


















































































 Биология
БиологияПохожие презентации:








")

Основы молекулярной биологии
1.
Крымский федеральный университетимени В.И. Вернадского
Медицинская академия имени С.И. Георгиевского
Кафедра биологии медицинской
Основы молекулярной биологии
Лекция №3 для фармацевтического
факультета
Агеева Елизавета Сергеевна
доктор медицинских наук,
доцент по кафедре патофизиологии,
Заведующий кафедрой биологии медицинской
Стоматологического факультета
2.
План лекцииОсновные понятия и определения
молекулярной биологии
Строение ДНК и РНК
Биосинтез белка
3.
Молекулярная биологияТермин ввел Фрэнсис Крик
Это раздел биологии который
раскрывает организацию и структуру
молекулярных механизмов важнейших
общебиологических явлений образует с
рядом других наук (биохимией) единую
систему знаний о механизмах хранения
и реализации генетической
информации
4.
1869 г. – открыты нуклеиновые кислотыИоганн Фридрих
Мишер
Открыл ДНК. Вначале новое вещество
получило название нуклеин, новое
фосфорсодержащее вещество,
соединенное с белками, которое назвал
нуклеином (от лат. nucleus – ядро).
Современное название нуклеина –
хроматин
Dahm RDev Biol. 2005 Feb 15;278(2):274-88.
5.
Нуклеиновая кислотаВ 1889 г. Рихард Альтман
разделил нуклеин на
нуклеиновую кислоту и
белок и предложил
термин «нуклеиновая
кислота», разработал
первый способ
выделения нуклеиновых
кислот, свободных от
белковых примесей
Рихард Альтман (1852-1900)
6.
1949г. Правила Чаргаффа (закон«эквивалентности» оснований)
Эрвин Чаргафф показал,
что ДНК из разных
биологических
источников содержит
равное коли- чество dT и
dA и равное количество
dC и dG, в то время как
соотношение количеств
этих пар бывает очень
Э. Чаргафф (р.1905)
американский биохимик
7.
1953 г. открыта модель молекулы ДНКПервая демонстрационная
модель двойной спирали
Сформулирован принцип
комплементарности
Фрэнсис Крик и Джеймс
Уотсон (лауреаты
Нобелевской премии)
8.
Физики рентгеноструктурщики –
Розалинда
Франклин и
Морис Уилкинс
получили первые
рентгенограммы
ДНК (19501953гг.)
Морис Уилкинс
Розалинда Франклин
9.
1961 год - структура генетическогокода
Дж. Генрих Маттеи
Маршалл Ниренберг
10.
1965 год - установление факта регуляциибактериального лактозного оперона у E. coli
Андре Львов • ЖАКОБ ФРАНСУА
Жак Моно
11.
1967 год - синтез биологическиактивной ДНК в системе in vitro1968 год - впервые осуществлен
химический синтез транспортной РНК
12.
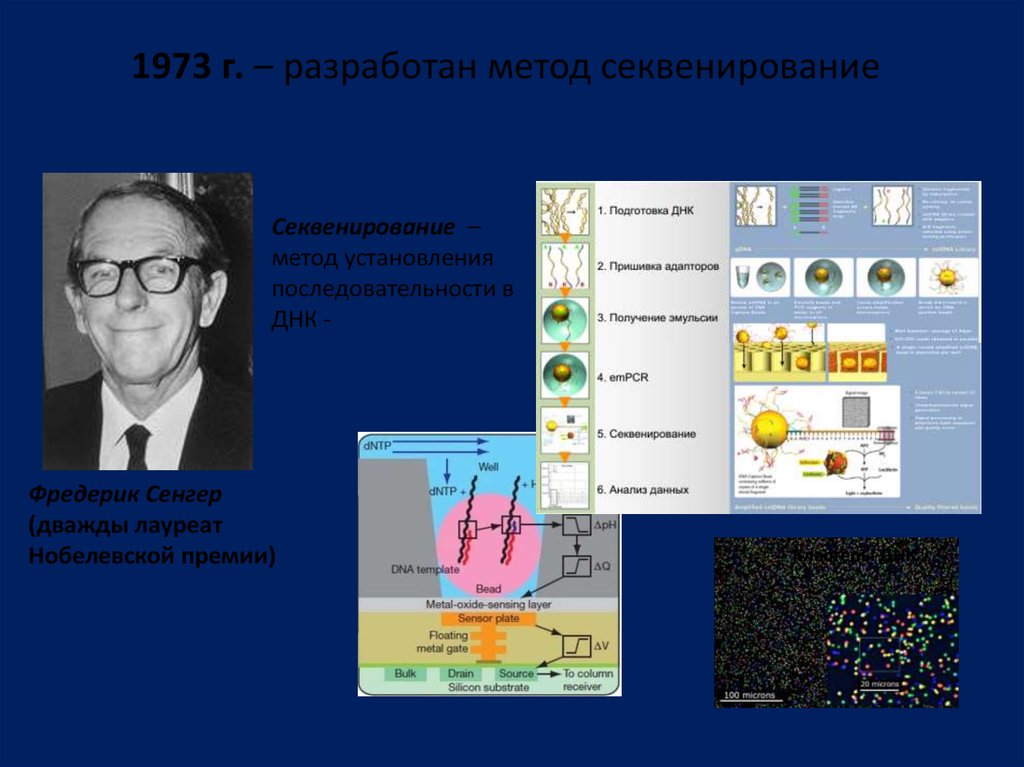
1973 г. – разработан метод секвенированиеСеквенирование –
метод установления
последовательности в
ДНК -
Фредерик Сенгер
(дважды лауреат
Нобелевской премии)
Кластеры ДНК
13.
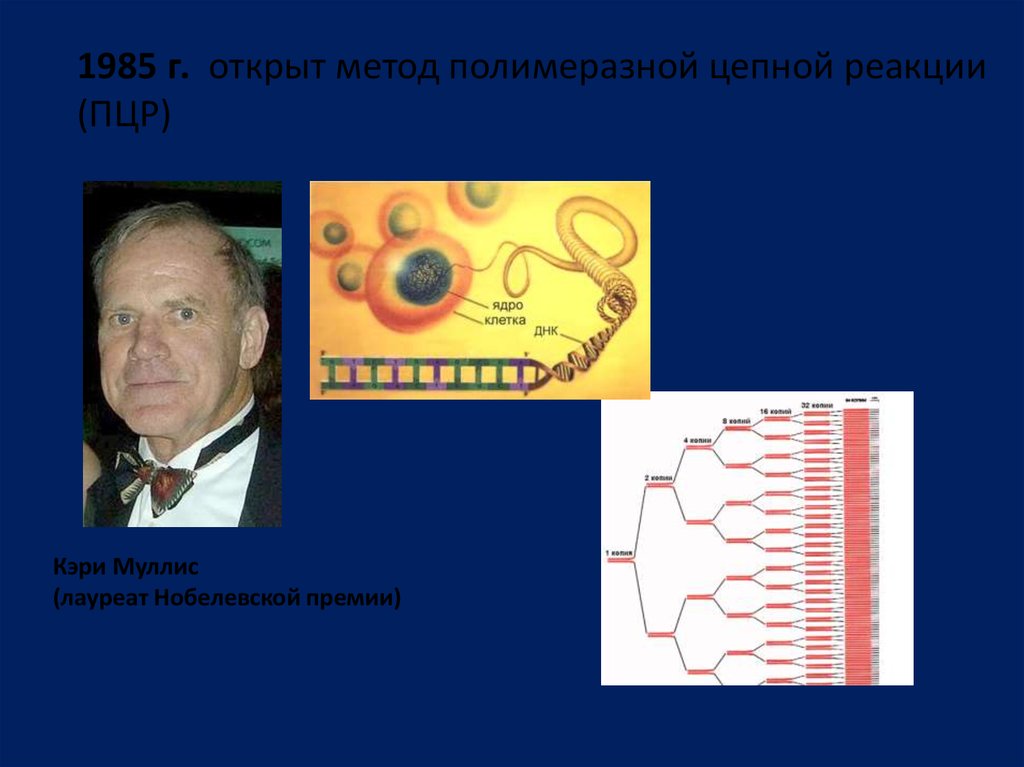
1985 г. открыт метод полимеразной цепной реакции(ПЦР)
Кэри Муллис
(лауреат Нобелевской премии)
14.
Современные представленияУстройство генетического материала у
высших организмов принципиально
отличается от низших .
Основные различия:
1. большее количество генов
2. сложность генома
3. принцип построения
15.
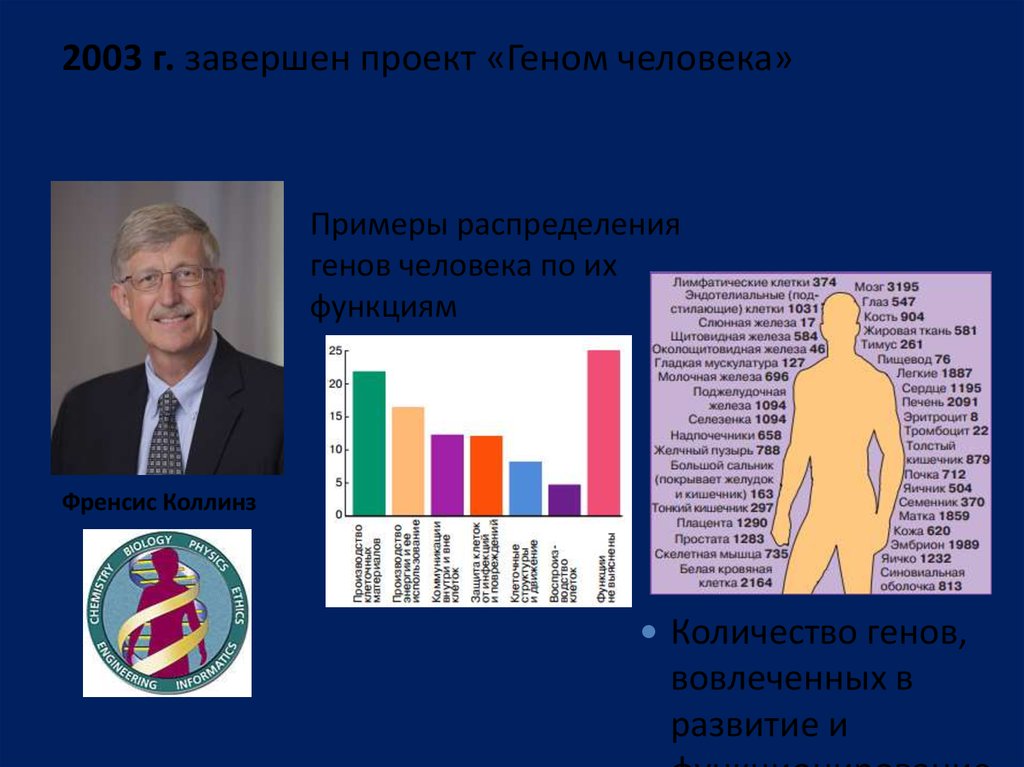
2003 г. завершен проект «Геном человека»Примеры распределения
генов человека по их
функциям
Френсис Коллинз
Количество генов,
вовлеченных в
развитие и
16.
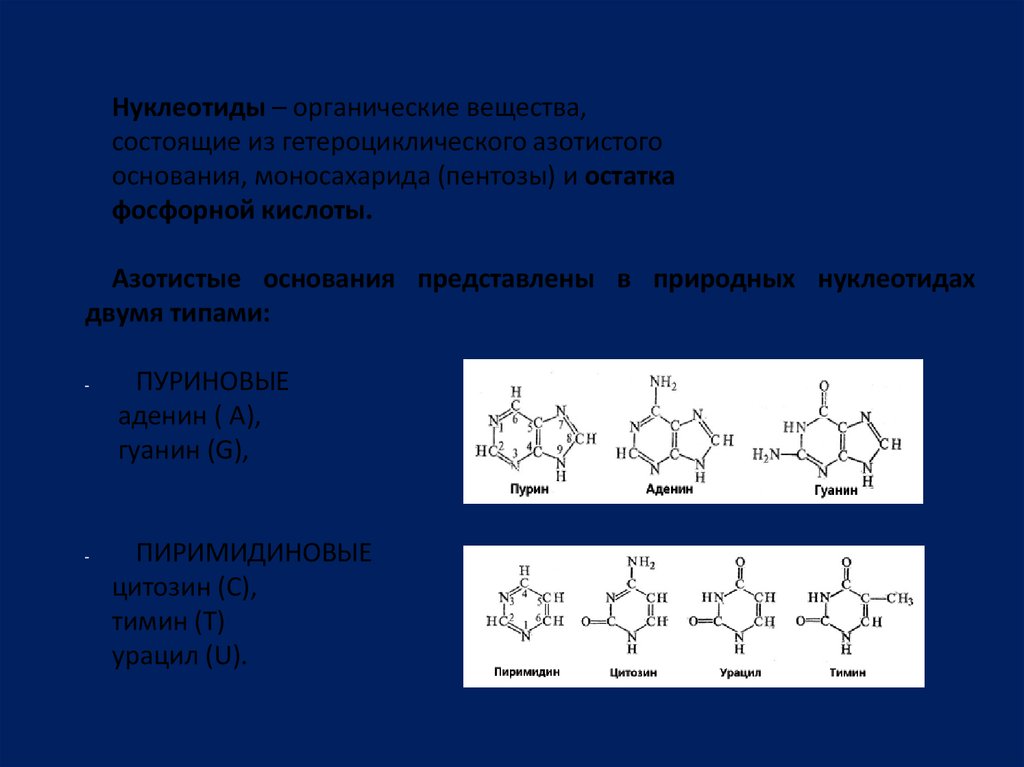
Нуклеотиды – органические вещества,состоящие из гетероциклического азотистого
основания, моносахарида (пентозы) и остатка
фосфорной кислоты.
Азотистые основания представлены в природных нуклеотидах
двумя типами:
-
-
ПУРИНОВЫЕ
аденин ( А),
гуанин (G),
ПИРИМИДИНОВЫЕ
цитозин (C),
тимин (Т)
урацил (U).
17.
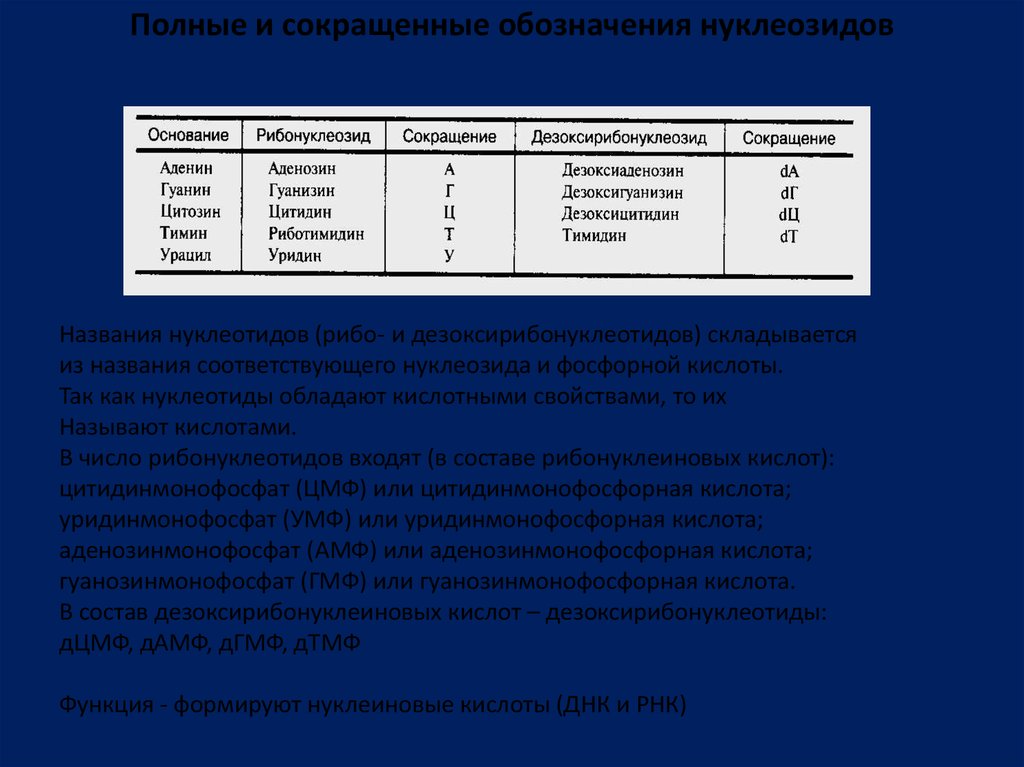
Полные и сокращенные обозначения нуклеозидовНазвания нуклеотидов (рибо- и дезоксирибонуклеотидов) складывается
из названия соответствующего нуклеозида и фосфорной кислоты.
Так как нуклеотиды обладают кислотными свойствами, то их
Называют кислотами.
В число рибонуклеотидов входят (в составе рибонуклеиновых кислот):
цитидинмонофосфат (ЦМФ) или цитидинмонофосфорная кислота;
уридинмонофосфат (УМФ) или уридинмонофосфорная кислота;
аденозинмонофосфат (АМФ) или аденозинмонофосфорная кислота;
гуанозинмонофосфат (ГМФ) или гуанозинмонофосфорная кислота.
В состав дезоксирибонуклеиновых кислот – дезоксирибонуклеотиды:
дЦМФ, дАМФ, дГМФ, дТМФ
Функция - формируют нуклеиновые кислоты (ДНК и РНК)
18.
Нуклеиновые кислоты являютсябиологическими полимерами.
Существует два типа нуклеиновых кислот:
ДНК (дезоксирибонуклеиновая кислота) и
РНК (рибонуклеиновая кислота), которая имеет
три разновидности:
информационная (матричная) РНК (и-РНК),
транспортная РНК (т-РНК)
рибосомальная РНК (р-РНК).
19.
СТРУКТУРА ДНКпоследовательност
ь нуклеотидов в 2-х
полинуклеотидных
цепях,
соединенных
между собой
фосфодиэфирными
связями в
3'-5'- направлении
20.
Двойная спираль ДНКрасположиться так, что против каждого нуклеотида
одной окажется селективно взаимодействующий с
ней другой нуклеотид.
Такие последовательности называются
комплементарными. направления цепей должны
быть противоположными (антипараллельными).
Примерами двух комплементарных
последовательностей могут служить фрагменты
ДНК
(5')dAdTdGdGdCdTdA(3') и (3')dTdAdCdCdGdAdT(5').
21.
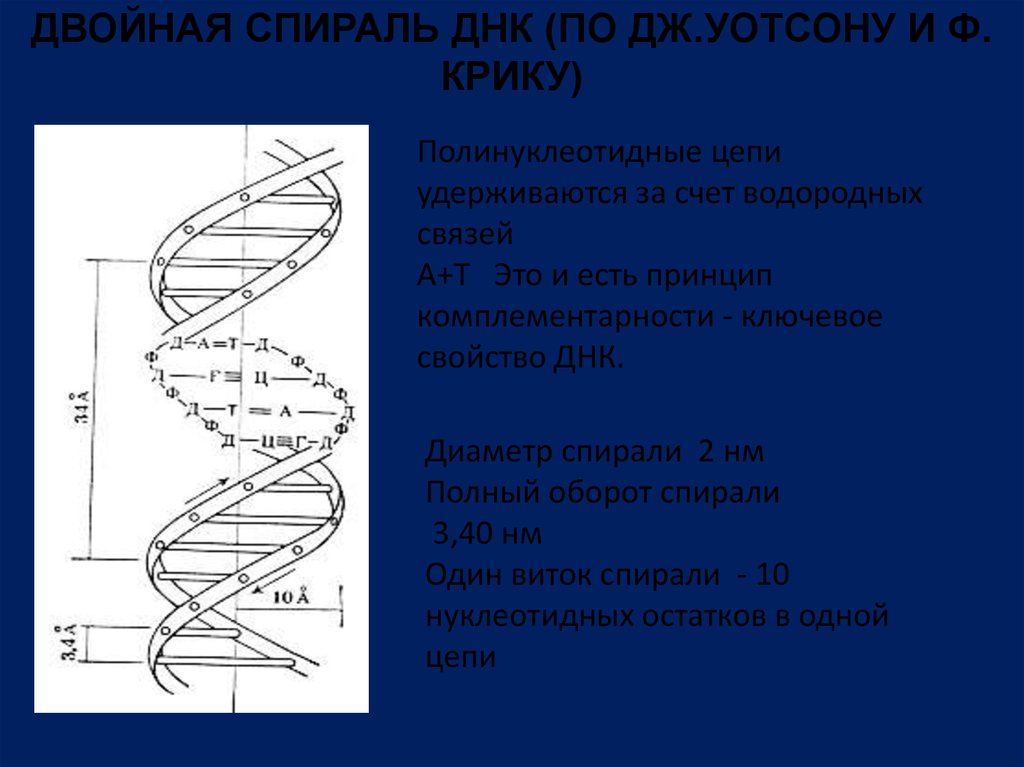
ДВОЙНАЯ СПИРАЛЬ ДНК (ПО ДЖ.УОТСОНУ И Ф.КРИКУ)
Полинуклеотидные цепи
удерживаются за счет водородных
связей
А+Т Это и есть принцип
комплементарности - ключевое
свойство ДНК.
Диаметр спирали 2 нм
Полный оборот спирали
3,40 нм
Один виток спирали - 10
нуклеотидных остатков в одной
цепи
22.
А-ДНКВ-ДНК
Формы ДНК
Z-ДНК
Молекулы ДНК могут находятся в различных
конформационных состояниях, зависящих от
степени обводненности биомолекулы, ионной
силы окружающей среды, типов катионов,
температуры. Правозакрученные спирали
образуют два семейства: А-семейство и В –
семейство
Z-форма ДНК выполняет какую-то регуляторную
роль в функциональных состояниях ДНК,
поскольку, возможны и обратные переходы из Zформы в В-форму
А-формой ДНК, выделенной при малой влажности.
имеет С3’-эндоконформацию сахара, что приводит к
уменьшению расстояния между фосфатными группами,
и уменьшению расстояния между нуклеотидными
парами вдоль оси спирали.
Увеличение количества нуклеотидов на виток спирали
(11 нуклеотидных остатков на виток вместо 10). Пары
оснований в А-форме образуют с осью спирали угол
около 20˚ и очень сильно отодвинуты от оси спирали к
периферии молекулы - сдвиг достигает 0,4 –0,5 нм т.е.
почти половины радиуса. Вследствие этого А-форма
ДНК при взгляде сверху- вдоль оси спирали, выглядит
как труба. В настоящее время показано наличие Аформы ДНК у некоторых бактерий, превращающихся в
споры
Считают, что в репликативных процессах
ДНК находится в В-форме
Полиморфизм – способность принимать различные
конформации
23.
В А-форме ДНК выполняет роль матрицы в
процессе транскрипции (синтез РНК на молекуле
ДНК).
В В-форме – роль матрицы в процессе
репликации (синтез ДНК на молекуле ДНК).
В С-форме – входит в состав надмолекулярных
структур хроматина и некоторых вирусов
(хранение информации).
Z-конформация – «горячие точки», где
реализуется участие ДНК в ряде метаболических
процессов.
24.
Третичная структура ДНК– это структура хроматина ядра и
хромосом.
Нуклеосома – элементарная
единица структуры хроматина,
компактная нуклеопротеидная
частица, состоящая из
нуклеосомного ядра – белкового
кора (коровые гистоныН2а,Н2в,Н3,Н4), вокруг которого
навита биспиральная ДНК (1.8
левых сверхвитка, ~150 н.п.).
ДНК, которая располагается между
нуклеосомами, называют линкерной ДНК
с длиной около 60 нуклеотидных пар.
Молекулы гистона Н1 «подтягивают»
отдельные нуклеосомы и тем самым
укорачивают размеры ДНК в 7 раз.
Такие архитектурные образования
защищают ДНК от действия нуклеаз.
25.
Суперспирализация ДНК формирует третичную структуру с помощьюразнообразных белков - гистоновых и негистоновых.
Гистоны - Н2А, Н2В, Н3 и Н4 в виде октамера образуют «нуклеосомный кор».
Молекула ДНК «накручивается» на поверхность гистонового октамера длиною
в 146 нуклеотидных пар, что составляет 1,75 оборота.
Такой комплекс белков с ДНК служит основной структурной единицей хроматина
и называется «нуклеосома».
Дальнейшие уровни компактизации молекулы ДНК происходят с участием
негистоновых белков, при этом каждый белок комплементарен определенной
последовательности нуклеотидов ДНК (сайт ДНК).
К этой группе относят семейство сайтспецифических белков.
К группе структурных и регуляторных белков относятся белки высокой
подвижности (HMG-белки), которые постоянно ассоциированы с хроматином.
Для них характерна маленькая молекулярная масса и высокое содержание
заряженных аминокислот.
К негистоновым белкам относятся ферменты
репликации, транскрипции, трансляции.
26.
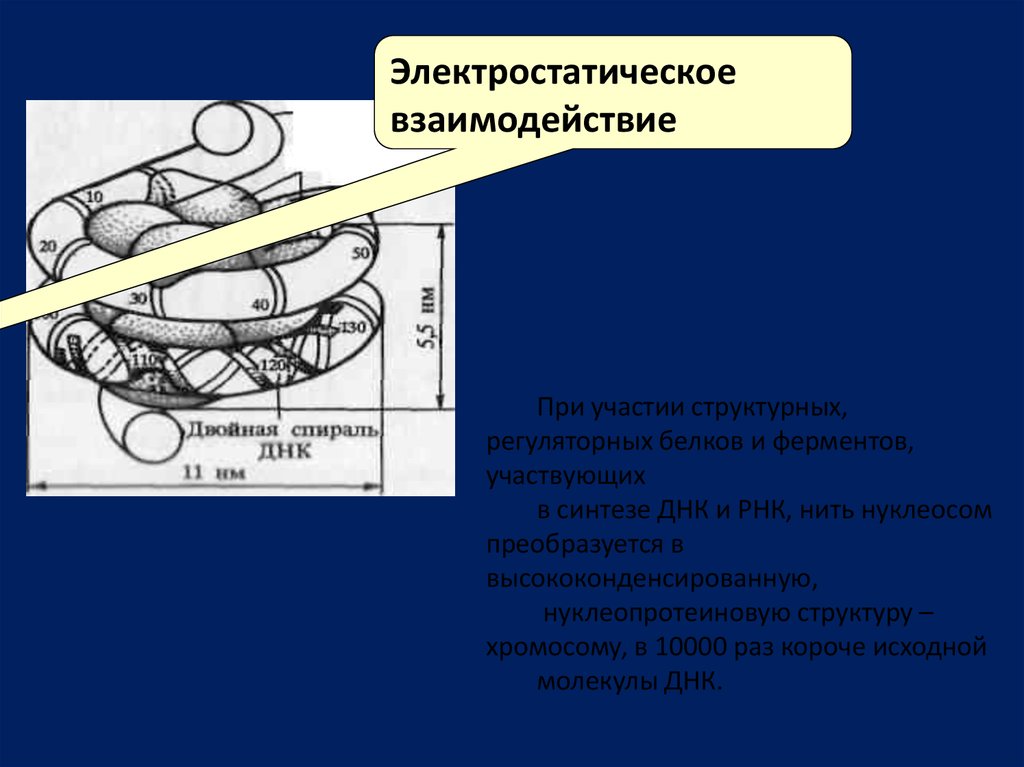
Электростатическоевзаимодействие
При участии структурных,
регуляторных белков и ферментов,
участвующих
в синтезе ДНК и РНК, нить нуклеосом
преобразуется в
высококонденсированную,
нуклеопротеиновую структуру –
хромосому, в 10000 раз короче исходной
молекулы ДНК.
27.
ФУНКЦИИ ДНК1. Аутосинтетическая – способность молекулы ДНК к
репликации, самоудвоение ДНК в S-период интерфазы
2. Гетеросинтетическая – участие ДНК в синтезе белка.
Основной
функцией ДНК
является
хранение и
передача
наследственной
информации в
ряду поколений
28.
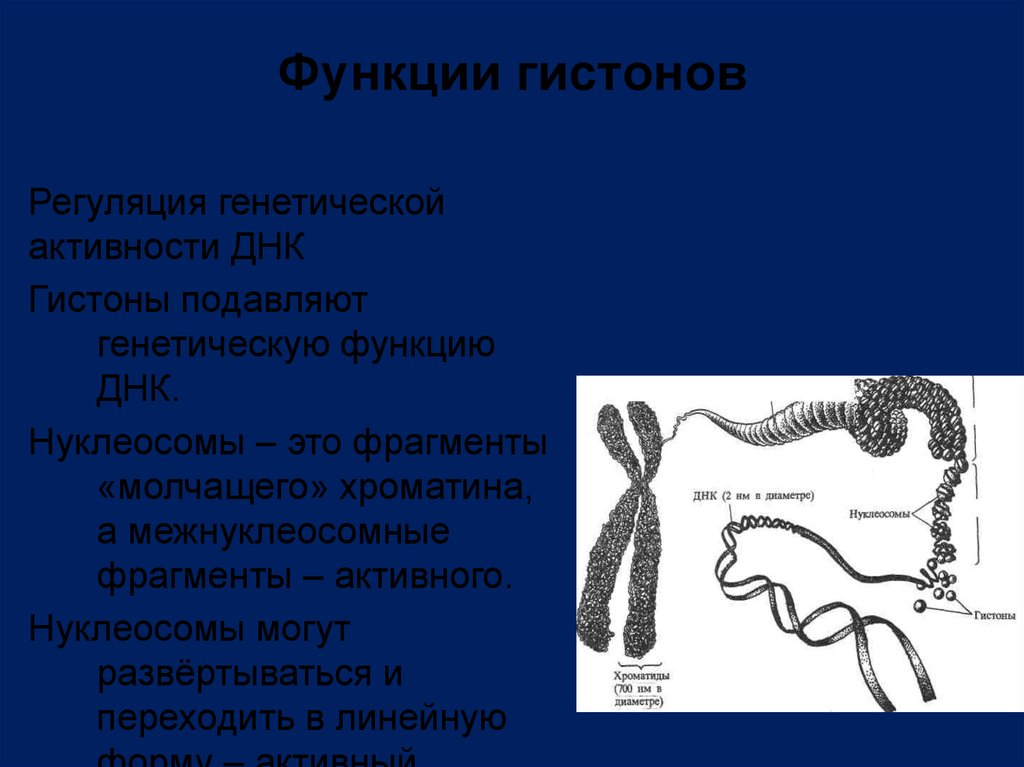
Функции гистоновРегуляция генетической
активности ДНК
Гистоны подавляют
генетическую функцию
ДНК.
Нуклеосомы – это фрагменты
«молчащего» хроматина,
а межнуклеосомные
фрагменты – активного.
Нуклеосомы могут
развёртываться и
переходить в линейную
29.
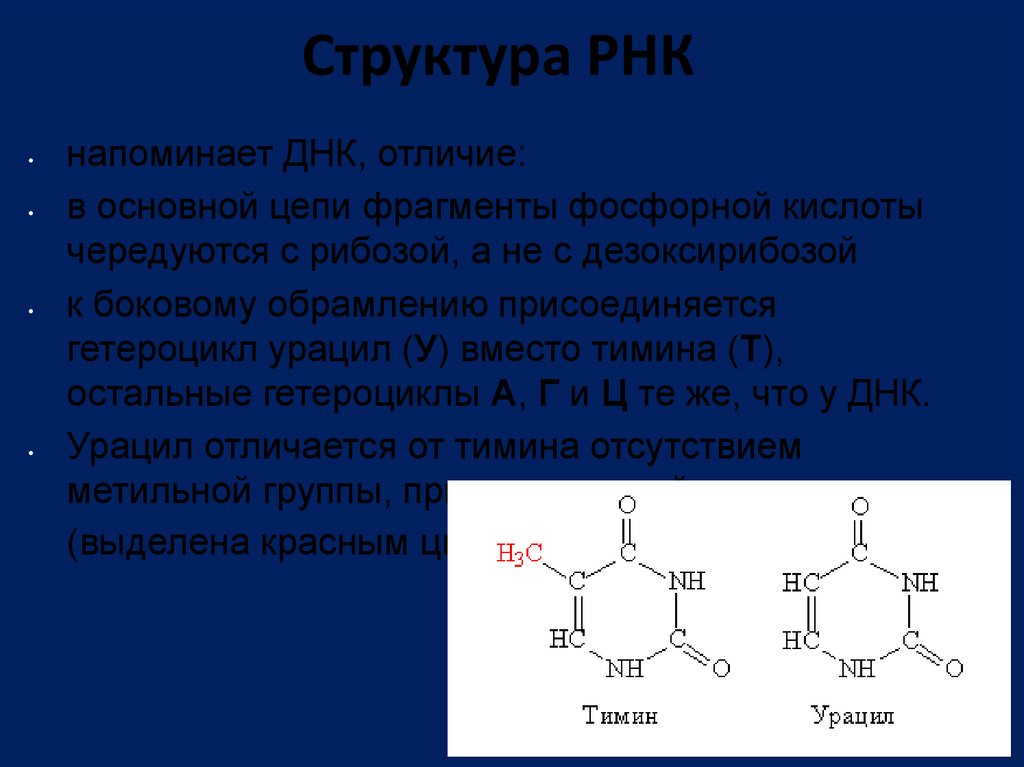
Структура РНКнапоминает ДНК, отличие:
в основной цепи фрагменты фосфорной кислоты
чередуются с рибозой, а не с дезоксирибозой
к боковому обрамлению присоединяется
гетероцикл урацил (У) вместо тимина (Т),
остальные гетероциклы А, Г и Ц те же, что у ДНК.
Урацил отличается от тимина отсутствием
метильной группы, присоединенной к циклу
(выделена красным цветом)
30.
Химические свойства РНКНапоминают свойства ДНК, однако наличие
дополнительных групп ОН в рибозе и меньшее (в
сравнении с ДНК) содержание стабилизированных
спиральных участков делает молекулы РНК
химически более уязвимыми.
При действии кислот или щелочей основные
фрагменты полимерной цепи Р(О)-О-СН2 легко
гидролизуются.
Группировки А, У, Г и Ц отщепляются легче.
используют деликатно действующие ферменты,
называемые рибонкулеазами.
31.
В цитоплазме клеток присутствуют три типа РНКтранспортные РНК (тРНК), матричные РНК
(мРНК) и рибосомальные РНК (рРНК).
они различаются по первичной структуре,
молекулярной массе , конформации,
продолжительности жизни и по выполняемым
функциям.
32.
Первичная структура РНК – порядок чередованиярибонуклеозидмонофосфатов в полинуклеотидной цепи.
Нуклеотиды в РНК связаны между собой как в ДНК - 3’,5’фосфодиэфирными связями.
На одном конце полинуклеотидной цепи находится фосфорилированная
ОН-группа 5’ углеродного атома, на другом – ОН-группа 3’-углеродного
атома рибозы.
Гидроксильная группа у 2’- углеродного атома рибозы делает молекулу
РНК нестабильной.
Для всех типов РНК характерно наличие специализированных участков.
Отдельные участки РНК образуют петли - «шпильки», за счет водородных
связей с комплементарными азотистыми основаниями.
Участки цепи РНК спиральных структурах антипараллельны, но не всегда
полностью комплементарны, в них встречаются неспаренные нуклеотидные
остатки или даже одноцепочечные петли, не образующие двойную спираль
33.
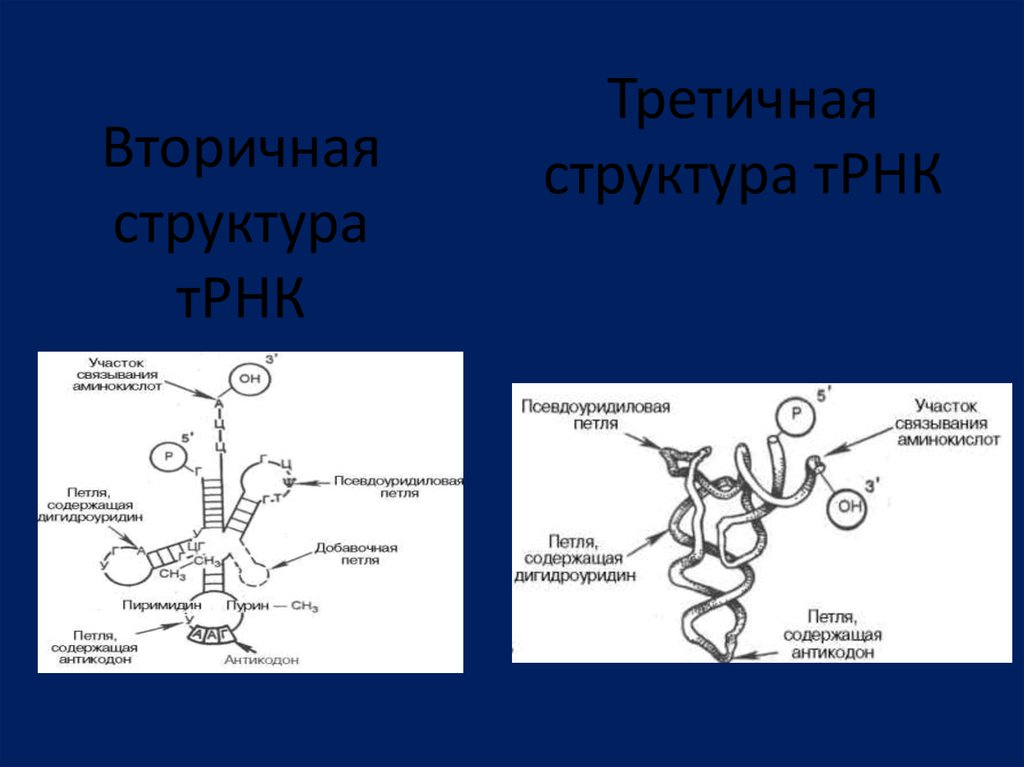
Транспортные РНКПространственная
структура
тРНК
описывается
универсальной
моделью
«клеверного листа».
Последовательность тРНК включают 70-90
нуклеотидов. Структура «клеверного листа»
состоит из 4-5 двухцепочечных спиральных
стеблей и трех петель.
Каждый стебель содержит 4–7 пар
комплементарных оснований.
Различают акцепторный, антикодоновый,
дигидроуридиловый,
псевдоуридиловый
и
добавочный ветви.
Акцепторная ветвь содержит 3’- и 5’- концы
полинуклеотидной цепи.
К 3’-концу присоединяется специфическая
аминокислота, отвечающая последовательности
антикодонового триплета в антикодоновой петле.
34.
Транспортные РНКГ-образная структура состоит из двух
спиралей расположенных почти
перпендикулярно одна относительно
другой. Одну спираль образуют уложенные
друг за другом антикодоновая (А) и
дигидроуридиловая (D) ветви, другую –
акцепторная и псевдоуридиловая (Т) ветви.
Антикодон имеет жесткую
архитектуру, которая позволяет ему быстро
считывать матричную РНК
35.
т-РНКНа долю тРНК приходится около
10% от общего содержания РНК в
клетке. Функции тРНК:
1) транспорт аминокислот к месту
синтеза белка, к рибосомам,
2) трансляционный посредник.
В клетке встречается около 40 видов
тРНК, каждый из них имеет
характерную только для него
последовательность нуклеотидов.
36.
Матричные Информационные РНК
разнообразны по содержанию нуклеотидов
и молекулярной массе (от 50 000 до
4 000 000).
На долю иРНК приходится до 5% от общего
содержания РНК в клетке.
Функции иРНК:
1) перенос генетической информации от
ДНК к рибосомам,
2) матрица для синтеза молекулы белка,
3) определение аминокислотной
последовательности первичной структуры
белковой молекулы.
37.
Рибосомные РНК содержат 3000–5000 нуклеотидов;
молекулярная масса — 1 000 000–1 500 000.
На долю рРНК приходится 80–85% от общего содержания РНК в
клетке. В комплексе с рибосомными белками рРНК образует
рибосомы — органоиды, осуществляющие синтез белка.
В эукариотических клетках синтез рРНК происходит в ядрышках.
Функции рРНК:
1) необходимый структурный компонент рибосом и, таким
образом, обеспечение функционирования рибосом;
2) обеспечение взаимодействия рибосомы и тРНК;
38.
Рибосомальные РНКВторичная
структура рРНК
Клетки прокариот и эукариот содержат рибосомы,
имеющие общий план строения.
В рибосомы входят высокомолекулярные рРНК,
дающие начало 30S-40S- и 50S –60S-субчастицам
рибосом; рРНК взаимодействуют с мРНК и
аминоацил-тРНК в процессе трансляции.
Низкомолекулярная 5S-рРНК в комплексе с
рибосомальными белками формирует комплекс,
который называют третьей субчастицей рибосомы, где
5S-рРНК выступает в роли посредника между
пептидилтрансферазным
центром
и
доменом
белкового фактора трансляции, обладающим ГТФазной активностью.
Вторичная структура рРНК характеризуется
спирализацией самой на себя полирибонуклеотидной
цепи.
39.
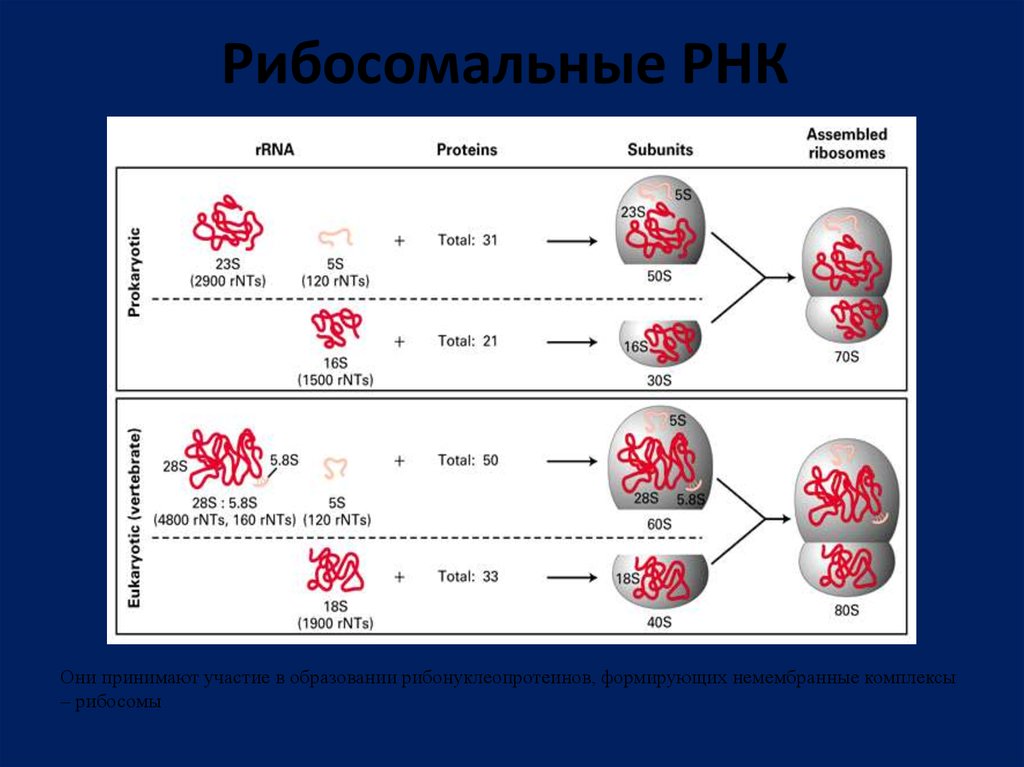
Рибосомальные РНКОни принимают участие в образовании рибонуклеопротеинов, формирующих немембранные комплексы
– рибосомы
40.
БИОСИНТЕЗ НУКЛЕИНОВЫХ КИСЛОТИ БЕЛКОВ (МАТРИЧНЫЕ
БИОСИНТЕЗЫ)
При биосинтезе новых молекул
нуклеиновых кислот и белков
носителями информации являются
нуклеиновые кислоты, в этой роли их
называют матрицами.
Матрица в ходе матричного синтеза не
расходуется и может использоваться
многократно.
41.
Генетическая информация
программирует синтез
специфических белков,
определяющих в свою очередь
специфичность структуры и
функции клеток, органов и
целостного организма (поток
информации):
ДНК –>РНК–>Белок–>Клетка –> Организм
42.
Биосинтез ДНКДНК-полимеразы обладают
3’ - 5’ экзонуклеазная
активонстью
1. Одноцепочечная – неспаренная ДНК
2. Дезоксирибонуклеозидтрифосфаты
3. Ферменты – ДНК-полимеразы,
топоизомеразы (гиразы), хеликазы,
лигазы, праймазы, ssb-белки
Различают три основных типа матричных биосинтезов
1) репликация ДНК - биосинтез ДНК с использованием в качестве матрицы уже существующих молекул ДНК;
2) транскрипция - биосинтез РНК по матрице ДНК;
3) трансляция - биосинтез белков с использованием в качестве матрицы мРНК.
43.
Репликация ДНКДвойная цепь ДНК служит матрицей для 2-х дочерних ДНК
Синтез происходит комплементарно
Репликация – процесс
удвоения хромосом в
течение S–фазы
клеточного цикла,
которая предшествует
делению клетки
Три механизма репликации
ДНК:
а – полукосервативный,
б – консервативный
в - дисперсивный
44.
1958г. М. Мезельсон и Ф. Сталь:
реплекация ДНК у эукариотов
осуществляется по полуконсервативному
механизму:
двойная спираль ДНК
сначала
раскручивается, цепи
расходятся, а затем
каждая
одноцепочечная
половина молекулы
ДНК достраивается до
45.
1963г. Ф. Жакоб, С. Бреннер, Ф. Кьюзен:Репликон – генетическая единица
репликации, то есть сегмент ДНК,
который автономно воспроизводится
(реплицируется) в процессе клеточного
роста и деления.
Каждый репликон имеет систему
«управления» собственной
репликацией.
46.
Для синтеза ДНК необходимы:Матрица ДНК (I цепочечная)
Субстраты - четыре dНТФ: dАТФ, dГТФ, dЦТФ,
dТТФ;
ДНК- pol
Mg2+;
РНК – «затравка»-праймер со свободным
3'-ОН группой.
47.
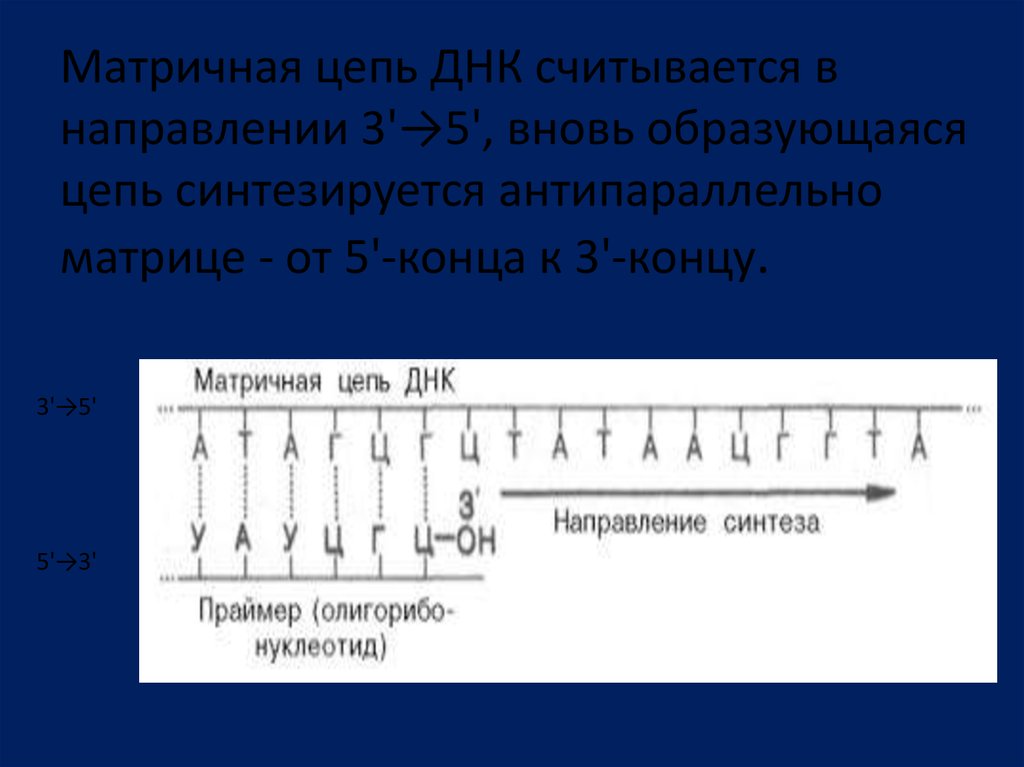
Матричная цепь ДНК считывается внаправлении 3'→5', вновь образующаяся
цепь синтезируется антипараллельно
матрице - от 5'-конца к 3'-концу.
3'→5'
5'→3'
48.
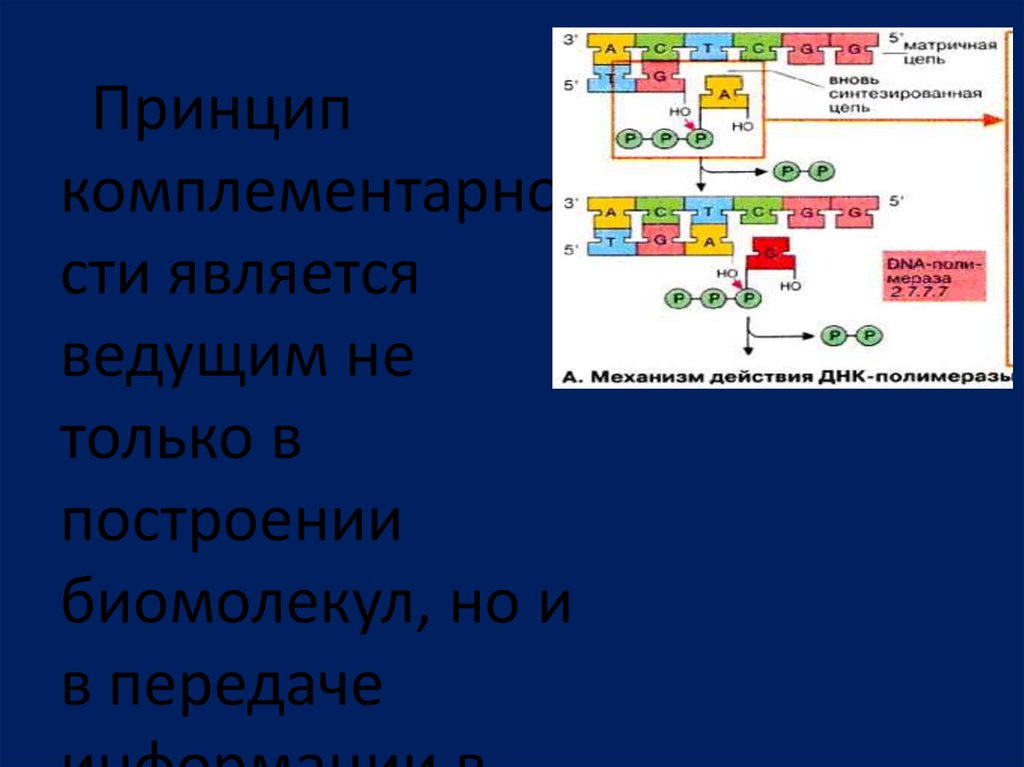
Принципкомплементарно
сти является
ведущим не
только в
построении
биомолекул, но и
в передаче
49.
Репликацию можно разделить на 3этапа:
инициация (начало) – образование
репликативной вилки.
элонгация (продолжение) – синтез
новых цепей.
терминация - завершение синтеза 2-х
дочерних цепей ДНК.
50.
В клетках эукариот имеютсяпять различных ДНК-зависимых
ДНК – полимераз: , , , ,
они могут синтезировать
нуклеотидную цепь только
направлении
5'→3'
51.
Инициация - созданиерепликативной вилки
Хеликаза - расплетение родительской ДНК,
на разделение каждой пары оснований
затрачивается 2АТФ.
Топоизомеразы (I,II,III) – обладая
нуклеазной активностью, участвуют в
регуляции суперспирализации ДНК.
52.
Образуется репликативный«глаз» - две репликативных
«вилки», каждая имеет
протяжённость до 2000
неспаренных оснований.
53.
Каждая из разделенных цепейсвязывается с ДНК- связывающими
белками (SSB), которые:
Стабилизируют одноцепочечные
участки ДНК, благодаря чему они
могут функционировать как матрица.
Защищают ДНК от внутриклеточных
нуклеаз.
54.
ДНК-зависимая РНК - роl(праймаза)
комплекса ДНК-полимераза праймаза синтезирует РНК - «затравки»
размером в 8-10 рибонуклеотидов,
которые затем элонгируются ДНКполимеразой , т.о., синтезируется
фрагмент ДНК (50 нуклеотидов).
55.
ДНК человека:Содержит 150 млн пар нуклеотидов
Скорость синтеза - 50 нуклеотидов в 1 мин.
Инициация синтеза происходит во многих
точках хромосомы, т.е.
эукариотические хромосомы имеют
полирепликонную организацию
56.
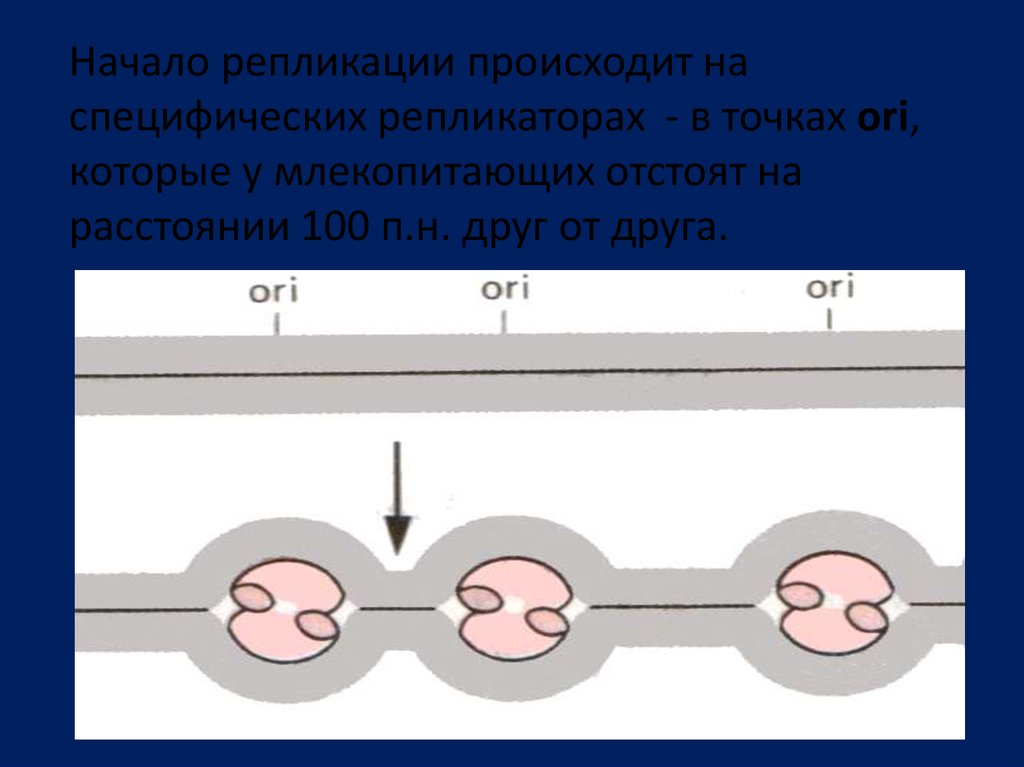
Начало репликации происходит наспецифических репликаторах - в точках ori,
которые у млекопитающих отстоят на
расстоянии 100 п.н. друг от друга.
57.
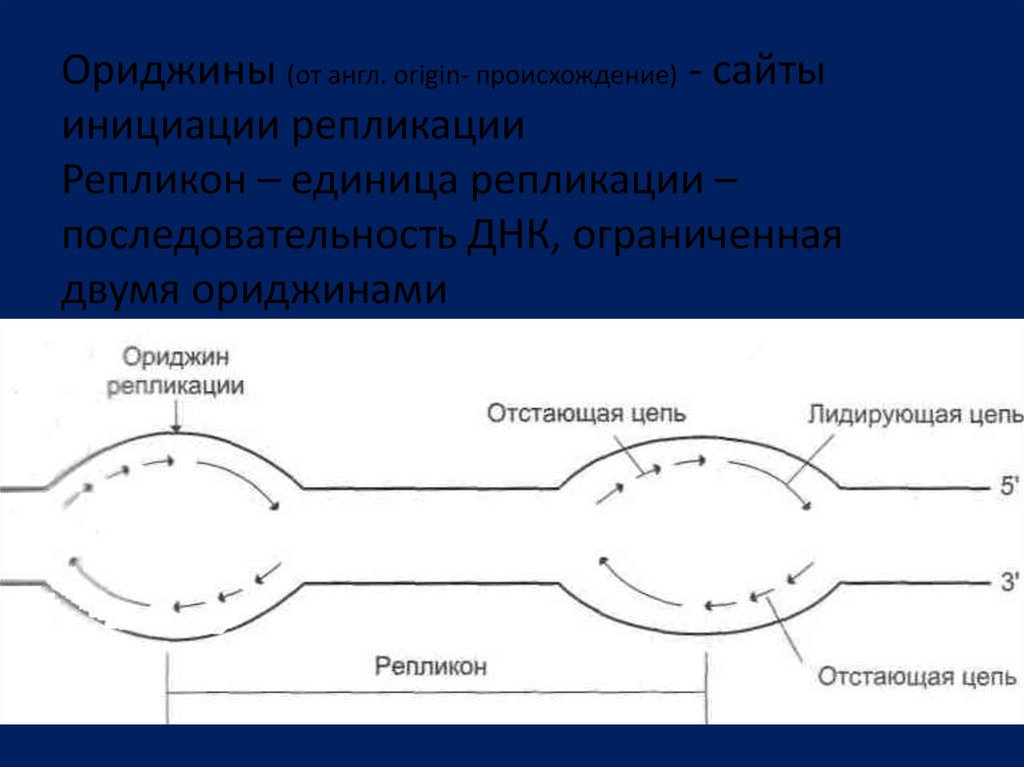
Ориджины (от англ. origin- происхождение) - сайтыинициации репликации
Репликон – единица репликации –
последовательность ДНК, ограниченная
двумя ориджинами
58.
Факторами, влияющими на клеточноеделение, являются:
а) факторы роста (пептиды, активирующие деление
определённого типа клеток);
б) наличие клеточных контактов, ингибирующих
клеточное деление;
в) уровень гормонов (половых стероидов, инсулина
и др.);
г) доступность пластических и энергетических
компонентов для деления клетки;
д) состояние клеточной мембраны, в том числе
количество в ней ХС.
59.
Известно 100 факторов роста и около 40факторов, подавляющих клеточное
деление
Механизмы действия факторов роста на
клетку аналогичны действию пептидных
гормонов:
фактор роста –> мембранный рецептор
–> сигнал, побуждающий клетку к началу
репликации.
60.
ЭЛОНГАЦИЯа) Синтез лидирующей цепи осуществляет ДНКполимераза , а отстающей ДНК-полимераза ε. Они
удлиняют «затравку», присоединяя к ней dНТФ один
за другим в направлении 5 3 .
б) по мере удлинения новой цепи ДНК, хеликазы и
топоизомеразы, обладающие обратимой
нуклеазной активностью, закручивают двойную
цепь ДНК в спираль;
61.
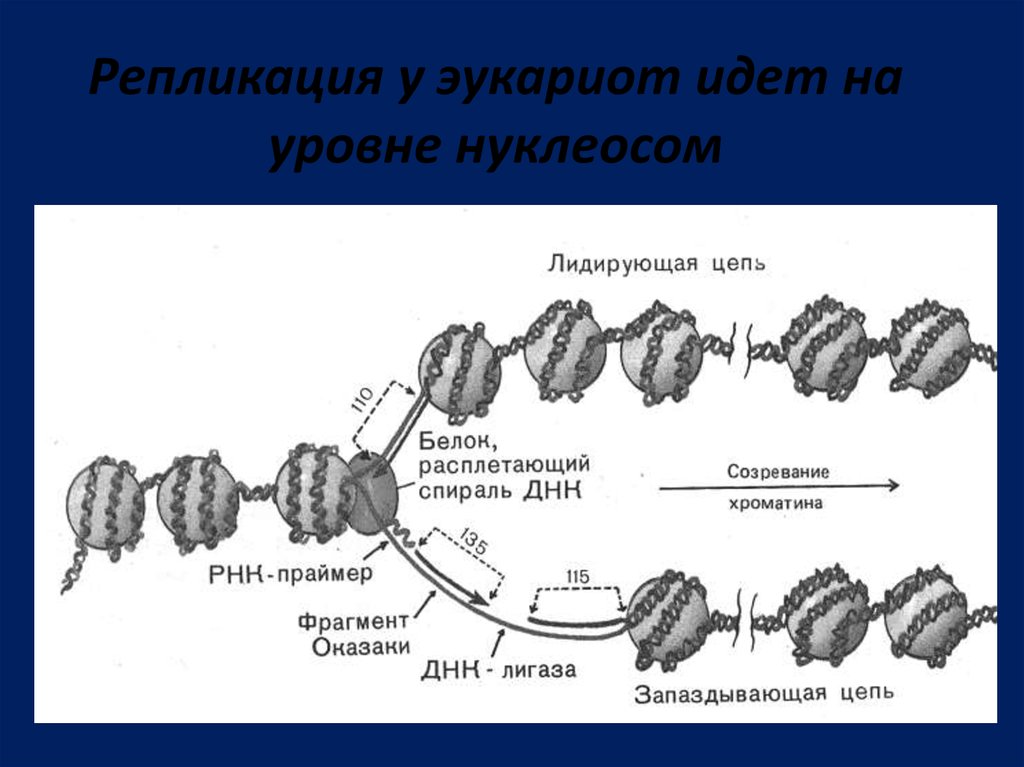
Репликация у эукариот идет науровне нуклеосом
62.
Элонгацияв) 5'-3' экзонуклеаза и РНК-аза Н1 удаляют
РНК-затравки (праймер).
г) ДНК-полимераза заполняет бреши.
д) ДНК-лигаза сшивает точечные разрывы,
образуя фосфодиэфирные связи.
63.
Потери генетической информациине происходит:
укорочение идет за счет теломер –
многократно повторяющейся на каждом
конце хромосомы последовательности
олигонуклеотидов – GGGTTA-.
Укорочение теломер в большинстве клеток
по мере их старения – фактор,
определяющий продолжительность жизни
организма.
64.
Механизм действия теломеразы:а) с помощью РНК фермент
комплементарно прикрепляется к
5' – концу недостроенной
дочерней цепи ДНК.
б) фермент по принципу
комплементарности удлиняет 5' –
конец цепи ДНК на один
гексануклеотид – GGGTTA- .
Синтез идет в направлении 3'→5'.
в) F смещается по цепи ДНК на
один теломер и начинается синтез
нового фрагмента -GGGTTA-
65.
Наибольшая активность теломеразыобнаружена в клетках с высокой
скоростью обновления:
Лимфоциты
Стволовые клетки костного мозга
Клетки эпидермиса кожи
Клетки эпителия и др.
66.
В соматических клеткахтеломераза неактивна
Соматическая клетка имеет длину
теломерной ДНК, достаточную для
времени жизни клетки и ее
потомства.
67.
Терминация репликацииПродвижение репликативной вилки
прекращается:
а) при столкновении с другой вилкой,
движущейся в противоположном
F
направлении,
б) по достижении конца хромосом.
68.
Точность репликации ДНК 1 ошибка на10
10 трансферазных реакций, однако
подобная ошибка обычно легко
исправляется за счет процессов
репарации.
69.
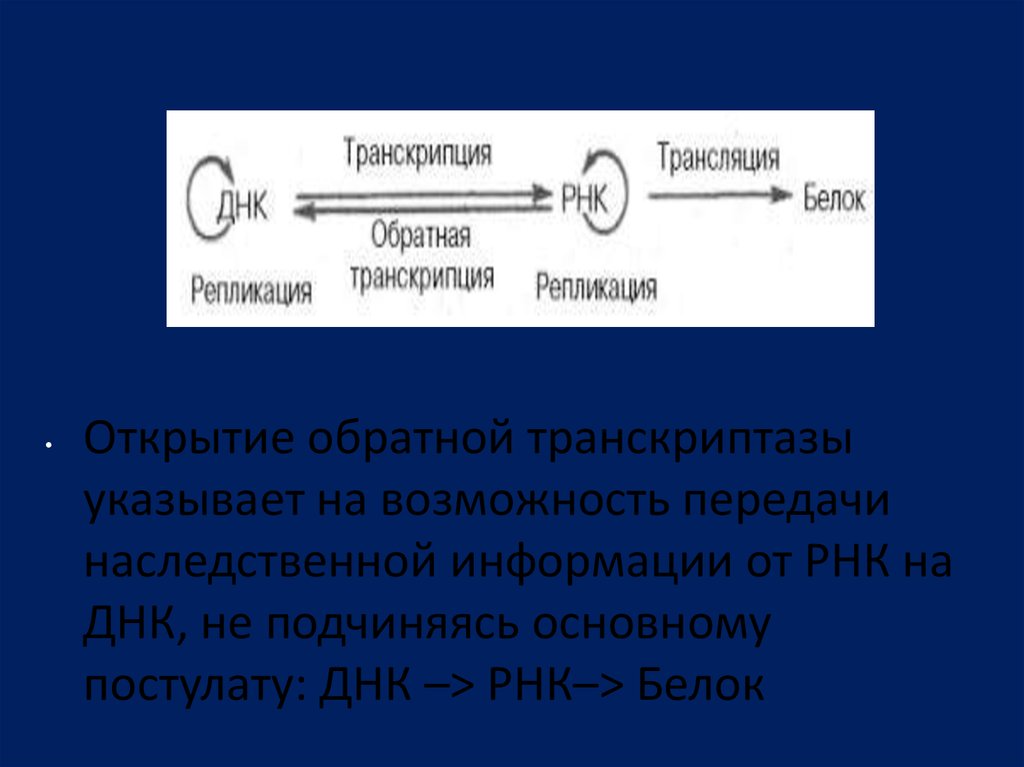
Открытие обратной транскриптазы
указывает на возможность передачи
наследственной информации от РНК на
ДНК, не подчиняясь основному
постулату: ДНК –> РНК–> Белок
70.
Экспрессия геновПроцесс транскрипции – биосинтез РНК на
матрице ДНК. Начальная стадия реализации
генетической информации. В результате
образуются тРНК, рРНК, и другие виды РНК,
кодирующие аминокислотные
последовательности.
Процесс трансляции – биосинтез белка на
мРНК.
71.
Транскрипцияпередача (переписывание)
информации, содержащейся в
последовательности нуклеотидных
остатков в ДНК-матрице, в
последовательность нуклеиновых
звеньев в молекуле новообразуемой
РНК.
72.
Транскриптон – часть молекулыДНК
транскрибируется в процессе
биосинтеза РНК на ней.
ограниченная промотором (точка начала
считывания информации) и сайтом
терминации, представляющая собой
единицу транскрипции.
73.
Независимая транскрипция гена – генобладает собственным промотором и
терминатором транскрипции.
Координированная транскрипция генов –
группа генов имеет общий промотор и
общий терминатор и составляет один
непрерывный участок ДНК, который
транскрибируется совместно (оперон).
74.
Ген эукариот является мозаичной структурой,содержащей наряду с кодирующими
(экзонами) также некодирующие (интроны)
последовательности
Экзоны (от англ. exit – выход) продукты транскрипции – мРНК выходят из
ядра в цитоплазму и выполняют функцию
матрицы в синтезе белка.
Интроны - вставочные, нетранслируемые
последовательности нуклеотидов в ДНК
эукариот.
75.
Соотношение информативной инеинформативной частей в
транскриптонах
Эукариотов – 1:9
Прокариотов – 9:1
76.
Соседние транскриптоны могут бытьотделены друг от друга
нетранскрибируемыми участками
Биологический смысл:
разделение ДНК на множество
транскриптонов позволяет осуществлять
с разной активностью индивидуальное
считывание (транскрипцию) разных генов.
77.
Ферменты транскрипцииВ 1960 г. открыт в двух лабораториях США (Дж. Хервиц и С.
Вейс)
ДНК - зависимые РНК – полимеразы –
нуклеотидилтрансферазы, в качестве субстратов используют
нуклеозид-5’-трифосфаты, транскриптаза).
Активны только присутствии ионов Mg2+, не нуждаются в
праймере
Рост цепи в направлении 5’-3’
Вместо основания Т в РНК включается У
78.
Фермент РНК-полимераза катализирует транскрипцию какэкзонов, так и интронов
Для синтеза РНК, катализируемого РНК pol необходимы:
ДНК-матрица.
Субстраты - НТФ: АТФ, ГТФ, ЦТФ, УТФ.
Mg 2+ или Мn 2+
79.
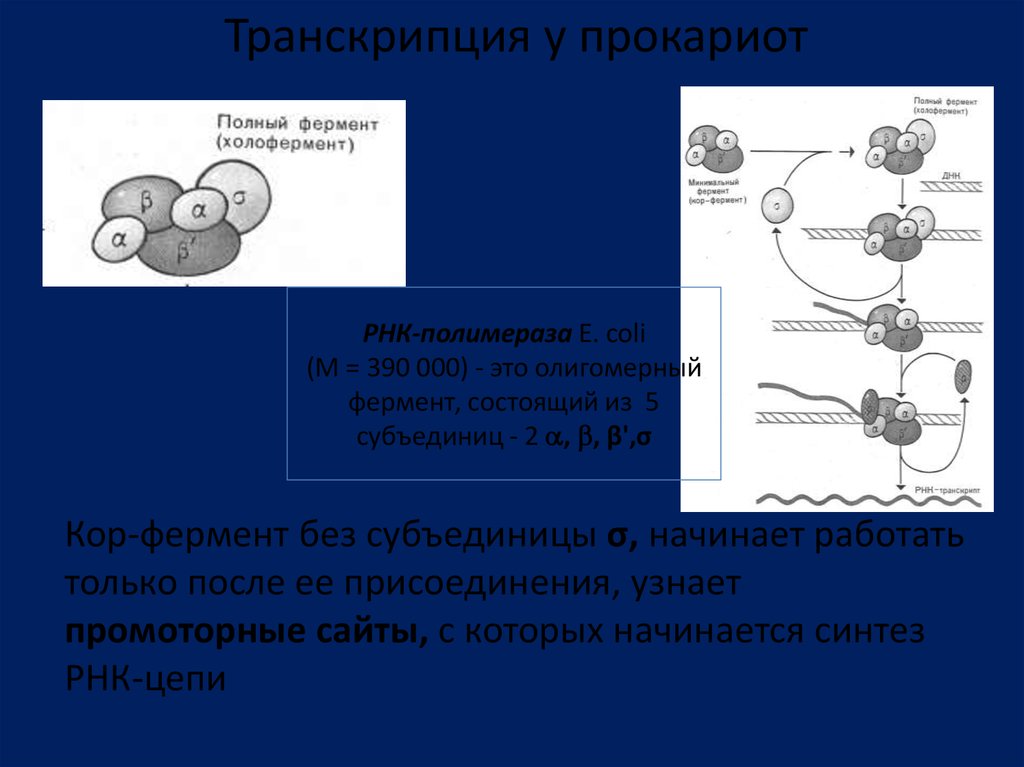
Транскрипция у прокариотРНК-полимераза Е. coli
(М = 390 000) - это олигомерный
фермент, состоящий из 5
субъединиц - 2 , , β',σ
Кор-фермент без субъединицы σ, начинает работать
только после ее присоединения, узнает
промоторные сайты, с которых начинается синтез
РНК-цепи
80.
ТранскрипцияИнициация
Элонгация
Терминация
-35
-10
+1
----TTGACA ----------------TATAAT-------------CAT--------AACTGT----------------ATATTA-------------GTA---16-19 bp
4-8 bp
Промотор – участок молекулы ДНК,
состоящий из 40 пар нуклеотидов и
расположен непосредственно перед
участком инициации.
Имеет 2 важные консервативные
последовательности:
ТАТААТ – расположена на расстоянии 10
п.н. от 1-го (1+) транскрибируемого
нуклеотида – Прибновобокс – РНК-pol
связывается с ДНК
(-35) – на расстоянии 35 п.н. до сайта
инициации – участок распознавания
промотора РНК-pol
81.
Факторы транскрипциирегуляторные белки обеспечивающие у
эукариот работу РНК-pol
ТАТА- связывающий белок (ТСБ, англ. ТАТАВох Binding Protein, TBP) взаимодействует с
основным регуляторным элементом
промотора -ТАТА-боксом, присутствующим в
большинстве генов
и облегчает взаимодействие РНК-pol c
промотором.
82.
РНК-pol связывается с промотором,происходит локальное раскручивание
двойной спирали ДНК (примерно 18 п.н.) и
образование открытого промоторного
комплекса – транскрипционный «глазок»
Нуклеотиды в области «глазка» становятся
доступными для спаривания с
рибонуклеозид-трифосфат – образуется
транскрипционная вилка
83.
Копирование смысловой (ДНК+) цепиНаправление 5’-3’
Первый нуклеотид всегда АТФ или ГТФ,
затем образуется первая
фосфодиэфирная связь
Отделение σ субъединицы, кор-фермент
перемещается по ДНК-цепи
84.
Элонгация транскрипцииФакторы элонгации (Е,Н,F) повышают активность
РНК-pol и облегчают расхождение цепей. Скорость
работы РНК-pol 50 нуклеотидов в секунду.
Растущий конец цепи РНК образует временную
гибридную спираль (12 н.п.) с ДНК-матрицей.
Наращивание РНК-цепи происходит на 3'-конце «замок молния»: впереди нее происходит
расхождение, а позади – восстановление двойной
спирали ДНК.
85.
Завершение синтеза РНКПроисходит в сайтах терминации (транскрипционные
терминаторы – GC – богатые участки) – строго
определенных участках матрицы.
3’-концы РНК-транскриптов образуют шпильки разной
длины, замедляет продвижение РНК – pol
Фактор терминации облегчает отделение первичного
транскрипта и РНК – pol от матрицы.
РНК – pol может вступить в следующий цикл
транскрипции после присоединения σ субъединицы.
86.
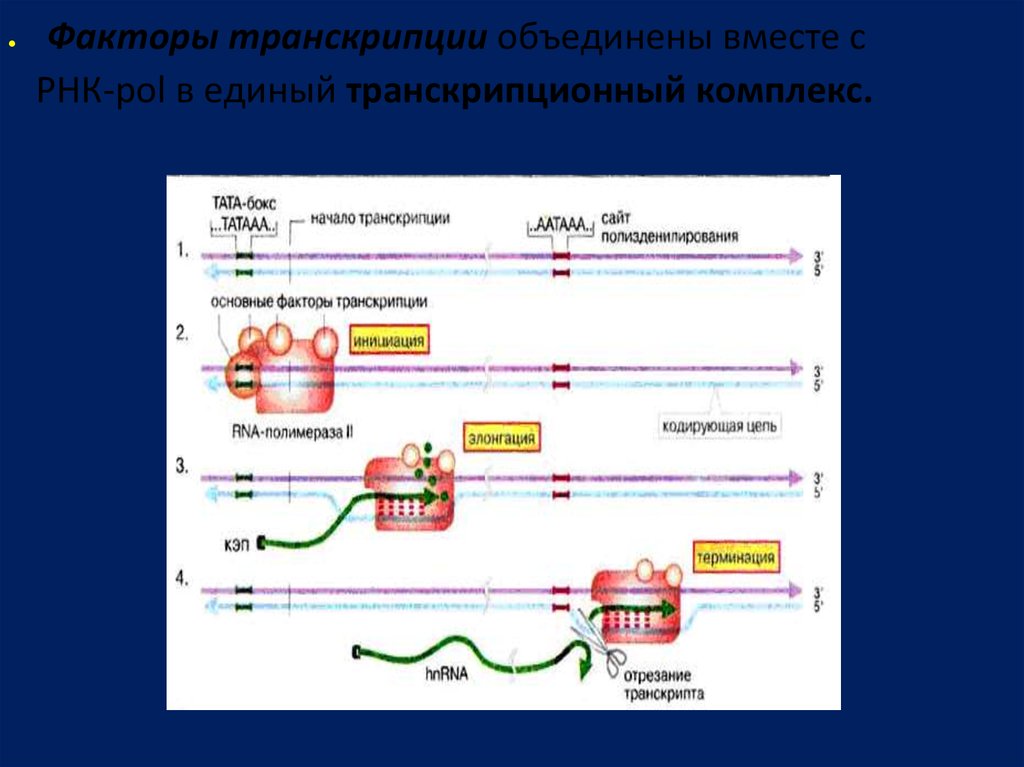
Факторы транскрипции объединены вместе сРНК-pol в единый транскрипционный комплекс.
87.
Транскрипция сходна с репликацейВ основе механизма процесса лежит принцип
комплементарности (А=У, Г=Ц) и
антипараллельности: синтез РНК идет в 5'→3‘
– направлении, а считывания информации с
матрицы – в 3'→5‘
2.
В основе процесса – полимеразная,
нуклеотидилтрансферазная реакция (реакция
элонгации)
3. Локализована в ядре
1.
88.
Транскрипция отличается отрепликации рядом особенностей:
1. Не требует синтеза праймера.
2. Использует не всю молекулу ДНК, а только ее
отдельные короткие сегменты (отдельные гены или
группы генов).
3. Требует наличия только одной из цепей ДНК в
качестве матрицы (матричная цепь), которая
полностью сохраняется (при репликации ДНК она
сохраняется наполовину).
89.
Регуляторные элементы,стимулирующие транскрипцию
связанных с ними генов, называются
энхансерами (усилителями, от англ.
enhancer).
подавляющие транскрипцию —
сайленсерами (успокоителями, от
англ. silencer).
90.
Транскрипционные факторы(Транс- действующие факторы)
это белки, т. е. продукты других,
независимых генов, поэтому их
называют опосредованно
действующими факторами
91.
Процессинг первичныхтранскриптов РНК
Между транскрипцией и трансляцией существует важное
звено – процессинг или созревание мРНК.
Созревание образованных РНК
У прокариот первичные транскрипты функционируют в виде
мРНК без последующей модификации или процессинга
У эукариот первичные транскрипты – гетерогенные ядерные
РНК (гяРНК) – содержт интроны, которые должны быть
удалены – сплайсинг (от англ. to splice - сплетать, сращивать).
Происходит ковалентное соединение участков РНК,
транскрибированных с экзонов
92.
Схема сплайсинга РНКИнтрон
Катализируется
протеиновым
комплексом,
сплайсосомой
Состав: белки, мяРНК
(рибозимы –
катализируют
расщепление
фосфодиэфирных
связей), богатые
урацилом
Экзон 1
Экзон 2
Реакция 1
Экзон 1
Интрон
Экзон 2
Реакция 2
Сплайсированная РНК
Экзон 1
Экзон 2
Интрон
93.
Регуляция генной экспрессииНе все белки синтезируются постоянно
Некоторые белки синтезируются только при
определенных условиях
Синтезируемые белки с постоянной скоростью
называются конститутивными (промоторы
генов не имеют «выключателя»)
Синтезируемые с изменяющейся скоростью в
зависимости от условий – адаптивные или
индуцибельные
94.
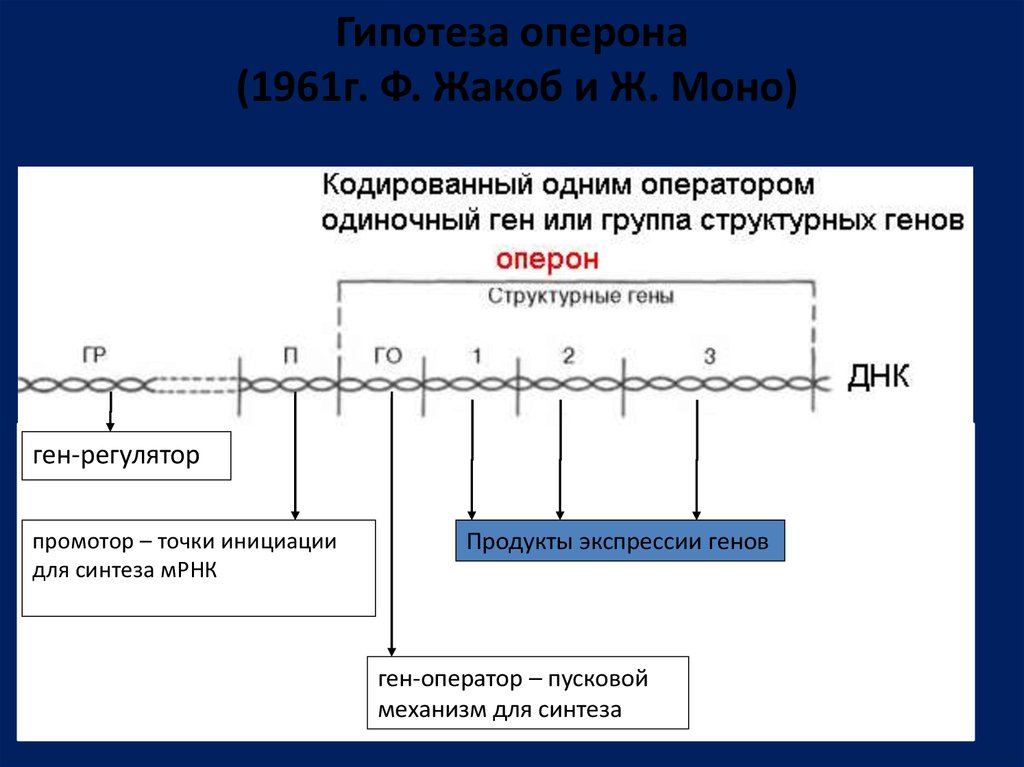
Гипотеза оперона(1961г. Ф. Жакоб и Ж. Моно)
ген-регулятор
промотор – точки инициации
для синтеза мРНК
Продукты экспрессии генов
ген-оператор – пусковой
механизм для синтеза
95.
Организация хроматина вдифференцированных клетках
многоклеточного организма
Гетерохроматин (стабильно
репрессированные участки)- ДНК упакована
компактно и недоступна для транскрипции.
Эухроматин (потенциально активные
участки) – ДНК имеет рыхлую укладку и
способна связывать РНК-pol.
96.
В разных типах клеток в областьэухроматина попадают разные
гены, а это означает, что в разных
тканях транскрибируются
разные участки хроматина.
(менее 1% генов доступно для
транскрипции)
97.
Регуляторные зоны ДНКЭнхансеры и сайленсоры
различны по числу и
расположению на молекуле ДНК
для различных генов в разных
тканях, то есть являются
тканеспецифическими
характеристиками
98.
Генетический кодЭто способ записи
информации об
аминоксилотной
последовательнос
ти белков с
помощью
последовательнос
ти нуклеотидов в
мРНК
99.
СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДАТриплетность - кодовое число = 3. Три
нуклеотидных остатка (триплет) кодируют одну
аминокислоту. Терминирующие триплеты – УАА, УАГ.
УГА не кодируют аминокислоты и являются сигналами
к прекращению синтеза белка.
Специфичность – каждый триплет кодирует только
одну аминокислоты.
Вырожденность – одну аминокислоту кодировать
могут (от 2 до 6) триплетов.
Универсальность – у всех видов организмов
биологический код одинаков
Колинеарность – последовательность кодонов в
зрелой мРНК соответсвтует последовательности
аминоксилот в синтезированном белке
100.
Код непрерывен, отсутствие «знаков препинания», т.е.
сигналов, указывающих на конец одного кодона и начало
другого.
Другими словами, код является линейным,
однонаправленным и непрерывающимся: АЦГУЦГАЦЦ.
101.
Вторичнаяструктура
тРНК
Третичная
структура тРНК
102.
Значение активирования АК:аатРНК - это транспортная форма АК
Каждая АК получила свой шифр в виде
триплета в антикодоновой петле тРНК
(который комплементарен кодону
мРНК).
Считывание информации при
биосинтезе белка идет путем
кодон-антикодонового
взаимодействия
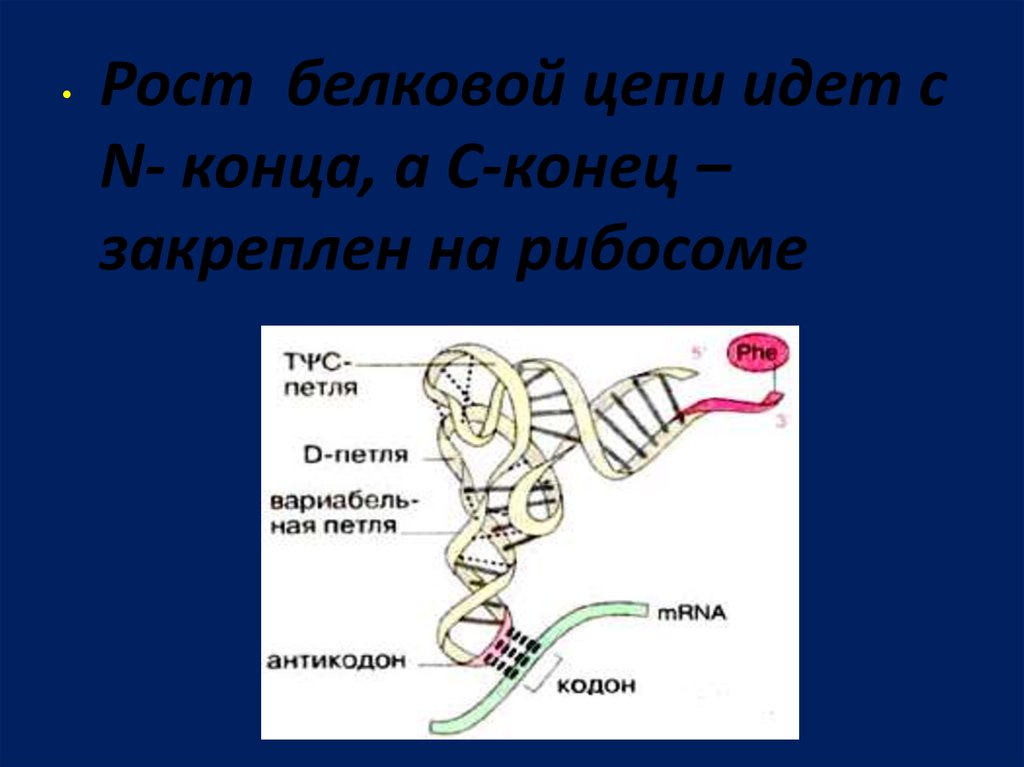
103.
Рост белковой цепи идет с
N- конца, а С-конец –
закреплен на рибосоме
104.
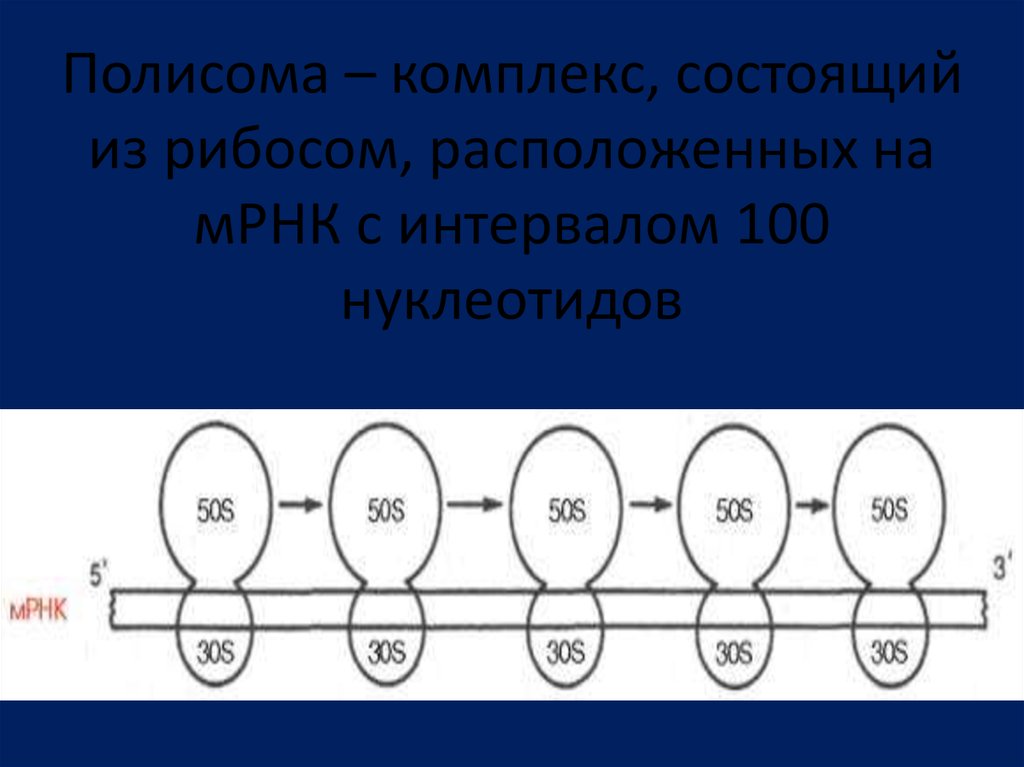
Полисома – комплекс, состоящийиз рибосом, расположенных на
мРНК с интервалом 100
нуклеотидов
105.
Функции субъединиц рибосомы впроцессе трансляции:
Малая субъединица присоединяет мРНК и
декодирует информацию с помощью тРНК
и механизма транслокации.
Большая субъединица ответственна за
образование пептидных связей.
рРНК вносит основной вклад в организацию и
проявление пептидилтрансферазной
активности.
106.
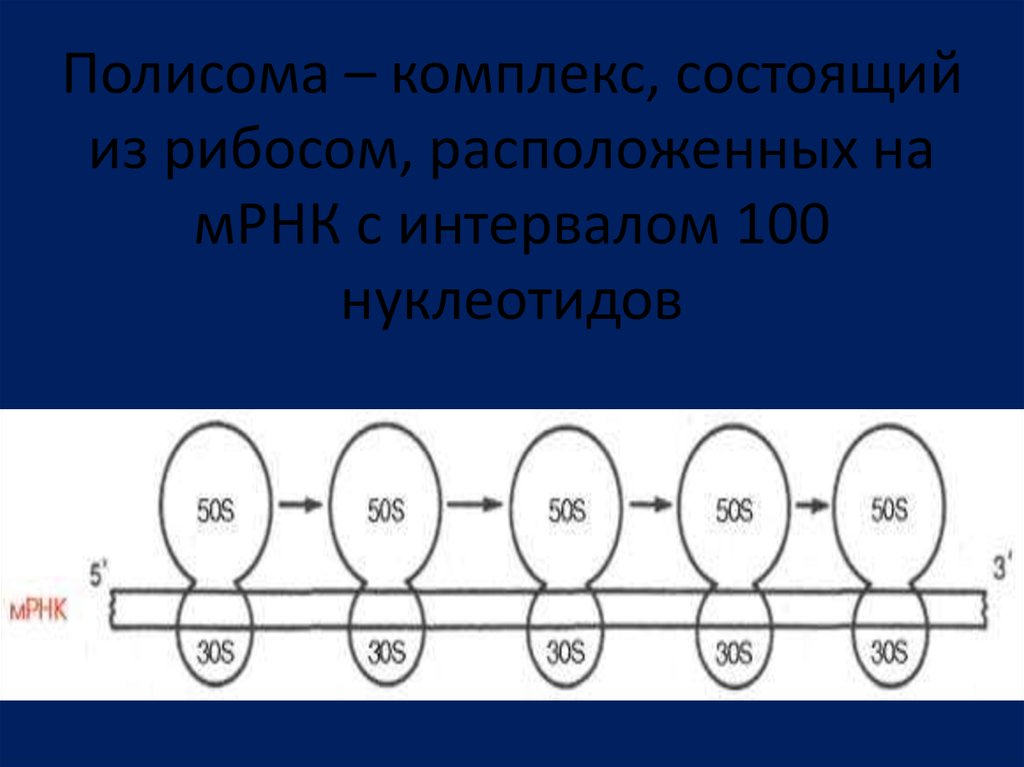
Полисома – комплекс, состоящийиз рибосом, расположенных на
мРНК с интервалом 100
нуклеотидов