
")




")













 является:")















 необходимы:")


")
































")
")




 Медицина
Медицина Биология
Биология Химия
ХимияПохожие презентации:

. Основы молекулярной генетики")








Обмен нуклеотидов. Матричные биосинтезы
1. КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ КАФЕДРА ФУНДАМЕНТАЛЬНОЙ И КЛИНИЧЕСКОЙ БИОХИМИИ
Лекция по биохимииТема:
«Обмен нуклеотидов.
Матричные
биосинтезы»
Краснодар
2017
2. Нуклеиновые кислоты (ДНК и РНК)
– это биополимеры(полинуклеотиды), состоящие из
мононуклеотидов, соединённых
фосфодиэфирными связями.
ДНК хранит наследственную
информацию, т.е. информацию о
первичной структуре белков
данного организма, а РНК (мРНК,
тРНК и рРНК) её реализуют, т.е.
участвуют в синтезе белков
3. Функции РНК:
м-РНК является копией гена иматрицей (планом) для синтеза
белка
т-РНК осуществляет транспорт
аминокислот к месту синтеза
белка и встраивает аминокислоту
в полипептидную цепочку в
соответствии с кодоном
р-РНК вместе с белками
формирует рибосому – место
синтеза белка
4. Строение мононуклеотида
остатокфосфорной
кислоты
азотистое
основание
пентоза
нуклеозид
НУКЛЕОТИД
5. Пуриновые основания нуклеиновых кислот
NH2N
N
N1
2
6
3
5
4
N
N
N
7
9
NH
аденин (А)
8
NH
OH
пурин
N
N
H2N
N
гуанин (G)
NH
6. Пиримидиновые основания нуклеиновых кислот
OHN
HO
N3
2
4
1
OH
N
урацил (U)
5
CH3
N
6
HO
N
NH2
пиримидин
N
HO
N
цитозин (С)
N
тимин (Т)
7. Углеводы нуклеотидов (пентозы)
рибоза(РНК)
дезоксирибоза
(ДНК)
8. Гидролиз нуклеопротеинов
ротоваяполость
ДНП и РНП пищи
белки
НСl, пепсин
желудок
(протамины, гистоны)
ДНК, РНК
(полинуклеотиды)
РНК-азы, ДНК-азы
ДПК
(эндонуклеазы)
аминокислоты
Н2О
олигонуклеотиды
фосфодиэстеразы
(экзонуклеазы)
Н2О
мононуклеотиды
тонкий
кишечник
ткани
нуклеотидазы
(фосфатазы)
Н2О
Н3РО4
нуклеозиды
нуклеозидазы
пурины,
пиримидины
(азотистые основания)
рибоза,
дезоксирибоза
(пентозы)
пентозофосфатный
путь
9. Катаболизм пуриновых оснований
В нормесодержание
мочевой
кислоты в крови
0,14-0,5 ммоль/л
10. Нарушения обмена пуринов
11. Ингибитор ксантиноксидазы
OHOH
N
N
N
N
H
гипоксантин
N
N
N
H
N
аллопуринол
12. Распад пиримидиновых оснований
13. Особенности синтеза нуклеотидов
1.2.
3.
4.
Синтез идёт из обычных простых
предшественников (ак, углекислого
газа и т.п.)
Синтезируются не отдельные
азотистые основания, а сразу
нуклеотиды
Синтезируются общие
предшественники (для пуриновых
нуклеотидов инозинмонофосфат –
ИМФ, для пиримидиновых –
уридинмонофосфат – УМФ)
Синтез протекает ферментативно, с
большой затратой энергии
14.
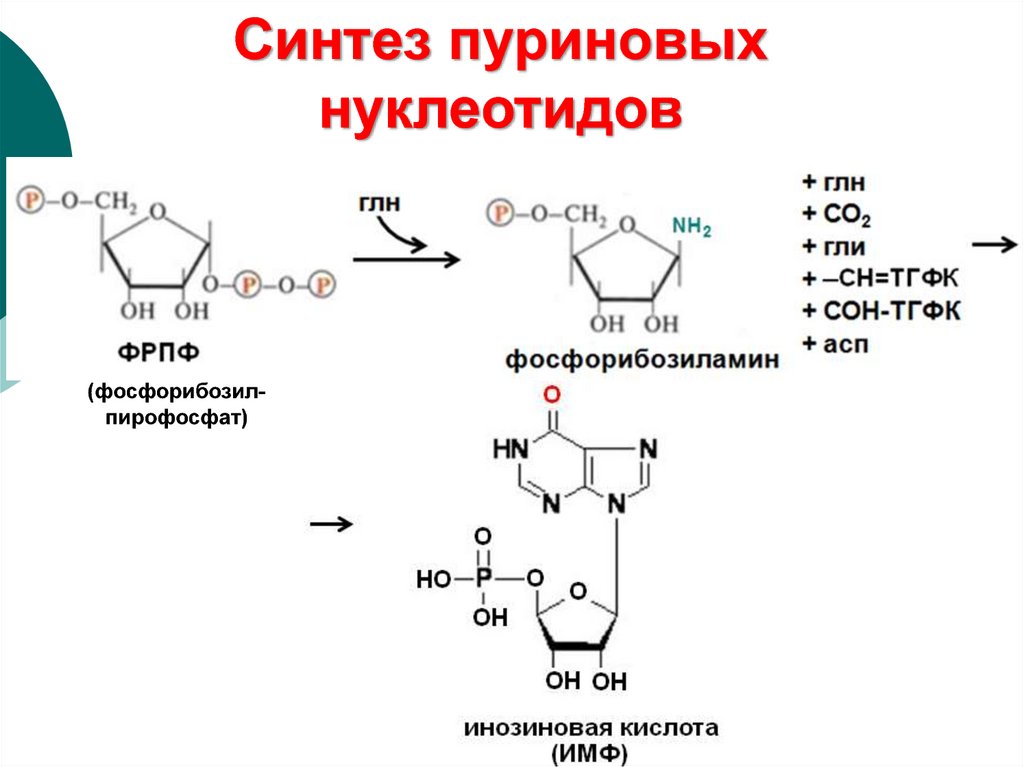
Синтез пуриновыхнуклеотидов
(фосфорибозилпирофосфат)
15. Происхождение атомов пуринового кольца
16. Синтез пуриновых нуклеотидов
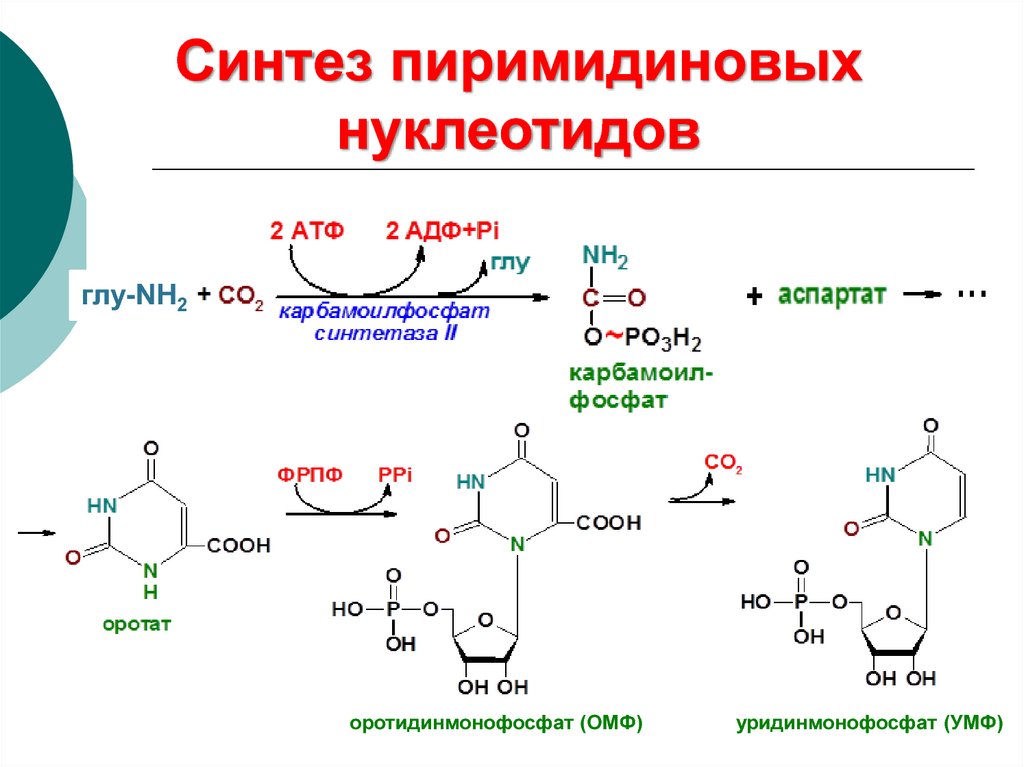
17.
Синтез пиримидиновыхнуклеотидов
глу-NH2
оротидинмонофосфат (ОМФ)
уридинмонофосфат (УМФ)
18. Синтез пиримидиновых нуклеотидов
19. Синтез дезоксирибонуклеотидов
20. Ингибиторы синтеза нуклеиновых кислот
21. Нарушения обмена нуклеотидов
Нарушение синтеза пиримидинов –оротацидурия – дефект ОМФдекарбоксилазы (недостаток синтеза
пиримидиновых нуклеотидов, снижение
синтеза нуклеиновых кислот)
Нарушения обмена пуринов:
– ксантинурия – дефект ксантиноксидазы
(увеличение содержания ксантина в моче,
возможно образование ксантиновых
камней)
– подагра
– синдром Лёша-Нихана
22. Первичная структура нуклеиновых кислот
основаниеоснование
фосфодиэфирная
связь
основание
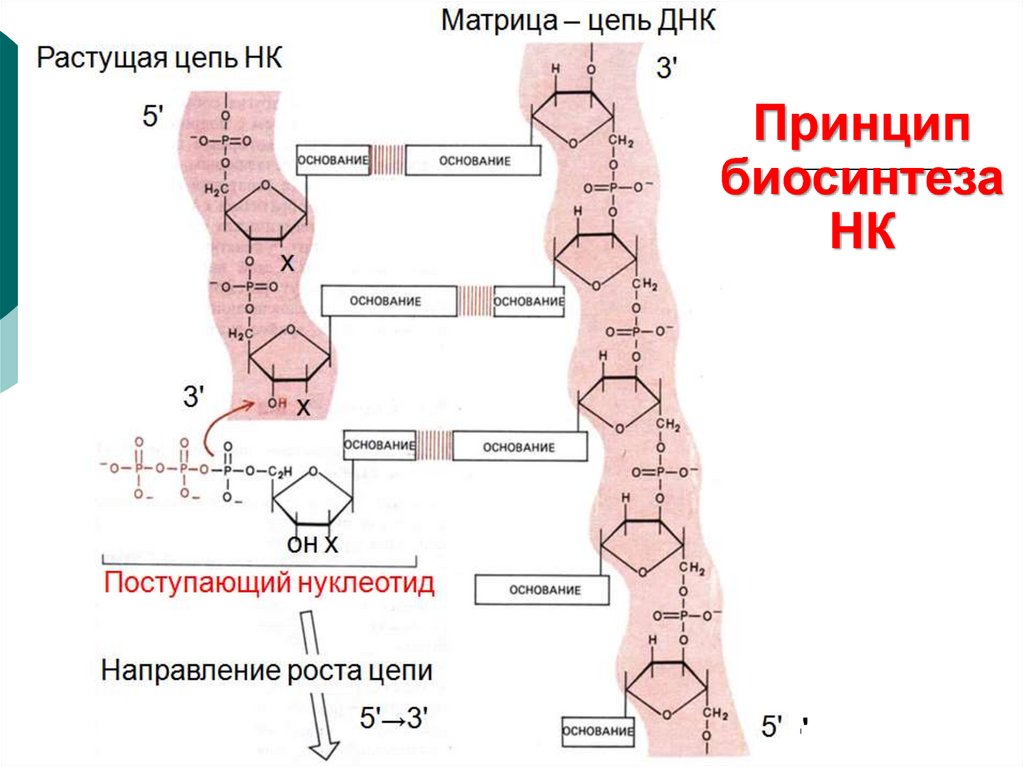
23.
Матрица – цепь ДНКРастущая цепь НК
3'
Принцип
биосинтеза
НК
5'
х
3'
х
х
Поступающийоннуклеотид
Направление роста цепи
5'
→3'
5'
24. Биосинтез ДНК (репликация) является:
матричным (матрица – обе нити ДНК)комплиментарным
фрагментарным (нити ДНК
синтезируются в виде фрагментов,
которые затем соединяются между
собой)
полуконсервативным (в каждой из
образовавшихся молекул ДНК одна
нить исходная – материнская, а одна
– вновь синтезированная – дочерняя)
25. Полуконсервативность биосинтеза ДНК
26.
РЕПЛИКАТИВНАЯ СИСТЕМА1. МАТРИЦА – ОБЕ НИТИ ДНК НА ВСЕМ ПРОТЯЖЕНИИ
2. СТРОИТЕЛЬНЫЙ МАТЕРИАЛ:
ДЛЯ СИНТЕЗА ПРАЙМЕРА – АТФ, ГТФ, ЦТФ, УТФ
ДЛЯ СИНТЕЗА ДНК – дАТФ, дГТФ, дЦТФ и дТТФ
3. ФЕРМЕНТАТИВНЫЕ БЕЛКИ
ТОПОИЗОМЕРАЗА (ГИРАЗА)
ХЕЛИКАЗА
ДНК - ПОЛИМЕРАЗЫ
α - праймаза
β - фермент репарации
γ - митохондриальный фермент
δ - строит ведущую цепь
ε - строит отстающую цепь
ДНК –ЛИГАЗА - сшивает фрагменты Оказаки
4. ДНК-СВЯЗЫВАЮЩИЕ БЕЛКИ (SSB-белки)
5. РЕГУЛЯТОРНЫЕ БЕЛКИ: факторы инициации,
элонгации, терминации
27. Особенности репликации
ДНК-полимеразы δ и ε не могутсоединять между собой два
мононуклеотида, а только достраивают
имеющуюся нить
Синтез идёт только в направлении от 5´к 3´-концу (т.е. в разных направлениях
на разных нитях материнской ДНК)
Репликативная вилка движется только в
одном направлении
Синтез ДНК начинается одновременно в
нескольких точках (ориджинах
репликации), участок ДНК между двумя
ориджинами называется «репликон»
28. Этапы репликации
1. Инициация:
Топоизомераза находит точку начала
репликации, гидролизует одну
фосфодиэфирную связь и даёт
возможность компоненатам
репликативной системы разомкнуть нити
ДНК и образовать репликативную
«вилку», а затем вновь соединяет связь
между мононуклеотиджами
Хеликаза разрывает водородные связи
между нитями ДНК
29.
Этапы репликацииДНК-связывающие белки (SSB-белки)
стабилизируют репликативную вилку, не
давая восстанавливаться водородным связям
между комплиментарными нуклеотидами
ДНК-полимераза α (праймаза) строит
праймер («затравку») из 8-10
рибонуклеотидов и 40-50
дезоксирибонуклеотидов, а ДНК-полимераза
δ достраивает нить из
дезоксирибонуклеотидов на лидирующей
нити, а ДНК-полимераза ε – на отстающей
нити ДНК
30. Инициация репликации
ДНК-полимераза αДНК-полимераза δ
ДНК-полимераза ε
31. Инициация репликации
Ориджинрепликации
репликон
32. Этапы репликации
2. ЭлонгацияДНК-полимераза
δ продолжает
удлинять нить из
дезоксирибонуклеотидов на
лидирующей нити, а ДНКполимераза ε – фрагменты
(фрагменты Оказаки) на
отстающей нити ДНК по мере
движения репликативной вилки
33. Элонгация репликации
ДНК-полимераза δДНК-полимераза
α (праймаза)
ДНК-полимераза ε
34. Элонгация репликации
35.
Этапы репликацииТерминация
ДНК-полимераза
β (фермент
репарации) удаляет праймеры и
достраивает фрагменты ДНК
ДНК-лигаза соединяет фрагменты
между собой
36.
Функции ДНК-полимеразы β37. Репарация ДНК
38. Вторичная структура ДНК
39. Строение нуклеосомы
40. Для биосинтеза РНК (транскрипции) необходимы:
•МАТРИЦА – участок одной из нитей ДНК– (транскриптон)
•СТРОИТЕЛЬНЫЙ МАТЕРИАЛ:
– АТФ, ГТФ, ЦТФ, УТФ
•ФЕРМЕНТАТИВНЫЕ БЕЛКИ
ДНК-зависимые РНК-полимеразы
I — для синтеза р-РНК
II — для синтеза м-РНК
III — для синтеза т-РНК
•РЕГУЛЯТОРНЫЕ БЕЛКИ: факторы инициации,
элонгации, терминации
41. Биосинтез РНК
42. Биосинтез РНК
РНК-полимеразаТранскрибируемая нить
ДНК
Нетранскрибируемая нить
мононуклеотиды
пре-РНК (первичный транскрипт)
Сайт терминации
43. Процессинг РНК (1. сплайсинг)
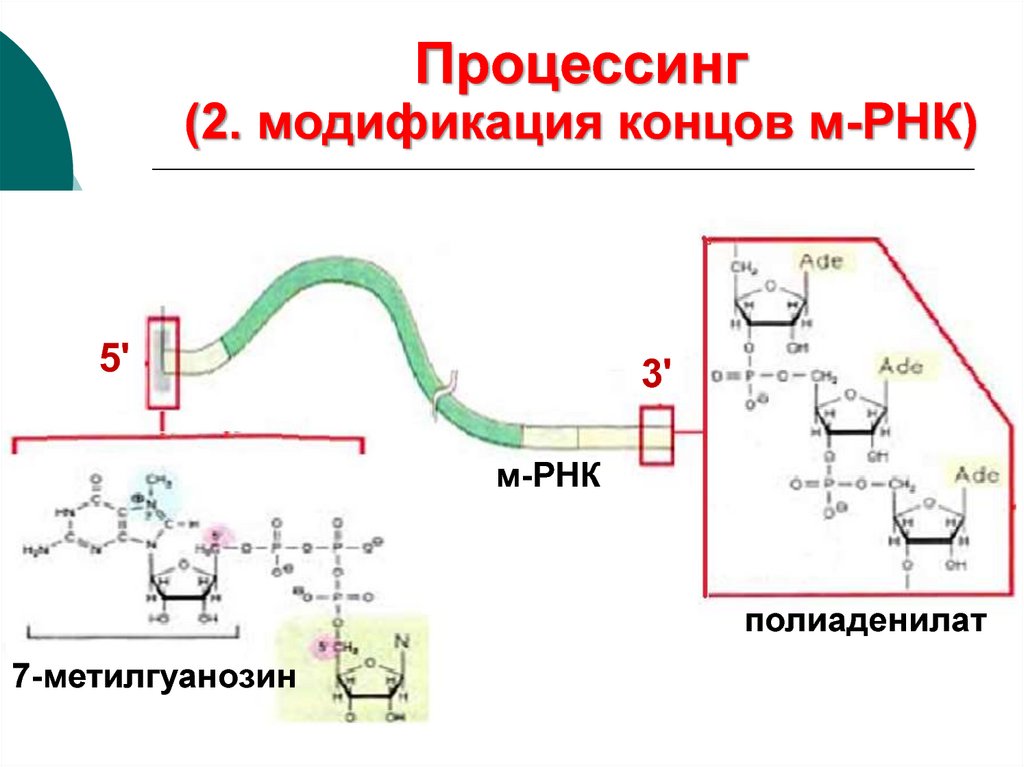
44.
Процессинг(2. модификация концов м-РНК)
5'
3'
м-РНК
полиаденилат
7-метилгуанозин
45. Состав зрелой м-РНК
5'-конец"колпачок" (кэп)
5'-нетранслируемый
участок
инициирующий кодон
кодирующая часть
терминирующий кодон
3'-нетранслируемый
участок
3'-конец
поли(А)-фрагмент
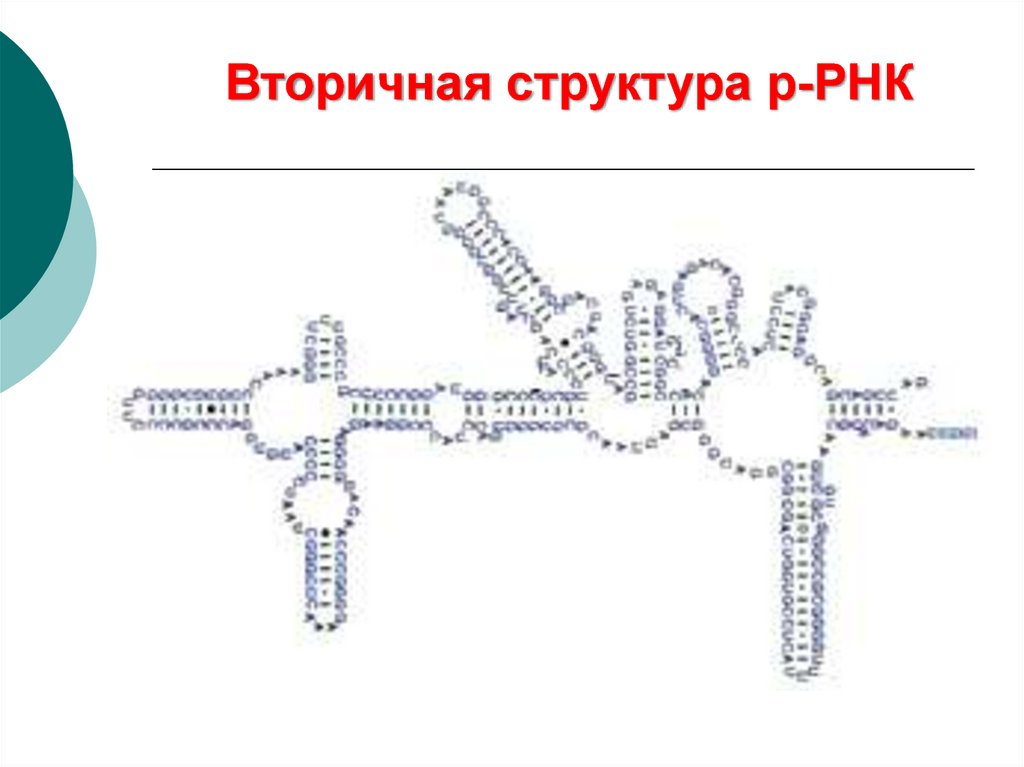
46.
Вторичная структура р-РНК47. Вторичная структура т-РНК
антикодон48. Общая схема биосинтеза белка
49. Компоненты белоксинтезирующей системы
мРНК20 Аминокислот
20 Аминоацил-тРНК синтетаз (АРС-аз)
Изоакцепторные тРНК
Рибосомы в виде полисом
Источники энергии (АТФ, ГТФ) и Мg2+
Белковые факторы регуляции: факторы
инициации, элонгации, терминации
Специальные ферменты
посттрансляционного процессинга
полипептидной цепи
50. Ядерные стадии биосинтеза белка
БиосинтезмРНК (транскрипция)
Созревание мРНК
(посттранскрипционный
процессинг)
51. Функции т-РНК
Акцепторная: связывает аминокислоту итранспортирует её к месту синтеза белка
Адаптерная: встраивает аминокислоту на
соответствующее место в образующемся
полипептиде (за счёт взаимодействия
антикодона в т-РНК с кодоном в м-РНК)
52. Строение т-РНК
антикодон53. Таблица генетического кода
УУУУУЦ
УУА
УУГ
ЦЦУ
ЦУЦ
ЦУА
ЦУГ
АУУ
АУЦ
АУА
АУГ
ГУУ
ГУЦ
ГУА
ГУГ
Фен
Лей
Иле
Мет+иниц
Вал
+ иниц
УЦУ
УЦЦ
УЦА
УЦГ
ЦЦУ
ЦЦЦ
ЦЦА
ЦЦГ
АЦУ
АЦЦ
АЦА
АЦГ
ГЦУ
ГЦЦ
ГЦА
ГЦГ
Сер
Про
Тре
Ала
УАУ
УАЦ
УАА
УАГ
ЦАУ
ЦАЦ
ЦАА
ЦАГ
ААУ
ААЦ
ААА
ААГ
ГАУ
ГАЦ
ГАА
ГАГ
Тир
Терм
Гис
Глн
Асн
Лиз
Асп
Глу
УГУ
УГЦ
УГА
УГГ
ЦГУ
ЦГЦ
ЦГА
ЦГГ
АГУ
АГЦ
АГА
АГГ
ГГУ
ГГЦ
ГГА
ГГГ
Цис
Терм
Три
Арг
Сер
Арг
Гли
54. Характеристика генетического кода
Триплетность (1 аминокислота кодируется 3нуклеотидами)
Специфичность (каждому кодону соответствует
только 1 аминокислота)
Вырожденность (1 аминокислота может кодироваться
более чем 1 триплетом)
Линейная запись (прочтение кода «без знаков
препинания»)
Универсальность (одинаков для всех живых существ)
Наряду со значимыми есть и «бессмысленные»
кодоны (терминирующие – УАА, УАГ, УГА)
Колинеарность (соответствие линейной
последовательности кодонов гена и
последовательности АК в кодируемом белке)
55. Колинеарность генетического кода
Глу-тРНК56. Цитоплазматические стадии биосинтеза белка
Активацияаминокислот, или
образование аминоацил-тРНК
Инициация
Элонгация
Терминация
Посттрансляционная
модификация
57. Синтез аминоацил-тРНК
АТФR СН COOH
РPi
R СН CO~О АМФ
NH2
NH2
аминокислота
аминоацилтРНК-синтетаза
аминоациладенилат
т-РНК
АМФ
R СН CO~ т-РНК
NH2
аминоацил-тРНК
58. Рибосома эукариотов
59. Функции активной рибосомы
Связывание и удерживаниебелоксинтезирующей системы
Транслокация – перемещение м-РНК
через рибосому каждый раз на один
триплет
Замыкание пептидной связи между
аминокислотами (каталитическая
функция)
60. Функционирующая рибосома
61. Образование инициирующего комплекса
40S-субъединицамРНК
(eIF-3)
(40S) (мРНК)
(eIF-2, ГТФ,
eIF-1)
Мет-тРНКМет
(40S) (мРНК) (Мет-тРНКМет)
60S-субъединица
(40S) (мРНК) (Мет-тРНКМет) (60S)
62. Инициация:
К малой субъединице рибосомыприкрепляется м-РНК
К инициирующему кодону прикрепляется
Мет-т-РНК
К образовавшемуся комплексу
присоединяется большая субъединица таким
образом, что Мет-т-РНК оказывается в
пептидильном центре, а в аминоацильном –
кодон, соответствующий второй
аминокислоте
63. Инициация трансляции
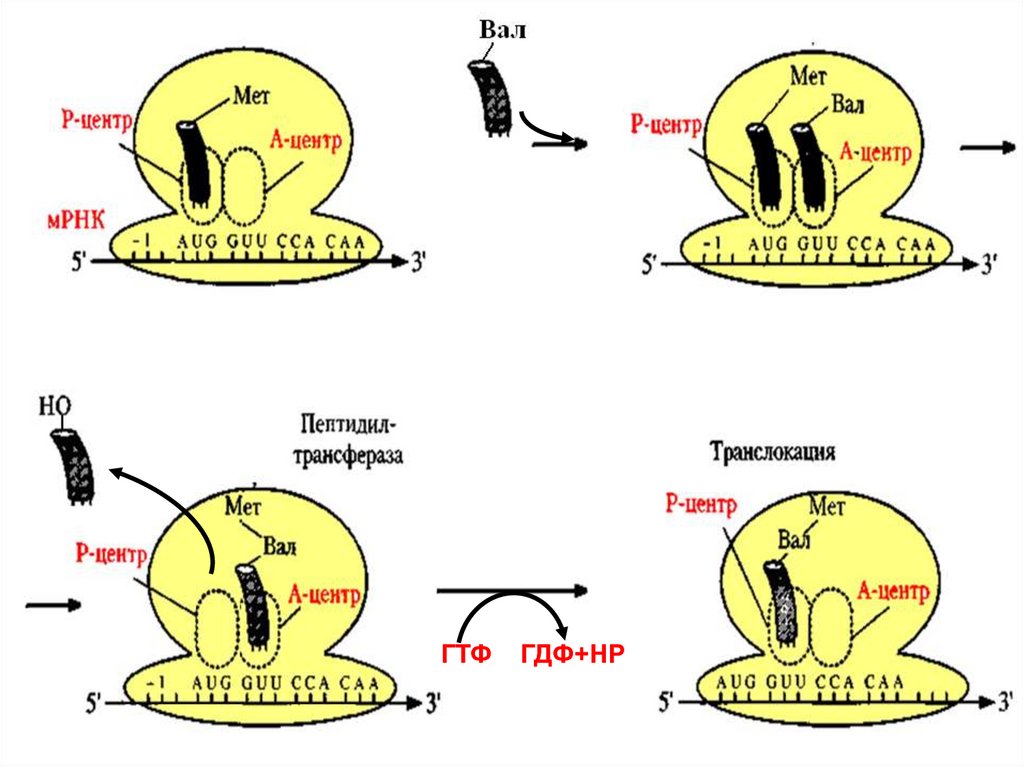
64. Элонгация:
В пептидильном центре находится Мет-т-РНК, а ваминоацильном – аа-тРНК, соответствующая второму
кодону.
Мет отрывается от т-РНК и перебрасывается в
аминоацильный центр
С помощью пептидилтрансферазы замыкается пептидная
связь между карбоксильной группой Мет и аминогруппой
второй аминокислоты (требуется энергия ГТФ и
регуляторный фактор Т)
Происходит передвижение рибосомы по м-РНК на один
триплет (транслокация)
В освободившийся аминоацильный центр входит аа-тРНК, соответствующая третьему кодону и т.д.
65.
ГТФГДФ+НР
66. Строение полирибосомы
67. Посттрансляционный процессинг
Модификация N-конца полипептидной цепиФолдинг (формирование пространственной
структуры)
Химическая модификация
(гидроксилирование, гликозилирование и др.)
Присоединение простетических групп (у
гетеропротеинов)
Объединение протомеров при образовании
олигомерных белков
Присоединение сигнальных пептидов для
выхода белка из клетки
68. Регуляция биосинтеза
1: РНК полимераза, 2: Репрессор, 3: Промотор,4: Оператор, 5: Лактоза, 6, 7, 8: гены, кодирующие
лактазу.
69. Действие регуляторных белков
70. Регуляция биосинтеза
Энхансеры – участки ДНК,присоединение к которым
регуляторных белков усиливает
транскрипцию
Сайленсеры – участки ДНК,
присоединение к которым
регуляторных белков уменьшает
транскрипцию
71. Амплификация
72. Типы генов в геноме
Структурные гены (кодируютбелки)
Регуляторные гены:
Гены-регуляторы (регулируют
работу структурных генов)
Процессинг-гены (регулируют
посттранскрипционные и
посттрансляционный процессинг)
Темпоральные гены (включают в
работу структурные гены в ходе
клеточной дифференцировки)
73. Клеточная дифференцировка
Hb P (Е)Hb F
эмбриональный
Нb плода
α2 ε2
α2 γ 2
Hb A
Hb A2
(гемоглобин взрослых)
α2 β2
α2 δ 2
74. Классификация мутаций
мутацииспонтанные
генные
моногенные
индуцируемые
хромосомные
геномные
по
локализации
полигенные
таутомерные
превращения
замещения
нейтральные
по
условиям
включение
аналогов
вставки
делеции
по
механизму
по типу
повреждения
по
проявлению
положительные отрицательные
"молчащие"
патологические
75. Типы мутаций
Тип мутацийГеномные
Характер мутационных
изменений
Изменение числа хромосом
Хромосомные Общее число хромосом не
меняется. Наблюдается
перестройки хромосом,
обычно видимые при
микроскопическом
исследовании
Генные
Изменения затрагивают один
кодон или небольшой отрезок
гена и не обнаруживаются
цитогенетически
76.
Типы генных мутацийНОРМА
ЗАМЕЩЕНИЕ
ДЕЛЕЦИЯ
ВСТАВКА
ИНВЕРСИЯ
АТЦ
АТТ
АТГ
АТА
АЦТ
ГЦА
ГЦА
ЦАА
ЦГЦ
ГЦА
ААТ
ААТ
АТ
ААА Т
ААТ
77. Проявления мутаций
Вид мутацииИзменения в
структуре ДНК
Изменения в
структуре белка
Замена:
нейтральная без
Замена в кодоне
Белок не изменён
изменения смысла одного нуклеотида
кодона
другим
«миссенс-мутация»
Замена одной
с изменением
аминокислоты на
смысла кодона
другую
«нонсенс-мутация»
На мутантном
с образованием
триплете синтез
терминирующего
пептидной цепи
кодона
прекращается
78.
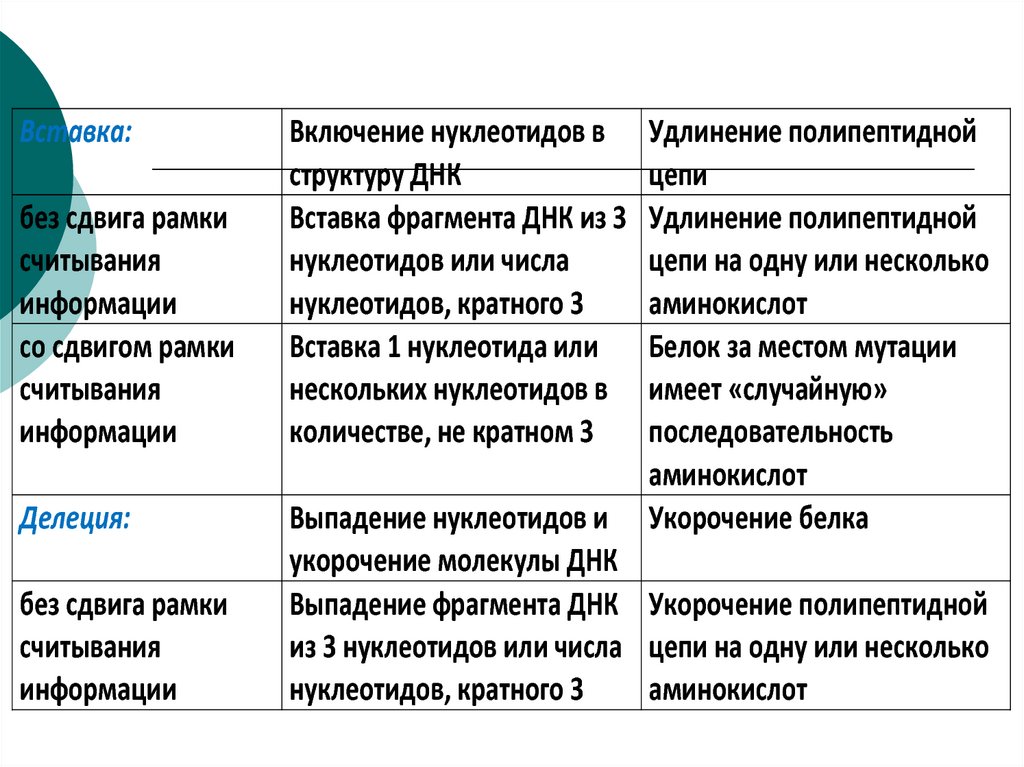
Вставка:без сдвига рамки
считывания
информации
со сдвигом рамки
считывания
информации
Делеция:
без сдвига рамки
считывания
информации
Включение нуклеотидов в
структуру ДНК
Вставка фрагмента ДНК из 3
нуклеотидов или числа
нуклеотидов, кратного 3
Вставка 1 нуклеотида или
нескольких нуклеотидов в
количестве, не кратном 3
Удлинение полипептидной
цепи
Удлинение полипептидной
цепи на одну или несколько
аминокислот
Белок за местом мутации
имеет «случайную»
последовательность
аминокислот
Укорочение белка
Выпадение нуклеотидов и
укорочение молекулы ДНК
Выпадение фрагмента ДНК Укорочение полипептидной
из 3 нуклеотидов или числа цепи на одну или несколько
нуклеотидов, кратного 3
аминокислот
79. Механизмы увеличения числа и разнообразия генов в геноме
1. Полная редубликация гена с последующими независимымимутациями.
Синтез цепей гемоглобина
ген
ген
ген
ген
ген
ген
ген
2. Полная редубликация с последующим слиянием одной
пары
3. Кроссинговер
4. Амплификация
80. Механизмы увеличения числа и разнообразия генов в геноме (кроссинговер)
81. Механизмы увеличения числа и разнообразия генов в геноме (амплификация)
82. Полиморфизм белков –
изменение первичной структуры белка в пределаходного вида без изменения функций. В его основе
лежит генетическая гетерогенность молекул ДНК.
полиморфизм лактатдегидрогеназы
Н Н
Н Н
Н Н
Н М
М М
Н Н
Н М
М М
М М
М М
ЛДГ1
ЛДГ2
ЛДГ3
ЛДГ4
ЛДГ5
полиморфизм гаптоглобинов
Hp =
S
F
83. Схема возникновения наследственных болезней
ДНКСтруктурные гены
м-РНК
т-РНК
р-РНК
Регуляторные гены
(не кодируют структуру, а
контролируют
экспрессию генов)
дефект на уровне
транскрипции трансляции
синтезируется дефектный
белок
изменяется количество
белка
протеинопатия
ферментопатия
неферментная
протеинопатия
метаболические
неметаболические
нарушения
нарушения
наследственная болезнь
84. Биохимические проявления наследственных болезней
ген АА
ЕА
ген В
В
ЕВ*
С
ген C
ген D
ЕС
ЕD
D
K
Возможные проявления нарушений:
1. накопление В
2. отсутствие С
3. появление К
F