")




























 Биология
БиологияПохожие презентации:




")





Взаємозв’язок процесів обміну речовин в організмі (інтеграція метаболізму)
1. ВЗАЄМОЗВ’ЯЗОК ПРОЦЕСІВ ОБМІНУ РЕЧОВИН В ОРГАНІЗМІ (ІНТЕГРАЦІЯ МЕТАБОЛІЗМУ)
2.
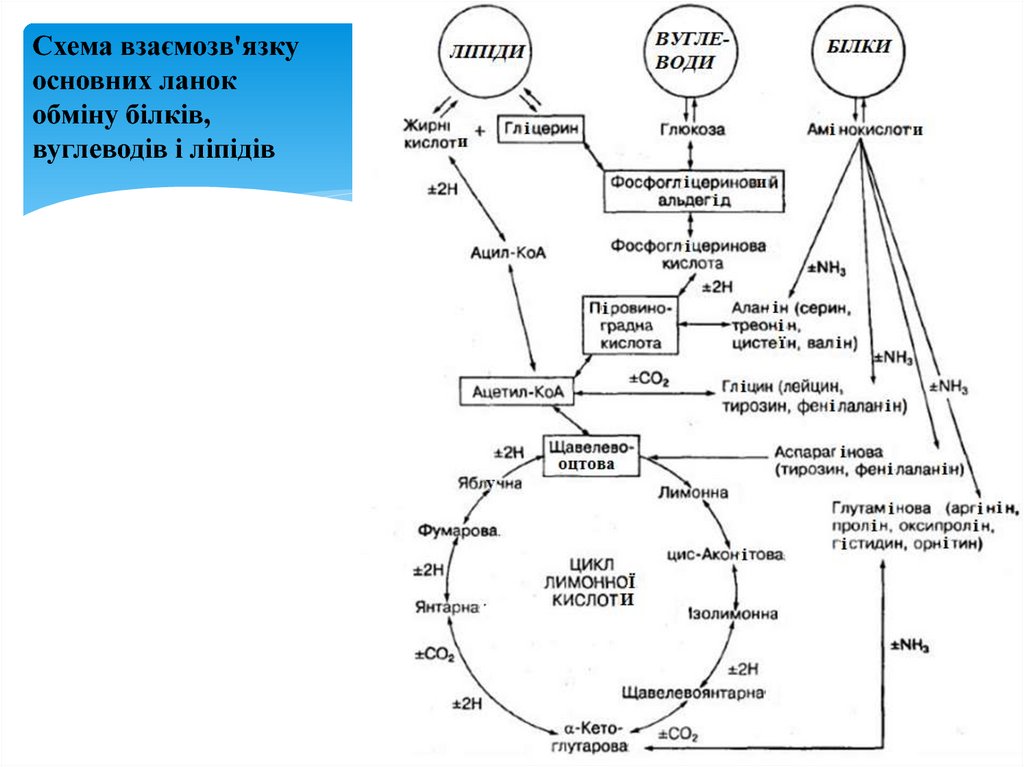
Схема взаємозв'язкуосновних ланок
обміну білків,
вуглеводів і ліпідів
3. Взаємозв'язок між обміном вуглеводів і ліпідів
Взаємозв'язокобміну
вуглеводів
і
ліпідів
розкривається в двох напрямках:
• перетворення вуглеводів у жири (головне значення);
• перетворення жирів у вуглеводи (обмежене
значення).
4.
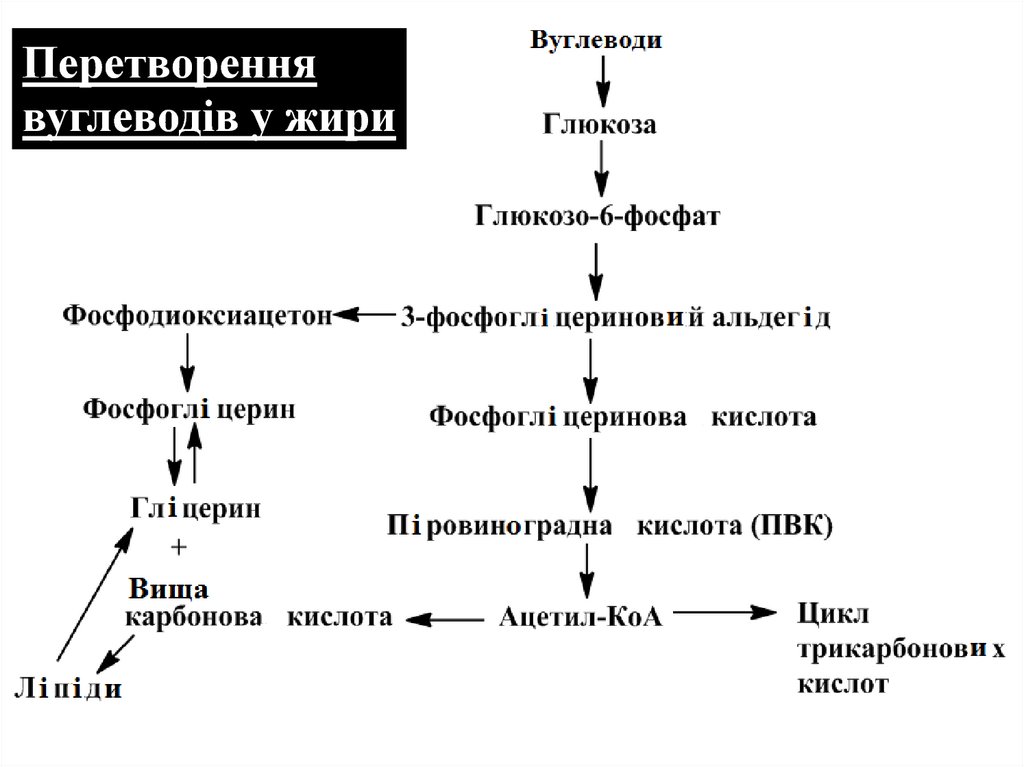
Перетвореннявуглеводів у жири
5. Перетворення вуглеводів у жири
При згодовуванні тваринам кормів, які містять великукількість вуглеводів (кукурудза, картопля та ін.), у них
відкладається жир.
Утворення гліцерину як складової молекули жиру
відбувається в результаті відновлення фосфодиоксиацетону,
який є загальним проміжним продуктом обміну вуглеводів і
ліпідів. Фосфогліцерин, що утворився, є активною формою
гліцерину і в подальшому може взаємодіяти з активними
формами вищих жирних кислот (ацилами). Таким шляхом
синтезуються жири і складні ліпіди (гліцерофосфоліпіди).
6.
Биосинтез высших жирных кислот происходит изактивных остатков уксусной кислоты, образующейся при
окислительном
декарбоксилировании
пировиноградной
кислоты, т.е. центрального промежуточного продукта обмена
углеводов.
Углеводы являются также важным источником атомов
водорода, необходимых для биосинтеза высших жирных кислот
из уксусной кислоты. В состав природных жиров входят
преимущественно
высшие
одноосновные
кислоты
(стеариновая, пальмитиновая и др.), которые содержат в своей
молекуле значительно больше атомов водорода, чем углеводы.
Для синтеза высших жирных кислот используются атомы
водорода восстановленных форм кофермента НАДФ, которые
образуются за счет расщепления углеводов в пентозофосфатном
пути.
7. Превращение жиров в углеводы
Образование углеводов из других продуктов (белков,жиров) называется глюконеогенезом или гликонеогенезом.
При β-окислении высших жирных кислот образуется
ацетил-КоА. Эта же кислота возникает при окислительном
декарбоксилировании пировиноградной кислоты. Именно
ацетил-КоА составляет важную молекулярную основу
превращения углеводов в липиды и липидов в углеводы.
Небольшое
количество
углеводов
может
синтезироваться из глицерина, окислением его в
фосфодиоксиацетон и фосфоглицериновый альдегид,
которые являются промежуточными продуктами гликолиза и
пентозофосфатного пути окисления углеводов.
8.
Глюконеогенез имеет важное значение дляжвачных животных. У них в рубце под влиянием
бактериальной
микрофлоры
сбраживаются
моносахариды с образованием летучих карбоновых
кислот, которые поступают в кровь. У жвачных в
печени активно протекает глюконеогенез из
пропионовой кислоты, образующейся в рубце, по
следующей схеме:
Пропионат → сукцинат → малат → глюкоза → гликоген
Пропионовая кислота образуется также при
катаболизме валина и изолейцина и при конверсии
холестерина в желчные кислоты.
9. НАРУШЕНИЯ УГЛЕВОДНОГО ОБМЕНА
Выраженныенарушения
углеводного
обмена
наблюдаются, в частности, при кетозах у крупного
рогатого скота. При этом отмечается снижение
концентрации глюкозы в крови (гипогликемия) с
повышением
концентрации
кетоновых
тел
(гиперкетонемия). Гипогликемия отмечается также и при
родильном парезе, атониях преджелудков у коров.
10.
Гипогликемия наблюдается при болезнях печени, пригликогенной болезни, когда клетки печени теряют
способность расщеплять гликоген с освобождением
глюкозы. Низкий уровень глюкозы в крови имеет место
при
гипокортицизме,
когда
снижен
синтез
глюкокортикоидов коркового слоя надпочечников, а также
при опухолях островковых клеток поджелудочной
железы, вырабатывающих инсулин, при передозировках
противодиабетических препаратов, голодании, при
поражении почек, когда глюкоза выделяется с мочой.
11.
Повышенное содержание глюкозы в крови(гипергликемия) часто указывает на наличие
сахарного диабета. При сахарном диабете
нарушается поступление глюкозы в клетки, что и
приводит к гипергликемии. Причиной этого является
или неспособность поджелудочной железы выделять
инсулин в кровь, или же неспособность инсулина
связываться с соответствующими рецепторами и
оказывать свое биологическое действие (перенос
глюкозы
в
клетки
и
последующее
ее
фосфорилирование).
12.
Нарушения углеводного обмена при диабете приводят красстройствам жирового и белкового обменов. Жирные
кислоты быстро поступают в митохондрии, так как
карнитинацилтрансфераза является высоко активной.
При этом происходит накопление молекул ацетил-КоА в
результате β-окисления жирных кислот. Большинство
молекул ацетил-КоА не может поступать в ЦТК, так как
проявляется
недостаточность
щавелево-уксусной
кислоты, а идут в реакции конденсации (синтеза
кетоновых тел).
Большинство
кислых
продуктов
нормального
метаболизма в виде СО2 быстро экскретируется легкими.
Кетоновые тела не экскретируются легкими, и при их
высокой концентрации они нарушают кислотнощелочное равновесие в почках, что приводит к развитию
комы, ацидоза (при инсулин-зависимом сахарном
диабете), ацетонурии.
13.
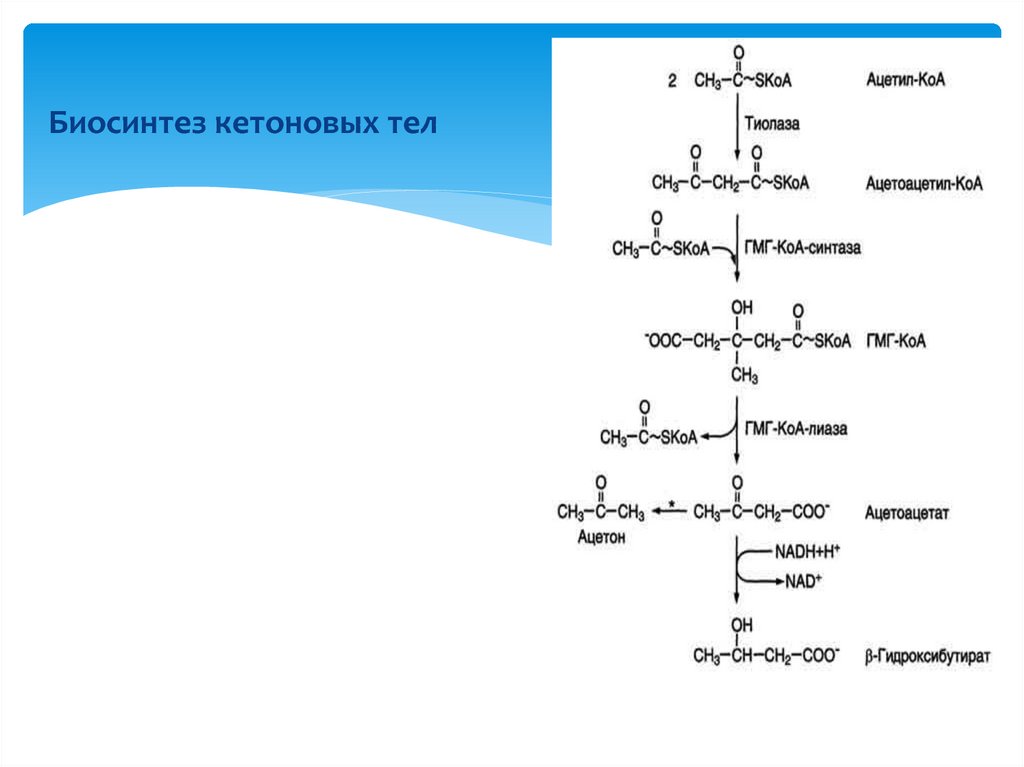
Биосинтез кетоновых тел14. Взаимосвязь между обменов углеводов и белков
Взаимосвязьмежду
обменом углеводов
и
белков
(аминокислот) включает две главные закономерности:
превращение белков в углеводы (основное значение в
организме);
превращение углеводов в белки (ограниченное значение).
15. Превращение белков в углеводы
Аминокислоты после дезаминирования и трансаминированияпревращаются в ЩУК, α-КГ кислоту и ПВК. Из них ПВК
является главным соединением превращения аминокислот в
углеводы. Подвергаясь окислительному декарбоксилированию
она превращается а ацетил-КоА, который является общим
продуктом обмена углеводов, жиров и белков. Ацетил-КоА
вступает в реакцию со ЩУК и окисляется в ЦТК до СО2 и
Н2О. В процессе окисления синтезируются α-КГ и ЩУК,
являющиеся общими продуктами обмена белков и углеводов.
Обе они после декарбоксилирования (однократного или
повторного) могут превращаться в ПВК, которая является
источником для синтеза углеводов.
16.
В этом процессе главную роль играют теаминокислоты, которые после дезаминирования
могут легко превращаться в ПВК или близкие к
ней соединения (например,
фосфоенолпировиноградную кислоту). К таким
«гликогенным» аминокислотам относят: аланин,
серин, аспарагиновая и глутаминовая кислоты,
цистеин, глицин, гистидин, метионин, аргинин,
пролин, триптофан, валин, треонин.
Менее интенсивно превращаются в углеводы
фенилаланин, тирозин, изолейцин.
17. Специфические пути катаболизма аминокислот
18. Специфические пути катаболизма аминокислот
CH2SH
CH
CH
NH
NH22
CH2
COOH
CH2
SH
COOH
Цистеин
Цистеин
Цистеїн
CH2
Н
Н22S
S +
+ C
C NH
NH22
ЦистеинЦистеинЦистеїндесульфогидраза
десульфогидраза
десульфогідраза
COOH
COOH
CH3
CH3
CH3
C
C
CH3
Н22О
О
NH ++ Н
NH
COOH
COOH
+ NH
C O
C
O + NH33
COOH
COOH
19. Гликогенные и кетогенные амнокислоты
ГликогенныеАланин
Аргинин
Аспарагиновая
кислота
Цистеин
Глутаминовая
кислота
Глицин
Аспарагин
Кетогенные Гликогенные
и кетогенные
Гистидин Лейцин
Метионин
Пролин
Серин
Треонин
Глутамин
Валин
Изолейцин
Лизин
Фенилаланин
Тирозин
Триптофан
20.
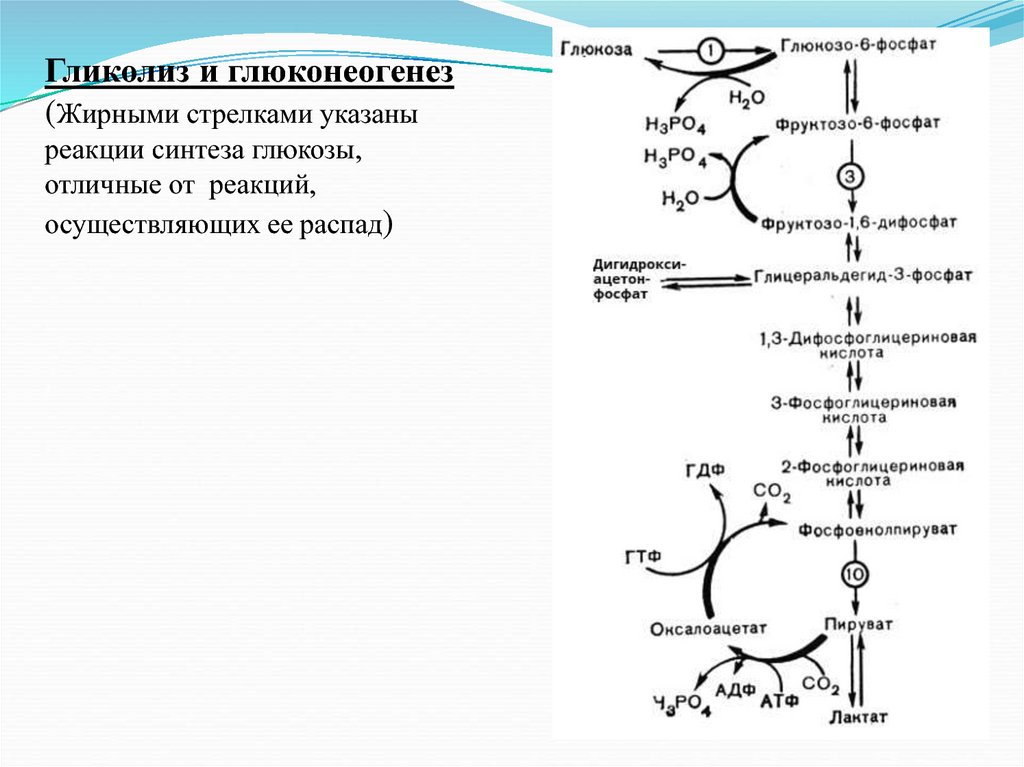
Гликолиз и глюконеогенез(Жирными стрелками указаны
реакции синтеза глюкозы,
отличные от реакций,
осуществляющих ее распад)
21.
Глюконеогенез является одним из важных процессовобеспечения
организма
углеводами.
Значение
глюконеогенеза возрастает при физических нагрузках,
при резком снижении температуры окружающей среды,
при реакциях организма на травму и др. В подобных
ситуациях резервы гликогена быстро расходуются и
возрастает роль глюконеогенеза.
Глюконеогенез активно протекает в печени и почках. На
каждые 100 г белка в среднем может синтезироваться
около 50 г углеводов. Этот процент возрастает при
некоторых заболеваниях, в частности, при сахарном
диабете до 70-75 % белков, могут превращаться в
углеводы.
22. Превращение углеводов в белки
Углеводы являются основнымматериалом,
из которого
в
организме образуется ПВК, а также ЩУК и α-КГ кислота.
Эти кетокислоты путем аминирования, преимущественно за
счет аминосахаров и их производных, превращаются в
соответствующие аминокислоты: аланин, аспарагиновую и
глутаминовую. Из глутаминовой кислоты изменением
структуры ее углеродной цепи также может синтезироваться
ряд аминокислот: пролин, гидроксипролин, орнитин.
Углеводы также являются источником атомов водорода для
синтеза
аминокислот
за
счет
атомов
водорода
восстановленной формы НАДФ, образующейся при
окислении углеводов в пентозофосфатном пути.
В организме животных синтез аминокислот (белков) за
счет углеводов ограничен и не имеет существенного
биологического значения.
23. Взаимосвязь между обменом углеводов и нуклеиновых кислот
Пентозофосфатный путь окисления являетсяисточником пентозофосфатов: рибулозо-5-
фосфата и рибозо-5-фосфата, используемых в
организме для синтеза нуклеотидов и
нуклеиновых кислот. Синтез
нуклеотиддифосфатов (УДФ, АДФ и др.) и
трифосфатов (УТФ, ГТФ, АТФ и др.) зависит от
интенсивности окисления углеводов в клетке,
сопряженного с окислительным
фосфорилированием в дыхательной цепи.
24.
При расщеплении нуклеиновых кислот инуклеотидов
освобождается
определенное
количество пентоз, которые могут включаться в
разные звенья обмена, в том числе превращаться в
гексозы, в частности, в глюкозу. Глюкоза
используется
для
синтеза
гликогена,
мукополисахаридов, лактозы и др. веществ.
В пентозофосфатном пути образуется большая
часть восстановленных форм НАДФ, которые
являются необходимыми донорами водорода для
синтеза различных веществ.
25. Взаимосвязь между обменом белков и липидов
Взаимосвязь между обменом белков (аминокислот) илипидов включает две главные закономерности:
превращение белков в липиды (основное значение в
организме);
превращение липидов в белки (ограниченное
значение).
26. Превращение белков в липиды
Преобладание в рационе животных кормов, содержащихпреимущественно белки, вызывает активный синтез
жиров.
При дезаминировании некоторых аминокислот (аланина,
серина) образуется ПВК, которая декарбоксилируется с
образованием ацетил-КоА, который образуется также из
жиров. Из молекул ацетил-КоА могут синтезироваться
высшие жирные кислоты. Кроме того ПВК и ацетил-КоА
могут образовываться из аспарагиновой и глутаминовой
кислот
после
их
дезаминирования
и
декарбоксилирования.
27.
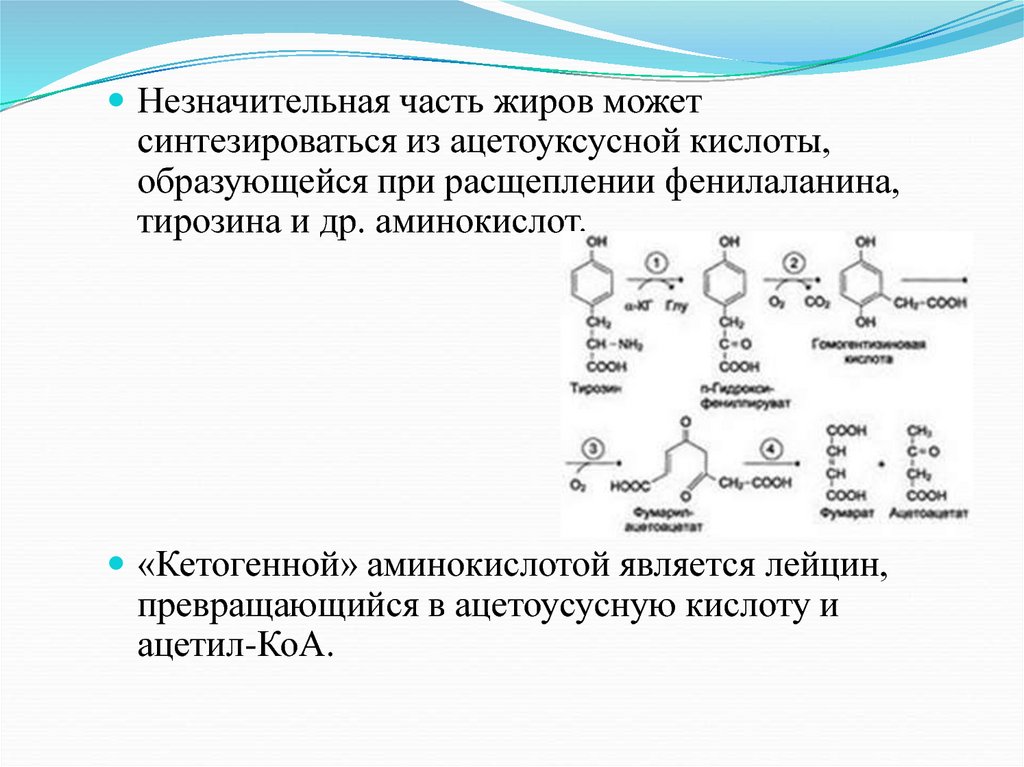
Незначительная часть жиров можетсинтезироваться из ацетоуксусной кислоты,
образующейся при расщеплении фенилаланина,
тирозина и др. аминокислот.
«Кетогенной» аминокислотой является лейцин,
превращающийся в ацетоусусную кислоту и
ацетил-КоА.
28.
Глицерин синтезируется из аланина.Азотистая часть сложных липидов образуется за
счет серина, холина и коламина.
29. Превращение жиров в белки
Этот процесс ограничен, т.к. основным источникомазотсодержащих веществ у животных являются только
белки. Незаменимые аминокислоты вообще не могут
синтезироваться у высших организмов. Заменить белки в
диете углеводами или липидами нельзя, потому что это
вызовет белковое голодание. Частичное образование
заменимых аминокислот из жиров возможно из ПВК,
ЩУК, α-КГ кислоты, т.е. тех кислот, которые легко
дезаминируются и подвергаются трансаминированию.
Для биосинтеза аминокислот из кетокислот необходим
аммиак, а он освобождается при обмене белков.
Источником
аммиака
также
могут
быть
глицеринфосфолипиды, содержащие в своем составе
серин, коламин и холин.
30.
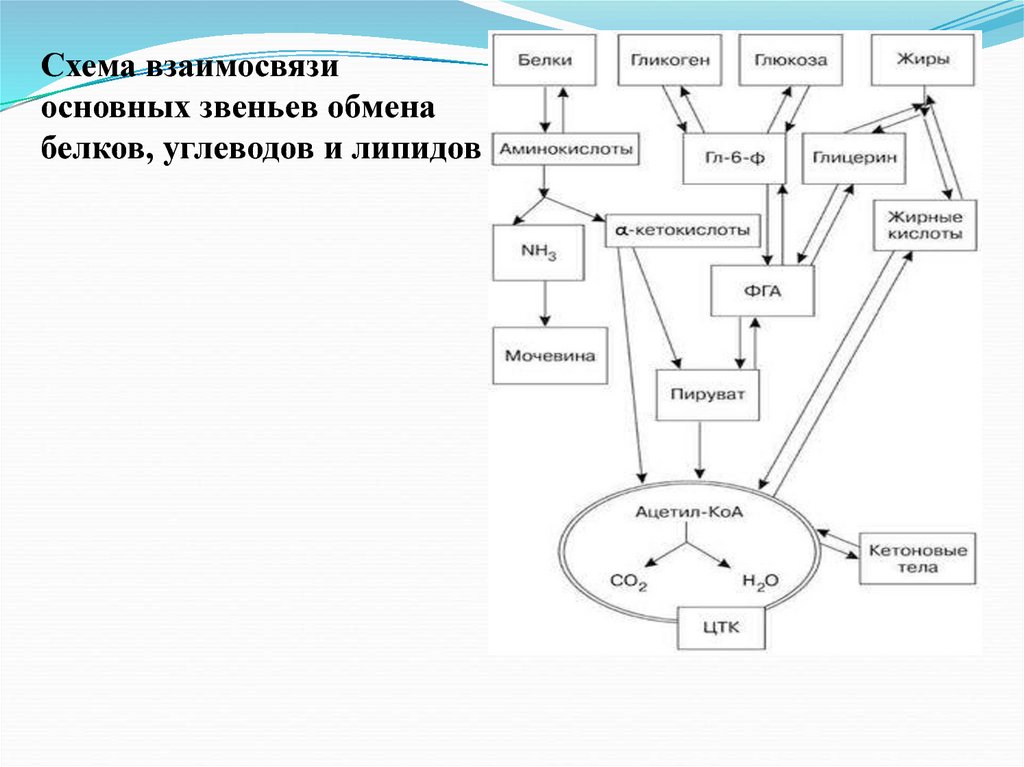
Схема взаимосвязиосновных звеньев обмена
белков, углеводов и липидов
31. Взаимосвязь между обменом органических и неорганических веществ
Обменорганических и неорганических веществ
неразрывно связаны между собой. Все биохимические
процессы протекают в водных растворах. Вода является
той дисперсной системой, которая обусловливает
химические и физико-химические процессы в живом
организме. Без воды не возможен обмен веществ,
поскольку многие реакции обмена (процессы
гидролиза, окисления, биосинтеза жирных кислот)
осуществляются путем гидратации, дегидратации,
гидрирования и т.д. Большая часть СО2, выделяющегося
из организма, образуется также за счет кислорода воды.
32.
Без фосфорной кислоты не может происходитьобмен углеводов и образовываться АТФ.
Без
железа
не
синтезируются
молекулы
гемоглобина, миоглобина, цитохромов и др.
соединений.
Многие ферменты проявляют свою активность при
наличии определенных ионов металлов. Тромбин не
катализирует превращение фибриногена в фибрин
при отсутствии ионов Са2+. Карбоангидраза не
расщепляет угольную кислоту на Н2О и СО2 при
отсутствии ионов Zn2+. Различные АТФ-азы не
расщепляют АТФ, если нет ионов К+, Na+, Са2+ и
Mg2+.
33. Катаболизм и анаболизм: взаимосвязь и особенности
Связь между катаболизмом и анаболизмом проявляется натрех уровнях – источников углерода, энергетическом и
восстановительных реакциях анаболизма.
На уровне источников углерода: Промежуточные продукты
центральных путей катаболизма становятся субстратами для
анаболических реакций, в процессе которых образуются
структурные блоки, необходимые для синтеза макромолекул.
На энергетическом уровне: В процессе катаболизма
вырабатывается метаболическая энергия в форме АТФ;
анаболические же процессы, как правило, являются
эндергоническими и потребляют АТФ.
34.
На уровне восстановительной способности:Катаболические
процессы являются в основном
окислительными
и
служат
донорами
высокоэнергетических электронов, для анаболизма
характерно обратное. Основным донором электронов в
восстановительных реакциях биосинтеза являются
НАДФ∙Н+Н+, восстановление которого происходит в
реакциях
катаболизма,
большей
частью
пентозофосфатном пути окисления глюкозы.
35. Основные пути использования атомов водорода восстановленных форм НАДФ
Синтез высших жирных кислот;Синтез холестерина;
Синтез нуклеотидов, в том числе коферментов и нуклеиновых кислот;
Синтез стероидных гормонов (половых, кортикостероидных);
Восстановление фолиевой кислоты в тетрагидрофолиевую;
Биосинтез метионина из гомоцистеина;
Биосинтез тимина и урацила;
Получение глюкозамина;
Аминирование α-кетоглутаровой кислоты в глутаминовую;
Превращение пировиноградной кислоты в щавелевоуксусную;
Восстановление глутатиона.