
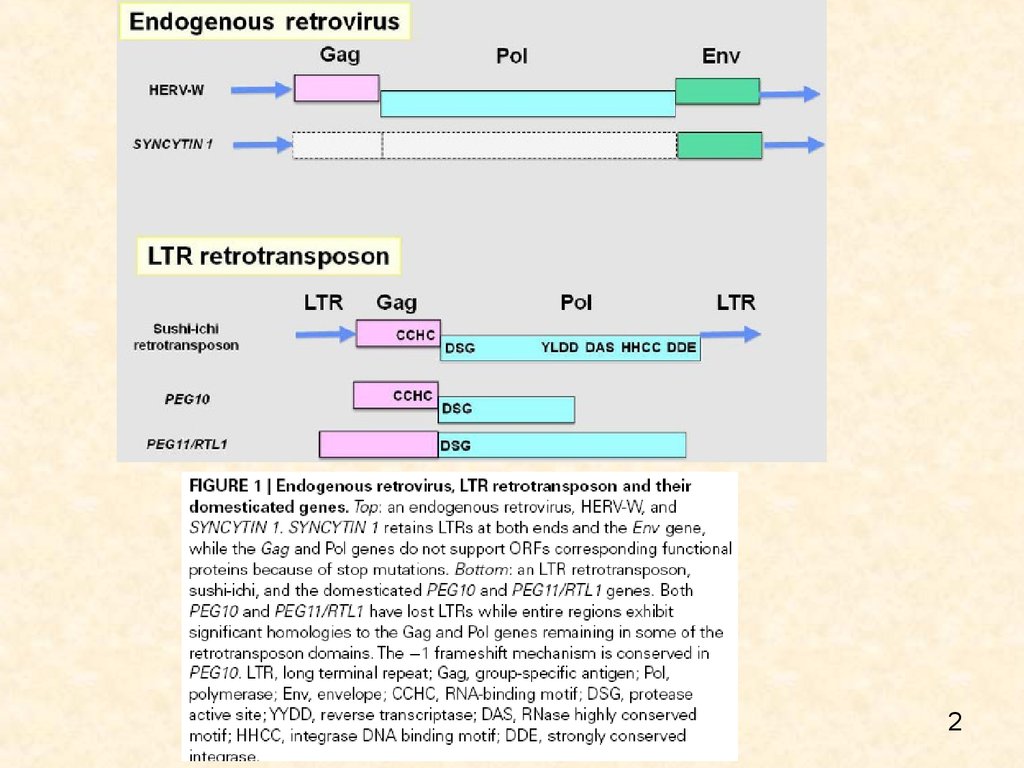
































































































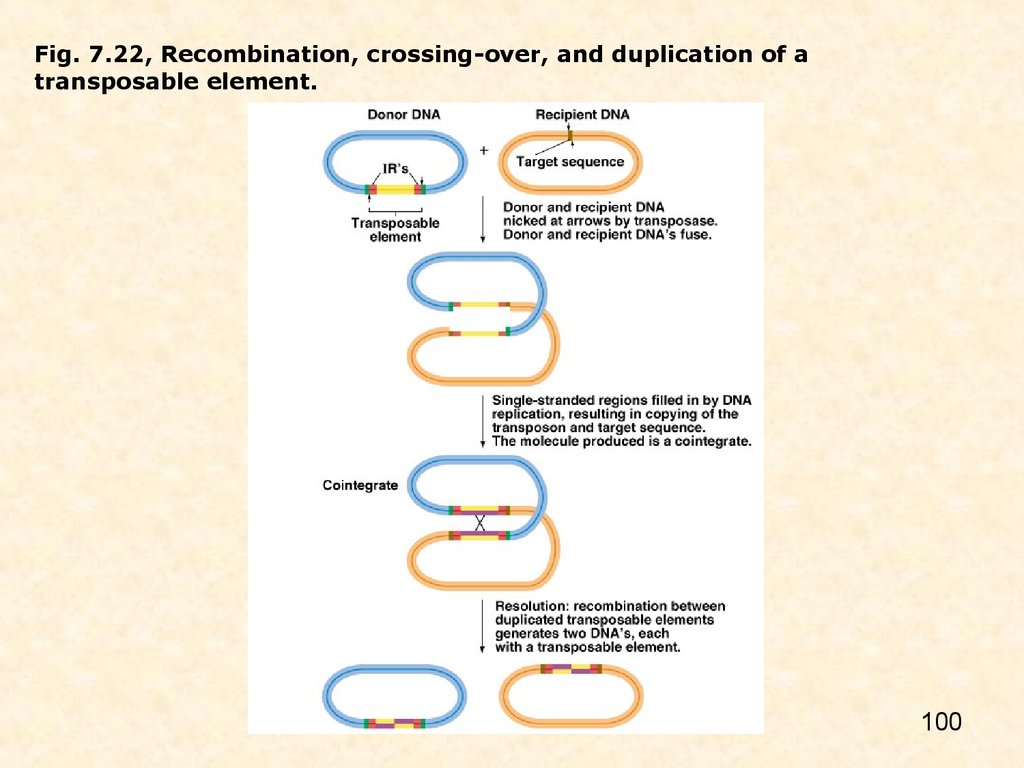




















 Биология
БиологияПохожие презентации:










Ретротранспозоны и ДНК транспозоны
1. Ретротранспозоны и ДНК транспозоны
12.
23.
34.
45. Мобильные генетические элементы
• В 40-х годах Барбара Мак-Клинток, американскийгенетик, обнаружила мозаичность окраски зерен у
кукурузы, небъяснимую законами Менделя и
мутационной теорией.
• Она предположила, что некоторые гены могут
менять свое место в геноме. В 70-х годах это
предположение получило экспериментальное
подтверждение.
5
6.
Первое описание элементов геномной ДНК, характеризующихся
наследуемой способностью передвигаться внутри и между
геномами, было сделано в 1951 г. прошлого века Барбарой МакКлинток для кукурузы. Благодаря развитию технологий
геномного секвенирования, к настоящему времени эти мобильные
генетические элементы (МГЭ) обнаружены в геномах самых
разных видов, часто получая названия, отражающие их
способность к передвижению (hobo – бродяга, gypsy – цыган, flea –
блоха и т.д.). Содержание МГЭ варьирует и может достигать 85%
(Zea mays) генома, причем максимальное разнообразие МГЭ
демонстрируют беспозвоночные. В малых геномах число копий
МГЭ на семейство низкое, но в больших геномах оно значительно
варьирует.
6
7.
• Все МГЭ по способу перемещения могут бытьсгруппированы в два основных класса:
1 - ретротранспозоны, перемещающиеся с помощью РНК-посредника
(«copy and paste»), и
2 - ДНК транспозоны (перемещается непосредственно ДНК).
Ретротранспозоны обнаружены у всех эукариот, от грибов до
млекопитающих и, по-видимому, эти МГЭ связаны с половым
размножением.
ДНК транспозоны – самая древняя группа МГЭ, распространенная
среди про- и эукариот.
Интересно, что МГЭ первого класса были обнаружены как у
плоских, так и круглых червей, а МГЭ второго класса оказались
типичными для паразитических нематод, но не для трематод и
ленточных червей. К настоящему времени только у гельминтов
млекопитающих описано более 30 видов МГЭ.
7
8.
Оба класса делятся последовательно на подклассы, суперсемейства исемейства, согласно механизму их интеграции.
Ретротранспозоны обнаружены у всех эукариот от грибов до млекопитающих; они
включают
- LTR-ретротранспозоны,
- non-LTR ретротранспозоны и
- тирозин-рекомбинантные ретротранспозоны.
• LTR ретротранспозоны объединяют пять групп: Copia, BEL, Gypsy, эндогенные
ретровирусы (ERV) и DIRS.
• non-LTR ретротранспозоны включают автономные LINES (перемещающиеся
самостоятельно) и неавтономные SINES (требующие для своего перемещения
наличия автономных МГЭ) элементы.
• Ретротранспозоны, по-видимому, связаны с половым размножением, поскольку
среди представителей 24 типов животных они не были обнаружены только у
размножающихся бесполым путем коловраток.
• Для своей экспансии ретротранспозоны вначале транскрибируются РНК
полимеразой хозяина, а затем копируются собственной обратной транскриптазой.
Поскольку оба фермента не обладают редактирующей способностью, эти копии
имеют тенденцию приобретения случайных замен оснований. Такие замены часто
инактивируют последовательности-потомки и приводят к утрате их мобильности
(“dead-on-arrival”).
• ДНК транспозоны – самая древняя группа МГЭ, распространенная среди про- и
эукариот; по механизмам транспозиции подразделяется на «cut-and-paste»
транспозоны, самореплицирующиеся транспозоны и транспозоны «катящегося
кольца» (полурепликативная транспозиция).
8
9.
Мутации, создаваемые инсерцией ретротранспозонов, более
стабильны, чем мутации, вызванные ДНК транспозонами,
поскольку в отличие от последних, ретротранспозоны,
встроившись в геном уже никогда не исчезают.
Передвигаясь по геному и интегрируя в новые сайты, МГЭ могут
обеспечить потенциально вредную генетическую изменчивость,
от полиморфизма простой последовательности, до кардинальных
изменений в структуре хромосом, их целостности и инактивации
соседних генов. Показано, например, что все non-LTR
ретротранспозоны или ретротранспозон-подобные
последовательности трипаносом T. brucei и T. cruzi, а также
лейшмании Leishmania major имеют тенденцию локализации в
местах хромосомных инверсий, «strand-switch» регионов и
хромосомных концов.
9
10.
Вырезание и встраивание МГЭ индуцирует мутагенез, уровень
которого может в тысячу раз превышать скорость спонтанных
мутаций. Поэтому для подавления неконтролируемой
пролиферации МГЭ сформировались специальные защитные
механизмы против их транспозиции, включающие
эпигенетические процессы, такие как система
транскрипционного и посттранскрипционного сайленсинга генов.
Однако различные виды физиологического стресса, например,
температурные изменения или колонизация новых мест обитания
хозяевами, могут активировать МГЭ и привести к их
пролиферации, разрушив эпигенетический сайленсинг.
С другой стороны, есть данные, предполагающие коэволюцию
МГЭ и геномов хозяев, положительное влияние и важную роль
этих элементов в формировании и поддержании хромосом
хозяина, модификации генной экспрессии и участии в разделении
генома на хромосомные домены с эпигенетическими метками.
10
11. МГЭ и трансгенные технологии
Накопление знаний о структуре и активности МГЭ может
пролить свет на эволюционную историю и филогенетические
отношения самих элементов и генома хозяина (в частности, дать
информацию об эволюции генов в сайтах интеграции МГЭ), а
также будет способствовать развитию трансгенных технологий,
полезных для изучения структуры, функции и контроля генов
паразитических видов.
Виды Platyhelminthes рассматриваются в настоящее время как
объекты трансфекции для изучения тканевой регенерации,
дизайна новых и более эффективных антигельминтных
препаратов и исследования молекулярных связей между
паразитом и хозяином. Недавно, при изучении регенеративных
процессов и МГЭ у саламандры Ambyostoma mexicanum и
голотурии Holothuria glaberrima, была высказана гипотеза, что
транскрипционная активность ретротранспозонов может
специфически контролироваться хозяином и играть
определенную роль в регенерации органов животных.
11
12.
Способность свободноживущих плоских червей к регенерации
значительных участков тела (подразумевающая присутствие
зародышевых клеток) привлекает внимание исследователей на
протяжении нескольких последних десятилетий. Понимание
основ регенирации у животных, в том числе плоских червей,
может внести важный вклад в регенеративную медицину
человека. Планарии, свободноживущие филогенетические
родственники трематод, способны регенирировать целых червей
из небольшого фрагмента их тела. Клетки, ответственные за эти
регенеративные свойства, были названы «необластами».
• Однако для изучения данного феномена необходимы новые более
эффективные методы для интродукции чужеродных генов и
векторы для стабильной трансфекции. Методом электропарации
в геном необластов планарии Dugesia japonica были эффективно
интродуцированы три транспозона: mariner, Herms и Piggybac, два
из которых хорошо экспрессировались через восемь месяцев
после трансфекции. Содержание паразитических видов червей в
лабораторных условиях, а также генетические манипуляции с
ними осуществлять сложнее. Тем не менее, уже известны
успешные эксперименты по получению генетически
модифицированных гельминтов - трематод, цестод и нематод.
Практическое применение данных исследований лежит в сфере
сайленсинга генов, вовлеченных в жизненно важные
физиологические процессы, посредством РНК интерференции.
12
13.
Помимо фундаментального значения, практический интерес
может иметь способность ретроэлементов выполнять важные
функции в эмбриогенезе, в частности, влиять на
плюрипотентность клеток.
Ретротранспозоны часто видо- или родоспецифичны, что
открывает перспективу разработки чувствительных и
специфичных молекулярных диагностикумов, особенно
необходимых для паразитических видов.
Кроме того, изучение МГЭ представляет определенный интерес
для эпидемиологических и медицинских исследований, поскольку
они могут генерировать специфические генетические паттерны,
ассоциированные с клиническими проявлениями заболеваний.
13
14.
Как членов фракции повторов, МГЭ вначале считали
«мусорной», «эгоистичной» ДНК, или «геномными
паразитами». Однако накопление весомых свидетельств о
важной роли МГЭ в эволюции генов и геномов приводит к
пониманию того обстоятельства, что помимо вредных
эффектов эти геномные элементы вносят позитивный вклад в
эволюцию генома их хозяев и поэтому могут быть
переименованы в «потенциально полезные
доместицированные элементы».
14
15.
• Геномная доместикация может приводить ксущественной функциональной эволюции МГЭ.
- Яркий пример – влияние этих элементов на эволюцию адаптивной
иммунной системы челюстных позвоночных. Домен Rag1,
участвующий в обеспечении образования огромного разнообразия
антител, высоко гомологичен транспозазе ДНК транспозонов
transib, которые были обнаружены в геномах беспозвоночных, но
не идентифицированы у позвоночных.
- Предполагают, что ретротранспозон LITc генома трипаносомы
Trypanosoma cruzi может участвовать в репарации ДНК, а
- ген SETMAR, участвующий в каскаде репараций двойных разрывов
ДНК, произошел от транспозазы mariner.
- Есть основания полагать, что теломераза, по-видимому, произошла
от обратной транскриптазы ретротранспозонов.
- Считается, что доместицированные гены ретроэлементов PNMA,
SIRH и SYNCYTIN, необходимые для нормального развития
плаценты, значительно повлияли на радиацию млекопитающих.
- Наконец, описано большое число генов, произошедших от
транспозонов, которые приобрели функцию регуляторов
транскрипции.
15
16. МГЭ и горизонтальный перенос генов
Мобильные элементы генома являются основными кандидатами
на роль переносчиков в рамках гипотезы горизонтального
переноса ДНК (Horizontal Gene Transfer, HGT) между
репродуктивно изолированными видами. События HGT
рассматриваются сейчас, как важная движущая сила эволюции
эукариотического генома. Причем считается, что система
паразит-хозяин является оптимальной моделью для изучения
данного феномена. Приобретение МГЭ de novo считается
доказанным, если гомология первичных последовательностей в
геномах неродственных организмов составляет более 90%.
16
17.
В геноме F. hepatica среди ретротранспозонов обнаружен повтор
BovB (67 тыс. полных или частичных копий). Ранее для него был
описан горизонтальный перенос между змеями и жвачными, а
наличие данного элемента у F. hepatica предполагает, что перенос
BovB между различными таксонами позвоночных может
усиливаться трематодной инфекцией.
В геноме анкилостомы Ancylostoma caninum содержится ДНК
транспозон bandit. Его ближайшим филогенетическим
родственником является ДНК транспозон человека Hsmar, что
предполагает возможность горизонтального переноса между
анкилостомами и их хозяевами-млекопитающими.
17
18.
Имеющийся объем знаний по изучению HGT позволяет сделать
ряд важных заключений:
(1) для HGT не требуется долговременной физической ассоциации
между видами, но физический контакт может увеличить
вероятность HGT;
(2) смена хозяев может затруднить идентификацию HGT,
ассоциированную с паразитизмом;
(3) обнаружение МГЭ часто идентифицируется как HGT, однако
следует проверять, нет ли первичного механизма, обусловившего
его распространение;
(4) HGT часто подвергается сомнению из-за потенциально
возможной контаминации, и эта проблема не должна оставаться
без внимания.
Дальнейшие исследования HGT могут в значительной мере
пополнить наши представления о геномной коэволюции и дать
эволюционое понимание эпидемиологии паразитарных инфекций.
18
19.
1920.
2021. Ретроэлементы
• Ретроэлементы (RE) составляют значительную часть(до 50%) генома человека. RE несут в себе большое
количество различных регуляторных элементов и
способны перемещаться по геному при помощи
ретротранспозиции. Ретроэлементы способны
оказывать влияние как на экспрессию близлежащих
генов, так и на функционирование генома в целом.
В геноме человека обнаружено большое количество
интеграций RE, полиморфных в различных
человеческих популяциях.
21
22.
Полиморфные интеграции ретроэлементов все шире используютсяв качестве молекулярно-генетических маркеров (МГМ)
в популяционно- и медико-генетических исследованиях,
например для определения генетических взаимоотношений между
отдельными популяциями или ассоциативного анализа
интеграционного полиморфизма с развитием некоторых
заболеваний. МГМ на основе RE обладают рядом преимуществ:
1) присутствие инсерции ретроэлемента в данном геномном локусе
всегда говорит об общем происхождении, так как вероятность
независимой интеграции RE в точно такое же место генома
исчезающе мала;
2) инсерции RE, как правило, достаточно стабильны, так как в
геноме не существует специального механизма для бесследного
удаления интеграций;
3) из всех известных типов полиморфизмов инсерционная
вариабельность ретроэлементов наиболее легко детектируется
экспериментально.
22
23.
• LINE• Полноразмерные LINE элементы (Long Interspersed
Nuclear Elements) имеют длину 4-6 тыс. п. о. и содержат
две открытые рамки считывания (ORF - Open Reading
Frame) и поли А последовательность на 3' конце.
Нуклеотидная последовательность полноразмерного
LINE кодирует ферменты, необходимые для их
размножения, таким образом, этот класс элементов
считается автономным. По краям LINE элементов
имеется короткий прямой повтор длиной 7-20 п. о.,
представляющий собой дупликацию сайта мишени.
23
24.
SINEВ отличие от LINE нуклеотидная последовательность
SINE (Short Interspersed Nuclear Elements) элементов
лишена каких-либо ORF. В ее состав входит
внутренний промотор для РНК-полимеразы III, поли А
последовательность на 3' конце и короткий прямой
повтор такой же длины как у LINE по краям. Для
размножения они используют ретротранспозиционный
аппарат LINE элементов.
В геноме приматов обнаружена немногочисленная
группа ретроэлементов SINE-R/SVA, имеющих
смешанное происхождение, но размножающиеся при
помощи ретротранспозиционного аппарата LINE.
24
25.
SINEсоставляют около 13-14% человеческого генома.
Этот класс ретроэлементов включает в себя неавтономные ретротранспозоны длиной менее
500 п. о., имеющие в своем составе промотор и поли А последовательность, и лишенные
каких-либо открытых рамок считывания.
В человеческом геноме, как и в геноме других приматов, SINE
представлены в основном двумя семействами:
• MIR (Mammalian-wide Interspersed Repeats) и
• Alu.
Семейство MIR представлено в геноме человека 300 - 400 тыс.
копий, что составляет около 2,5% генома. Как и многие
другие SINE элементы, обнаруженные в геномах многих
эукариотических организмов, MIR произошли от тРНК.
Считается, что представители семейства MIR активно
размножались в геноме еще до расхождения основных
эволюционных ветвей млекопитающих.
25
26.
AluСемейство Alu представлено более чем миллионом копий
в человеческом геноме, что составляет порядка 10 %
всех геномных последовательностей. Полноразмерный Alu
элемент имеет длину около 300 п. о. и представляет собой гомодимер. В 5'
концевой части Alu элемента находится расщепленный промотор,
узнаваемый РНК полимеразой III и состоящий из боксов А и В. В средней
части ретроэлемента содержится короткая А/Т богатая
последовательность, а в 3' концевой части поли А последовательность
различной длины. В отличии от большинства других
семейств SINE, ведущих свое начало преимущественно
от тРНК, Alu элементы произошли от 7SL РНК,
входящей в состав рибонуклеопротеиновых частиц
SRP (Signal Recognition Particles).
26
27.
Наиболее известными повторами семейства SINE в
геноме грызунов являются B1 и B2 семейства.
27
28.
Схема строения Alu элемента.А и В - бокс А и бокс В - части расщепленного промотора для РНК
полимеразы III;
А5ТАСА6 последовательность, разделяющая две половины
элемента;
(А)n - поли А последовательность на 3' конце. Белые треугольники
обозначают дупликации сайта мишени в результате интеграции.
28
29.
Все Alu-повторы разделяются на три основныхсемейства, различающиеся по времени возникновения,
и обозначающиеся заглавными латинскими буквами в
алфавитном порядке:
старое (junk, J),
среднее (standard, S) и
молодое (young, Y).
Каждое семейство подразделяется
на ряд подсемейств. AluJ разделяют на два подсемейства
AluJo и AluJb. Последовательность AluJo наиболее
близка к 7SL RNA. S подсемейства Sx, Sp, Sq, Sg и Sc средние по возрасту подсемейства Alu возникли от 31 до
48 миллионов лет назад.
29
30.
Времявозникновения и
представленность
основных семейств
Alu.
Ряд молодых
подсемейств AluY,
например, Yb9,
Ya5a2, Yc1 и др.,
является человек
специфическим.
Некоторые
интеграции
представителей
молодых
подсемейств Alu
остаются
полиморфными в
популяциях
человека.
30
31. LINE
В геноме встречаются в основном 3 класса LINE:L1 (LINE1)
L2 (LINE2)
L3 (LINE3)
Полноразмерные представители LINE имеют длину
несколько тысяч п. о. и включают в свой состав две
ORF, однако большинство содержащихся в геноме
копий укорочены со стороны 5' конца. К LINE
элементам можно отнести также HAL1 элементы,
имеющие только одну ORF, сходную по структуре с
ORF1. Считается, что LINE элементы возникли еще в
докембрии и являются ровесниками эукариот, так как
встречаются в геномах практически всех
эукариотических организмов. L2 и L3 очень "старые"
группы, которые были активны еще до возникновения
млекопитающих. Считается, что последние
представители L2 были активны 80-100 млн. лет назад
и принимали участие в размножении MIR элементов. 31
32.
LINE-последовательности заключают в себе гены обратных
транскриптаз, что является признаком ретротранспозонов (мобильных
генетических элементов животных, обладающих структурным сходством
с геномом ретровирусов ), но для них характерно отсутствие
последовательностей длинных концевых повторов ( LTR - long terminal
repeats), типичных для ретротранспозонов.
• В качестве примера LINE-последовательности можно
упомянуть LINE1 повтор мыши и L1-повтор человека.
Длинные повторы изначально были описаны как последовательности
ДНК длиной более 5 kb и представленные более чем 104 копиями на
геном. В геноме человека было обнаружено только одно семейство
повторов LINE - L1. Консенсусная последовательность этих повторов
имеет длину около 6 kb, так называемый poly-A "хвост" на одном конце и
фланкирована короткими прямыми повторами обычно менее 20 н.п.
длиной. Число копий - примерно 100,000 на гаплоидный геном.
Большинство этих повторов усечены на 5'-конце.
32
33.
Семейство L1 ретротранспозонов является наиболеемолодой и широко распространенной группой LINE
элементов. Около полумиллиона копий
представителей L1 образуют не менее 17 %
человеческого генома.
Полноразмерный элемент имеет длину около 6 тысяч п. о. и включает в
свой состав 5' нетранслируемую область (5' UTR - 5' Untranslated Region)
длиной около 900 п. о., содержащую внутренний промотор, ORF1 (длиной
около 1 т. п. о.) и ORF2 (длиной 4 т. п. о.), разделенные спейсером (63 п.
о.), короткую 3' нетранслируемую область (3' UTR - 3' Untranslated
Region) длиной около 200 п. о., за которой следует сигнал
полиаденилирования и поли А последовательность. В геноме содержится
всего лишь около 5000-7000 полноразмерных копий
L1, в то время как остальные копии укорочены со стороны 5' конца.
Благодаря этому средняя длина L1 элемента в геноме составляет около
900 п. о.
Считается, что человеческий геном содержит 60-100
L1, сохранивших способность к автономной
ретротранспозиции.
33
34.
Схема строение LINE 1 элемента5' UTR и 3' UTR - 5' и 3' нетранслируемые области;
ORF1 и ORF2 - открытые рамки считывания LINE;
EN – эндонуклеазный домен, RT - домен с активностью обратной
транскриптазы, С - С-концевой домен белка ORF2p;
Pr - промотор;
ААТААА - сигнал полиаденилирования;
(А)n - поли А последовательность на 3' конце.
Белые треугольники обозначают дупликации сайта мишени в
результате интеграции L1.
34
35. LTR содержащие ретротранспозоны
3536.
Еще одной обширной группой ретроэлементов являются LTRсодержащие ретротранспозоны.
Эта достаточно разнородная группа включает в себя
повторяющиеся элементы, объединяемые наличием LTR,
указывающего на ретровирусное происхождение. При этом
LTR (Long Terminal Repeat) определяется как длинный
концевой повтор, имеющий по краям инвертированную
последовательность TG/CA и содержащий промотор и сайт
полиаденилирования. Все LTR-ретропозоны составляют
около 8 % человеческого генома и представлены 450
тысячами копий.
36
37.
Систематическое исследование и каталогизация базы данныхRepbase, содержащей все обнаруженные повторяющиеся генетические
элементы многих организмов, выявило около 200 семейств
ретровирусных и LTR подобных повторяющихся элементов в
геноме человека.
К этой группе также относится суперсемейство MaLR
(Mammalian apparent LTR retrotransposons) с наиболее
многочисленным семейством THE-1.
Последовательность MaLR, находящаяся между двумя LTR, содержит
единственную ORF.
Для некоторых MaLR показана гомология их единственной ORF либо
ретровирусному гену pol, либо gag, в то время как остальные лишены
такого структурного сходства с HERV, что затрудняет объяснение их
происхождения. По-видимому, MaLR возникли 80-100 млн. лет
назад, о чем свидетельствует их наличие в геномах млекопитающих
различных отрядов, таких как грызуны, парнокопытные и др.
37
38.
Эндогенные ретровирусы (HERVs-Human EndogenousRetroviruses) - это повторяющиеся мобильные
генетические элементы геномов млекопитающих,
сходные по структуре с интегрированной формой
персистирующих экзогенных ретровирусов.
По современным представлениям HERV являются
отпечатками древних экзогенных ретровирусов,
заразивших клетки линии зародышевого пути и
закрепившихся в геноме.
38
39.
Строение провируса ERVБелыми прямоугольниками обозначены ORF, соответствующие
ретровирусным генам:
gag - group specific antigen кодирует белок основного капсида (CA), белок
ядерного капсида (NC) и белок матрикса (MA);
prt - кодирует дУТФазу (DU) и протеазу (PR);
pol - полимераза кодирует обратную транскриптазу (RT), РНКазу Н
(RNH) и интегразу (IN);
env - липопротеиновая оболочка кодирует трансмембранный (TM) и
поверхностный (SU) белки.
Черными прямоугольниками обозначен ген corf, кодирующий
вспомогательный белок
CORF. PBS - участок связывания тРНК затравки;
LTR - длинный концевой повтор.
Белые треугольники по краям обозначают дупликации сайта мишени
39
(TSD).
40.
4041.
Мобильные генетические элементы (МГЭ) - это
последовательности нуклеотидов, меняющие свою локализацию
и копийность в геноме.
Выделяют следущие классы МГЭ:
1. IS - вставочные элементы у прокариот.
2. Tn - транспозоны у прокариот.
3. Эписомы у прокариот.
4. Некоторые умеренные фаги.
5. Контролирующие элементы кукурузы.
6. Мобильные диспергированные гены у дрозофилы, мыши,
человека.
7. Провирусы.
В литературе "транспозоны" - все мобильные генетические
элементы. Это элементы генома, которые меняют свое положение
и копийность в геноме.
41
42.
Major Types of Mobile DNA ElementsType
Structural Features
Mechanism of Movement Examples
DNA-MEDIATED TRANSPOSITION
Bacterial
insertion
sequences
(IS
elements)
≈50-bp inverted repeats
flanking region encoding
transposase and, in some,
resolvase
Excision or copying of
DNA and its insertion at
target site
IS1, IS10
Bacterial
transposons
Central antibioticresistance gene flanked by
IS elements
Copying of DNA and its
insertion at target site
Tn9
P element
Eukaryotic
transposons
Inverted repeats flanking
coding region with introns
Excision of DNA and its
insertion at target site
(Drosophila);
Ac and Ds
elements
(corn)
42
43.
Major Types of Mobile DNA ElementsType
Structural Features
Mechanism of Movement Examples
RNA-MEDIATED TRANSPOSITION
≈250- to 600-bp direct
terminal repeats (LTRs)
Viral
flanking region encoding
retrotranspo
reverse transcriptase,
-sons
integrase, and retrovirallike Gag protein
Transcription into RNA
from promoter in left LTR
by RNA polymerase II
followed by reverse
transcription and insertion
at target site
Ty
elements
(yeast);
Copia
elements
(Drosophila)
F and G
elements
Of variable length with a 3′
Nonviral
A/T-rich region; fullretrotranspo
length copy encodes a
-sons
reverse transcriptase
Transcription into RNA
from internal promoter
;folding of transcript to
provide primer for reverse
transcription followed by
insertion at target site
(Drosophila);
LINE and
SINE
elements
(mammals);
Alu
sequences
43
(humans)
44. IS-элементы
•Это самые простые транспозоны. Другое название – инсертосомы.Размер IS-элементов ~1000 п.н. На концах они содержат
инвертированные повторы (~20 п.н.). IS- элементы содержат только
один ген - ген транспозазы, фермента, обеспечивающего
перемещение IS-элемента по геному.
•Транспозаза - это обобщеный термин.
Разные транспозазы работают по-разному, но смысл общий:
транспозаза вырезает ДНК в одном месте и вставляет в другое
место генома.
•Перед геном в IS-элементе имеется промотор, за геном - слабый
терминатор транскрипции. Не всегда РНК-полимераза
останавливается на нем, она может продолжать транскрибировать
и рядом стоящий участок генома до сильного терминатора.
44
45. Tn-транспозоны
Такие транспозоны содержатпо краям IS-элементы.
• Помимо гена транспозазы Tn-транспозоны содержат один или несколько
генов лекарственной устойчивости. Копийность транспозона возрастает
при наличии провокационного фона (например, наличие в среде
антибиотика, ген устойчивости к которому кодируется в транспозоне).
• Есть транспозоны, не содержащие гена транспозазы. Такие транспозоны
содержат по краям IS-элементы. Размер транспозонов - 2.5-10 тыс. п. н.
• Всем транспозонам свойственно наличие прямых повторов, LTR (long
terminal repeats - длинные концевые повторы)
• После перехода из основной ДНК в плазмиду транспозон может попасть с
ней в другую бактерию, придавая новому хозяину ранее отсутствующую
лекарственную устойчивость. Кроме того, при вырезании транспозона из
геномной бактериальной ДНК захватываются участки генома одной
бактерии, которые вместе с транспозоном переносятся в другую
бактерию. Захват ДНК происходит, если эта ДНК находится между двумя
45
транспозонами.
46. Умеренные фаги
• Фаги – бактериальные вирусы, осуществляющие инфекционный процесс . Фагможет захватывать часть генетической информации одной бактерии и переносить ее в
другую. Поэтому сегодня говорят о едином генофонде прокариот.
Вирусы
• Некоторые вирусы - на самом деле тоже мобильные элементы. В форме
провируса они находятся в геноме клетки хозяина, а потом могут начать
перемещаться. Это, например, онкорнавирусы, являющиеся ретровирусами.
С открытием ретровирусов стал вопрос о возможности переноса генетического
материала от одного организма к другому без полового процесса. При заражении
вирусом генетический материал может переходить от вида к виду, минуя барьеры
межвидовой изоляции. Такой перенос был назван горизонтальным.
Его могли осуществлять клещи, паразитирующие на разных видах дрозофилы (работа
наподобие иглы со шприцом). Такой перенос может осуществляться, когда хищник
поедает жертву (показано, что ДНК переваривается не до конца, отдельные молекулы
могут попадать из кишечника в клетку и ядро, а затем интегрироваться в хромосому).
Эписомы прокариот
Эписома – плазмида, способная интегрировать в бактериальную
ДНК. Плазмиды способствуют конъюгации бактерий.
46
47. Мобильные элементы эукариот
4748.
• Контролирующие элементы кукурузыГеном кукурузы содержит несколько семейств, члены каждого
семейства могут быть подразделены на два класса:
автономные – способны вырезаться и транспозироваться и
не автономные – передвигаются, если в определенной области
генома есть автономный член того же семейства
• Элементы Ty дрожжей
• Напоминают бактериальные транспозоны. Способны вырезаться
при участии гомологичной рекомбинации; траскрибируются.
48
49. Мобильные элементы дрозофилы
CopiaFB
p
Число
копий
Длина,
п.н
Длина
терминально
го прямого
повтора
Длина
Длина
терминального повтора
инвертированн мишени
ого повтора
20-60
~30
~50 или 0
5 000
500-5
000
500-2
900
276
Нет
Нет
17
250-1 250
31
5
9
?
49
50.
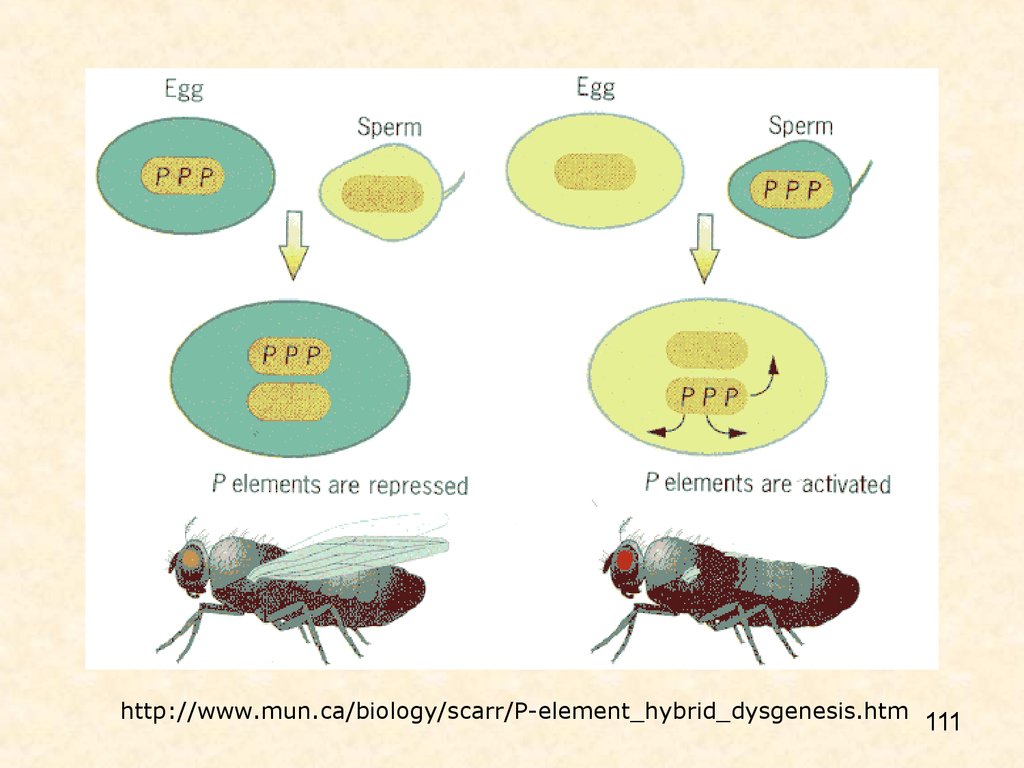
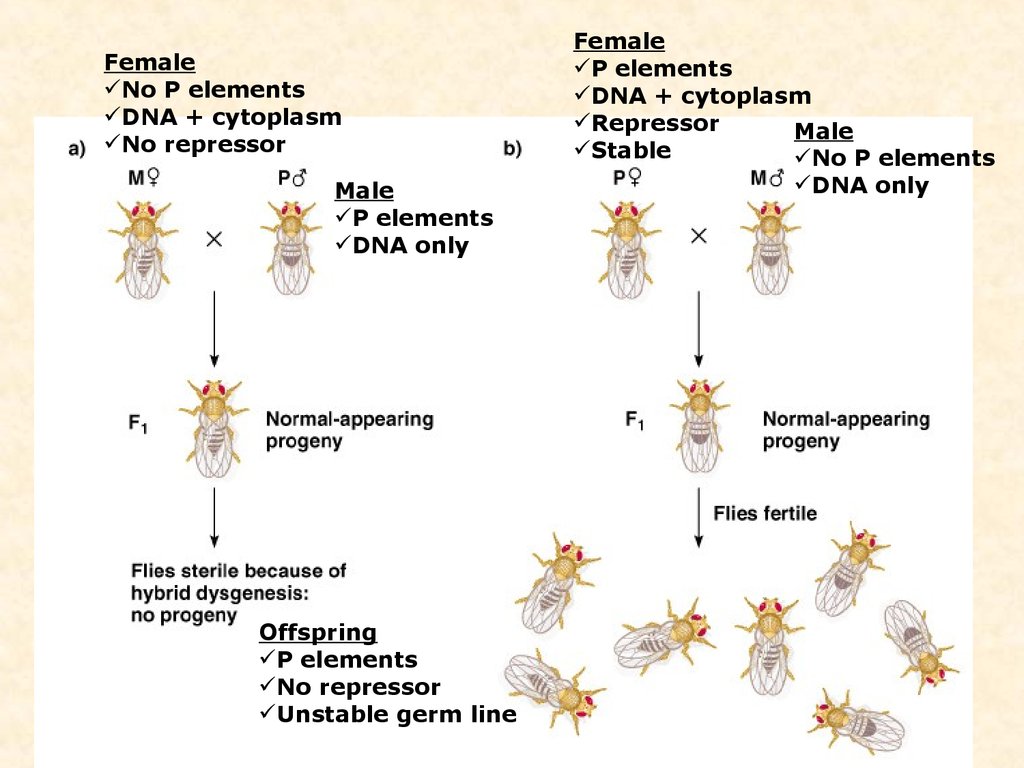
гибридизация при 290С: ♀ норма х ♂ с активным р элементом → все потомствостерильное;
♀ с активным р элементом х ♂ норма → все потомство нормальное
P-elements are transposable elements that carry genes for transposase activity that cause the elements to move, and repressor activity
that prevents expression of transposase.
In a cross between a P-element-carrying female and a laboratory male [left], repressors in the maternally - derived cytoplasm repress
expression of the maternally - inherited P elements. The resulting offspring show the wild-type phenotype.
50
51.
Contribution of mobile genetic elements to the evolution of pathogenic E. coli.
51
52.
The Tn916/Tn1545 Family of Conjugative Transposons
http://www.landesbioscience.com/curie/chapter/5268/
52
53.
Haploid Pokey number inside and outside of rDNA in D. pulex and D. pulicaria. L isolates are from lakes (D.
pulicaria) and P isolates are from ponds (D. pulex). (a) gPokey and rPokey number in each isolate. rPokey is inserted
in 28S genes. gPokey was calculated as (total
Pokey- rPokey). (b) Correlation between gPokey and rPokey. D. pulicaria (-3) refers to the analysis done after
excluding the three isolates with high Pokey load
53
54.
Mobile Genetic Elements in Metazoan Parasites http://www.landesbioscience.com/curie/chapter/3124/
It has become apparent that the schistosome genome is replete with numerous copies of diverse repetitive
sequences. Mobile genetic elements constitute much of this repetitive component of the schistosome genome. Of
these mobile elements, an expanding number of discrete schistosome retrotransposons—long terminal repeat
(LTR) retrotransposons, non-LTR retrotransposons and SINEs—have been identified in recent years. The identity,
structure, phylogenetic relationships, and size contributions of the schistosome non-LTR retrotransposons
reported to date are reviewed here. (Other chapters in this book focus on LTR retrotransposons of schistosomes
[Copeland, Laha, Brindley] and schistosome SINES [Ferbeyre].) These elements include members of the CR1,
RTE and R2 clades, including elements termed SR1, SR2, SR3, pido and Perere-3 and Perere-9 from Schistosoma
mansoni, S. japonicum and S. haematobium. An enhanced understanding of these elements will increase\r\nour
understanding on the evolutionary and phylogenetic relationships of schistosomes and Platyhelminths at large,
will facilitate the annotation of the soon-to-be-reported draft genome sequences of S. mansoni and S. japonicum,
and can be expected to have practical implications including the development of transgenesis vectors.
54
55.
The impact of retrotransposons on
human genome evolution
http://www.nature.com/nrg/journal/v10/n10
/full/nrg2640.html
55
56.
This visualiztion provides
an overview of the
composition of LTR
retrotransposons across
the entire genome, and
quickly allows the user to
see that LTR
retrotransposons have
preferentially accumlated
in pericentromeric
heterochromatin.
This image illustrates an
overview of 1) the distribution
of the LTR retrotransposons
coverage for the 10
chromosomes in the maize
genome, 2) the histogram of
the percent coverage of all 1
MB bins in the genome, 3) the
empiriculative cumulative
distirbution coverage for these
data, and 4) color assignment
of these data into ten color
classes using an euqal interval
clustering approach. The
above visualization was
generated by hand in the R
statistical programming
language, and providing a tool
for quick generation of
intuitive visualizations such as
these could facilitate
exploratory spatial data
analysis. Author: James Estill
56
http://gencart.sourceforge.net/
57.
Distribution of
different clades of
non-LTR
retrotransposons
among nematode
Caenorhabditis
elegans, fruit fly
Drosophila
melanogaster,
malaria mosquito
Anopheles
gambiae, sea squirt
Ciona intestinalis,
fungi, and
vertebrata.
http://retrotranspos
ons.org/index_resea
rch_other.html
57
58.
Age and Number of Mobile elementsHumans contain a higher percentage of mobile elements (active and incactive) relative to three other
fully-sequenced genes (Fly, worm, mustard):
Element
Human
Fly
Worm
Arabodopis
LINE/SINE
33.4%
0.7%
0.4%
0.5%
LTR
8.1%
1.5%
0.0%
4.8%
DNA
2.8%
0.7%
5.3%
5.1%
All TEs
44.4%
3.1%
6.5%
10.5%
However, human elements are older.
Even when compared to mice, again human
elements are older.
Hence, relative to certain "model" organisms,
humans have a higher percentage of their
genome from mobile elements, but the vast 58
majority of these are old, no longer functional,
elements.
59.
59Evolutionary Implications of Mobile Elements
60. Структура, механизмы перемещения и роль подвижных элементов в поддержании целостности хромосом
6061.
http://www-old.agr.unipi.it/typo3temp/pics/a46ce
df7ff.jpg
http://sete.nichd.nih.gov/current_research.htm
61
62. Перемещение транспозона
Концы транспозона
(инвертированные повторы)
показаны направленными
навстречу стрелками.
Дочерние нити ДНК после
репликаии изображены
разным цветом
Внизу на схеме направленные
навстречу стрелки указывают
положение транспозона в
районе «красной»двойной
спирали.
Синими стрелками изображен
синтез комплементарных
нитей.
62
63. Перемещение ретротранспозонов
6364. Сохранение концов хромосомы в процессе повторных актов репликации
6465. Устранение повреждений в одной из комплементарных цепей ДНК и залечивание двухнитиевого разрыва ДНК с помощью ретротранспозона
6566. Роль МГЭ в регуляции активности генов и эволюции генома
6667. Нарушение работы гена в результате внедрения подвижного элемента
6768. След, оставляемый транспозоном в районе промотора
6869.
Variegation in sectors of wildflower petals. A wildflower blooms all over the middle
Atlantic states in the USA in late May and June. My neighbors call this wild flox. It
is an invasive plant, but it is pretty when it blooms. It has two predominant flower
colors, purple (A) and white (B). However, a casual examination of the plants
reveals sectored petals at a moderate frequency (C-E). This is a variegating
phenotype of unknown origin. It can produce white sectors on purple petals (C) or69
purple sectors on white petals (D, E).
70.
Three-dimensional structure of the Tn5 transposase in complex with Tn5
transposon DNA. A. The dimer of the Tn5 transposase is shown bound to a fragment of duplex DNA
from the end of the transposon. Alpha helices are green cylinders, beta sheets are yellow-brown, flat arrows
and protein loops are blue wires. The DNA is a duplex of two red wires, one for each strand. B. The DNA is
shown without the protein and with the nucleotides labeled. The end of the DNA at the top of this panel is
oriented into the active site in the middle of the protein in panel A. The structure was determined by Davies
DR, Goryshin IY, Reznikoff WS, Rayment I. (2000) Three-dimensional structure of the Tn5 synaptic
complex transposition intermediate. Science 289:77-85. These images was obtained by downloading the
atomic coordinates from the Molecular Modeling Database at NCBI, viewing them with CN3D 3.0 and
saving static views as screen shots. The file for observing a virtual three-dimensional image is available at
the course website.
70
71.
The mobility of genetic elements called transposable elements (TEs) was discovered half a century ago by
Barbara McClintock. Although she had recognized them as chromosomal controlling elements, for much of the
consequent time TEs were primarily considered as parasites of the host genome. However the recent explosion of
discoveries in the fields of genomics and epigenetics have unambiguously shown the importance of TEs in
genome function and evolution. Bursts of endogenous TEs have been reported in plants with epigenetic
misregulation, revealing the molecular mechanisms underlying their control. We review here the different steps
in TE invasion of the host genome involving epigenetic control and environmental stress responses. As TEs
propagate in plant genomes and attract epigenetic marks, their neo-insertions can lead to the formation of new,
heritable epigenetic variants (epialleles) of genes in their vicinity and impact on host gene regulatory networks.
71
The epigenetic interplay between TE and genes thus plays a crucial role in the TE-host co-evolution.
72.
7273. Перестройки хромосом, обусловленные присутствием в хромосомах одинаковых повторяющихся последовательностей, представленных подвижными
Перестройки хромосом,обусловленные присутствием
в хромосомах одинаковых
повторяющихся
последовательностей,
представленных
подвижными элементами
73
74. Эффекты, вызываемые мобильными элементами
- Внедрение
мобильных элементов внутрь гена приводит к
выключению гена.
- Может нарушаться регуляция гена, если мобильный элемент
внедряется между оператором и цистроном (у мобильного элемента
есть свой промотор).
- Вставка мобильного элемента может привести к экспрессии генов,
которые не должны в данное время работать.
Наличие мобильных элементов является фактором,
способствующим незаконной рекомбинации.
При незаконной рекомбинации перетасовываются гены, не
имеющие отношения друг к другу.
Мобильные элементы провоцируют образование делеций,
инверсий, дупликаций. Все это - хромосомные мутации.
74
75. Обратная транскрипция
Обратная транскрипция - это синтез ДНК по матрице РНК.Обратную транскрипцию обнаружили в 1970 г. Темин, Балтимор, Дульбеко, работавшие с
вирусом саркомы Рауса (ВСР). Этот вирус вызывает саркому у кур. Это онкорнавирус
(oncoRNA) - относится к ретровирусам.
Ретровирусы - это РНК-содержащие вирусы, в жизненный цикл которых входит
стадия образования ДНК обратной транскриптазой и внедрение ее в геном
клетки хозяина в форме провируса.
Предпочтительного места внедрения провируса в геном нет. Это позволяет
отнести его к мобильным генетическим элементам.
В состав ретровируса входят две идентичные молекулы РНК. На 5'-конце имеется Сap, на 3'конце - поли А-хвост. Фермент обратную транскриптазу вирус "носит" c собой.
Геном ретровируса содержит 4 гена: gag - белок нуклеоида,
pol - обратная транскриптаза,
env - белок капсида (оболочки),
onc - онкоген, ответственный за злокачественную трансформацию клетки.
str5 = str3 - (short terminal repeat) короткий концевой повтор;
U5, U3 - уникальные последовательности (U5 - 80 н., U3 - 200 н.);
75
PB (primer binding site) - участок связывания затравки.
76.
7677.
Ретровирусные РНК кончаются прямыми повторами (R), а свободные линейные
ДНК-последовательностями LTR; концы провирусов представляют собой LTRпоследовательности, укороченные на два основания.
77
78.
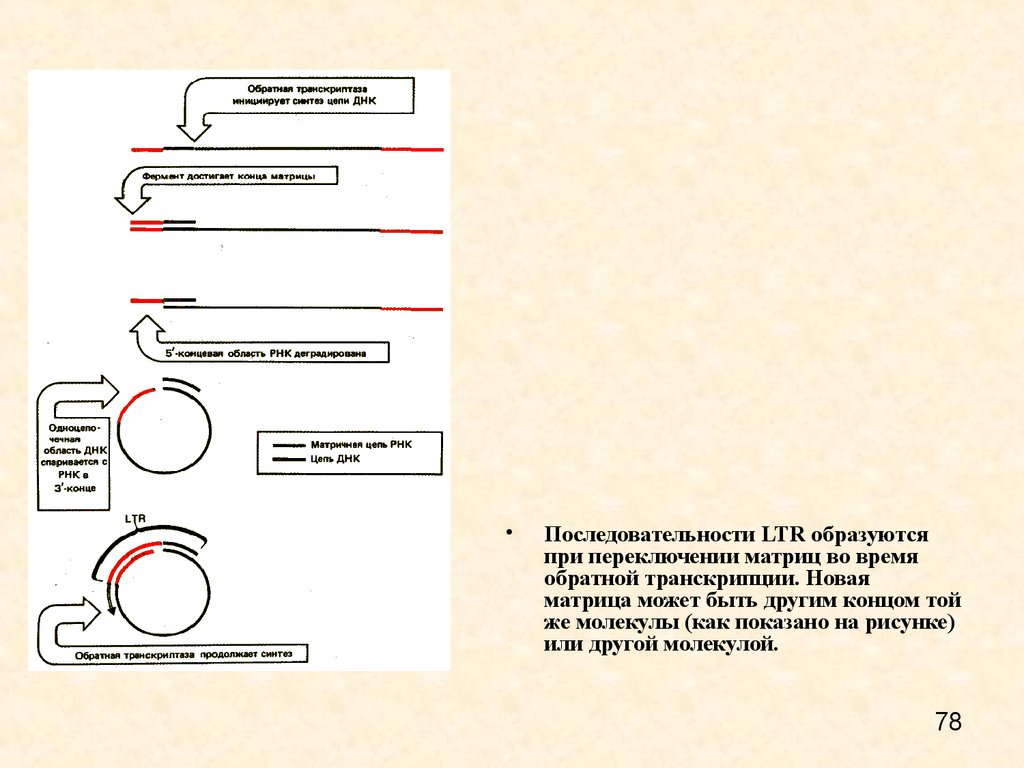
Последовательности LTR образуются
при переключении матриц во время
обратной транскрипции. Новая
матрица может быть другим концом той
же молекулы (как показано на рисунке)
или другой молекулой.
78
79. у дефектных в отношении репликации трансформирующих вирусов часть вирусной последовательности замещена последовательностью клетки. Дли
у дефектных в отношении репликациитрансформирующих вирусов часть
вирусной последовательности
замещена последовательностью
клетки. Длина замещенной
последовательности характерна для
каждого вируса
Интеграция ретровируса сопровождается
образованием двух типов кольцевых молекул
ДНК
79
80.
В отдельных случаях последовательность РНК служит предшественником
геномной последовательности (ДНК). Вероятно, РНК должна была превратиться в
двухцепочечную ДНК, которая затем внедрялась в геном с помощью события,
напоминающего транспозицию. Прямое указание на существование такого пути
получено при изучении ретровирусов, у которых такой путь представляет собой часть
обычного инфекционного цикла. Структура некоторых псевдогенов и других
последовательностей (включая членов семейства Alu) свидетельствует, что
аналогичный путь может быть пройден клеточной последовательностью (достаточно
редко). В обоих случаях внехромосомный элемент дает начало геномной
последовательности.
По всей вероятности, транспозоны не способны к существованию вне генома.
Изменения в относительном соотношении компонентов генома иногда
происходят во время соматического развития. Хорошо известно, например,
Независимо от того, являются ли они компонентами, играющими важную роль в
жизни клетки, или ее паразитами, занятыми только своим собственным
воспроизведением, свободных молекул ДНК они не образуют.
увеличение числа копий определенных генов у личинок насекомых. Благодаря
возможности отбирать варианты клеток с увеличенным числом копий определенного
гена показана случайная амплификация генов в культуре клеток млекопитающих.
Инициируемое внутри генома событие амплификации способствует созданию
дополнительных копий гена, которые существуют либо в составе хромосомы, либо в
форме внехромосомных элементов.
Открытию транспозиции и других перестроек в ДНК в какой-то мере
способствовали наши знания об удивительной способности
последовательностей ДНК приспосабливаться.
80
• Льюин
81.
8182. ЛОКАЛИЗАЦИЯ МОБИЛЬНЫХ ЭЛЕМЕНТОВ В ЛОКУСАХ рДНК
•R – недлинные терминальные повторыретротраннспозоны, в то время как Pokey
являются ДНК опосредованным элементом.
• R2, R4, R5 и R8 найдены во многих
линиях животных, а R1, R6, R7 и RT
представляют независимые линии элементов
у артропод; они перемещаются иногда за
пределы локусов рРНК.
•R1 и R2 обнаружены в большом
колич