



























































































 Медицина
МедицинаПохожие презентации:










Функциональная биохимия нервной ткани
1.
ЗАПОРОЖСКИЙ ГОСУДАРСТВЕННЫЙМЕДИЦИНСКИЙ УНИВЕРСИТЕТ
Кафедра клинической лабораторной диагностики
ФУНКЦИОНАЛЬНАЯ
БИОХИМИЯ НЕРВНОЙ
ТКАНИ
кандидат медицинских наук, доцент
Беленький Сергей Андреевич
ЗАПОРОЖЬЕ
2016
2.
3.
Морфо-функциональныеособенности нервной
ткани.
4.
Ткань – это филогенетическисложившаяся система
обладающих общностью
строения (а в ряде случаев и
общностью происхождения)
клеток и неклеточных
структур, специализированных
на выполнении определенных
функций.
5.
В любой системе все её элементыупорядочены в пространстве и
функционируют согласованно друг
с другом.
Система в целом приобретает
при этом свойства, не присущие
ни одному из ее элементов,
взятому в отдельности !
Соответственно и в каждой
ткани ее строение и функции
несводимы к простой сумме
свойств отдельных входящих
в нее клеток.
6.
Морфофункциональнаяклассификация тканей
1. Эпителиальные
2. Ткани внутренней
среды
3. Мышечные
4. Нервная
7.
НЕРВНАЯ ТКАНЬ – этоткань
эктодермального происхождения,
представляющая собой сложнейшую систему взаимосвязанных
специализированных структур
(нейронов, элементов глии, рецепторов,
проводников, синапсов и др.),
образующих основу нервной
системы и создающих условия
для реализации её
специфических функций:
восприятие раздражений, преобразование
их в нервный импульс (возбуждение) и
передача его к эффектору .
8.
НЕРВНАЯСИСТЕМА –
чрезвычайно сложная в структурном и функциональном отношении
система организма, интегрирующая и
регулирующая все происходящие в
нем процессы. Она обеспечивает:
связь
организма с внешней средой;
функционирование
его
как
единого
целого
(взаимосвязь
и
согласованную
работу тканей, органов и систем)
переработку и хранение информации;
процессы (обучение, внимание, мышление, чувства, речь), позволяющие
человеку
познавать
окружающую среду и активно её изменять.
когнитивные
9.
Нейрогуморальная регуляция:1. Гуморальная – изменение
физиологической активности
организма под влиянием
химических веществ.
Источники передачи информации:
– утилизоны – продукты метаболизма (СО2, глюкоза, жирные кислоты)
– информоны – гормоны желез
внутренней секреции, местные
или тканевые гормоны.
10.
2. Нервная – изменение физио-логической активности организма при помощи электрохимических потенциалов, распространяющихся по нервным
волокнам. Её особенности:
–
более
поздний
продукт
эволюции;
–
обеспечивает
быструю
регуляцию;
– имеет
точного
адресата
воздействия;
– экономичный способ регуляции;
–
высокая
надежность
передачи
информации.
11.
Нервной ткани из-за специфики еёфункций, кроме характерных для
любой ткани свойств, присущи:
особенности химического состава и характера метаболизма
наличие сложных компенсаторноприспособительных механизмов на
различных уровнях:
– молекулярном (специфические
рецептор- и канал-образующие белки,
ферментативные системы)
– клеточном (взаимодействие
«нейрон – глия»)
– тканевом (ГЭБ, ликвор и др.)
12.
Энергообеспечение нервнойткани
Основной особенностью обмена
веществ в нейронах является
преобладание аэробных
процессов и высокая их
интенсивность.
Вес мозга взрослого человека к
весу тела – 2%, а потребление
им кислорода у взрослых 2025%, а у детей – 50% от
общего его потребления в покое!
13.
Энергообеспечение нервнойткани
Основной субстрат дыхания мозга –
глюкоза, постоянно поступающая
из крови. Мембраны нейронов не
имеют рецепторов к инсулину и
свободно проницаемы для глюкозы (перемещение по градиенту
концентраций). Концентрация
глюкозы в нейронах четко
коррелирует с концентрацией в
плазме. Утилизируется она в них
также без участия инсулина.
14.
Энергообеспечение нервнойткани
Мозгу человека нужно 100 г
глюкозы в сутки (90% её
окисляется до СО2 и Н2О в ЦТК).
100 г ткани мозга потребляют
5 мг глюкозы в минуту.
Концентрация глюкозы в клетках
мозга – около 50 мг на 100 г ткани,
т.е. её количества в мозгу
достаточно
на 10 минут жизни (!)
15.
Высокая скорость потребленияглюкозы клетками мозга
обеспечивается работой двух
изоформ высокоактивной
гексокиназы.
Её активность в клетках мозга –
350-450 мкмоль / г / час.
(мышцы – 100-120 мкмоль / г / час,
печень – 25-30 мкмоль / г / час).
Фермент в нейронах расположен вблизи митохондрий или
прямо на их внешней мембране(!)
16.
Из поступившей в клетки мозгаглюкозы за счет гексокиназной
реакции образуется
90-95% глюкозо-6-фосфата
(в других тканях источники этого
промежуточного продукта – это
гликогенолиз и
глюконеогенез !!!)
Наблюдаются существенные
отличия путей дальнейшего
метаболизма этого интермедиата.
17.
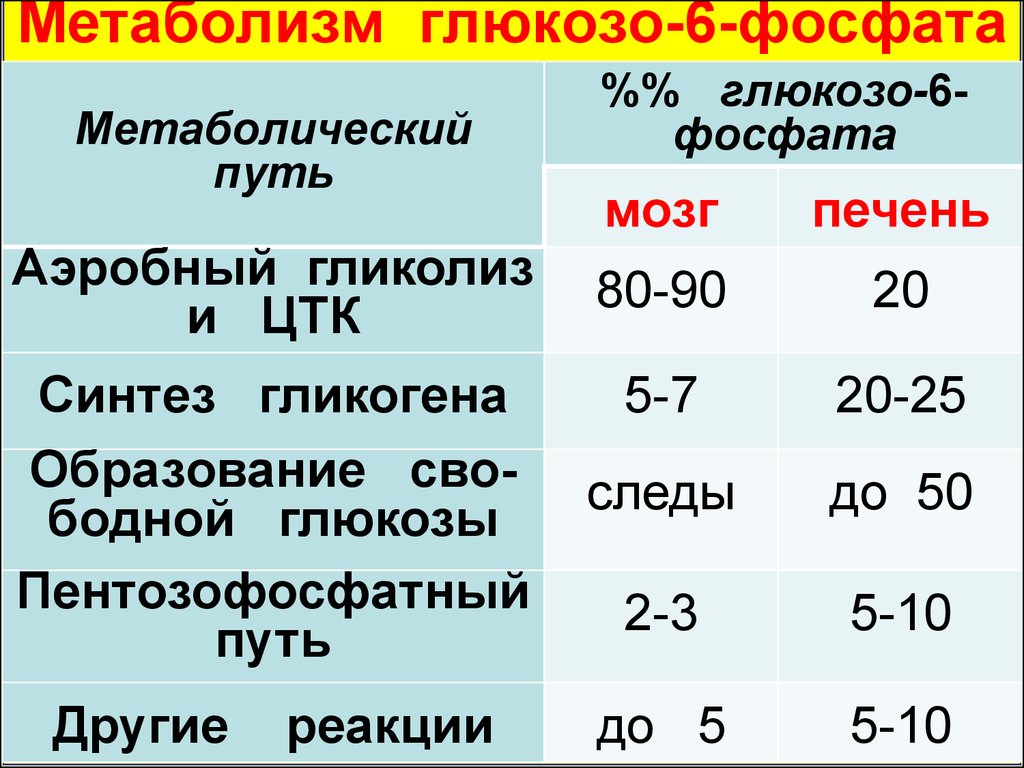
Метаболизм глюкозо-6-фосфатаМетаболический
путь
Аэробный гликолиз
и ЦТК
Синтез гликогена
Образование свободной глюкозы
Пентозофосфатный
путь
Другие
реакции
%% глюкозо-6фосфата
мозг
печень
80-90
20
5-7
20-25
следы
до 50
2-3
5-10
до 5
5-10
18.
Уменьшение соотношения АТФ/АДФпри возбуждении нейронов активирует ключевые ферменты основных путей энергообмена мозга –
гликолиза и ЦТК –
фосфофруктокиназу и
изоцитратдегидрогеназу.
Активность изоцитратдегидрогеназы
максимальна даже при утилизации
глюкозы в состоянии покоя,
поэтому при повышении
энергопотребления нет
возможностей ускорения ЦТК (!)
19.
Окислительноедекарбоксилирование пирувата –
(конечного продукта аэробного гликолиза)
это единственный источник
ацетил-КоА для ЦТК – основного
пути энергообеспечения мозга.
Поэтому нейроны очень чувствительны к нарушениям функционирования любого из компонентов
пируватдегидрогеназного комплекса
(напр., к дефициту
тиаминпирофосфата при гипоили авитаминозе В1).
20.
ГликолизЦТК
21.
Мозговая ткань также способна ик анаэробному гликолизу,
причём в клетках мозга около
10% общей активности
лактатдегидрогеназы
проявляется в митохондриях (!),
что способствует более полному
и эффективному использованию
конечных продуктов гликолиза.
В нейронах преобладает
«аэробная» изоформа ЛДГ1, а в
клетках глии «анаэробная» ЛДГ5.
22.
Нервная ткань состоит из трехосновных типов клеток:
нейронов или нейроцитов
(собственно нервных клеток ),
нейроглии (макроглии),
заполняющей промежутки между
ними,
мезенхимных элементов
(микроглии, включающей, в частности, глиальные макрофаги клетки Ортеги).
Основная масса мозга
представлена первыми двумя
типами клеточных элементов.
23.
Нейроны – структурная и функциональная единица нервнойсистемы; высокоспециализированные не делящиеся клетки.
Функциональные части:
воспринимающая – дендриты
и мембрана сомы;
интеграционная – сома с аксонным холмиком;
передающая
аксонный холмик с аксоном.
24.
Свойства нейронов:способность воспринимать
раздражения
переходить в состояние
возбуждения
генерировать и проводить
электрохимические импульсы
передавать их в местах
межклеточных контактов (синапсах) с помощью нейротрансмиттеров, синтезируемых
самими нейронами.
25.
Плазмалемма сомы:состоит из липидного бислоя
со встроенными в него белками
формирует электротонический
потенциал и распространяет его
к аксонному холмику.
26.
Функции мембранныхбелков:
– канальные белки,
перемещающие ионы по
градиенту концентраци
(определяют избирательную проницаемость мембраны);
– белки-«насосы», перемещающие ионы и молекулы против градиента
концентрации, используя
энергию АТФ;
27.
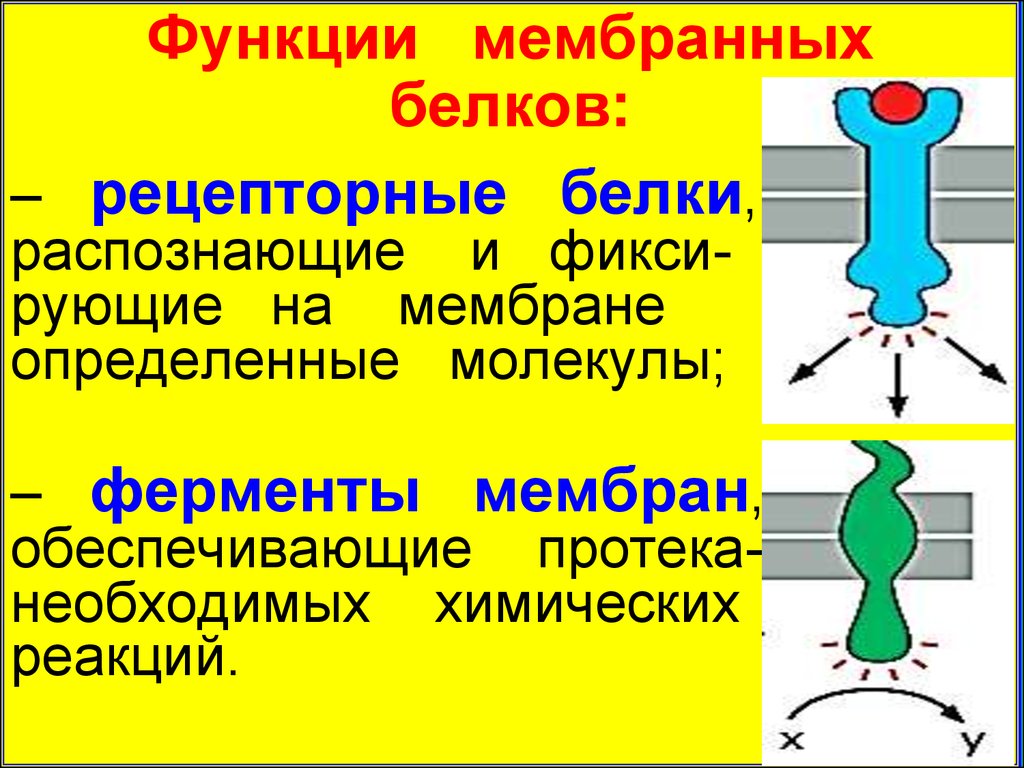
Функции мембранныхбелков:
– рецепторные белки,
распознающие и фиксирующие на мембране
определенные молекулы;
– ферменты мембран,
обеспечивающие протеканеобходимых химических
реакций.
28.
Структура ионо- и метаботропных белков-рецепторовН-холинорецептор
(ионотропный)
М-холинорецептор
(метаботропный)
29.
Структура димерногоГАМКВ – рецептора
Субъединица
GABABR2 при
взаимодействии с
ГАМК при участии
α-субъединицы
другого мембранного Gi-белка
ингибирует
аденилатциклазу
Субъединица
GABABR1 при
взаимодействии с
ГАМК при
участии βγсубъединиц
мембранного Gбелка активирует
К+-канал
30.
Эмитоходрии
ЭПШР
(тигроид)
ядро с РНП
комплекс
Гольджи
лизосомы
31.
Ядро нейрона содержит генетический материал, определяляющий порядок дифференцирования, конечную форму,типичные для него связи.
Ядрышки с большим
количеством РНК обеспечивают
образование и накопление
субстанции Ниссля. Существует
определенная зависимость между
развитием в онтогенезе ядрышек
и формированием первичных
поведенческих реакций.
32.
Субстанция Ниссля (тигроид) –хорошо развитая гранулярная
эндоплазматическая сеть с правильно ориентированным расположением мембран, содержащая
много РНК, липидов, гликогена.
Тигроид – показатель функциональной активности нейрона.
У новорожденных большое его
количество в нейронах, обеспечивающих врожденные
жизненно важные рефлексы,
а в нейронах лобной доли коры
его практически нет (!!!).
33.
Классификация глиальныхэлементов:
Макроглия:
астроциты
олигодендроциты
эпендимоциты
Микроглия –
тканевые макрофаги
34.
микроглияиммунокомпетентные
клетки ЦНС, противостоящие
вторжению чужеродных
веществ и лизирующие
погибшие нейроны.
При ишемии индуцируют синтез
нейротоксинов, сигнальных
молекул, клеточных регуляторов, трофических факторов,
уменьшают зону рубцевания.
35.
НЕЙРОГЛИЯГлавные отличия от нейронов:
выполняют роль межклеточного вещества соединительной
ткани;
активно делятся (именно с
этим связано возникновение
ляющего числа опухолей
невозбудимы (в
ЦНС);
их мембранах
потенциал-зависимых
для Ca2+ и Na+).
очень мало
каналов
подав-
36.
Функции клеток нейроглии:создают для нейронов специфическую среду и условия
генерации и передачи нервных
импульсов;
препятствуют гиперактивности
нейронов и восстанавливают их
готовность к восприятию новых
импульсов;
регулируют состав внеклеточной жидкости (содержание амино
кислот, глюкозы, депо и буфер К+)
37.
Функции клеток нейроглии:обеспечивают нейроны пита-
тельными веществами и выводят
продукты метаболизма;
непосредственно осуществляют
значительную часть метаболических процессов нейрона;
участвуют в условно-рефлекторной деятельности мозга и
в процессах формирования
памяти
38.
Функции клеток нейроглии:структурный компонент гематоэнцефалического барьера –
ферментативный барьер
(высокая активность
холинэстеразы, моноаминоксидаз,
катехол-О- метилтрансфераз,
фосфатаз, ДОФА-декарбоксилазы,
γ-глутамилтрансферазы
и других ферментов).
39.
Олигодендроцитысодержат большое количество
рибосом и отвечают за
образование миелина.
Кроме этого, они секретируют
нейротрофические факторы,
участвующие в процессах
регенерации и дегенерации
нервных волокон, а также в
обмене веществ в них.
40.
Миелин – особый видмембраны, обеспечивающий:
эффективную изоляцию аксона
высокое сопротивление и
малую емкость
препятствие продольному
распространению импульса
сальтаторное проведение
импульса, увеличение его
скорости
трофическую, барьерную и
опорную функции
41.
Химический состав миелина:Это сложный белковолипидный комплекс.
Белки составляют 25-30%
массы сухого вещества
миелиновой оболочки, а
липиды – 70-75%
(содержание липидов
лине спинного мозга
в миевыше !).
Миелин также содержит гликопротеины и гликолипиды.
42.
Бόльшую часть ЛИПИДОВмиелина составляют
фосфолипиды (43%), а все
остальное – холестерол
(28%) и цереброзиды
(галактосфинголипиды) – 29%.
В липидных слоях
миелиновых оболочек
молекулы различных
липидов имеют четкое
определенное положение.
43.
Белки миелина выполняютструктурную, стабилизирующую
и транспортную функции;
проявляют выраженные
иммуногенные свойства.
Выделено около 30 белков,
80% из них составляют:
оснόвные белки миелина;
протеолипидний комплекс
Фолча;
миелин-ассоциированный
гликопротеин P0.
44.
Особенности оснόвныхбелков миелина
(3 изоформы – 17,5; 18,5; 21,5 кД)
значительное содержание
оснόвных аминокислот (25% –
аргинин, лизин и гистидин);
очень высокая изоэлектрическая точка (рI = 12-13);
высокая степень гомологии
последовательности аминокислот у разных видов (до 90%).
45.
Особенности оснόвныхбелков миелина
(3 изоформы – 17,5; 18,5; 21,5 кД)
будучи поликатионами, образуют стабильные комплексы
с –СООН группами кислых
мембранных липидов и с
другими полярными липидами,
находящимися преимущественно
в форме цвиттер-ионов
(сфингомиелином, фосфатидилэтаноламином).
46.
Протеолипидныекомплексы Фолча
чрезвычайно
гидрофобны.
В липофилине (≈30 кД) 65%
полипептидной цепи
составляют неполярные
гидрофобные аминокислоты.
Для него характерна определённая избирательность контактов с липидами, в частности, вытеснение холестерина.
47.
Миелин-ассоциированныйгликопротеин P0
в
ЦНС
до миелинизации
(3 изоформы – 92; 107; 113 кД),
в миелине ПНС (50% всех его
белков – изоформа 107 кД).
Богат глутаматом и аспартатом;
достаточно низкое содержание
(30%), но полный набор моносахаров: галактоза, манноза,
фукоза, N-ацетилглюкозамин,
N-ацетилнейраминовая кислота.
48.
Нарушения структуры миелинагенетического, аутоиммунного,
воспалительного или иного
характера – причина тяжёлых
заболеваний ЦНС:
рассеянный склероз;
наследственная сенсомоторная нейропатия (невральная
амиотрофия Шарко-Мари-Тута);
воспалительная демиелинизирующая полирадикулоневропатия Гийена-Барре.
49.
Рассеянный склероз –следствие дезиминирования
аргинина с образованием
цитруллина в молекулах
оснόвных белков миелина (!)
Дефект гена гликопротеина P0 –
причина аутоиммунного заболевания внутреннего уха.
50.
Белки и аминокислотынервной ткани.
51.
Химический состав нервнойткани (в %%)
Химический
состав
Серое
Белое
вещество вещество
Вода
Сухой остаток,
в т.ч.
Белки
84
70
16
30
8
9
Липиды
Минеральные
вещества
5
17
1
2
52.
Белки нервной тканиПринципы классификации:
химический состав
(простые и сложные);
физико-химические свойства
(растворимые и нерастворимые, кислые и оснόвные и др.)
локализация (региональная,
клеточная и субклеточная);
функциональная роль;
метаболическая активность.
53.
Белки нервной тканиПростые белки:
нейроальбумины (фосфопротеины);
нейроглобулины (липопротеины);
катионные белки
(гистоны);
нейросклеропротеины (фибриллярные белки – нейроколлагены, нейроеластины, нейроке-
54.
Белки нервной тканиСложные белки:
липопротеины и протеолипиды А, В, С;
фосфопротеины (в
мембра-нах ядер и ядрышек);
гликопротеины (межклеточные контакты нейронов, процессы хранения информации);
нуклеопротеины
хромопротеины
55.
Белки нервной тканиСложные белки:
В клетках нервной ткани часто
образуются ещё более сложные
надмолекулярные комплексы,
непосредственно участвующие
в выполнении её
специфических функций
липонуклеопротеины,
липогликопротеины,
липонуклеогликопротеины.
56.
Нейроспецифические белки(нейрональные
и
глиальные) –
характерные только для
нервной ткани, прямо или
опосредованно участвующие в
генерации и проведении
нервных импульсов,
переработке и хранении
информации, синаптической
передаче, клеточном узнавании
и адгезии, рецепции,
образовании миелина.
57.
Нейроспецифические белки:2+
семейство Са –
связывающих белков S-100;
MAG (мyelin-associated glycoprotein);
N-CAM (neural cells adhesion
molecule);
NG-CAM (neuralglial cells
adhesion molecule);
мозговые изоферменты (альдолаза С, ВВ-КФК, -енолаза).
58.
Аминокислоты нервнойткани
Свободные аминокислоты
играют важнейшую роль в
поддержании функциональной
активности мозга, являясь:
источником синтеза белков;
источником синтеза гормонов
белковой (пептидной) природы;
нейротрансмиттерами;
источником синтеза нуклеотидов;
59.
Аминокислоты мозгаСвободные аминокислоты
играют важнейшую роль в
поддержании функциональной
активности мозга, являясь:
источником синтеза биологически активных аминов;
источником синтеза производных витаминов (НАД-Н);
средствами нейтрализации
аммониака;
источниками энергии.
60.
АминокислотыМозг Плазма Ликвор
Сравнительное
Глутамат
10,6содержание
0,05
0,23
аминокислот
N-ацетиласпартат 5,7
Глутамин
(мкмоль/г)
-
4,3
0,7
0,03
ГАМК
2,3
-
-
Аспартат
2,2
0,01
0,01
Цистатионин
1,9
-
-
Таурин
1,9
0,1
-
Глицин
1,3
0,4
0,01
Другие
3,8
2,11
0,27
61.
Глутаминовая кислотазанимает центральное место в
обмене аминокислот мозга, т.к.
она:
является нейромедиатором;
участвует в синтезе ГАМК;
участвует в синтезе
глутатиона
непосредственно временно
обезвреживает аммониак;
используется в реакциях
трансаминирования;
тесно связана с промежуточ-
62.
Другие аминокислоты мозгаметионин (синтез адреналина,
ацетилхолина, лецитина);
цистатионин (синтез сульфатидов, сульфатированных ГАГ);
цистеин (синтез таурина);
фенилаланин (синтез катехоламинов);
триптофан (синтез серотонина
и мелатонина).
63.
Нарушение метаболизмааминокислот сопровождается
значительными нарушениями
функций мозга:
фенилпировиноградная олигофрения (фенилаланин);
паркинсонизм (катехоламины);
печёночная энцефалопатия
(синтез псевдомедиаторов –
октопамина и фенилэтиламина);
цистинурия (цистатионин)
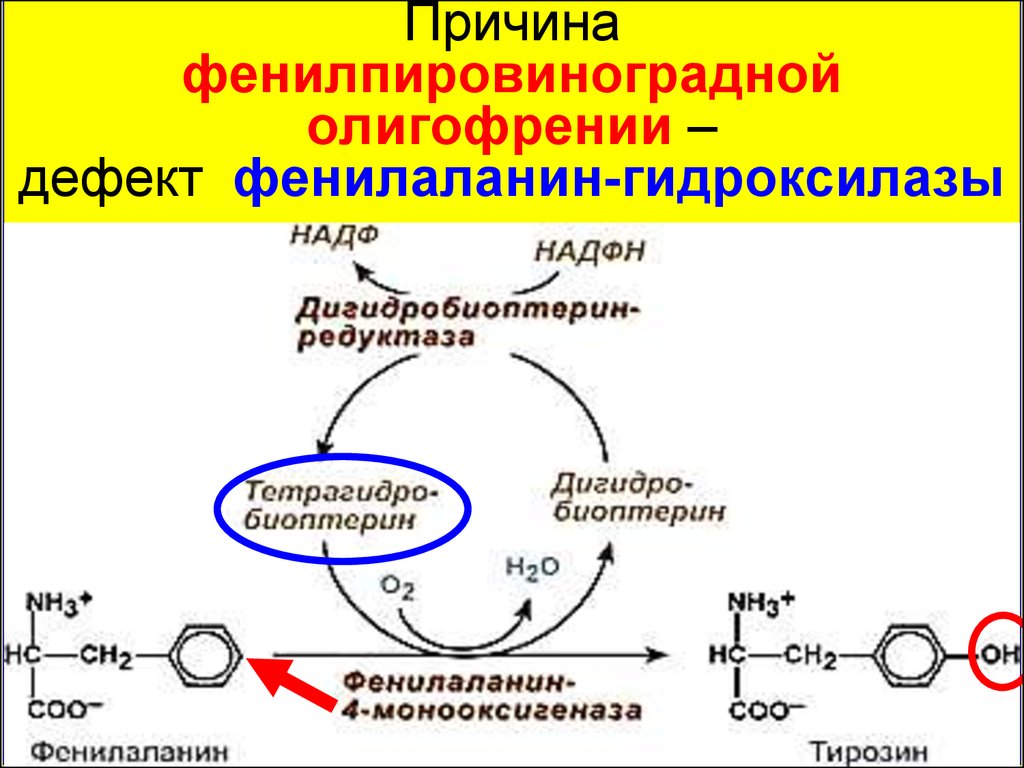
64.
Причинафенилпировиноградной
олигофрении –
дефект фенилаланин-гидроксилазы
65.
Обмен фенилаланинаПечень
(фумарат, ацетоацетат)
66.
Катаболизм фенилаланина67.
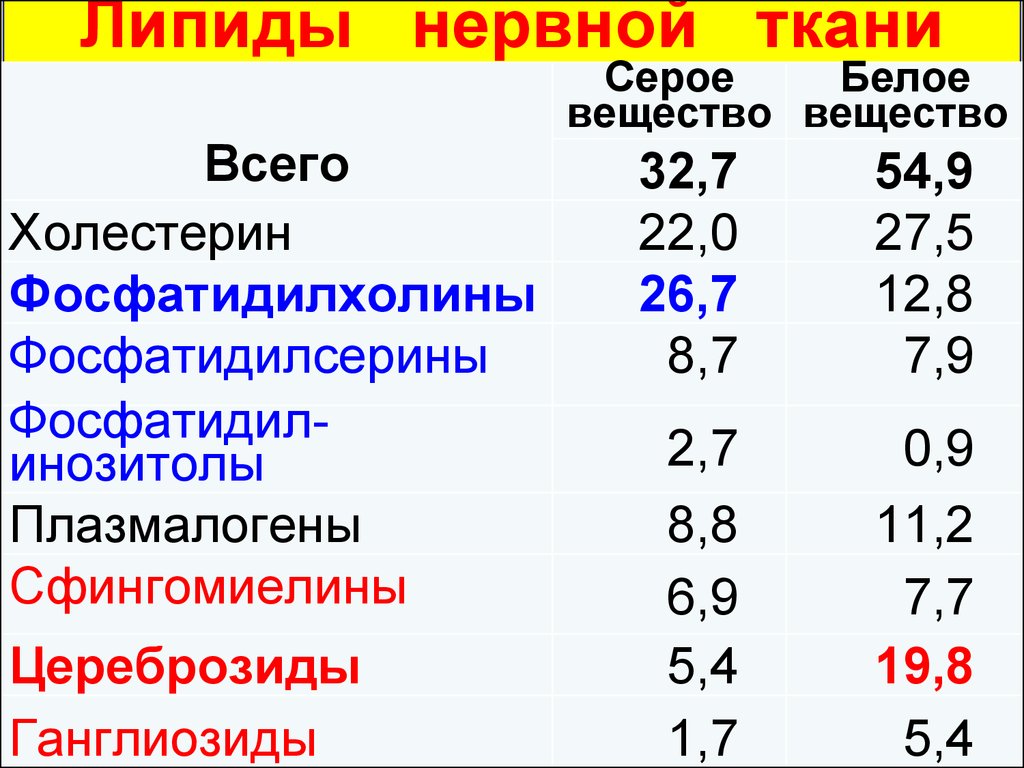
Липиды нервнойткани.
68.
Липиды нервной ткани.
Всего
Холестерин
Фосфатидилхолины
Фосфатидилсерины
Фосфатидилинозитолы
Плазмалогены
Сфингомиелины
Цереброзиды
Ганглиозиды
Серое
Белое
вещество вещество
32,7
22,0
26,7
8,7
54,9
27,5
12,8
7,9
2,7
8,8
6,9
5,4
1,7
0,9
11,2
7,7
19,8
5,4
69.
Фосфолипиды играют особуюроль в построении мембран,
имея следующие свойства:
амфифильность;
четкая ориентация на
границе раздела фаз;
способность к самопроизвольному плотному упаковыванию с формированием барьера
для диффузии молекул;
возможность образования
мицелл различной формы.
70.
Липидный состав мембрандетерминирован генетически.
Они располагаются в мембране
в соответствии с их конфигурацией, зарядом, особенностями
состава, степенью гидратации
полярных групп, что создает
структурно-функциональную
асимметрию мембран
(66% ненасыщенных ВЖК,
бόльшая часть фосфатидилэтаноламинов и фосфатидилсеринов – во внутреннем слое).
71.
Сфинголипиды играют важнуюроль в коммуникации нервной
клетки с окружающей средой,
участвуя в передаче сигналов.
Вариабельность углеводной части делает их носителями
специфичности и информации.
У взрослых почти все
цереброзиды находятся в
миелиновых оболочках, а
ганглиозиды – в нейронах.
72.
Биологическая рольсфинголипидов нервной ткани:
рецепторы внешних сигналов, в т.ч. некоторых опасных
токсинов – ботулизма (связывается с GT1), столбняка (с GD1);
отвечают за специфичность
клеточной поверхности, распознавание и адгезию клеток;
участвуют в синаптической
передаче, в реакциях адаптации и приспособления;
73.
Биологическая рольсфинголипидов нервной ткани:
связывают катионы и другие положительно заряженные
лиганды;
обнаруживают умеренные
свойства гаптенов (аллергические
и иммунологические процессы).
74.
Сфинголипиды – это производныеаминоспирта сфингозина, активно синтезирующегося нейронами
из пальмитоил-КоА и серина.
N-ацил-сфингозин (церамид) –
исходный субстрат для синтеза
сфингомиелина, ганглиозидов и
цереброзидов – образуется из
сфингозина и соответствующего
ацил-КоА.
75.
Сфинголипиды• гидрофобная часть – церамид
• гидрофильная часть – остаток,
присоединённый к ОН-группе С1
атома церамида.
Сфинголипиды в значительных
количествах содержатся в мембранах
клеток мозга, эритроцитов, эпителии.
76.
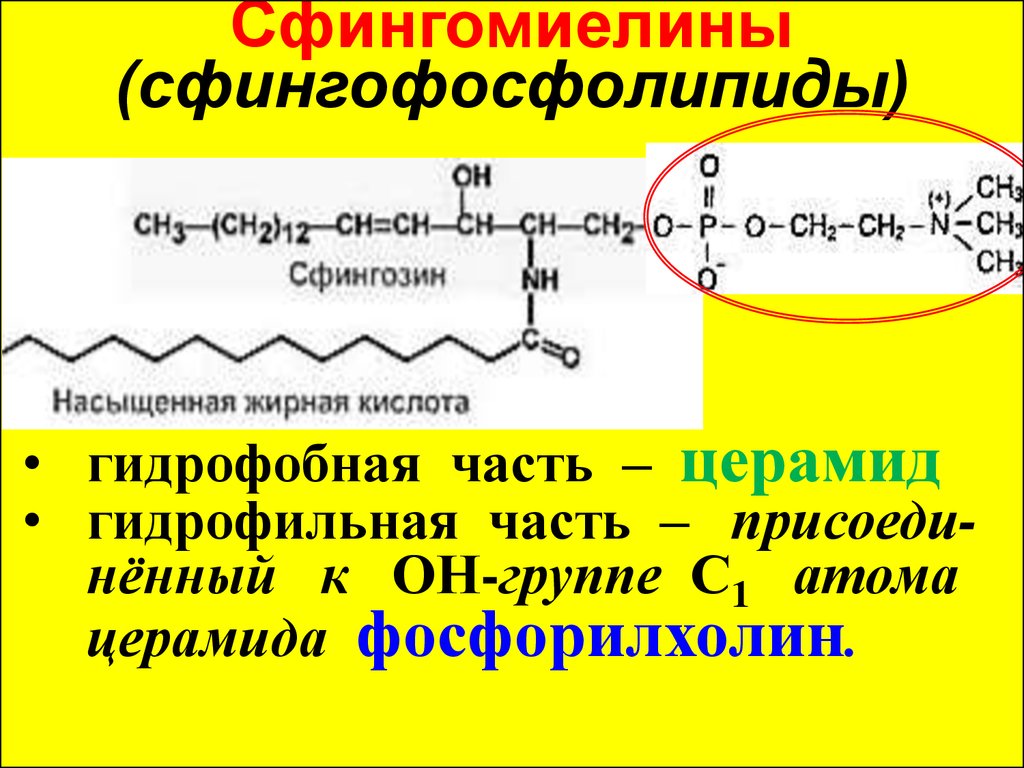
Сфингомиелины(сфингофосфолипиды)
• гидрофобная часть – церамид
• гидрофильная часть – присоединённый к ОН-группе С1 атома
церамида фосфорилхолин.
77.
Сфингогликолипиды• гидрофобная часть – церамид
• гидрофильная часть – присоединённый к ОН-группе С1 атома
церамида моно-(олиго-)сахарид.
цереброзиды –
ганглиозиды –
содержат моносодержат разветвили
лённый олигосахарид,
олигосахаридный включающий N-ацетилостаток
нейраминовую кислоту
78.
Липиды нервной тканиСхема структуры ганглиозида GM1
79.
Синтез цереброзидов и ганглиозидов из церамида происходитпутем последовательного
присоединения УДФ-моносахаридов
соответствующими полиферментными комплексами мембраносвязанных гликозилтрансфераз.
Ганглиозиды находятся
преимущественно в сером
веществе. В настоящее время
их известно около 15, наиболее
исследованные из них –
GM1, GD1a, GD1b и GT1.
80.
–Синтез (церебро/ганглио-)зидов
Гал–ГалNAc– Гал– Гл–церамид
глюкогалактоцерамид
глюкоцерамид
сиаловая
к-та
GМ2
GМ3
GМ1
(цереброзид)
(цереброзид)
81.
Номенклатураганглиозидов
–
–
–
Гал–ГалNAc– Гал– Гл–церамид
сиаловая сиаловая
к-та
к-та
сиаловая
к-та
GМ1
GD1b
GD1a
82.
Номенклатураганглиозидов
–
–
–
Гал–ГалNAc– Гал– Гл–церамид
сиаловая сиаловая
к-та
к-та
сиаловая
к-та
GD1b
GТ1
83.
Катаболизм ганглиозидов происходит при участии лизосомальныхгликозидаз и нейраминидазы.
Нарушение их активности – причина
сфинголипидозов и ганглиозидозов:
болезнь Тея-Сакса (дефект
гексозаминидазы);
болезни Гоше и Краббе
(дефект β-глюко- или β-галактоцереброзидаз соответственно);
болезнь Фарбера (дефект
церамидазы);
болезнь Нимана-Пика (дефект сфингомиелиназы).
84.
Катаболизм ганглиозида GM1болезнь
Краббе
дефект
болезнь
Тея-Сакса
ганглиозидоз
GM1
болезнь
Фарбера
–––
сиалидоз
– дефект
болезнь
Гоше
––дефект
β-галактцереброзидазы
дефект
гексозаминидазы)
дефект
β-галактозидазы
дефект
церамидазы
нейраминидазы
β-глюкоцереброзидазы
накопление
накопление
накопление
накопление
лактоGM3
глюкоGM2
церамида
церамида
церамида
85.
Генерация и проведениенервного импульса.
Морфо-функциональная
структура синапсов.
Нейротрансмиттеры.
86.
87.
Структура различныхрецепторов:
Н-ХР
М-ХР
88.
Схема взаимодействия АцХи холиноблокаторов с Н-ХР
89.
Структура АцХ, атропина ифосфорорганического эфира
90.
Структура различныхрецепторов:
Адренергичекие
синасы
ГАМКрецептор
91.
Критерии (признаки)медиаторов :
– избирательность их локализации
в нервных окончаниях;
– присутствие в пресинаптических
терминалях ферментов их синтеза;
– Са2+-зависимое высвобождение
медиаторов при стимуляции нервных окончаний в количествах, соответствующих величине стимулов;
– наличие на постсинаптической
мембране особых чувствительных
к медиатору участков – рецепторов;
92.
Критерии (признаки)медиаторов:
– наличие в синапсах и нервных
терминалях ферментов, участвующих в разрушении медиаторов;
– наличие системы обратного захвата медиатора или его составляющих в пресинаптические терминали;
– возможность влияния на эффекты медиатора с помощью фармакологических средств.
93.
Химическая классификациямедиаторов.
1. Сложные эфиры – ацетилхолин.
2. Биогенные амины:
– катехоламины (дофамин, норадреналин, адреналин);
– серотонин;
– гистамин.
3. Аминокислоты:
– γ-аминомасляная кислота (ГАМК);
– глутаминовая кислота;
– глицин;
– аргинин;
– таурин.
94.
Химическая классификациямедиаторов.
4. Пептиды:
– опиоидные пептиды (эндорфины,
мет-энкефалин, лей-энкефалин);
– вещество «P»;
– вазоактивный интестинальный
пептид;
– соматостатин.
5. Пуриновые соединения: АТФ.
6. Неорганические вещества с малой молекулярной массой:
– NO;
– CO.
95.
Функциональнаяклассификация медиаторов.
1.
–
–
–
2.
–
–
–
–
–
–
Возбуждающие медиаторы:
ацетилхолин;
глутаминовая кислота;
аспарагиновая кислота.
Тормозные медиаторы:
ГАМК;
глицин;
вещество «P»;
дофамин;
серотонин;
АТФ.
96.
Функциональнаяклассификация медиаторов.
3. Возбуждающие
медиаторы:
– адреналин;
– норадреналин;
– гистамин.
и тормозные
97.
Нейротрансмиттеры:Характер
действия
Главная функция
возбуждение торможение
Глутамат
НейроАспартат
медиаторы Ацетилхолин
ГАМК
Глицин
Серотонин
Нейро-
Норадреналин
Аденозин
модуляторы
Серотонин
Дофамин
98.
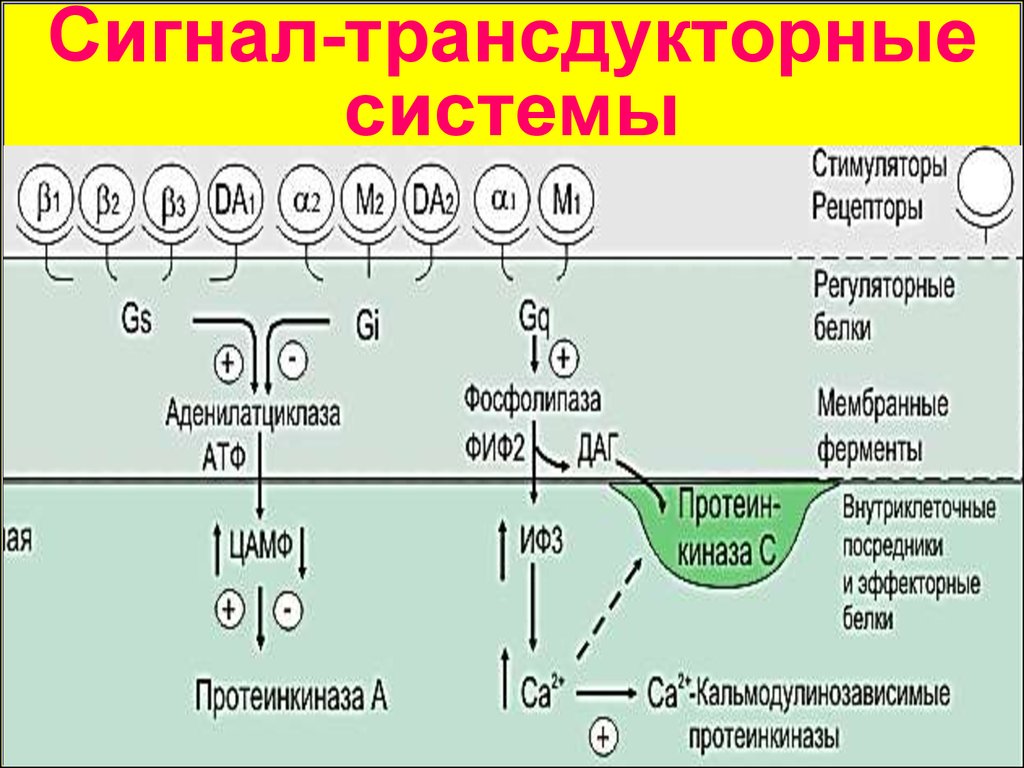
Сигнал-трансдукторныесистемы
99.
Сигнал-трансдукторные системыВторичные посредники
ИФ 3
Арахидоновая
кислота
НА (α1)
гистамин
(Н1)
Gs (β1, β2)
Gi (α2, М2)
Gq
–
Аденилатциклаза
ФЛ С
ФЛ А2
ц-АМФ
ИФ-1,4,5
и ДАГ
Арахидоновая к-та
ПК А
Ca2+
ПК С
А1, А2а,
А2b, А3
ц-АМФ
Медиаторы/
Рецепторы
Первичный
посредник
Первичный
эффектор
Вторичный
посредник
Вторичный
эффектор
НА (α2, β1-2)
АцХ (М2)