
 является:")











 необходимы:")


")































")
")



















")























 Биология
Биология Химия
ХимияПохожие презентации:

. Основы молекулярной генетики")





")

Матричные биосинтезы
1. КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ КАФЕДРА ФУНДАМЕНТАЛЬНОЙ И КЛИНИЧЕСКОЙ БИОХИМИИ
Лекция по биохимииТема:
«Матричные
биосинтезы»
Краснодар
2016
2.
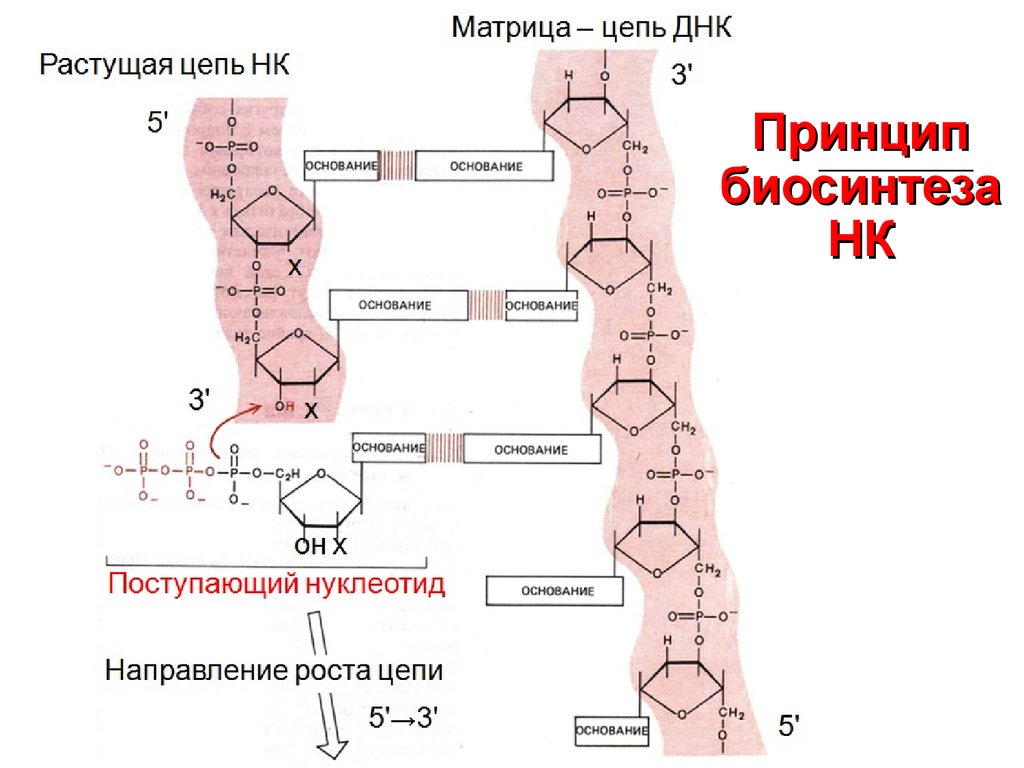
Матрица – цепь ДНКРастущая цепь НК
3'
Принцип
биосинтеза
НК
5'
х
3'
х
х
Поступающийоннуклеотид
Направление роста цепи
5'
→3'
5'
3. Биосинтез ДНК (репликация) является:
матричным(матрица – обе нити ДНК)
комплиментарным
фрагментарным (нити ДНК
синтезируются в виде фрагментов,
которые затем соединяются между
собой)
полуконсервативным (в каждой из
образовавшихся молекул ДНК одна
нить исходная – материнская, а одна –
вновь синтезированная – дочерняя)
4. Полуконсервативность биосинтеза ДНК
5.
6. Особенности репликации
7. Этапы репликации
1. Инициация:Топоизомераза находит точку начала
репликации, гидролизует одну
фосфодиэфирную связь и даёт возможность
компонентам репликативной системы
разомкнуть нити ДНК и образовать
репликативную «вилку», а затем вновь
соединяет связь между мононуклеотидами
Хеликаза разрывает водородные связи
между нитями ДНК
8.
Этапы репликацииДНК-связывающие белки (SSB-белки)
стабилизируют репликативную вилку, не давая
восстанавливаться водородным связям между
комплиментарными нуклеотидами
ДНК-полимераза α (праймаза) строит праймер
(«затравку») из 8-10 рибонуклео-тидов и 4050 дезоксирибонуклеотидов, а ДНКполимераза δ достраивает нить из
дезоксирибонуклеотидов на лидирующей нити,
а ДНК-полимераза ε – на отстающей нити ДНК
9. Инициация репликации
ДНКполимераза αДНКполимераза δ
ДНК-полимераза ε
10. Инициация репликации
Ориджинрепликации
репликон
11. Этапы репликации
2. ЭлонгацияДНК-полимераза
δ продолжает
удлинять нить из
дезоксирибонуклеотидов на
лидирующей нити, а ДНК-полимеразы
α и ε – строить фрагменты из
праймеров и дезоксирибонуклеотидов
(фрагменты Оказаки) на отстающей
нити ДНК по мере движения
репликативной вилки
12. Элонгация репликации
13.
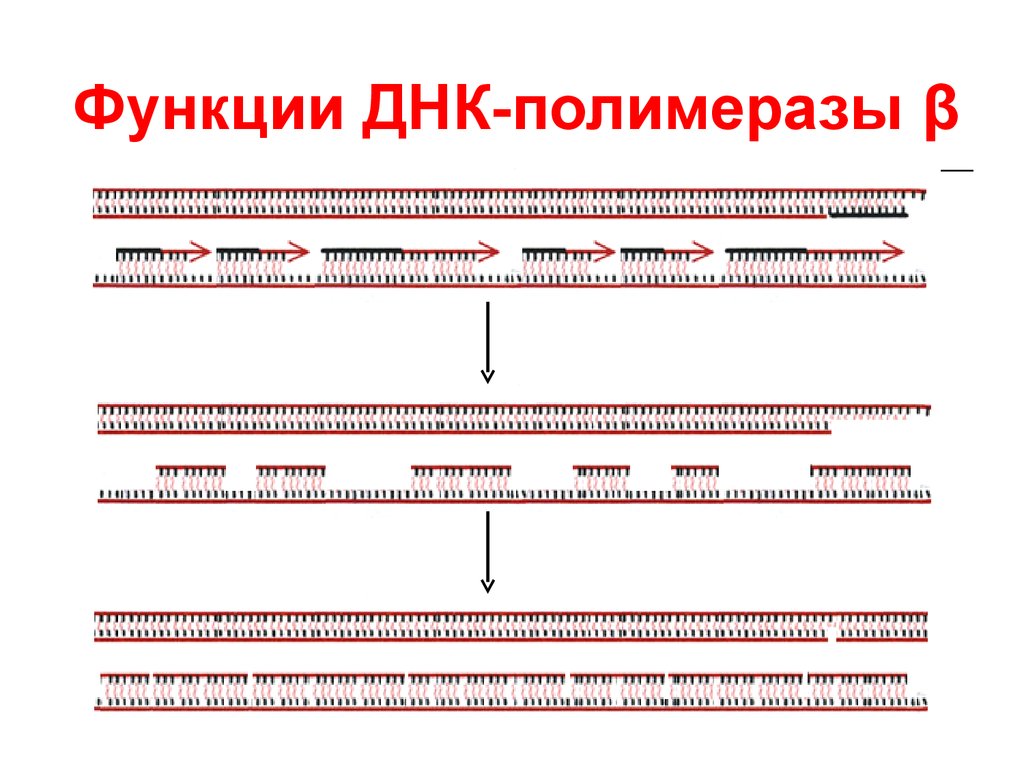
Этапы репликации3. Терминация
ДНК-полимераза
β (фермент
репарации) удаляет праймеры и
достраивает фрагменты ДНК
ДНК-лигаза
соединяет фрагменты
между собой
14.
Функции ДНК-полимеразы β15. Репарация ДНК
16. Для биосинтеза РНК (транскрипции) необходимы:
•МАТРИЦА – участок одной из нитей ДНК– (транскриптон)
•СТРОИТЕЛЬНЫЙ МАТЕРИАЛ:
– АТФ, ГТФ, ЦТФ, УТФ
•ФЕРМЕНТАТИВНЫЕ БЕЛКИ
ДНК-зависимые РНК-полимеразы
I — для синтеза р-РНК
II — для синтеза м-РНК
III — для синтеза т-РНК
•РЕГУЛЯТОРНЫЕ БЕЛКИ: факторы инициации,
элонгации, терминации
17. Биосинтез РНК
18. Биосинтез РНК
РНК-полимеразаБиосинтез РНК
Транскрибируемая нить
ДНК
Нетранскрибируемая нить
мононуклеотиды
пре-РНК (первичный транскрипт)
Сайт терминации
19. Процессинг РНК (1. сплайсинг)
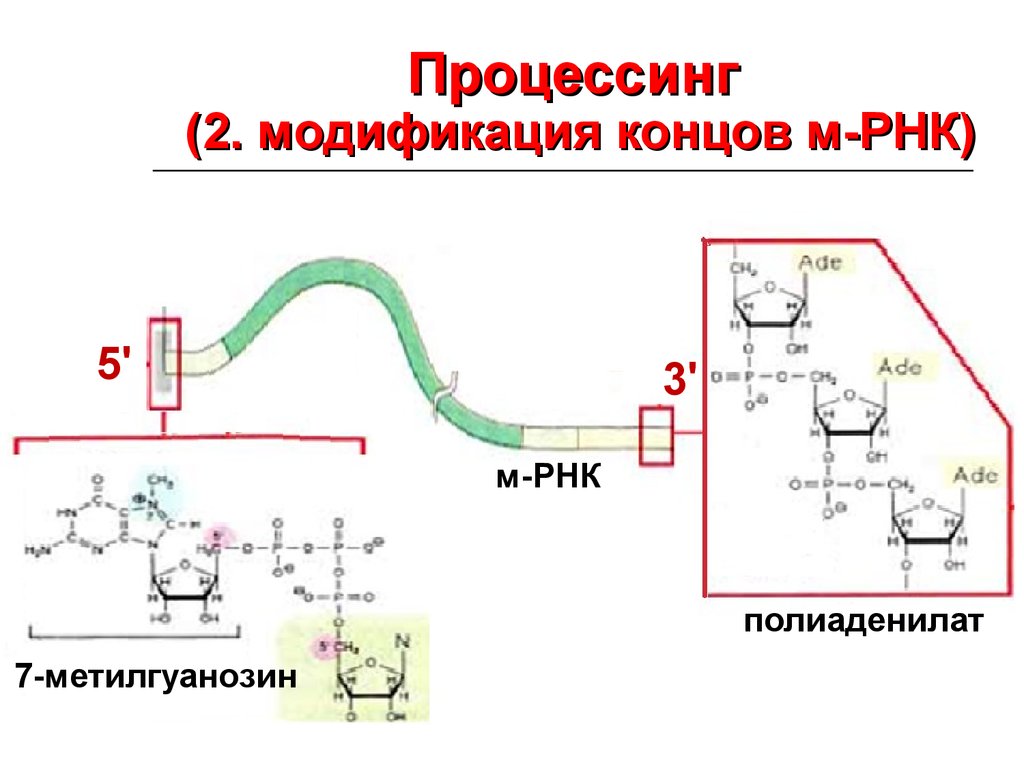
20.
Процессинг(2. модификация концов м-РНК)
5'
3'
м-РНК
полиаденилат
7-метилгуанозин
21. Состав зрелой м-РНК
22. Общая схема биосинтеза белка
23. Компоненты белоксинтезирующей системы
мРНК20 Аминокислот
20 Аминоацил-тРНК синтетаз (АРС-аз)
Изоакцепторные тРНК
Рибосомы в виде полисом
Источники энергии (АТФ, ГТФ) и Мg2+
Белковые факторы регуляции: факторы
инициации, элонгации, терминации
Специальные ферменты посттрансляционного
процессинга полипептидной цепи
24. Ядерные стадии биосинтеза белка
БиосинтезмРНК (транскрипция)
Созревание мРНК
(посттранскрипционный
процессинг)
25. Функции т-РНК
Акцепторная: связывает аминокислоту итранспортирует её к месту синтеза белка
Адаптерная: встраивает аминокислоту на
соответствующее место в образующемся
полипептиде (за счёт взаимодействия
антикодона в т-РНК с кодоном в м-РНК)
26. Строение т-РНК
антикодон27. Таблица генетического кода
28. Характеристика генетического кода
Триплетность (1 аминокислота кодируется 3нуклеотидами)
Специфичность (каждому кодону соответствует только 1
аминокислота)
Вырожденность (1 аминокислота может кодироваться
более чем 1 триплетом)
Линейная запись (прочтение кода «без знаков
препинания»)
Универсальность (одинаков для всех живых существ)
Наряду со значимыми есть и «бессмысленные» кодоны
(терминирующие – УАА, УАГ, УГА)
Колинеарность (соответствие линейной
последовательности кодонов гена и последовательности
АК в кодируемом белке)
29. Колинеарность генетического кода
Глу-тРНК30. Цитоплазматические стадии биосинтеза белка
Активацияаминокислот, или
образование аминоацил-тРНК
Инициация
Элонгация
Терминация
Посттрансляционная
модификация
31. Синтез аминоацил-тРНК
32. Рибосома эукариотов
33. Функции активной рибосомы
Связывание и удерживаниебелоксинтезирующей системы
Транслокация – перемещение м-РНК
через рибосому каждый раз на один
триплет
Замыкание пептидной связи между
аминокислотами (каталитическая
функция)
34. Функционирующая рибосома
35. Образование инициирующего комплекса
36. Инициация:
К малой субъединице рибосомы прикрепляетсям-РНК
К инициирующему кодону прикрепляется Мет-тРНК
К образовавшемуся комплексу присоединяется
большая субъединица таким образом, что Метт-РНК оказывается в пептидильном центре, а в
аминоацильном – кодон, соответствующий
второй аминокислоте
37. Инициация трансляции
38. Элонгация:
В пептидильном центре находится Мет-т-РНК, а ваминоацильном – аа-тРНК, соответствующая второму кодону.
Мет отрывается от т-РНК и перебрасывается в аминоацильный
центр
С помощью пептидилтрансферазы замыкается пептидная связь
между карбоксильной группой Мет и аминогруппой второй
аминокислоты (требуется энергия ГТФ и регуляторный фактор Т)
Происходит передвижение рибосомы по м-РНК на один триплет
(транслокация)
В освободившийся аминоацильный центр входит аа-т-РНК,
соответствующая третьему кодону и т.д.
39.
ГТФГДФ+НР
40. Строение полирибосомы
41. Посттрансляционный процессинг
Модификация N-конца полипептидной цепиФолдинг (формирование пространственной
структуры)
Химическая модификация (гидроксилирование,
гликозилирование и др.)
Присоединение простетических групп (у
гетеропротеинов)
Объединение протомеров при образовании
олигомерных белков
Присоединение сигнальных пептидов для выхода
белка из клетки
42. Регуляция биосинтеза
1: РНК полимераза, 2: Репрессор, 3: Промотор,4: Оператор, 5: Лактоза, 6, 7, 8: гены, кодирующие
лактазу.
43. Действие регуляторных белков
44. Регуляция биосинтеза
Энхансеры – участки ДНК,присоединение к которым
регуляторных белков усиливает
транскрипцию
Сайленсеры – участки ДНК,
присоединение к которым
регуляторных белков уменьшает
транскрипцию
45. Амплификация
46. Типы генов в геноме
Структурные гены (кодируют белки)Регуляторные гены:
Гены-регуляторы (регулируют работу
структурных генов)
Процессинг-гены (регулируют
посттранскрипционные и
посттрансляционный процессинг)
Темпоральные гены (включают в работу
структурные гены в ходе клеточной
дифференцировки)
47. Клеточная дифференцировка
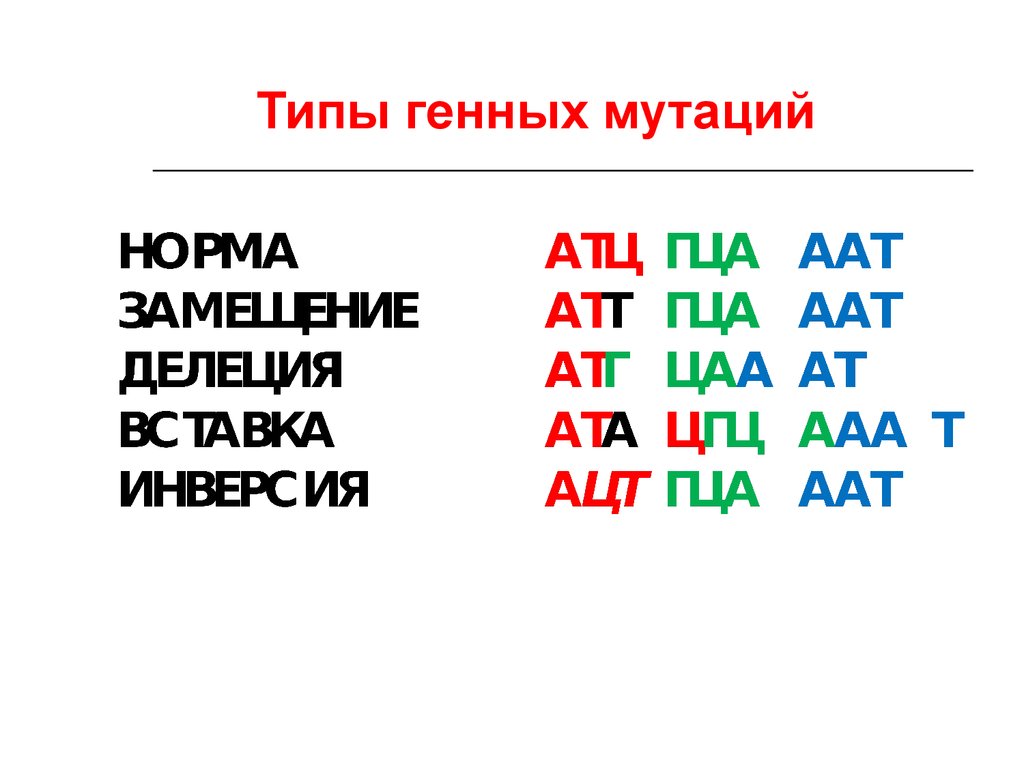
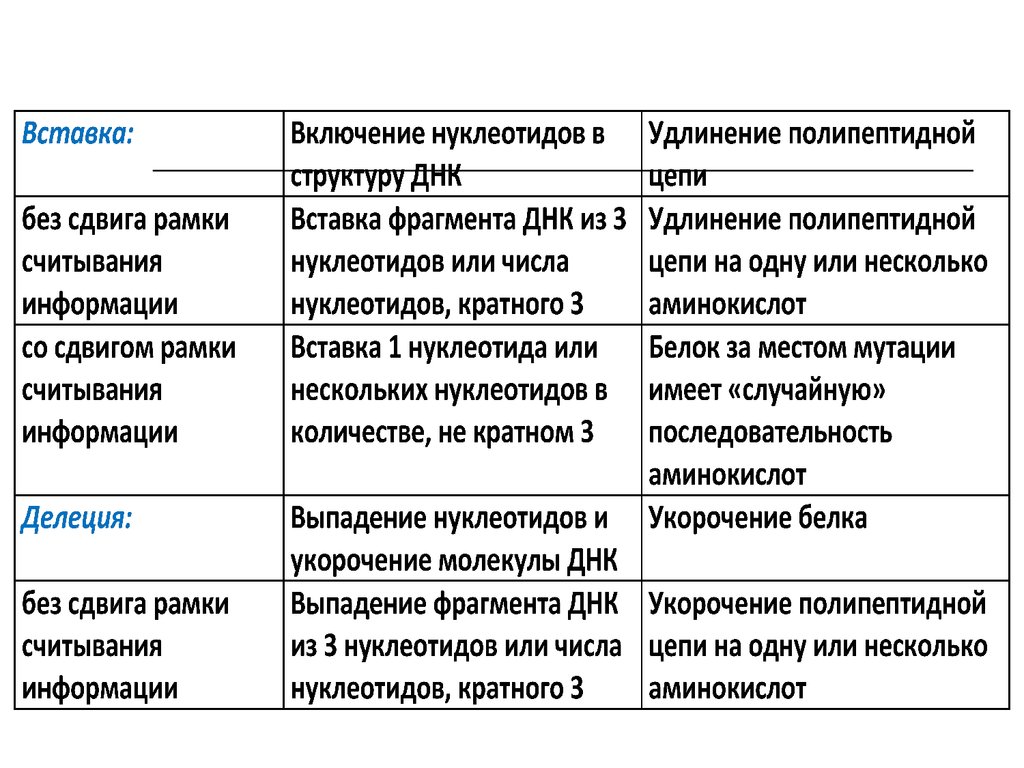
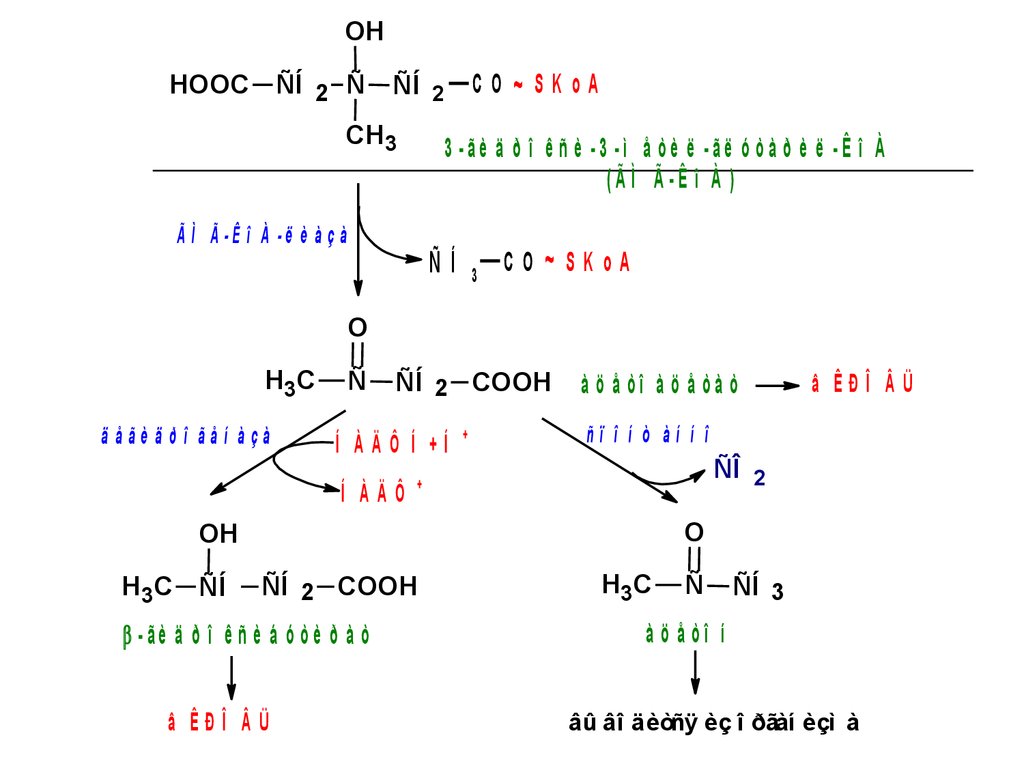
48. Классификация мутаций
49. Типы мутаций
50.
Типы генных мутаций51. Проявления мутаций
Вид мутацииИзменения в
структуре ДНК
Изменения в
структуре белка
Замена:
Замена в кодоне
Белок не изменён
нейтральная без
изменения смысла одного нуклеотида
другим
кодона
«миссенс-мутация»
с изменением
смысла кодона
Замена одной
аминокислоты на
другую
«нонсенс-мутация»
с образованием
терминирующего
кодона
На мутантном
триплете синтез
пептидной цепи
прекращается
52.
53. Механизмы увеличения числа и разнообразия генов в геноме
54. Механизмы увеличения числа и разнообразия генов в геноме (кроссинговер)
55. Механизмы увеличения числа и разнообразия генов в геноме (амплификация)
56. Полиморфизм белков –
изменение первичной структуры белка в пределаходного вида без изменения функций. В его основе
лежит генетическая гетерогенность молекул ДНК.
57. Схема возникновения наследственных болезней
58. Биохимические проявления наследственных болезней
59. Клинические проявления наследственных болезней
60. КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ КАФЕДРА ФУНДАМЕНТАЛЬНОЙ И КЛИНИЧЕСКОЙ БИОХИМИИ Лекция по БИОХИМИИ тема:
«Обмен липидов1»Краснодар
2016
61. Биологическая роль липидов
Резерв и источникэнергии
62.
Классификация липидов63. Классификация простых липидов
Простые липиды: сложные эфиры жирныхкислот с различными спиртами
Ацилглицеролы (нейтральные жиры) сложные эфиры трехатомного спирта
глицерина и высших жирных кислот.
Воска - сложные эфиры одноатомных или
двухатомных длиноцепочечных спиртов и
высших жирных кислот
Стериды - сложные эфиры циклического
спирта холестерола и высших жирных кислот
64. Строение простых липидов
СН2 O CО R1СН O CО R2
СН2 O СО R3
триацилглицерол
(нейтральный жир)
65. Классификация фосфолипидов
66. Строение глицеролфосфолипидов
СН 2 OCО R1
СН2 O
CО R1
СН O
CО R2
СН O
CО R2
OH
OH
СН 2 O P OH
СН2 O P O СН2 СН NH2
фосфатидная кислота
СН2 O
CО R1
СН O
CО R2
OH
СН2 O P O СН2 СН2 NH2
O
фосфатидилэтаноламин
COOH
O
O
фосфатидилсерин
СН2 O
CО R1
СН O
CО R2
OH
CH3
СН2 O P O СН2 СН2 N+ CH3
O
фосфатидилхолин
CH3
67. Строение сфингофосфолипидов
HN СО RH3C (CН 2)12 СН СН
СН
СН
O
OH
HO
сфингомиелин
СН2
P O
CH3
O СН2 СН2 N+ CH3
CH3
68. Классификация гликолипидов
гликолипидыглицеролгликолипиды
сфингогликолипиды
ганглиозиды
олигоцерамиды
цереброзиды
глюкоцереброзиды
галактоцереброзиды
цереброзидсульфатиды
69. Строение сфингогликолипидов
HN СО RHN СО R
H3C (CН2)12 СН СН
СН
СН2
СН
H3C (CН2)12 СН СН
СН 2ОН
O
СН 2ОН
O
HO
OH
глюкоцереброзид
СН2
СН
OH
O
OH
СН
O
OH
HO
OH
галактоцереброзид
OH
70. Жирные кислоты тканей человека
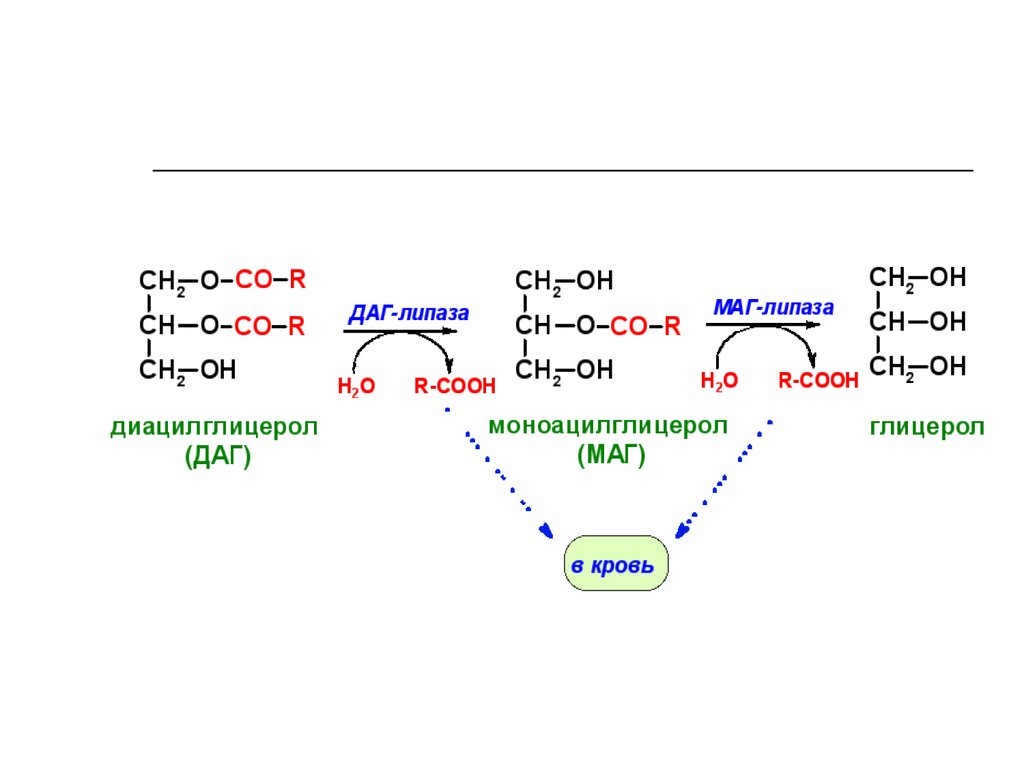
71. Переваривание нейтрального жира
СН2O СО R1СН O СО R2
СН2O СО R3
триглицерид
Н2О
R1 COOH
липаза
СН2OH
СН O СО R2
СН2O СО R3
диглицерид
Н2О
R3 COOH
СН2OH
СН O СО R2
липаза
СН2OH
моноглицерид
Н2О
липаза
R2 COOH
СН2OH
СН OH
СН2OH
глицерол
72. Строение желчных кислот
73. Переваривание фосфолипидов
фосфолипаза А1СН2 O CO R1
СН O CO R2
СН2 O PO3H
фосфолипаза А2
фосфолипаза D
N
фосфолипаза С
R1–COOH – насыщенная жк
R2–COOH – ненасыщенная жк
74. Переваривание стеридов
Í 2ÎR CO O
ýô èð õî ëåñòåðèí à
R-COOH
õî ëåñò åðî ëýñò åðàçà HO
õî ëåñòåðèí
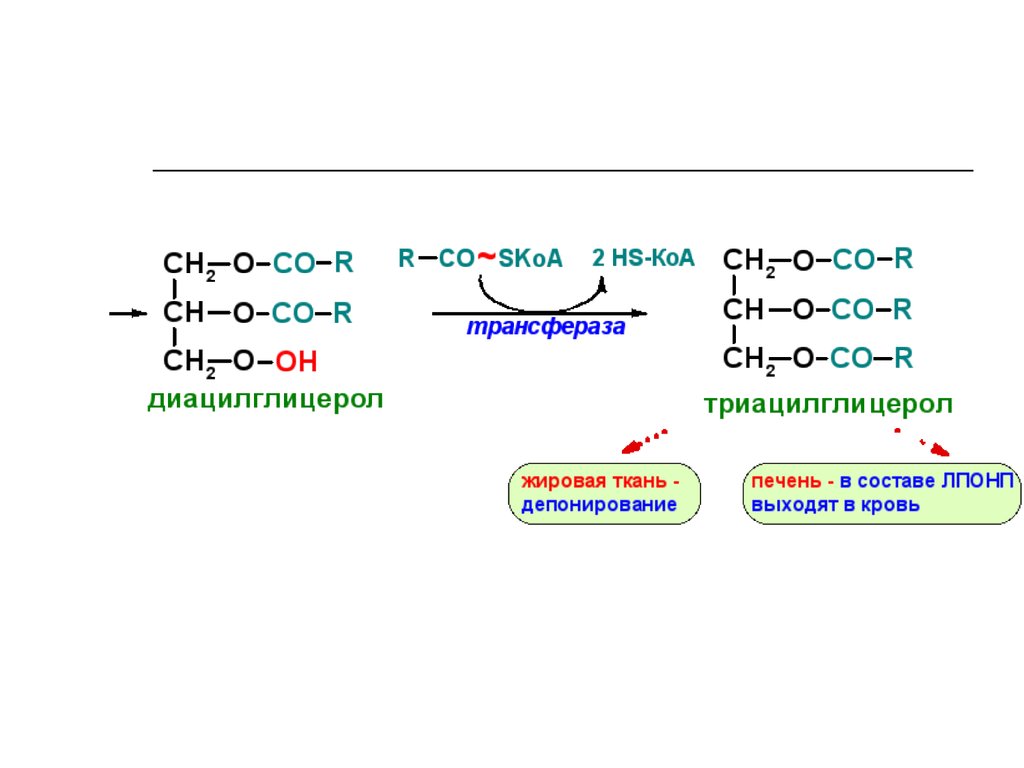
75. Ресинтез нейтрального жира (II)
ÑÍ2
ÑÍ
ÑÍ
OH
R CO ~SKoA
O CO
OH
2-ì î í î àöèëãëèöåðî ë
2
R
Í S-Êî À
ò ðàí ñô åðàçà
ÑÍ
2
O CO R
ÑÍ
ÑÍ
O CO R
2
OH
äèàöèëãëèöåðî ë
R CO ~SKoA
ò ðàí ñô åðàçà
Í S-Êî À
ÑÍ
2
ÑÍ
ÑÍ
O CO R
O CO R
O CO R
òðèàöèëãëèöåðî ë
2
76. Строение хиломикрона
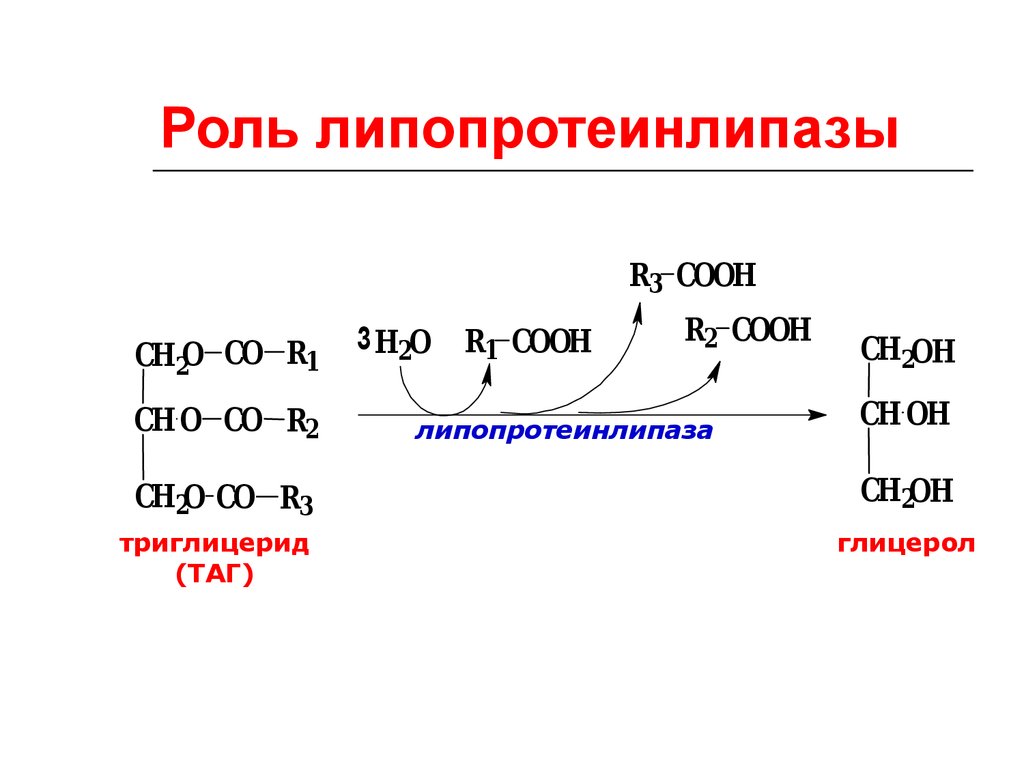
77.
Роль липопротеинлипазыR3 COOH
СН2O СО R1
СН O СО R2
СН2O СО R3
триглицерид
(ТАГ)
3 Н2О
R1 COOH
R2 COOH
липопротеинлипаза
СН2OH
СН OH
СН2OH
глицерол
78. СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ В ПЕЧЕНИ И ЖИРОВОЙ ТКАНИ
79.
80. МОБИЛИЗАЦИЯ ТРИАЦИЛ-ГЛИЦЕРОЛОВ
МОБИЛИЗАЦИЯТРИАЦИЛГЛИЦЕРОЛОВ
81.
82. Активация жирных кислот
Í S -Ê î ÀR
COOH
À ÒÔ
ÀÌ Ô + ÐÐi
à ö è ë -Ê î À ñè í ò åò àçà
2 R
CO ~ SKoA
à ö è ë -Ê î À
83. ТРАНСПОРТ ЖИРНЫХ КИСЛОТ В МИТОХОНДРИИ
+N (Ñ Í
ÑÍ
R
CO ~ SKoA
à ö è ë -Ê î À
+
3)3
O H
ÑÍ
ÑÍ
2
ì àò ð è êñ
ì èò î õî í ä ðè è
+
N (Ñ Í
ÑÍ
CO ~ SKoA
à ö è ë -Ê î À
àö è ë êàð í è ò è í ò ð àí ñô åð àçà
ö è ò î ï ë àçì àò è ÷ åñêàÿ
êà ð í è òè í
ö è ò î ï ë àçì à
+
ÑÍ
3)3
2
CO O H
êà ð í è òè í
2
ÑÍ
Î
ÑÍ
Î Ñ
R
2
CO O H
+
Í S -Ê î À
N (Ñ Í
ÑÍ
O H
3 )3
à ö è ë -ê à ð í è òè í
2
ÑÍ
N (Ñ Í
ÑÍ
2
CO O H
R
Í S -Ê î À
àö è ë êàð í è ò è í ò ð àí ñô åð àçà
ì èò î õî í ä ðèàëüí àÿ
2
ÑÍ
ÑÍ
3)3
Î
Î Ñ
R
2
CO O H
à ö è ë -ê à ð í è òè í
84. β-ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
RÑÍ 2 ÑÍ 2 CO ~ SKoA
àöèë-Êî À
ÔÀÄ
äåãèäðî ãåí àçà
ÔÀÄÍ
R
ÑÍ
ÑÍ
ãèäðàò àçà
R
2
CO ~ SKoA
åí î èë-Êî À
Í 2Î
ÑÍ
ÑÍ 2 CO ~ SKoA
OH
ãèäðî êñèàöèë-Êî À
2 АТФ
85.
R CH ÑÍ 2 CO ~ SKoAOH
ãèäðî êñèàöèë-Êî À
äåãèäðî ãåí àçà
Í ÀÄ+
Í ÀÄÍ + Í
+
3 АТФ
R Ñ ÑÍ 2 CO ~ SKoA
O
ò èî ëàçà
R CO ~ SKoA
àöèë-Êî À (nÑ-2)
êåòî àöèë-Êî À
Í S-KoA
CH3 CO ~ SKoA
àöåòèë-Êî À
следующий цикл β-окисления
ЦТК
12 АТФ
86. ОБЩАЯ СХЕМА ЦИКЛА β–ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ
дегидрогеназа2 АТФ
гидратаза
дегидрогеназа
3 АТФ
тиолаза
12 АТФ
87. ЭНЕРГЕТИЧЕСКИЙ ИТОГ β-ОКИСЛЕНИЯ
n – количество С-атомов в жирной кислоте;
n/2 – количество молекул ацетил-КоА,
образованных в процессе β-окисления;
12 – количество АТФ, синтезирующихся при
окислении ацетил-КоА в ЦТК;
(n/2 – 1) – количество циклов β-окисления;
5 – количество молекул АТФ, образованных в
каждом цикле за счёт двух реакций
дегидрирования;
1 – затрата 1 молекулы АТФ на активацию
жирной кислоты
88. Окисление ненасыщенных жирных кислот
Цис-транс-…
β-окисление
89. ИСТОЧНИКИ АЦЕТИЛ-КоА
Ï ÂÊÆÊ
( î êèñëåí èå)
êåò î ãåí í û å
ÀÊ
êåò î í î âû å
ò åëà
àöåòèë-Êî À
ÖÒÊ
õî ëåñò åðèí
æèðí û å
êèñëî ò û
êåò î í î âû å
ò åëà
90. Строение пальмитоилсинтетазы
Å2Å3
Å4
Å5
Å1
ÀÏ Á SH
Å6
SH
Å1 - òðàí ñô åðàçà
Å2 - òðàí ñô åðàçà
Å3 - ñèí òàçà
Å4 - ðåäóêòàçà
Å5 - ãèäðàòàçà
Å6 - ðåäóêòàçà
91. Биосинтез пальмитиновой кислоты
CH3 ÑÎ ~ SÊîÀ + ÑÎàöåòèë-Êî À
2
+ ÀÒÔ
àöåò èë-Êî Àêàðáî êñèëàçà
êî ô åðì åí ò áèî ò èí
HOOC ÑÍ 2 ÑÎ ~ SÊîÀ + ÀÄÔ + Ði
ì àëî í èë-Êî À
92. Схема биосинтеза пальмитиновой кислоты
93. Схема биосинтеза пальмитиновой кислоты
94. Суммарное уравнение биосинтеза пальмитиновой кислоты
95. Регуляция биосинтеза и окисления жирных кислот
активнаянеактивная
96. Удлинение жирных кислот
H3C (CН2)14 СО ~ SKoAпальмитоил-КоА
НАДФН+Н+
HOOC CН2 СО ~ SKoA
малонил-КоА
конденсация
элонгаза
(ферментный
комплекс)
восстановление
дегидратация
восстановление
H3C (CН2)16 СО ~ SKoA
стеарил-КоА
деацилаза
Н2О
НS-КоА
H3C (CН2)16 COOH
стеариновая кислота (стеарат)
97. Биосинтез непредельных жирных кислот
H3C (CН2)14 СО ~ SKoAпальмитоил-КоА
О2 + НАДФН+Н+
десатураза
2 Н2О + НАДФ+
H3C (CН2)5 СН СН (CН 2)7 СО~ SKoA
пальмитоолеил-КоА
деацилаза
Н2О
НS-КоА
H3C (CН2)5 СН СН (CН 2)7 COOH
пальмитоолеиновая кислота
(пальмитоолеат)
98. Синтез кетоновых тел
ÑÍ3
CO
~ SKoA +
ÑÍ
ò è î ë àçà
CH3 ÎÑ
3
~ SKoA
CO
2 à ö å òè ë -Ê î À
Í S -Ê î À
ÑÍ 2 C O
ÑÍ
Í 2Î
Ã Ì Ã -Ê î À -ñ è í ò à ç à
3
~ SKoA
CO
à ö å òî à ö å òè ë -Ê î À
~ SKoA
Í S -Ê î À
OH
HOOC ÑÍ 2 Ñ
ÑÍ 2
CH3
CO
~ SKoA
3 -ãè ä ð î ê ñ è -3 -ì å òè ë -ãë ó òà ð è ë -Ê î À
(ÃÌ Ã-Ê î À )
99.
OHHOOC ÑÍ 2 Ñ
CO
ÑÍ 2
CH3
~ SKoA
3 -ãè ä ð î ê ñ è -3 -ì å òè ë -ãë ó òà ð è ë -Ê î À
(ÃÌ Ã-Ê î À )
Ã Ì Ã -Ê î À -ë è à ç à
ÑÍ
3
CO
~ SKoA
O
H3C
ä åãè ä ðî ãåí àçà
Ñ
ÑÍ 2 COOH
Í ÀÄÔ Í +Í
Í ÀÄÔ
+
à ö å òî à ö å òà ò
ñï î í ò àí í î
ÑÎ 2
+
O
OH
H3C ÑÍ
â ÊÐÎ Â Ü
ÑÍ 2 COOH
-ãè ä ð î ê ñ è á ó òè ð à ò
â ÊÐÎ Â Ü
H3C
Ñ
ÑÍ 3
à ö å òî í
âû âî äèòñÿ èç î ðãàí èçì à
100. Окисление кетоновых тел
101. Типы липопротеинов
Типы липопротеиновФункции
Хиломикроны
(ХМ)
ЛПОНП
Транспорт Транспорт
экзогенных эндогенлипидов
ных
липидов
Место
Эпителий
Клетки
образования тонкого
печени
кишечника
ЛППП
ЛПНП
ЛПВП
Промежу- Транспорт Удаление
точная
холестери- избытка
форма
на в ткани холестерин
а
Кровь
Кровь (из
ЛПОНП и
ЛППП)
Клетки
печени
Плотность,
г/мл
0,92-0,98
0,96-1,00
1,00-1,06
1,06-1,21
Диаметр
частиц, нм
>120
30-100
21-100
7-15
В-100
А-I
С-II
E
Основные
В-48
апопротеины С-II
E
В-100
С-II
E
В-100
E