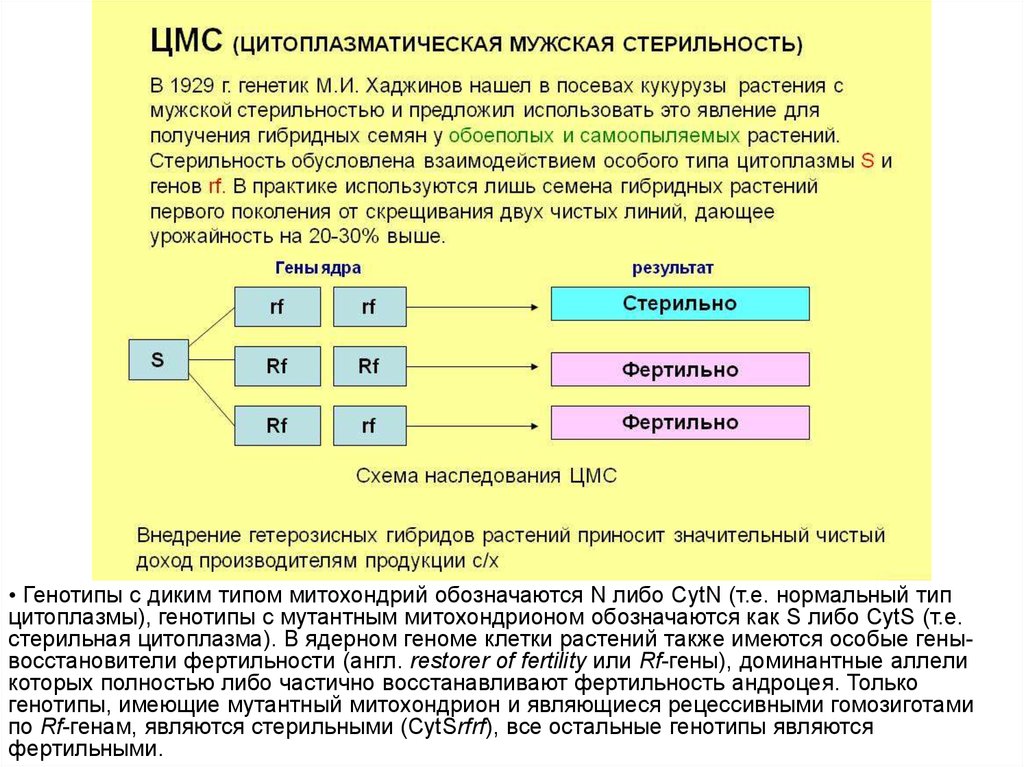
17. Мужская стерильность, цитоплазматическая мужская стерильность, ЦМС (male sterility, cytoplasmic male sterility, CMS)
стерильность, передаваемая мужскими цитоплазматическими факторами
наследственности; известна у многих растений (признак М.с. у кукурузы
сопровождается недоразвитием пыльников и формированием аномальной
пыльцы), а также у некоторых межвидовых гибридов насекомых.
Цитоплазматическая мужская стерильность проявляется во
взаимодействии ядерного генома с митохондрионом. Причиной
явления полной или частичной стерильности андроцея высших растений
является наличие особой мутации в митохондрионе,
фертильность растений восстанавливается полностью или частично при
наличии доминантного аллеля ядерного гена-восстановителя
фертильности.
18.
• Генотипы с диким типом митохондрий обозначаются N либо CytN (т.е. нормальный типцитоплазмы), генотипы с мутантным митохондрионом обозначаются как S либо CytS (т.е.
стерильная цитоплазма). В ядерном геноме клетки растений также имеются особые генывосстановители фертильности (англ. restorer of fertility или Rf-гены), доминантные аллели
которых полностью либо частично восстанавливают фертильность андроцея. Только
генотипы, имеющие мутантный митохондрион и являющиеся рецессивными гомозиготами
по Rf-генам, являются стерильными (CytSrfrf), все остальные генотипы являются
фертильными.
19. Практическая цель использования цитоплазматической мужской стерильности
- перевод линии на стерильную основу свосстановлением мужской
фертильности. Это предполагает
создание стерильного аналога
материнского отца и аналогавосстановителя родительского.
• Селекция кукурузы на гетерозис с
использованием ЦМС
осуществляется в следующей
последовательности:
• создание самоопыляющихся линий и
их оценка;
• создание и испытание гибридов;
• перевод перспективных гибридов на
стерильную основу с
восстановлением мужской
фертильности;
• размножение исходных форм
гибридов.
Использование цитоплазматической
мужской стерильности в
семеноводстве кукурузы позволяет
осуществлять получение гибридных
семян на больших площадях без
трудоемкой операции обрыва
метелок
20. ЦМС у кукурузы
У кукурузы (Zea mays L) известно несколько типовЦМС, наиболее изученными являются т.н. Т-тип
ЦМС (называемый также техасским), С-тип
ЦМС (т.н. чарруа тип, также называемый
парагвайским и колумбийским) и S-тип
ЦМС (называемый также молдавским или Мтипом ЦМС). Каждый тип ЦМС определяется
своей специфичной мутацией в митохондрионе и
восстанавливается своими генамивосстановителями фертильности.
S-тип ЦМС обусловлен мутацией открытых рамок
считывания orf355 и orf77, образуется химерная
нуклеотидная последовательность R orf355-orf77.
Геном-восстановителем фертильности
является Rf3, ввиду постмейотического
восстановления фертильности
генотипы Rf3rf3 образуют 50% фертильной
пыльцы. Было показано, что Rf3 оказывает
влияние на уровни экспрессии митохондриальных
и ядерных генов и
оказывает плейотропное действие на
уровне транскрипции. Возможным механизмом Sтипа ЦМС у кукурузы является изменение уровня
экспрессии генов у orf355-orf77 растений и
включении механизма программируемой
клеточной гибели, восстановление же
фертильности обусловлено нормализацией
уровня транскриптов в митохондриях и
ингибировании апоптотических механизмов.
21. Собственно цитоплазматическое наследование
Собственно цитоплазматическое
наследование
В некоторых случаях цитоплазма сама
по себе может детерминировать
наследуемые признаки, однако
наследование признака при этом
нестойкое и затухает в течение одного
или нескольких поколений.
Наиболее известным примером
собственно цитоплазматического
наследования является наследование
формы раковины у прудовика. Она
может быть правозакрученной (D,
доминантный аллель) или
левозакрученной (d, рецессивный
аллель). При этом сам генотип моллюска
никакого влияния на форму раковины не
оказывает. Это определяется свойствами
материнского организма, а именно
цитоплазмы яйцеклетки, которая и
обусловливает направление
закручивания раковины (как раз эти
свойства цитоплазмы и определяются
геном D). При этом у материнского
организма с генотипом dd все потомки
будут левозакрученными, а с
генотипом Dd или DD —
правозакрученными, даже если он сам
имеет левозакрученную раковину