













































")
")







 Биология
БиологияПохожие презентации:
")









Эндокринология. Общие свойства гормонов
1. Эндокринология Общие свойства гормонов
Лекция 6Леч.Фак
2.
23. Гормоны-определение
• Гормоны - это биологически активныевещества, которые синтезируются в
малых количествах в
специализированнных клетках
эндокринной системы и через
циркулирующие жидкости (например,
кровь) доставляются к клеткаммишеням, где оказывают свое
регулирующее действие.
3
4. Информация структуры Информация действия
СТРУКТУРАИНФОРМАЦИЯ
структуры
ЭНЕРГИЯ СТРУКТУРЫ
(гормоны)
ИНФОРМАЦИЯ ДЕЙСТВИЯ
ФУНКЦИЯ
(свет, ЭМП)
4
5. Гормоны, как и другие сигнальные молекулы, обладают некоторыми общими свойствами.
1. выделяются из вырабатывающих их клеток вовнеклеточное пространство;
2. не являются структурными компонентами клеток и не
используются как источник энергии;
3. способны специфически взаимодействовать с
клетками, имеющими рецепторы для данного
гормона;
4. обладают очень высокой биологической
активностью - эффективно действуют на клетки в
очень низких концентрациях (около 10-6 -10-11
моль/л), за счет взаимодействия с вторичными
посредниками.
5
6.
• Гормональная регуляция — пример эволюционнонаиболее древнего химического дистантного и
локального контроля.
• Термин гормон введен Р. Старлингом (1905) для
секретина, выделенного из двенадцатиперстной
кишки и активирующего через кровь секрецию
сока поджелудочной железы.
• Доказательствами гормональной природы того
или иного вещества служат следующие признаки:
выделение в кровь; наличие структуры —
«мишени»; существование специфического
рецептора; подавление синтеза гормона или
невозможность связи с рецептором должно
приводить к отсутствию гормонального эффекта.
6
7.
• Во второй половине XX века былопоказано, что в гипоталамической области
мозга существуют клетки способные
генерировать и распространять нервные
импульсы.
• Они демонстрируют свойства, присущие
эндокринным клеткам.
• Последние получили название
нейрогормонов, или нейропептидов.
• Это привело к развитию новой для своего
времени области знаний —
нейроэндокринологии
7
8.
• Оказалось, что нейроэндокринные клетки мозга иклетки иммунной системы функционируют в тесной
кооперации и проявляют признаки сходства в
своей организации.
• Иммунные функции стали рассматривать как
составляющую нейроэндокринной активности, а
сам по себе иммунный ответ невозможен без
содружественного участия нервной и эндокринной
систем.
• Столь тесные взаимосвязи основных
регулирующих систем стали основанием к
выделению интегральной медико-биологической
дисциплины нейроиммуноэндокринологии.
8
9.
• Было показано, что в ответ на действиепатогенных агентов из макрофагов выделяется
ИЛ-1.
• Этот иммунный пептид способен проникать в
мозг через гемато-энцефалический барьер.
• Попав в мозг, ИЛ-1 стимулирует секрецию
кортиколиберина.
• Кортиколиберин стимулирует секрецию АКТГ в
гипофизе, что приводит к стимуляции секреции
глюкокортикоидных гормонов в коре
надпочечников, тормозящих процесс
воспаления.
9
10.
• Взаимодействия нейроэндокринной ииммунной систем в различные периоды
жизни изменяются.
• С возрастом снижаются реакции
иммунной защиты и параллельно
снижается активность гормона роста.
Снижение функции иммунной защиты с
возрастом идет параллельно с
повышением секреции АКТГ.
10
11.
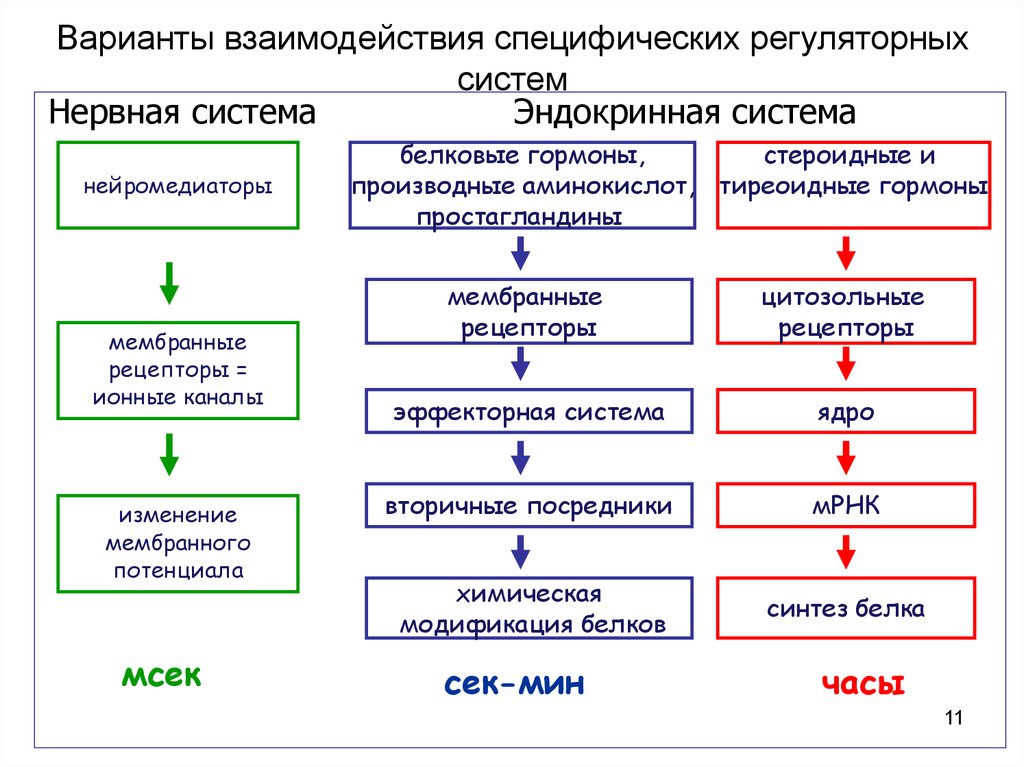
Варианты взаимодействия специфических регуляторныхсистем
Нервная система
Эндокринная система
нейромедиаторы
мембранные
рецепторы =
ионные каналы
изменение
мембранного
потенциала
мсек
белковые гормоны,
стероидные и
производные аминокислот, тиреоидные гормоны
простагландины
мембранные
рецепторы
цитозольные
рецепторы
эффекторная система
ядро
вторичные посредники
мРНК
химическая
модификация белков
синтез белка
сек-мин
часы
11
12.
• Эндокринная система организма животных ичеловека включает комплекс центральных и
периферических эндокринных желез, а также
другие гормон—продуцирующие структуры.
• Центральные выделяют тропные гормоны,
стимулирующие секреторную активность
периферических желез.
• Периферические — щитовидная,
паращитовидные, вилочковая (тимус),
поджелудочная железы, надпочечники и гонады.
• Гормон—продуцирующие структуры: сердце,
легкие, печень, почки, сенсорные и
репродуктивные органы.
12
13.
Особенности группирования разных типовэндокринных клеток
•Эндокринные клетки не эндокринных органов
(энтероциты ЖКТ, эндокринные нейроны
гипоталамуса)
•Эндокринные клетки желез смешанной секреции
(семенников, яичников, поджелудочной железы)
•Эндокринные клетки эндокринных желез
(надпочечников, щитовидной железы, паращитовидных
желез, аденогипофиза, эпифиза)
13
14. Гормоны, секретируемые эффекторными типами клеток
Клетки печени– ИФР-1
– Ангиотензиноген (прогормон)
Клетки почек
– Эритропоэтин
Клетки жировой ткани
– Лептин
– Адипонектин
Кардиомиоциты
– Атриопептиды
14
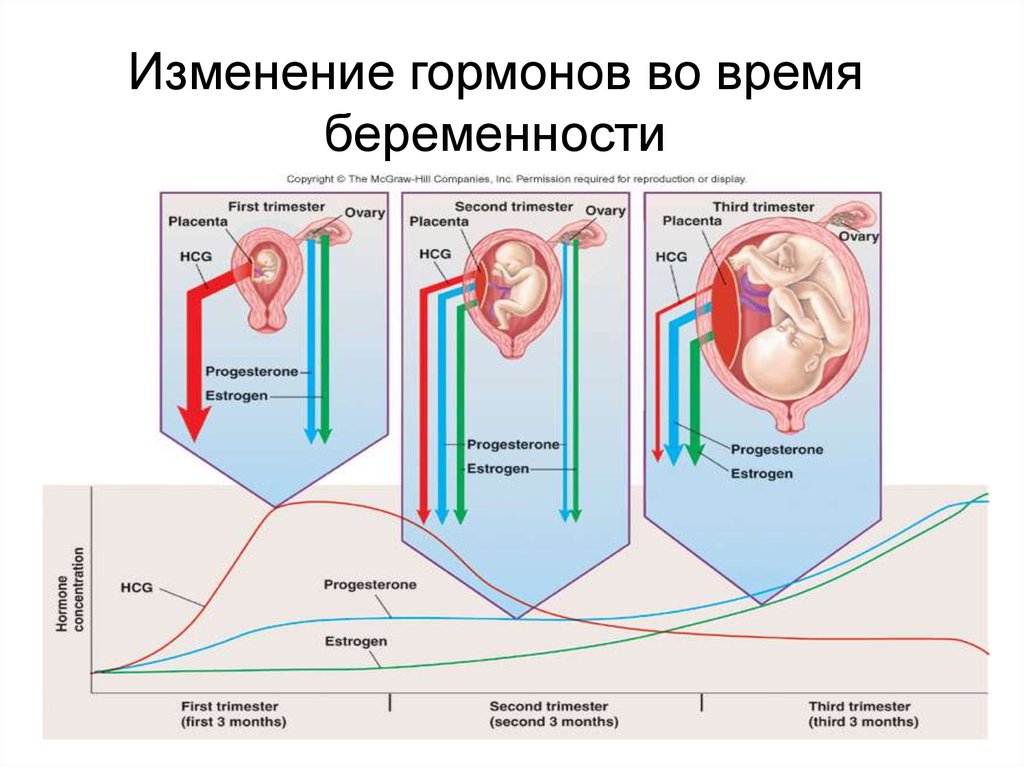
15.
• Эндокринной активностью обладаеттакже плацента.
• На разных этапах беременности её
значение и активность существенно
меняются.
15
16.
Изменение гормонов во времябеременности
17.
Гормональные изменения во времябеременности
18.
Биологический смысл интеграцииэндокринных клеток в эндокринную
железу –
создание более совершенного
механизма управления потоками
информации и эффекторными
функциями
18
19.
• Существуют три основных класса гормонов похимическому строению
1. Белки и полипептиды, включая гормоны,
секретируемые передней и задней долями гипофиза,
поджелудочной железой (инсулин, глюкагон),
околощитовидной железой (паратгормон) СТГ, ФСГ,
ЛГ, Пролактин.
• 2. Стероиды, секретируемые корой надпочечников
(кортизол и альдостерон), яичниками (эстрогены,
прогестерон), семенниками (тестостерон), плацентой
(эстрогены, прогестерон).
• 3. Производные аминокислоты тирозина,
секретируемые щитовидной железой (тироксин,
трийодтиронин) и мозговым веществом надпочечников
(адреналин и норадреналин).
• 4.Производные арахидоновой кислоты
19
20. По свойствам и механизму действия все гормоны можно разделить на 2 группы
1) гидрофильные (растворяющиеся в воде) – этопептиды и гормоны мозгового слоя
надпочечников. Срок их действия – от секунд-минут
до 1-2 часов. Они не могут пройти через мембрану
клетки, поэтому действуют на рецепторы мембраны,
которые активизируют систему вторичных
посредников.
2) гидрофобные – это гормоны щитовидной железы и
стероиды. Действуют дольше, чем гидрофильные –
до нескольких дней. Проходят через мембрану
внутрь клетки и регулируют там процессы
транскрипции и стабильность иРНК (например,
трийодтиронин проникает в ядро и регулирует
транскрипцию).
20
21. Гормональные эффекты
• срочные реакции - изменение проницаемостимембраны для ионов или глюкозы и аминокислот, что
приводит, например, к сокращению гладких мышц или
повышению темпа метаболизма клетки,
• отсроченные реакции, которые заключаются в
изменении активности уже существующих ферментов,
и в дополнение к темпу метаболизма может измениться
его направление (например, запасать глюкозу или
расходовать),
• длительные реакции - синтез новых ферментов и
структурных компонентов клетки - такие реакции
способны изменить и структуру, и функцию органа или
системы органов. (Кроссадаптация, формирование
функциональных систем, структурный след адаптации)
21
22.
Общие свойства гормонов:Строгая специфичность действия.
Высокая биологическая активность.
Дистантный характер действия.
Многие гормоны не имеют видовой
специфичности(инсулин)
5. Генерализованность действия.
6. Пролонгированность эффекта
1.
2.
3.
4.
22
23.
Этапы гуморальной регуляцииМесто выработки
Стимул для выделения
Транспорт
Рецепция и клеточные механизмы
действия
5. Физиологические эффекты
6. Метаболизм и выведение
1.
2.
3.
4.
23
24. Место выработки
• Эндокринная железа• Неэндокринные органы
• Одно или несколько мест выработки
24
25. Стимул для выделения
1.Нервный импульс2. Влияние другого гормона
3. Конкретное вещество, уровень
которого регулируется гормоном
4. Непосредственное раздражение
рецепторов
5. Повреждение ткани
25
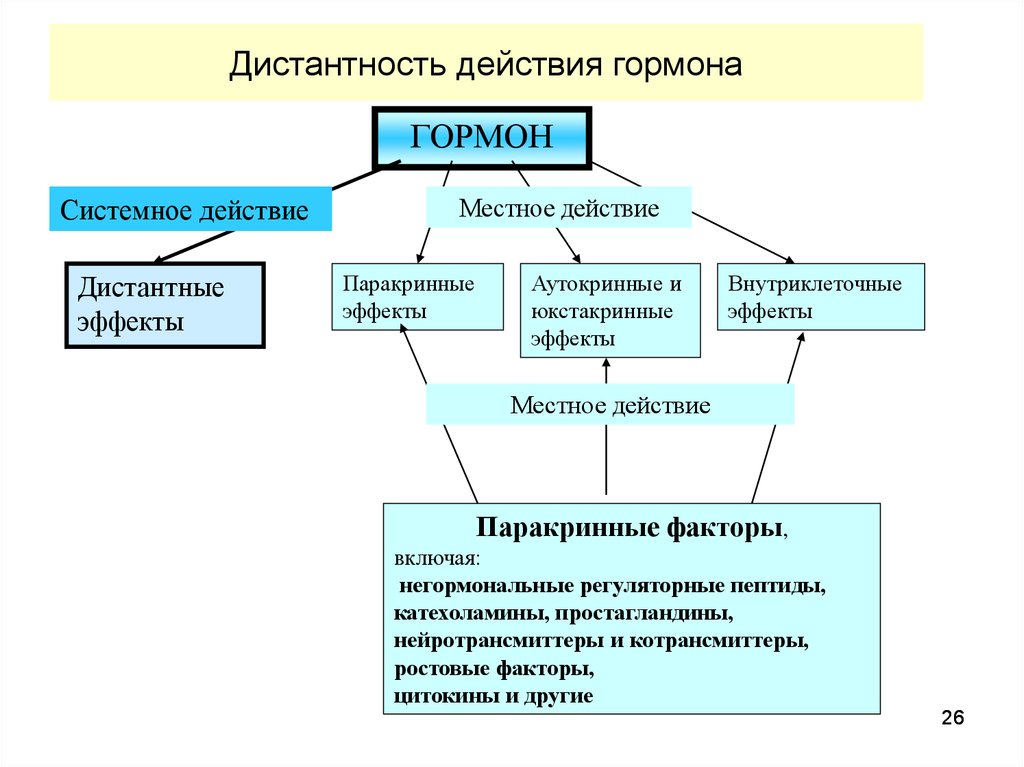
26.
Дистантность действия гормонаГОРМОН
Системное действие
Дистантные
эффекты
Местное действие
Паракринные
эффекты
Аутокринные и
юкстакринные
эффекты
Внутриклеточные
эффекты
Местное действие
Паракринные факторы,
включая:
негормональные регуляторные пептиды,
катехоламины, простагландины,
нейротрансмиттеры и котрансмиттеры,
ростовые факторы,
цитокины и другие
26
27.
Типы местного контроляВнутриклеточный
контроль
Региональный контроль
Аутокринный
Юкстакринный
С
Р
С
Паракринный
С
Р
Р Р Р
Р
С
Э
Э
Э Э Э
Э
27
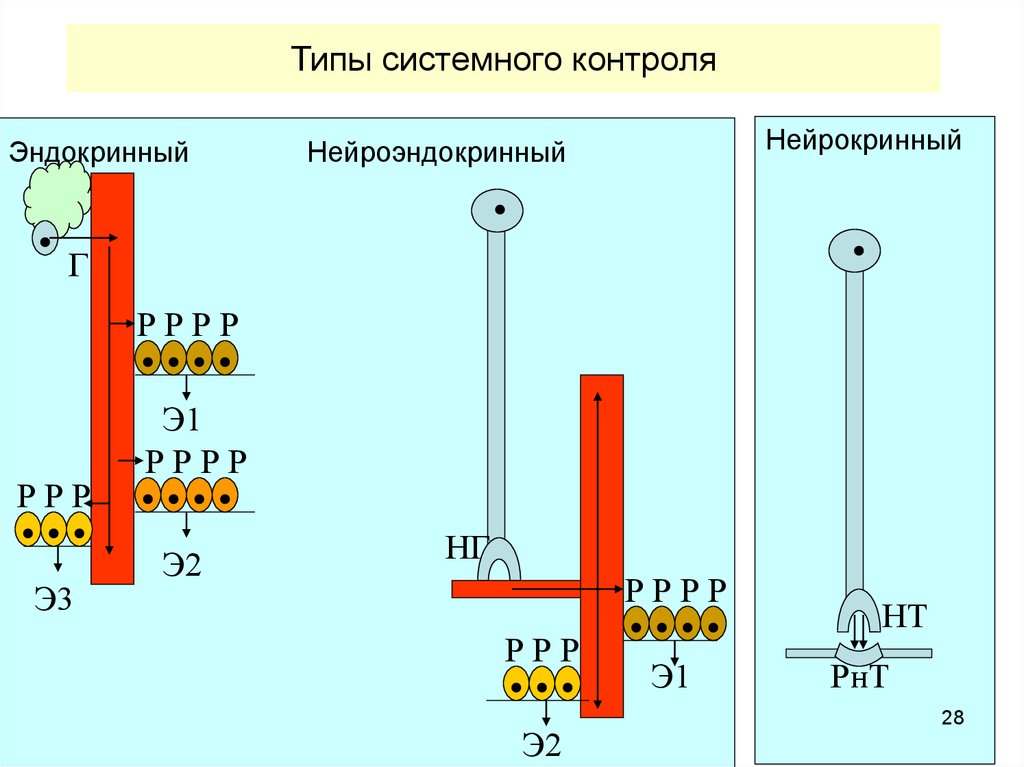
28.
Типы системного контроляЭндокринный
Нейрокринный
Нейроэндокринный
Г
РРРР
РРР
Э3
Э1
РРРР
Э2
НГ
РРРР
РРР
Э2
Э1
НТ
РнТ
28
29.
Транспорт1. Транспорта нет – паракринный эффект
2. Транспорт со специальным белком
3. Транспорт в неактивной или
малоактивной форме
4. Транспорт в свободном виде
29
30. Транспорт
• Водорастворимые гормоны раствореныв плазме.
• Стероидные, тиреоидные в связи со
специфическими белками
переносчиками, но около 10% в
свободном виде.
30
31.
Метаболизм гормонов1. Метаболизм в тканях с изменением
активности
2. Связывание в тканях
3. Разрушение в лизосомах
4. Катаболизм в печени и почках
5. Экскреция с желчью
Можно измерить количество продуктов
распада и оценить концентрацию гормона
31
32. Рецепция, клеточные и молекулярные механизмы действия
3233. Орган или ткань мишень
• Один• Несколько
• Ответ тканей-мишеней формируется
при образовании гормон-рецепторного
комплекса.
33
34.
Регуляция рецепцииКоличество рецепторов
Чувствительность рецепторов
Индуцибельность рецепторов
Типы рецепторов (мембранные,
цитоплазматические, ядерные)
34
35. Механизмы рецепции стероидных гормонов
3536.
• Стероидные соединения плохо растворяются вводе и хорошо в органических растворителях и
растительных маслах.
• Благодаря липофильности они легко
проникают через плазматические
липопротеидные мембраны и поэтому могут
свободно входить внутрь реагирующих клеток.
36
37. Рецепция и метаболизм жирорастворимых гормонов
3738. Механизмы рецепции гормонов белковой природы
3839. Клеточные рецепторы делятся на следующие классы
мембранные
рецепторы, сопряжённые с G-белками
рецепторные тирозинкиназы
ионные каналы.
цитоплазматические
ядерные
39
40. Значение вторичных посредников для рецепции белковых типов гормонов
• Эффективность действия малых доз гормонов• Универсальный ответ клетки при действии
различных гормонов белковой природы
• Вторичные посредники:цАМФ, цГМФ,
инозитолтрифосфат,диацилглицерин или ионы
кальция.
• Активация посредников приводит к выходу
ионов кальция и стимуляции протеинкиназы С.
40
41. Вторичные посредники
4142.
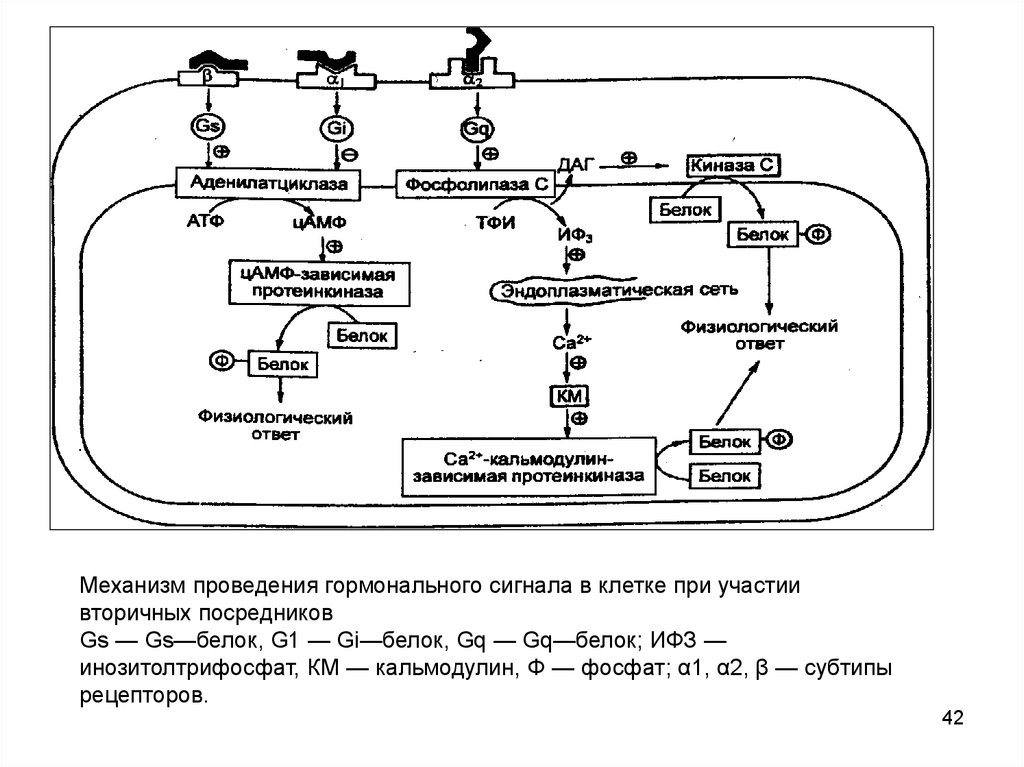
Механизм проведения гормонального сигнала в клетке при участиивторичных посредников
Gs — Gs—белок, G1 — Gi—белок, Gq — Gq—белок; ИФЗ —
инозитолтрифосфат, КМ — кальмодулин, Ф — фосфат; α1, α2, β — субтипы
рецепторов.
42
43.
• Вторичные посредники (англ. secondmessenger) — это низкомолекулярные
вещества, которые образуются или
высвобождаются в результате ферментативной
активности одного из компонентов цепи
передачи сигнала и способствуют его
дальнейшей передаче и амплификации.
• Вторичные посредники характеризуются
следующими свойствами: имеют небольшую
молекулярную массу и с высокой скоростью
диффундируют в цитоплазме; быстро
расщепляются и быстро удаляются из
цитоплазмы.
43
44. Вторичные посредники
• ионы кальция (Ca2+);• циклический аденозинмонофосфат (цАМФ) и
циклический гуанозинмонофосфат (цГМФ).
• инозитолтрифосфат
• липофильные молекулы (например,
диацилглицерол);
• оксид азота (NO) (эта молекула выступает и в
роли первичного посредника, проникающего
в клетку извне).
• Молекула СО
• Активные формы кислорода
44
45. Классификация вторичных посредников по растворимости в воде и размеру молекулы
• Гидрофобные молекулы — нерастворимые в водемолекулы, например, диацилглицерол,
инозитолтрифосфат, фосфатидилинозитолы —
связаны с мембраной и могут диссоциировать в
околомембранное пространство;
• Гидрофильные молекулы — водорастворимые
молекулы и ионы, например, цАМФ, цГМФ, Ca2+ —
находятся в цитозоле;
• Газы — оксид азота (NO), оксид углерода (CO),
АФК — могут переходить из цитозоля в
межклеточную среду через клеточные мембраны.
45
46. Аденилатциклазная система
4647. Аденилатциклазная система рассматривается на примере действия адреналина на клетки печени.
• Адреналин вызывает в организме эффект,называемый «fight or flight» (бой или
бегство) — усиливается тонус мышц,
увеличивается частота сердечных
сокращений. Для мобилизации организма
требуется повышение концентрации глюкозы
в крови.
• Связывание адреналина с рецепторами на
поверхности клеток печени запускает распад
гликогена, запасенного в клетках печени и
высвобождение глюкозы..
47
48. Особенность этой системы передачи сигнала в клетке
• Сигнал на большинстве этаповусиливается, например,
активированная аденилатциклаза
синтезирует множество молекул цАМФ.
• В результате взаимодействия одной
молекулы адреналина с рецептором в
плазматической мембране клетки
печени, в кровь выводится около
10 миллионов молекул глюкозы.
48
49.
• Таким образом, даже относительнослабый стимул может вызывать
значительный ответ.
• Это явление называется амплификацией
сигнала
50. Роль кальция как вторичного посредника. Кальмодулин. Кальций-кальмодулин-зависимые киназы.
Роль кальция как вторичногопосредника. Кальмодулин.
Кальций-кальмодулинзависимые киназы.
51.
• Выделяют шесть типовпотенциалзависимых кальциевых
каналов.
• Наиболее важными в сердечнососудистой системе являются L- и Ттипы.
• Каналы Т- и L-типов обнаружены в
миокарде, гладкой мускулатуре сосудов.
• L-каналы инактивируются медленно,
через них в клетку проникает большая
часть внеклеточного кальция.
• L-каналы чувствительны к действию
БКК.(Блокаторы кальциевых каналов)
51
52. Два типа передачи сигнала при посредстве ионов кальция (1)
• Первый из них осуществляется вэлектро-возбудимых, преимущественно
нервных клетках.
• В них деполяризация плазматической
мембраны вызывает поглощение
нервным окончанием кальция через
потенциал-зависимые кальциевые
каналы что приводит к секреции
нейромедиатора
53. Два типа передачи сигнала при посредстве ионов кальция (2)
• Второй способ передачи сигнала припосредстве ионов кальция осуществляется
практически во всех типах эукариотических
клеток.
• Сигнальная молекула связывается с
рецептором на поверхности клетки, что
приводит к синтезу вторичных посредников ,
высвобождению ионов кальция из
внутриклеточных депо , активации
эффекторных ферментов и запуску кальцийопосредованных внутриклеточных реакций.
54.
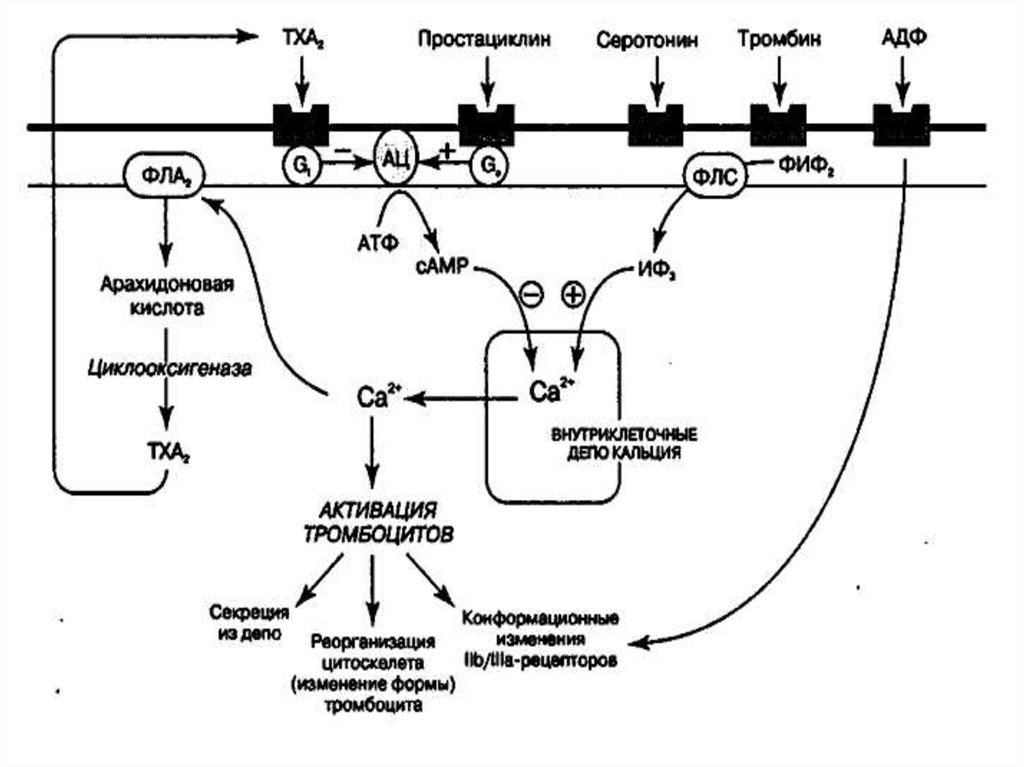
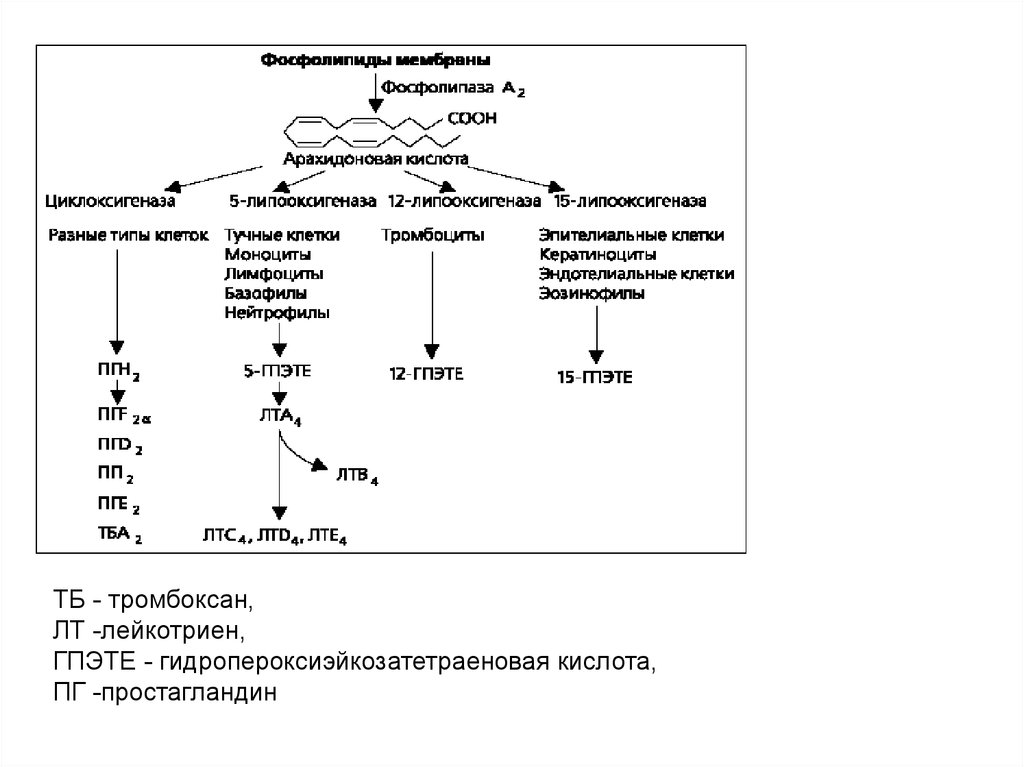
5455. Производные арахидоновой кислоты
5556. Арахидоновая кислота
• Существует два основных пути метаболизмаарахидоновой кислоты - циклоксигеназный и
липоксигеназный.
• Циклоксигеназный путь метаболизма арахидоновой
кислоты приводит к образованию простагландинов и
тромбоксана A2.
• Липоксигеназный - к образованию лейкотриенов.
• В тучных клетках легких синтезируются как
простагландины, так и лейкотриены, в базофилах только лейкотриены.
• Образующиеся в незначительном количестве 12гидропероксиэйкозотетраеновая кислота и 15гидропероксиэйкозотетраеновая кислота играют
важную роль в воспалении .
57.
ТБ - тромбоксан,ЛТ -лейкотриен,
ГПЭТЕ - гидропероксиэйкозатетраеновая кислота,
ПГ -простагландин
58. Роль простагландинов:
ТипМесто синтеза
Действие
ПГ Д2
Тучные клетки
Вазодилатация
Ингибирование агрегации
ТБЦ и лейкоцитов.
Уменьшение пролиферации
Т-клеток.
ПГ Е2
Сердце
Вазодилатация
Агрегация ТБЦ
Уменьшение пролиферации
Т-клеток
ПГ F2α
Сердце
Вазоконстрикция
ПГ Н2
Сердце
Вазоконстрикция
Агрегация ТБЦ
ПГ I2
Эндотелий
Вазодилатация
Ингибирование ТБЦ,
агрегация лейкоцитов
59. Тирозинкиназные рецепторы
60. Рецепторы с тирозинкиназной активностью
• Тирозиновые протеинкиназы - ферменты,фосфорилирующие специфические белки по
тирозину, подразделяют на 2 типа - мембранные
(рецепторные) и цитоплазматические.
• Внутриклеточные тирозиновые протеинкиназы
принимают участие в процессах передачи сигнала в
ядро.
• Рецепторные тирозиновые протеинкиназы участвуют
в трансмембранной передаче сигналов.
• Примером рецепторной тирозиновой протеинкиназы
может служить рецептор инсулина
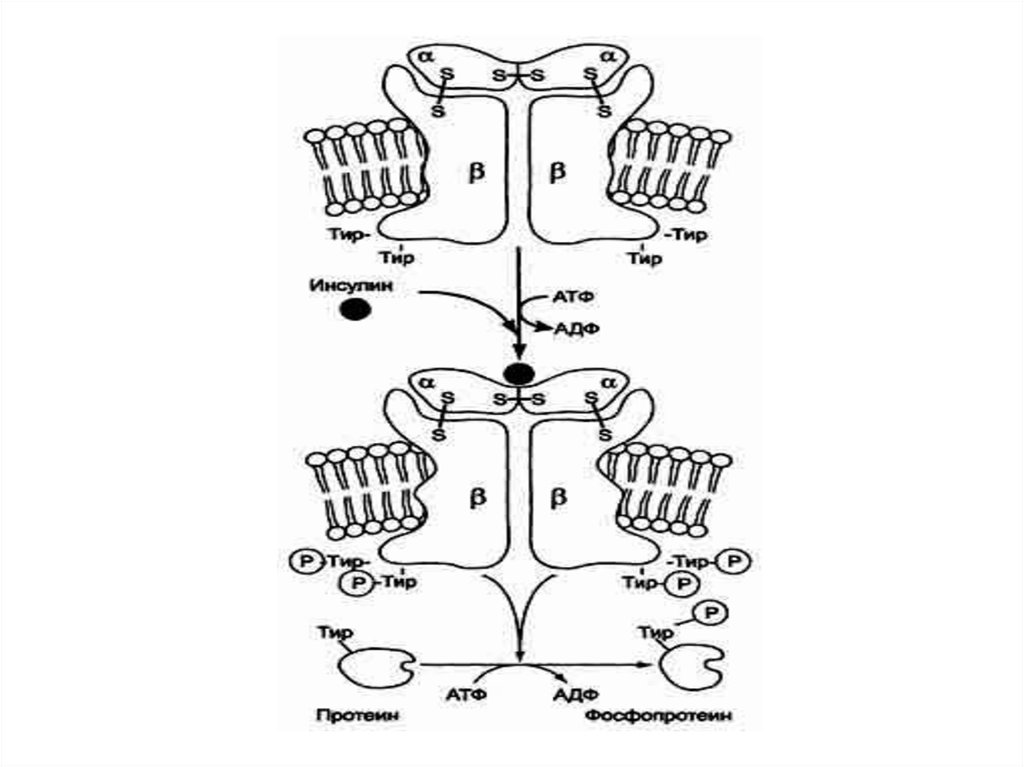
61.
• Рецептор состоит из двух α- и двух βсубъединиц, связанных дисульфиднымисвязями и нековалентными
взаимодействиями.
• α- и β-Субъединицы - гликопротеины с
углеводной частью на наружной стороне
мембраны.
• Вне мембраны на её поверхности находятся
α-субъединицы.
• Центр связывания инсулина образован Nконцевыми доменами α-субъединиц.
62.
63.
• Каталитический центр тирозиновой протеинкиназынаходится на внутриклеточных доменах β-субъединиц.
• В отсутствие гормона инсулиновые рецепторы не
проявляют тирозинкиназной активности.
• Присоединение инсулина к центру связывания на αсубъединицах активирует фермент, причём субстратом
служит сама тирозиновая протеинкиназа т.е. происходит
фосфорилирование β-субъединицы.
• Фосфорилирование β-субъединиц происходит по
механизму межмолекулярного трансфосфорилирования,
т.е. одна β-цепь фосфорилирует другую β-цепь той же
молекулы рецептора.
• Это, в свою очередь, приводит к изменению субстратной
специфичности тирозиновой протеинкиназы; теперь она
способна фосфорилировать другие внутриклеточные
белки.
• Активация и изменение специфичности обусловлены
конформационными изменениями рецептора инсулина
после связывания гормона и аутофосфорилирования.