























 Биология
БиологияПохожие презентации:










Слух. Орган слуха
1. Слух
2. Орган слуха
• Наружное ухо: «рупор» для сбора колебаний воздуха.• Среднее ухо: энергия колебаний воздуха улавливается
барабанной перепонкой и передается слуховыми
косточками на стенку улитки («овальное окно»). В
результате возникают колебания лимфы, наполняющей
улитку, что приводит к изгибу волосков и возбуждению
расположенных вдоль улитки рецепторов.
• Внутреннее ухо. Состоит из улитки (слуховая часть), а
также вестибулярных мешочков и каналов, наполненных
лимфой.
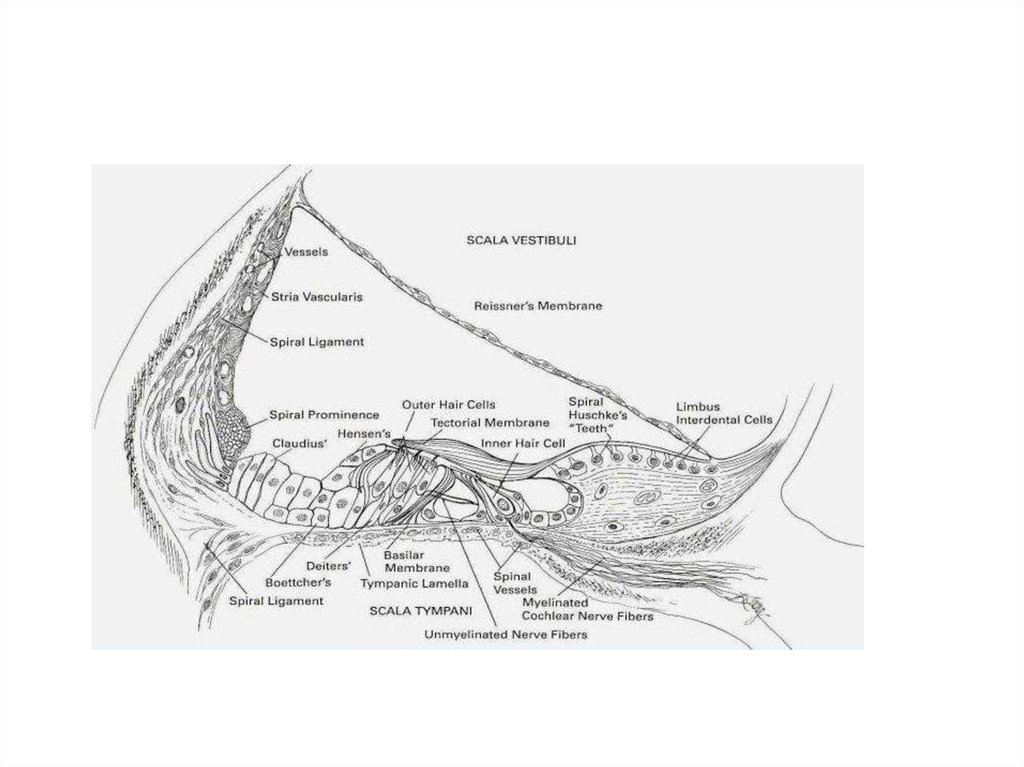
3. Улитка
Улитка: две мембраны, три канала.Лестница преддверия (верхний
канал), барабанная лестница
(нижний канал) и улитковый проток
(средний канал);
Рейснерова (верхняя) и базилярная
мембраны
+ текториальная мембрана над
волосковыми рецепторами.
4. Улитка
5. Как улитка позволяет различать тональность?
Механизм частотной избирательности, основанный на строении и свойствах улитки«арфа». Гельмгольц:
Вообще-то базилярная мембрана имеет треугольную форму (на рисунке- будто раскрутили
улитку). У хеликотремы широкая, а рядом с овальным окном- узкая. Получается
конструкция, которая натянута на костяных выростах. В самом широком месте натянуты
самые короткие струны арфы, резонирующие на высокие звуки, в узком месте (у овального
окна)- только длинные струны. Они резонируют на низких частотах. Работает физический
принцип резонанса.
6. Как улитка позволяет различать тональность?
Другой принцип- бегущей волны. Аналогия- раскачивание линейки. Раскачивание с
небольшой частотой- волны убегают далеко, с большой- волны затухают вблизи места
возникновения. Из-за того, что жесткость базилярной мембраны различна на ее
протяжении.
Эти механизмы сосуществуют в улитке.
Возможно, стереоцилии на протяжении баз. Мембраны также меняют жесткость.
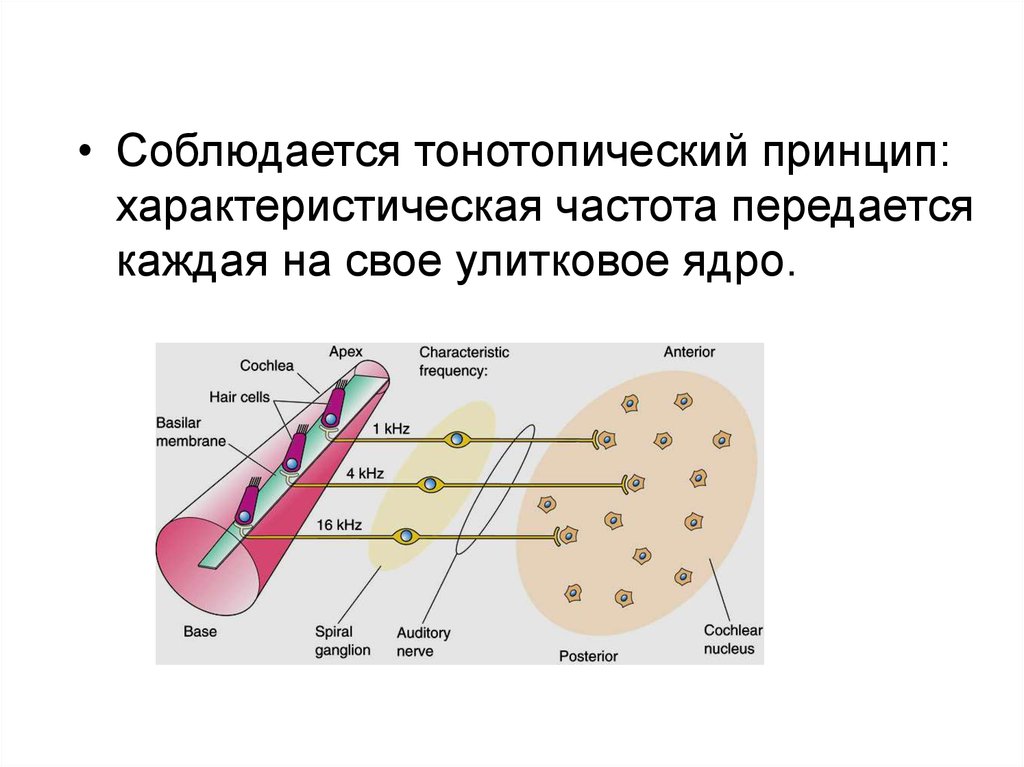
7.
• Соблюдается тонотопический принцип:характеристическая частота передается
каждая на свое улитковое ядро.
8. Кортиев орган
Кортиев органНаружные
волосковые
клетки
Чувствительные
волоски
Базилярная
мембрана
Текториальная
мембрана
Палочки
Корти
Внутренняя
волосковая
клетка
(organum spirale).
Внутренние ВК -1 ряд
(улавливают звук).
Наружные ВК – 3 ряда
(«сенсорно-двигательные» усиливают звук).
Ретикулярная
пластина
Спиральный ганглий
Слуховой нерв
9.
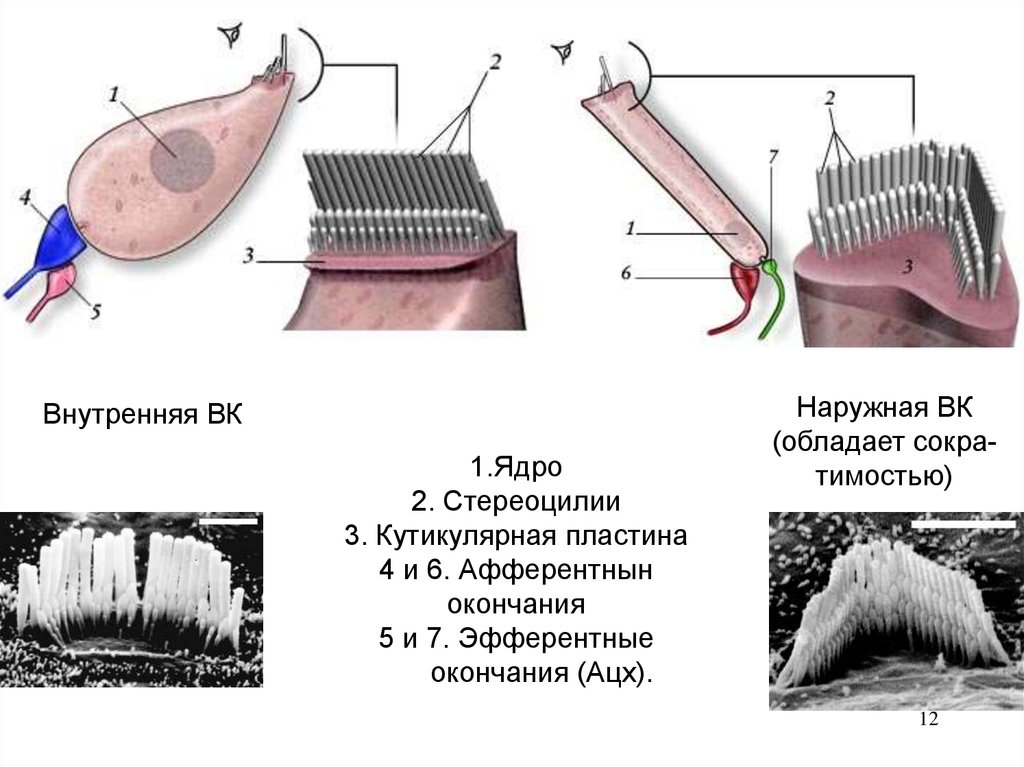
10. Рецепторы
Волосковые клетки- вторичные механорецепторы. На
апикальных концах клеток- стереоцилии (видоизмененные
микроворсинки). Располагаются на поддерживающих
клетках- фалангиоцитах. Делятся на 2 типа:
Внутренние ВК -1 ряд (улавливают звук).
Наружные ВК – 3 ряда («сенсорно-двигательные» усиливают звук). Их стереоцилии числом 100- 300 своими
вершинами прикасаются к внутренней поверхности
покровной мембраны.
11. Калиевые каналы волосковых клеток
• Характеристическая частота каждойклетки определяется плотностью и
кинетическими свойствами кальцийактивируемых калиевых каналов.
Называются они ВК-каналами (от big
K+) из-за большой проводимости. В
клетках с низкими частотами меньше Кпроводимость и медленнее активация.
12.
Внутренняя ВК1.Ядро
2. Стереоцилии
3. Кутикулярная пластина
4 и 6. Афферентнын
окончания
5 и 7. Эфферентные
окончания (Ацх).
Наружная ВК
(обладает сократимостью)
12
13. Трансдукция
В основе ориентационнойизбирательности трансдукции
лежит гипотеза воротной
пружины. Отклонение
волоскового пучка в позитивном
направлении (в сторону более
высоких волосков) увеличивает
расстояние между концами
волосков, растягивает воротную
пружину, открывая таким
образом ворота каналов
трансдукции. Когда пучок
смещается в сторону,
противоположную более
длинным волоскам, происходит
сжатие пружины, и каналы
закрываются.
14. Трансдукция
Открытие каналов → вход К +→деполяризация → вход Ca 2+ →
секреция глутамата → ПД на
окончаниях афферентного нерва
15. Трансдукция
16. Эфферентное торможение
• Нейроны комплекса верхних олив действуютна ипсилатеральную и контралатеральную
улитки и вызывает высвобождение Ацх в
эфферентных синапсах на наружных ВК.
Наружные ВК начинают укорачиваться и
«тянут на себя» текториальную мембрану.
Это укорочение вызвано прямым
воздействием электрического потенциала на
заряженные «моторные» протеины, а не
актин-миозином, как можно было ожидать.
17. Эфферентное торможение: значение
• 1) Понижает чувствительность улиткипри наличии фонового шума.
• 2) Расширяет кривую настройки
афферентов.
• 3) Защищает от повреждения громкими
звуками.
18. Адаптация
Мышца, напрягающаябарабанную перепонку
(m. tensor tympani)
и стременная мышца (m.
stapedius) –
адаптация к громким
звукам.
Овальное и круглое окна в
стенке улитки; наличие
круглого окна
предотвращает слишком
быстрое угашение колебаний
лимфы
19. Адаптация
2 механизма частотной избирательности, характерные в т.ч. для неживой улитки- «бегущая
волна» и «арфа»- обеспечивают плохую частотную избирательность, частотная кривая- с
тупым пиком. Более тонкая избирательность осуществляется из-за эфферентной связи.
Это происходит так: сигнал от всех рецепторов поступает в ядра ГМ, где происходит
латеральное торможение (в первую очередь, ядрами ромбовидной ямки). Обработанный
сигнал поступает в ГМ. ГМ решает, какую частоту надо выделить (например, хотим услышать
звук частотой 1000 Гц- вычленить голос друга из шума на дискотеке)- это работа для
третичной коры. Мозг посылает эфферентный сигнал наружным волосковым клеткам и
вызывает их сокращение. Эти клетки, сокращаясь, увлекают за собой, тянут текториальную
мембрану, увеличивая тем самым амплитуду ее колебаний. Основной сигнал собирается
внутренними волосковыми клетками. Такая настройка волевым усилием вычленяет
частотный диапазон с острым пиком, осуществляется системой возвратного возбуждения.
Нужная нам
частота, Гц
слуховые ядра ромбовидной ямки
20. Адаптация
Волосковые кл-и очень чувствительны. Нужны адаптивные процессы для
устранения фоновых стимулов. В процессе эксперимента доказано, что при
поддержании постоянного отклонении пучка ресничек трансдукционный ток
постепенно спадает при отрицательных значениях МП и менял направление не
спадал при положительных.
Почему?
21. Адаптация
Потому что происходит изменение жесткости пучка стереоцилий благодаря переустановке
жесткости воротной пружины.
Как это происходит?
Миозин движется вдоль актинового каркаса стереоцилии и смещает положение места
прикрепления концевой связи.
Более быстрая адаптация обусловлена ионами кальция, которые заставляют
трансдукционные каналы закрываться
22. Адаптация
• Латеральное торможение,осуществляемое ядрами ромбовидной
ямки и таламусом, см. обработка и
анализ в ЦНС.
23.
43
5
2
1
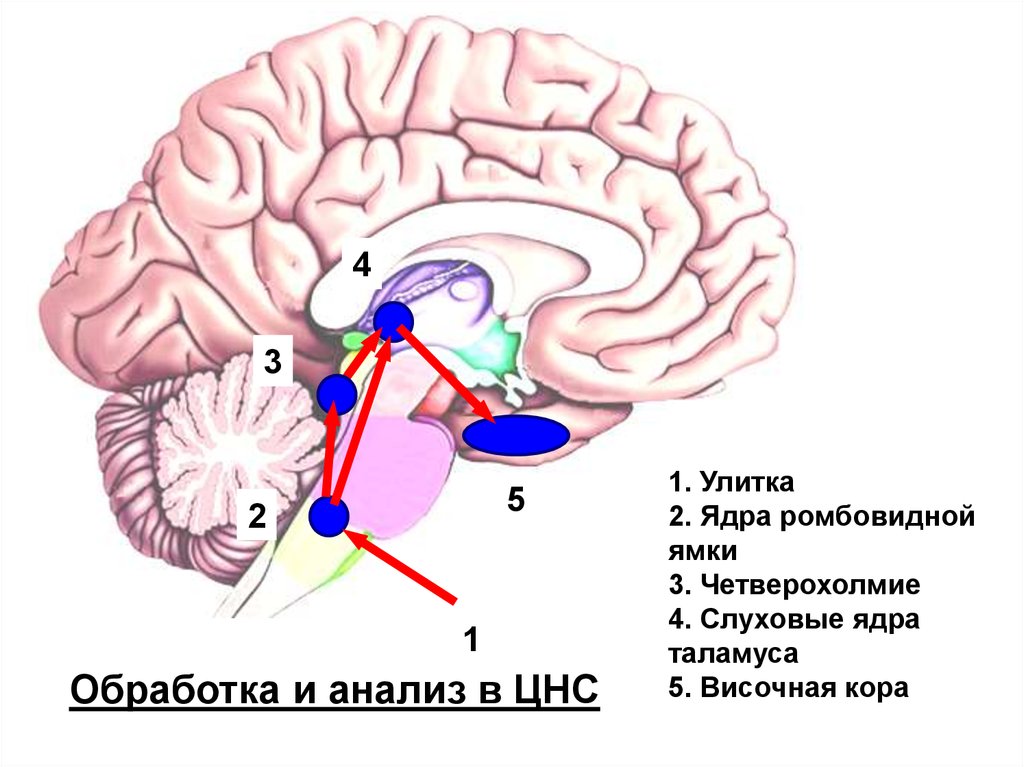
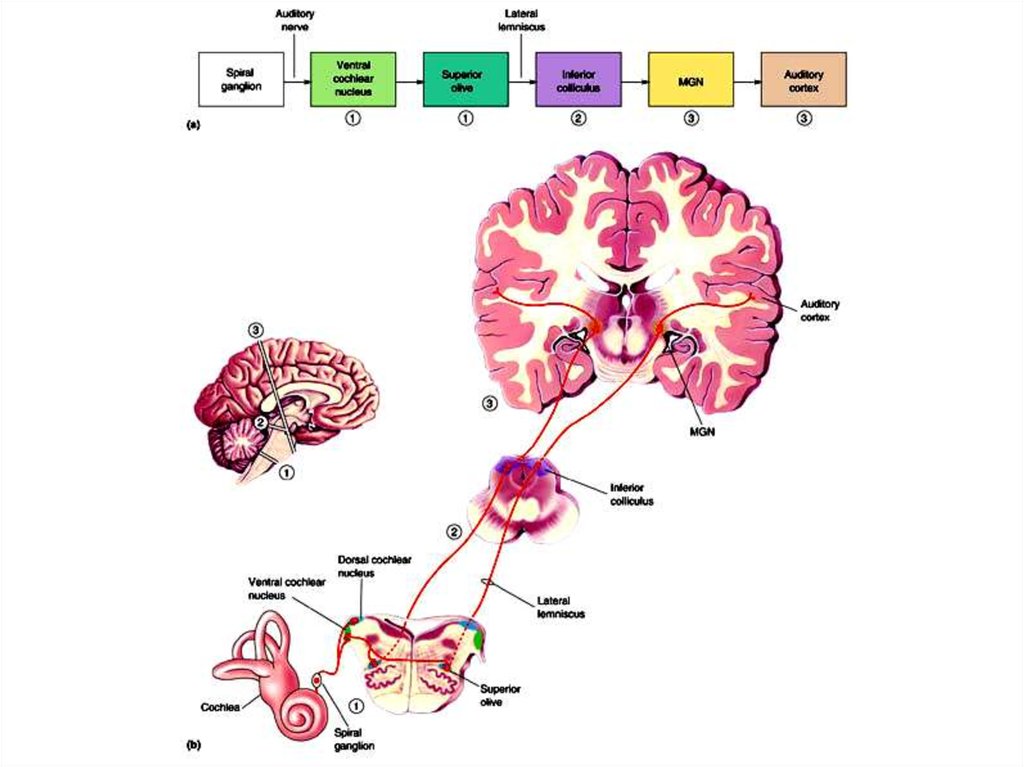
Обработка и анализ в ЦНС
1. Улитка
2. Ядра ромбовидной
ямки
3. Четверохолмие
4. Слуховые ядра
таламуса
5. Височная кора
24.
25. Слуховые центры головного мозга
Слуховые ядра вромбовидной ямке
(продолговатый
мозг/мост)
1) Дорзальные и вентральные
улитковые ядра; ядра верхней
оливы.
Взаимодействие с улиткой (с
наружными ВК)- доп.частотная
избирательность (лат.
торможение)
сравнение сигналов от правой и
левой улитки (по громкости звука
и моменту прихода/фазе
сигнала). Звук оттуда, где громче
и раньше. Животное
утанавливает голову так, чтобы
слышать с одинаковой
громкостью = определение
направления на источник звука.
На этой осно-ве – эхолокация.
26. Слуховые центры головного мозга
ДНТИ
2. Нижние холмики
четверохолмия: новизна
(изменение частоты и
громкости, движение
источника звука).
• нейроны-детекторы новизны
(ДН) – сравнение текущего
сигнала с тем, который был
«только что» (доли секунды
назад, передается через
тормозный интернейрон: ТИ).
• При несовпадении – запуск
ориентировочного рефлекса
(через глазодвигательные
центры и тектоспинальный
тракт; у животных – отдельно
двигаются ушные раковины).
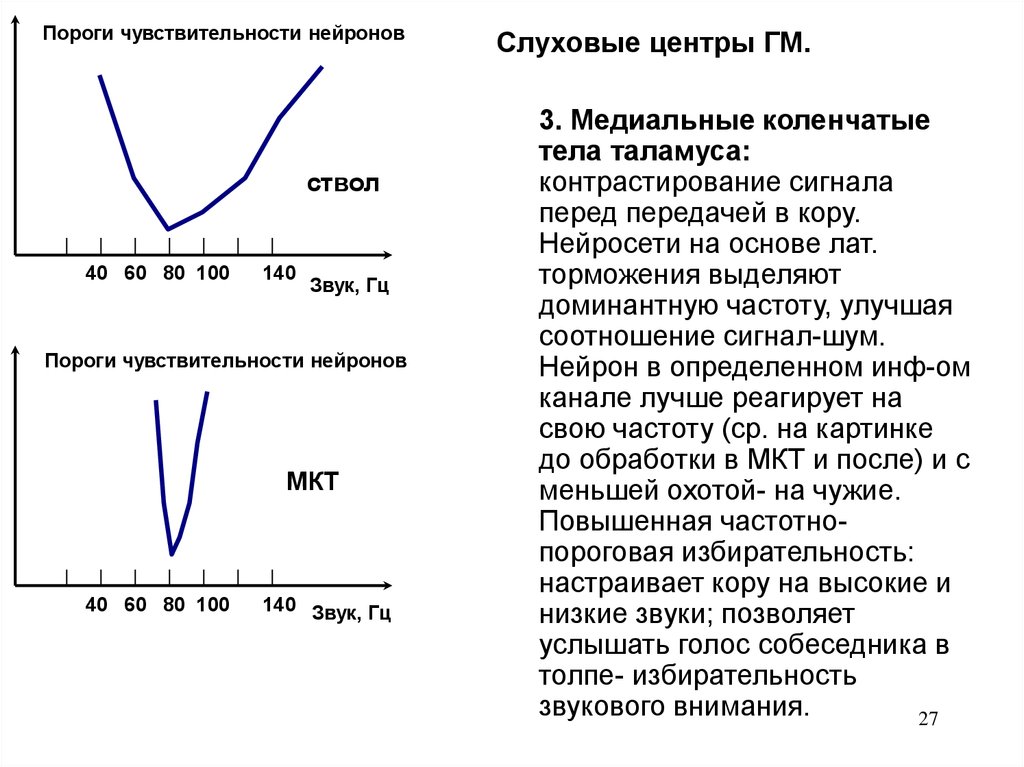
27.
Пороги чувствительности нейроновствол
40 60 80 100
140
Звук, Гц
Пороги чувствительности нейронов
МКТ
40 60 80 100
140 Звук, Гц
Слуховые центры ГМ.
3. Медиальные коленчатые
тела таламуса:
контрастирование сигнала
перед передачей в кору.
Нейросети на основе лат.
торможения выделяют
доминантную частоту, улучшая
соотношение сигнал-шум.
Нейрон в определенном инф-ом
канале лучше реагирует на
свою частоту (ср. на картинке
до обработки в МКТ и после) и с
меньшей охотой- на чужие.
Повышенная частотнопороговая избирательность:
настраивает кору на высокие и
низкие звуки; позволяет
услышать голос собеседника в
толпе- избирательность
звукового внимания.
27
28. Слуховые центры головного мозга
Первичная – височная доля,
по границе латеральной борозды, поля 41
и 42 по Бродману. Организована в виде
изочастотных полос или пластин коры,
идущих перпендикулярно к тонотопической
оси.
Завершение частотно-амплитудного
анализа, наиболее «детальная» тонотопическая карта (до этого момента, далее
нет).
Интересно: слуховая кора активируется и
при чтении по губам.
29. Слуховые центры головного мозга
• Ниже расположена вторичнаяслуховая кора – опознавание звуковых
образов как совокупности частот (шумы,
«звуки природы» и т.п.).
• Как правило, свойства нейронов этой
области – результат обучения.
• Невербальная коммуникация (плач,
смех и т.п.) опознается врожденно.
30. Слуховые центры головного мозга
Задняя часть височной доли – третичная слуховая кора:
узнавание наиболее сложных слуховых образов (музыки,
речи).
Узнавание речи на слух – зона Вернике (доминантное
полушарие)
Основная проблема: нужно реагировать не на частоты и их
совокупность, а общую форму спектра (вне зависимости от
конкретных частот) + это надо делать в реальном времени.
Спектр
слова