























































 Биология
БиологияПохожие презентации:



")






Обмен углеводов
1.
Тема лекции:ОБМЕН УГЛЕВОДОВ
Переваривание и всасывание углеводов в
желудочно-кишечном тракте поли- и
моногастричных животных. Гликолиз.
Суммарный энергетический эффект
анаэробного окисления глюкозы.
Лектор: доцент Васильева С.В.
2.
План лекции:1. Характеристика углеводов
2. Функции углеводов в организме
животных
3. Пищеварение углеводов у
моногастричных и полигастричных
животных
4. Гликолиз
5. Суммарный энергетический эффект
анаэробного окисления глюкозы
3.
Характеристика углеводов• Углеводы – многоатомные альдегидо- или
кетоспирты природного происхождения.
• По строению углеводы подразделяют на
моносахариды, дисахариды и полисахариды.
• Моносахариды – это семейство углеводов,
содержащих от 3 до 8 углеродных атомов. Они
имеют простое строение. Являются
структурными единицами для ди- и
полисахаридов.
• Самые распространённые в природе
моносахара – глюкоза, фруктоза, галактоза,
рибоза, дезоксирибоза.
4.
• Дисахариды – молекулы, состоящие издвух моносахаров, соединённые
гликозидной связью. Наиболее
распространённые – мальтоза, лактоза,
сахароза.
• Полисахариды – полимерные молекулы,
состоящие из множества циклических
моносахаридов, соединённых
гликозидными связями.
5.
Полисахариды:• Гомополисахариды (все мономеры
одинаковые)
• Гетерополисахариды (мономеры могут
быть разные)
К гомополисахаридам относят крахмал,
гликоген, инулин, целлюлозу.
К гетерополисахаридам –
гликозаминогликаны, гемицеллюлоза.
6.
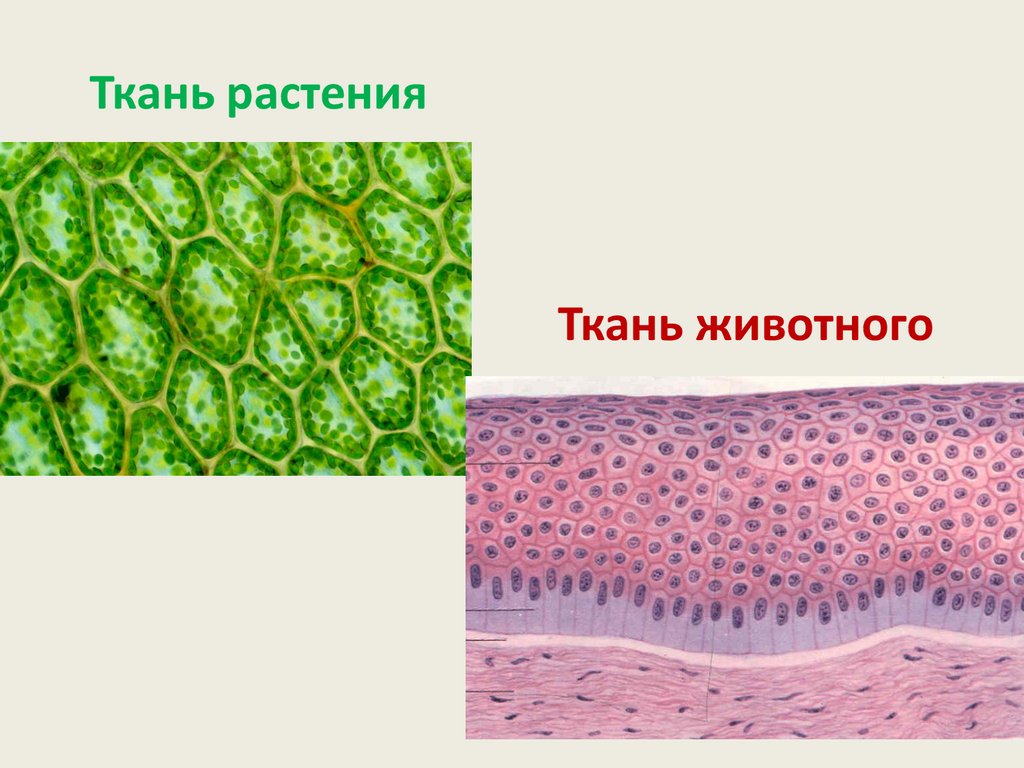
Обмен углеводов и их функции уживотных и растений различаются.
Существенно отличается и их
содержание в сухой массе.
• У растений углеводы составляют до 90% от
сухой массы
• У животных углеводы составляют порядка
2% от сухой массы
7.
• Это различие связано с тем, что у растенийуглеводы выполняют структурную
функцию, формируя клеточную стенку.
Прочная и толстая клеточная стенка
состоит преимущественно, из целлюлозы.
Именно целлюлоза обеспечивает роль
прочного каркаса растений, выполняя
механо-статическую функцию.
• В клетках животных клеточной стенки нет.
8.
Ткань растенияТкань животного
9.
Важнейшие функции углеводов уживотных:
• Энергетическая (аэробное и анаэробное
окисление глюкозы)
• Пластическая (в составе гликозаминогликанов в
соединительной ткани)
• Запасающая (гликоген)
• Рецепторная (клеточные рецепторы –
гликопротеины)
• Регуляторная (в составе гормонов –
гликопротеинов)
• Участие в составе нуклеотидов (в нуклеиновых
кислотах, макроэргах)
10.
Пищеварение углеводов уживотных
• Животные потребляют углеводы в составе кормов
растительного и животного происхождения. Их
пищеварение происходит с учётом особенностей
строения желудка. Но у всех животных углеводы в
пищеварительном тракте расщепляются до
моносахаридов.
• Процесс начинается в ротовой полости. Под
действием фермента альфа-амилазы слюны
начинается переваривание крахмала или
гликогена. Альфа-амилаза содержится в слюне
растительноядных и всеядных животных. У
плотоядных этого фермента практически нет.
• Альфа-амилаза гидролизует α-1,4 – гликозидные
связи крахмала или гликогена.
11.
Пищеварение углеводов умоногастричных животных
• Пищевой ком попадает в желудок, где
прекращается действие амилазы слюны
(при рН 1,5 – 2 фермент инактивируется).
Далее химус продвигается в тонкий
кишечник. В 12-типерстной кишке
открывается желчный проток, а вслед за
ним – проток поджелудочной железы. За
счёт желчи и сока поджелудочной железы
происходит защелачивание химуса (рН 7,5
– 8,0).
12.
• Поджелудочная железа секретирует впросвет кишечника альфа-амилазу,
аналогичную слюнной амилазе. Под
влиянием этого фермента молекулы
крахмала и гликогена частично
гидролизуются по α-1,4 –
гликозидным связям, образуя
декстрины с короткой цепью и
мальтозу.
13.
Далее эти фрагменты расщепляются доглюкозы с помощью кишечных
ферментов в процессе пристеночного
пищеварения:
• мальтаза – гидролизует
дисахарид мальтозу до двух
молекул глюкозы
14.
H2CH
H2C OH
OH
H
OH
O
H
H
OH
O
OH
H
H2C OH
H
H
O
H
OH
H
+
H
OH
OH
H
OH
H
H
O
H
OH
H
H
OH
+ H2O
HO
CH2 OH
O
H
H
OH
мальтаза
H
OH
OH
H
OH
15.
α-гликозидаза – гидролизует α-1,6 –гликозидные связи
O
O
-1,6-гликозидные
связи
O
O
O
O
O
O
O
O
гликозидные
связи
O
16.
В процессах пристеночного пищеваренияучаствуют и другие ферменты:
• сахараза – расщепляет сахарозу
до глюкозы и фруктозы
• лактаза – расщепляет лактозу до
глюкозы и галактозы
17.
• Целлюлозу переваривают толькомикроорганизмы, обитающие в желудочнокишечном тракте. Они вырабатывают
ферменты, расщепляющие β-гликозидные
связи. Целлюлозолитическая микрофлора у
моногастричных животных обитает в
толстом отделе кишечника. У лошадей
микрофлора есть и в желудке – в слепом
мешке. Бактерии расщепляют целлюлозу до
глюкозы, которая частично всасывается в
кровь, а частично утилизируется самими
бактериями и сбраживается до органических
кислот.
18.
Пищеварение углеводов уполигастричных животных
• У полигастричных животных
важнейшие пищеварительные
процессы происходят в рубце.
Здесь обитает огромное
количество амилолитических и
целлюлозолитических
микроорганизмов. К ним относят
бактерии, простейшие и грибы.
19.
• Амилолитические бактерии метаболизируютмолекулы крахмала кормов, но они не могут
разрушить целлюлозу, в которой углеводные
мономеры связаны бета-глюкозидными
связями. Амилолитические бактерии имеют
высокую толерантность к кислой среде. При
скармливании животным больших количеств
зерна, амилолитическая активность бактерий
резко возрастает и концентрация ионов
водорода в содержимом рубца увеличивается
в 10 – 50 раз.
20.
• Целлюлолитические бактерии за счетсвоих ферментов разрывают бета- 1,4глюкозидные связи, вызывая распад
полисахаров, которые составляют части
стенок клеток растений, таких как
целлюлоза (клетчатка), гемоцеллюлоза,
пектин. Получаемые при этом молекулы
глюкозы трансформируются затем в
летучие жирные кислоты. Распад
целлюлозы в рубце угнетается при рН
ниже 6,0.
21.
Глюкоза, образующаяся в рубце врезультате ферментации
полисахаридов и дисахаридов, вначале
превращается в молочную кислоту:
C6Н12О6
2 СН3 – СНОН – СООН
Молочная кислота
22.
Которая затем трансформируется вуксусную и пропионовую кислоты:
3СН3–СНОН–СООН
Молочная кислота
СН3–СООН +
Уксусная кислота
2СН3–СН2–СООН
+ Н2О + СО2
Пропионовая кислота
23.
Пример маслянокислогоброжения:
С6Н12О6 СН3–СН2–СН2–СООН + 2СО2 + 2Н2
Масляная кислота
24.
• В обычных условиях кормления в рубцелактирующей коровы в течение суток
образуются 2,5–5,0 кг ЛЖК, у бычков на
откорме 1,4–1,6 кг.
• Основные летучие жирные кислоты – это
уксусная, пропионовая и масляная
кислоты, являются конечными
продуктами метаболизма бактерий. Более
важным является тот факт, что летучие
жирные кислоты представляют основной
источник энергии кормов, используемый
жвачными животными.
25.
• На долю уксусной кислотыприходится 70%, пропионовой – 20%,
масляной – 8%.
• Отметим, что только пропионовая
кислота может быть
трансформирована в глюкозу в
клетках млекопитающих животных.
Летучие жирные кислоты
всасываются в кровь
непосредственно через стенки рубца.
26.
• У коров есть поджелудочнаяжелеза, в которой
вырабатывается амилаза, но с
низкой активностью. Она берёт на
себя вспомогательную роль,
воздействуя на непереваренные в
рубце остатки крахмала.
27.
Кишечная абсорбция продуктовпищеварения
Продукты гидролиза дисахаридов и
полисахаридов в основном в форме D-глюкозы,
D-галактозы, D-фруктозы хорошо всасываются в
тонком кишечнике на основе двух механизмов:
• Na+ - зависимого трансмембранного активного
транспорта с помощью Na+-K+-АТФ-азы
• Na+-независимого трансмембранного переноса
облегченной диффузии с участием
специальных переносчиков.
28.
ГЛИКОЛИЗ• Молекулы глюкозы, всосавшиеся в кровь
из кишечника, транспортируются по всему
организму и при участии гормона
инсулина переходят внутрь клеток.
• Для проникновение глюкозы в нервные
клетки и клетки крови инсулин не
требуется
• Первостепенная роль глюкозы –
окисление с целью получения энергии.
29.
• Гликолиз – это цепьферментативных реакций
окисления глюкозы до молочной
или пировиноградной кислоты.
• В анаэробных условиях конечным
продукта гликолиза является
лактат, в аэробных - пируват
30.
• Анаэробный гликолиз протекает собразованием только двух
молекул АТФ.
• Здесь АТФ образуется путём
субстратного фосфорилирования
(остаток фосфорной кислоты
передаётся на АДФ с
фосфорилированной молекулы
субстрата)
31.
Реакции гликолиза протекают вцитоплазме клетки
Гликолиз можно разделить на две фазы:
• В первой фазе происходит затрата двух
молекул АТФ на фосфорилирование
гексоз
• Во второй фазе происходит окисление
триоз и образование четырёх молекул
АТФ
32.
• Рассмотрим первую реакциюгликолиза. В ней происходит
фосфорилирование глюкозы с
участием АТФ по шестому
углеродному атому. Катализирует
реакцию фермент гексокиназа
33.
OH
O
C
C
H
C
OH
HO
C
H
C
OH
C
OH
H2C
OH
H
H
глюкоза
H
+
АТФ
гексокиназа
H
C
OH
HO
C
H
C
OH
C
OH
H
H
H2C
+
АДФ
O Р
глюкоза-6-фосфат
34.
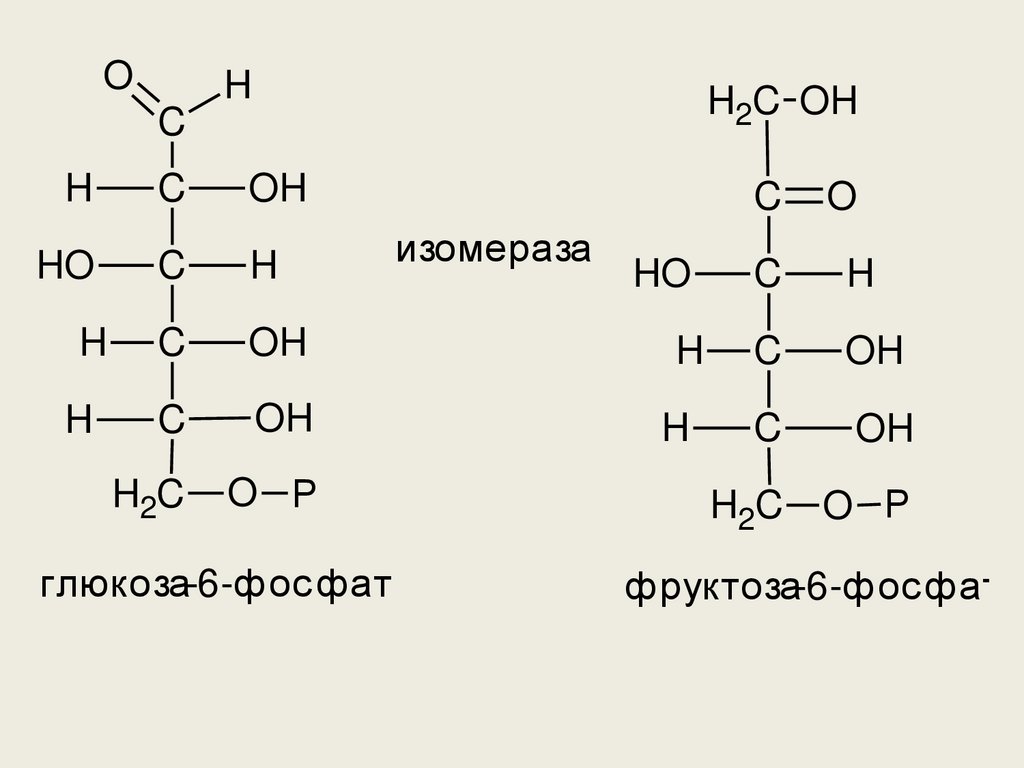
• Во второй реакции происходитизомерное превращение
глюкозы-6-фосфата во фруктозу-6фосфат. Фермент – глюкозо-6фосфат-изомераза
35.
OH
H2C OH
C
H
C
OH
HO
C
H
C
OH
C
OH
H2C
O Р
H
H
глюкоза-6-фосфат
C
изомераза
HO
H
H
O
C
H
C
OH
C
OH
H2C
O Р
фруктоза-6-фосфат
36.
• В третьей реакции затрачивается ещё однамолекула АТФ. Остаток фосфорной кислоты
переходит на молекулу при первом атоме
углерода. Образуется молекула –
фруктоза-1,6-дифосфат.
• Эта реакция является одной из ключевых и
катализируется аллостерическим ферментом
фосфофруктокиназой.
• Активность этого фермента подавляется
аллостерическим ингибитором – АТФ.
• Биологический смысл: при избытке АТФ в
клетке тормозится окисление глюкозы.
37.
H2CC
HO
OH
O
C
H
H
C
OH
H
C
OH
H2C
H2C
+
АТФ
O Р
фруктоза-6-фосфат
фосфофруктокиназа
HO
C
O Р
O
C
H
H
C
OH
H
C
OH
H2C
+ АДФ
O Р
фруктоза-1,6-дифосфат
38.
Четвёртую реакцию катализирует ферментальдолаза, который вызывает распад
молекулы гексозы на две триозы:
• 3-фосфоглицериновый альдегид
• Диоксиацетон-фосфат
Эти две триозы изомерны по отношению
друг к другу
39.
H2C OHH2C O Р
C
O
HO
C
H
H
C
OH
H
C
O Диоксиацетон-фосфат
C
альдолаза
OH
H2C O Р
фруктоза-1,6-дифосфат
H2C O Р
+
O
H
C
Фосфоглицериновый
H C OH
альдегид
CH2
OР
40.
Далее начинается вторая фазагликолиза
• Сначала диоксиацетон фосфат
превращается в 3фосфоглицериновый альдегид с
помощью триозофосфатизомеразы.
• Далее мы рассмотрим судьбу
3-фосфоглицеринового альдегида, но
будем помнить, что одна молекула
глюкозы породила две молекулы 3ФГА.
41.
• Следующие три реакции, которые мырассмотрим, являются этапами одной
ферментативной реакции,
катализируемой ферментом
глицеральдегид-3-фосфатдегидрогеназой
• Все изменения, которые мы
пронаблюдаем, будут происходить в
составе фермент-субстратного комплекса.
42.
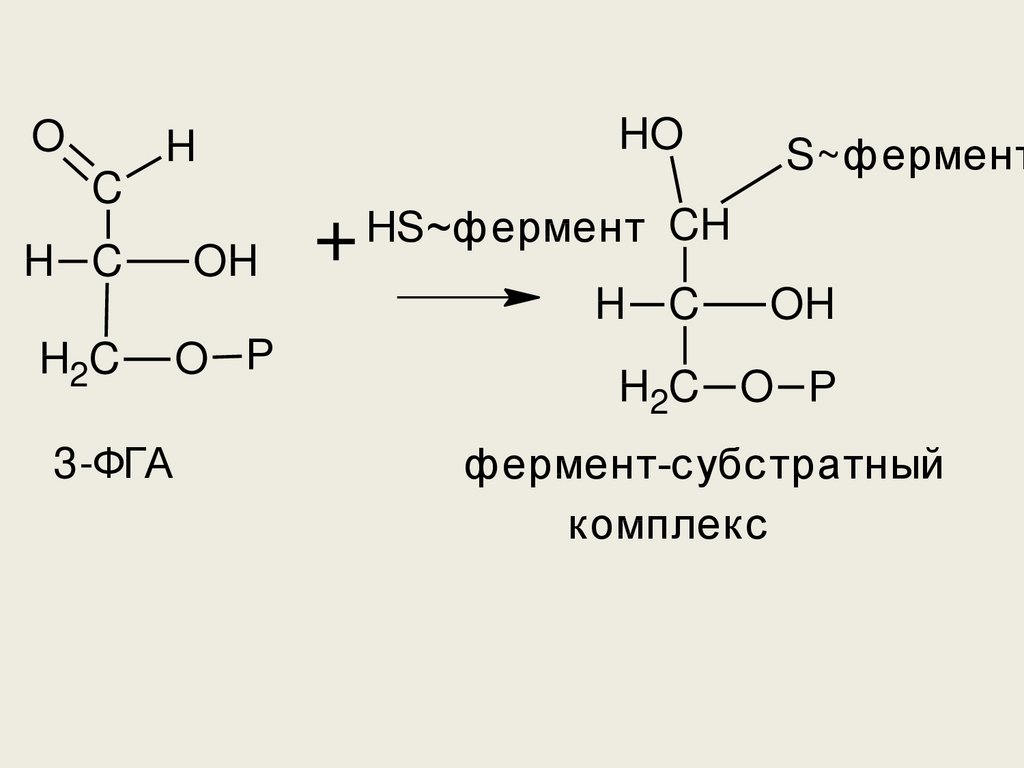
OH
C
H C
OH
HO
+ HS~фермент
CH
H C
H2C
3-ФГА
O Р
S~фермент
OH
H2C O Р
фермент-субстратный
комплекс
43.
HOS~фермент
CH
H C OH
H2C
O Р
+НАД
O
S~фермент
C
H C OH
H2C O Р
+ НАДН2
44.
• Важным моментом в даннойреакции является перенос двух
атомов водорода на НАД.
Восстановленная форма НАДН2 в
дальнейшем пойдёт на образование
молочной кислоты в конце
гликолиза.
• Фермент-субстратный комплекс
распадается в присутствии
неорганического фосфата
45.
OS~фермент
C
H
C
OH
+ Н3РО4
H2C O Р
фермент-субстратный
комплекс
O
O Р
C
H
C
OH
+ НS~фермент
H2C O Р
1,3-дифосфоглицериновая
кислота
46.
• Таким образом, при первом атомеуглерода появляется ещё один
остаток фосфорной кислоты, который
соединяется фосфоангидридной
связью.
• Расщепление этой связи является
высоко экзоэргическим процессом,
что и будет использовано для
образования молекулы АТФ в
следующей реакции
47.
OC
H
C
O
O Р
OH
C
+
АДФ
CH2 O Р
1,3-дифосфоглицериновая
кислота
OН
H C OH
+
АТФ
H2C O Р
3-фосфоглицериновая
кислота
48.
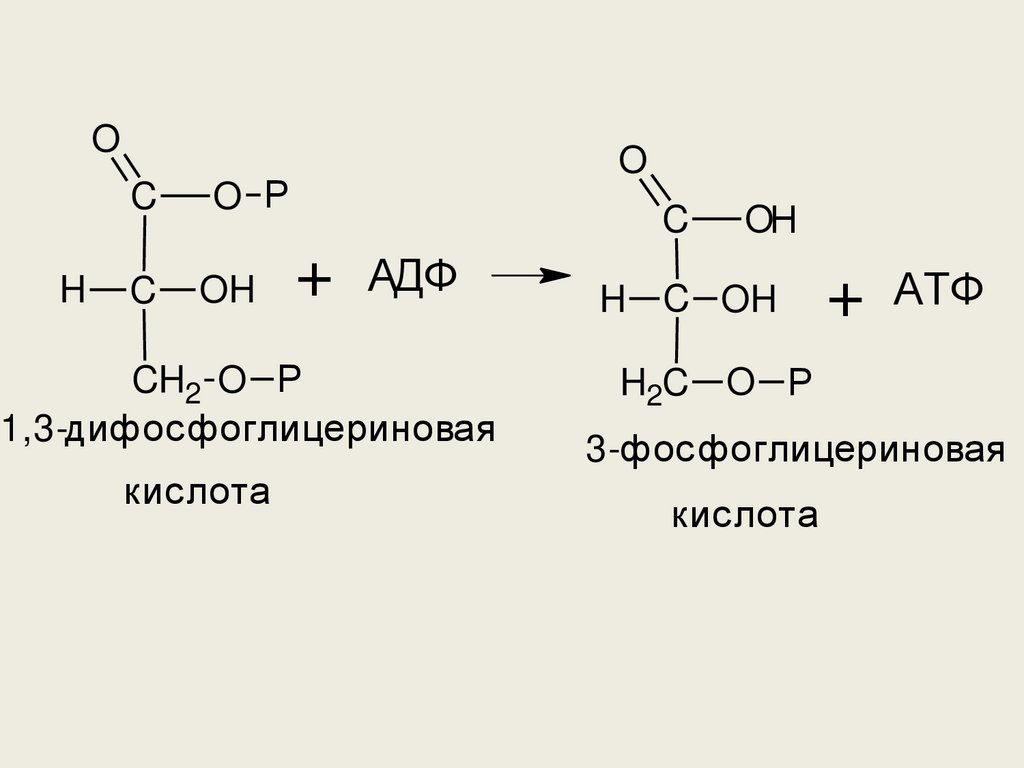
• Здесь мы видим первую реакциюобразования АТФ путём субстратного
фосфорилирования. Избыток
энергии, выделяющийся при
окислении альдегидной группы до
карбоксильной кумулируется в
фосфоангидридной связи. Энергия
гидролиза этой связи и используется
для образования АТФ.
49.
• Сейчас, после этой реакции можноутверждать, что возместились
затраченные в первой фазе гликолиза две
молекулы АТФ.
• Не забываем, что мы рассматриваем
судьбу только половины молекулы
глюкозы. Поэтому умножаем количество
АТФ на два.
50.
• Следующая реакция представляетсобой изомерное превращение 3фосфоглицериновой кислоты в 2фосфоглицериновую кислоту.
• Реакцию катализирует фермент
фосфоглицеромутаза в присутствии
катионов магния.
51.
OO
C
OH
фосфоглицеромутаза
H C
OH
2+
Mg
CH2O Р
3-фосфоглицериновая
кислота
C
H C
H2C
OH
O Р
OH
2-фосфоглицериновая
кислота
52.
• Следующая реакция протекает сучастием фермента енолазы,
который способствует отнятию
воды от 2-фосфоглицериновой
кислоты. В результате образуется
молекула фосфоенолпирувата.
53.
OO
C
OH
- H 2O
H C
O
Р
енолаза
H2C OH
2-фосфоглицериновая
кислота
C
OH
C
O
Р
CH2
2-фосфоенол-ПВК
54.
• Молекула фосфоенол-ПВК нестабильна,так как енольные формы стремятся
перейти в более энергетически выгодные
кето-формы.
• Для реализации кето-енольной
таутомерии молекула фосфоенол-ПВК
должна лишиться фосфатной группы,
после чего она легко перейдёт в
стабильную кето-форму.
55.
• Это используется для второйреакции субстратного
фосфорилирования, которая
протекает при участии фермента
пируваткиназы
56.
OO
C
OH
C
O Р
+АДФ
CH2
2-фосфоенол-ПВК
++
Mg
C
OH
C O
H3C
ПВК
+АТФ
57.
• Если гликолиз идёт в анаэробныхусловиях, то молекула ПВК
восстанавливается до молочной кислоты
с помощью НАДН2 (вспоминаем пятую
реакцию гликолиза, где от 3фосфоглицеринового альдегида водород
передаётся на НАД). При отсутствии
ферментов биологического окисления
этот же водород и восстанавливает ПВК
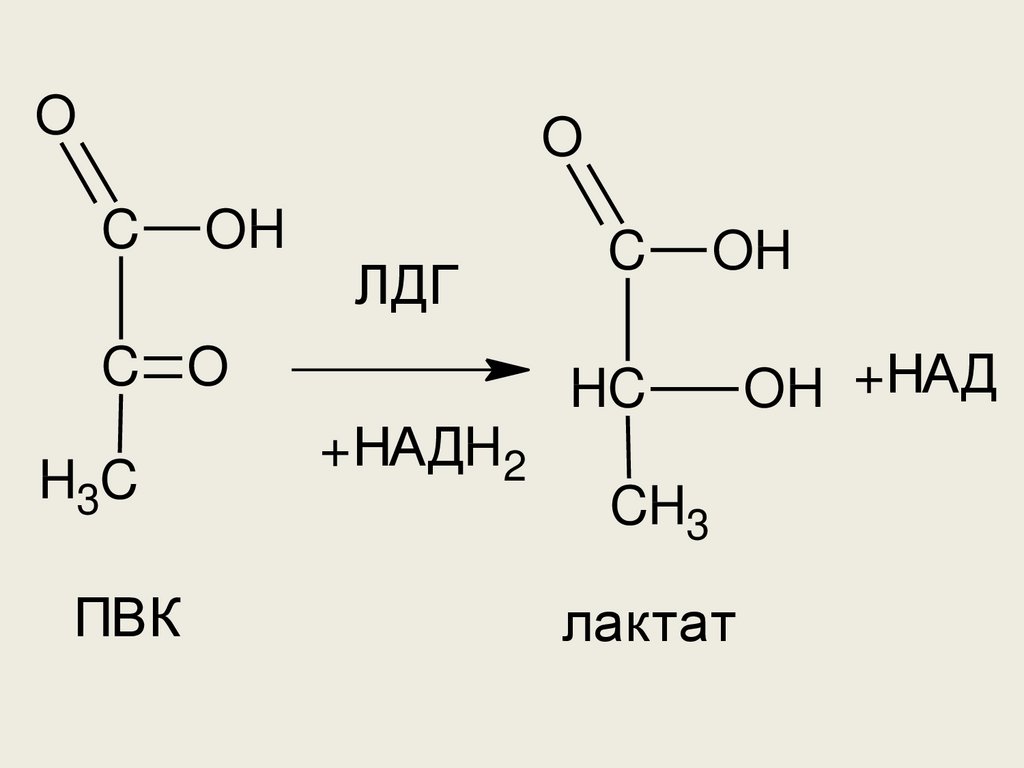
до лактата. Необходим фермент
лактатдегидрогеназа (ЛДГ)
58.
OO
C
OH
ЛДГ
C O
H3C
ПВК
C
OH
HC
+НАДH2
CH3
лактат
OH +НАД
59.
Суммарный энергетический эффектгликолиза можно выразить схемой:
С6Н12О6 + 2АТФ → 2 С3Н6О3 + 4 АТФ
«Чистая прибыль» – 2 АТФ
60.
• При окислении глюкозы по аэробномупути сначала проходит стадия гликолиза,
но образование молочной кислоты не
происходит. Пировиноградная кислота не
восстанавливается до лактата, так как
молекула НАДН2 идёт на цепь
биологического окисления.