


















 Биология
БиологияПохожие презентации:










Потенциал покоя. Потенциал действия. Проведение нервного импульса
1.
Тема:Потенциал покоя. Потенциал действия.
Проведение нервного импульса
Задачи:
Дать характеристику мембранному потенциалу и проведению
нервного импульса
Пименов А.В.
2.
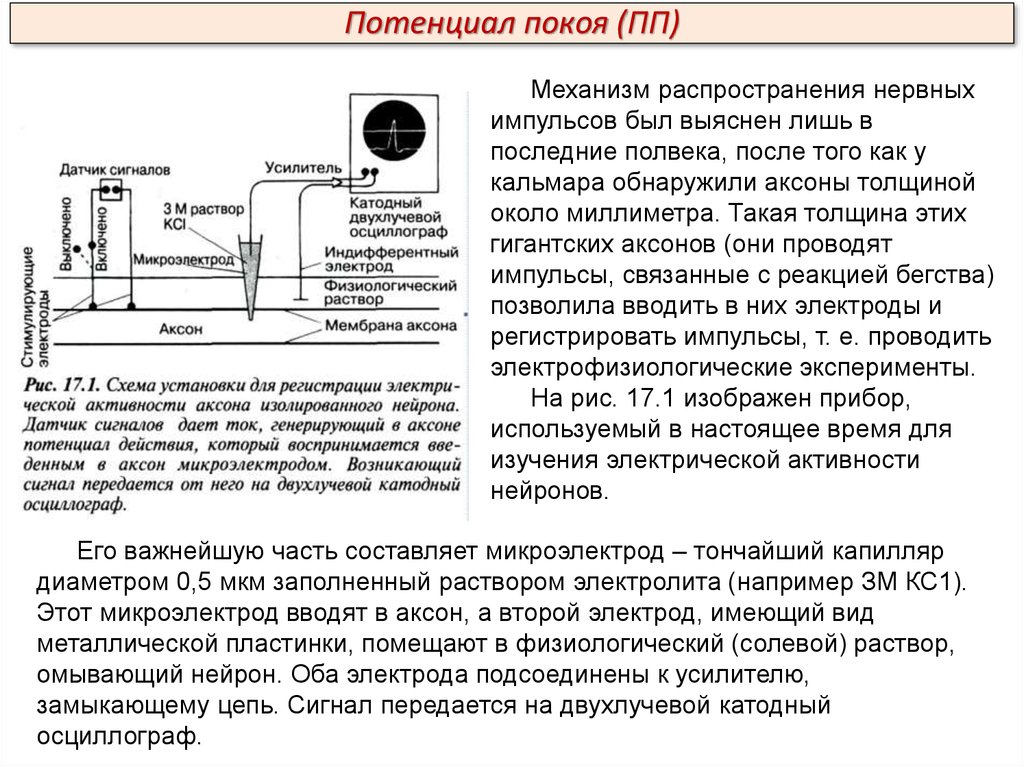
Потенциал покоя (ПП)Механизм распространения нервных
импульсов был выяснен лишь в
последние полвека, после того как у
кальмара обнаружили аксоны толщиной
около миллиметра. Такая толщина этих
гигантских аксонов (они проводят
импульсы, связанные с реакцией бегства)
позволила вводить в них электроды и
регистрировать импульсы, т. е. проводить
электрофизиологические эксперименты.
На рис. 17.1 изображен прибор,
используемый в настоящее время для
изучения электрической активности
нейронов.
Его важнейшую часть составляет микроэлектрод – тончайший капилляр
диаметром 0,5 мкм заполненный раствором электролита (например ЗМ КС1).
Этот микроэлектрод вводят в аксон, а второй электрод, имеющий вид
металлической пластинки, помещают в физиологический (солевой) раствор,
омывающий нейрон. Оба электрода подсоединены к усилителю,
замыкающему цепь. Сигнал передается на двухлучевой катодный
осциллограф.
3.
Потенциал покоя (ПП)Когда кончик микроэлектрода
проходит сквозь плазматическую
мембрану аксона, лучи осциллографа
раздвигаются. Расстояние между
лучами на экране показывает разность
потенциалов между электродами в
цепи. Разность потенциалов
покоящейся клетки называется
потенциалом покоя. В сенсорных,
железистых клетках, нейронах и
мышечных клетках измеряемая
описанным способом величина
меняется в зависимости от их
активности, поэтому такие клетки
называют возбудимыми.
На мембранах всех остальных живых клеток тоже существует подобная
разность потенциалов, известная как мембранный потенциал, но в этих
клетках она остается постоянной, поэтому их называют невозбудимыми
клетками.
4.
Потенциал покоя (ПП)Нейроны
Секреторные клетки
Мышечные клетки
Рецепторные клетки
4
5.
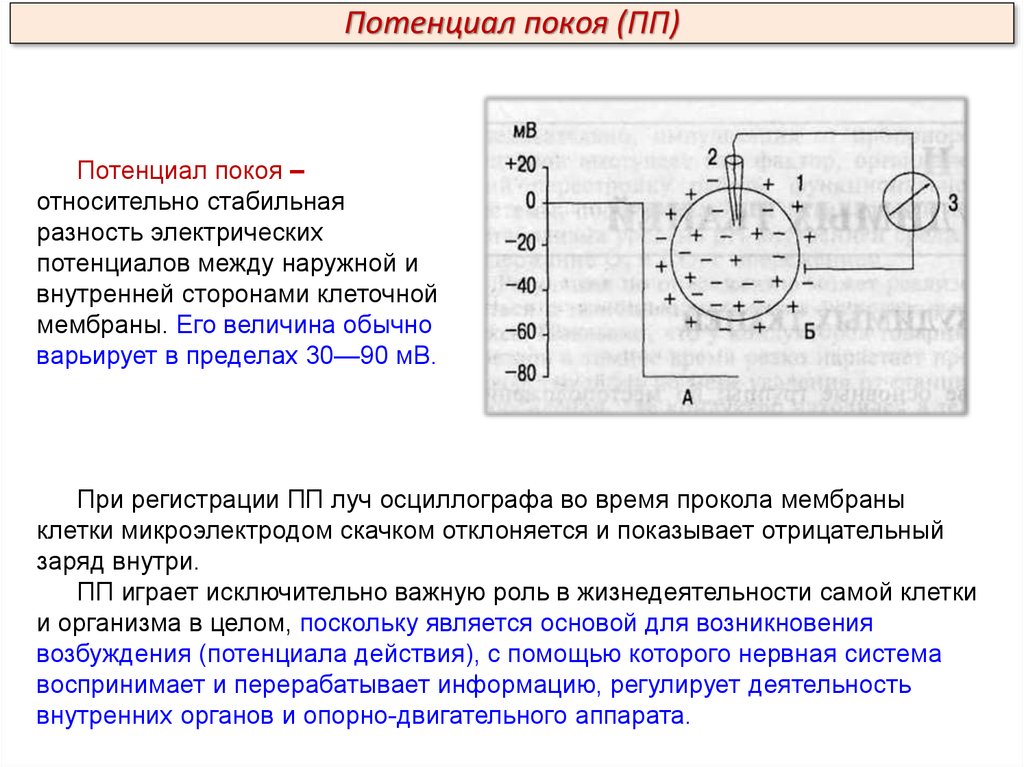
Потенциал покоя (ПП)Потенциал покоя –
относительно стабильная
разность электрических
потенциалов между наружной и
внутренней сторонами клеточной
мембраны. Его величина обычно
варьирует в пределах 30—90 мВ.
При регистрации ПП луч осциллографа во время прокола мембраны
клетки микроэлектродом скачком отклоняется и показывает отрицательный
заряд внутри.
ПП играет исключительно важную роль в жизнедеятельности самой клетки
и организма в целом, поскольку является основой для возникновения
возбуждения (потенциала действия), с помощью которого нервная система
воспринимает и перерабатывает информацию, регулирует деятельность
внутренних органов и опорно-двигательного аппарата.
6.
Потенциал покоя (ПП)В нервных и мышечных клетках концентрация К+ внутри клетки в 30-40 раз
больше, чем вне клетки; концентрация Na+ вне клетки в 10-12 раз больше,
нежели в клетке. Ионов Сl- вне клетки в 30-50 раз больше, чем внутри клетки.
В клетке имеется небольшое количество ионов Mg2+, Са2+. Кальций в
свободном состоянии находится в основном вне клетки. Он содержится также
в эндоплазматическом ретикулуме; в гиалоплазме его очень мало.
Это обусловливается
отчасти активным транспортом
Са2+ наружу через клеточную
мембрану, отчасти
поглощением его
эндоплазматическим
ретикулумом (это резервуар
для Са2+) и другими
органеллами, например
митохондриями.
7.
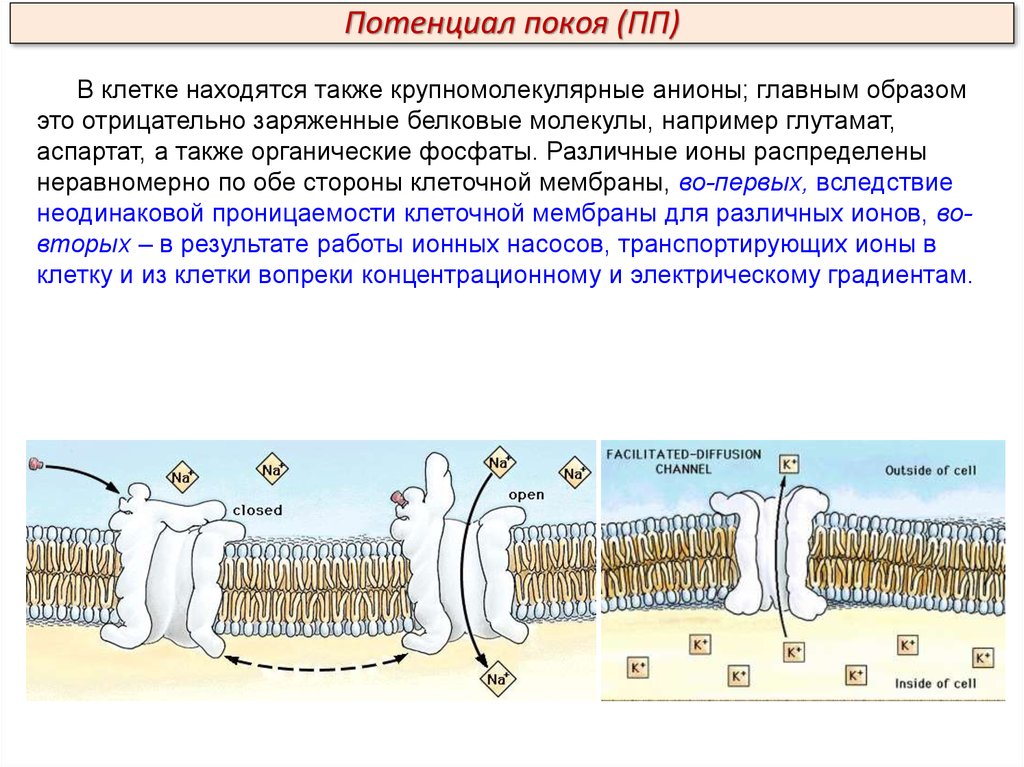
Потенциал покоя (ПП)В клетке находятся также крупномолекулярные анионы; главным образом
это отрицательно заряженные белковые молекулы, например глутамат,
аспартат, а также органические фосфаты. Различные ионы распределены
неравномерно по обе стороны клеточной мембраны, во-первых, вследствие
неодинаковой проницаемости клеточной мембраны для различных ионов, вовторых – в результате работы ионных насосов, транспортирующих ионы в
клетку и из клетки вопреки концентрационному и электрическому градиентам.
8.
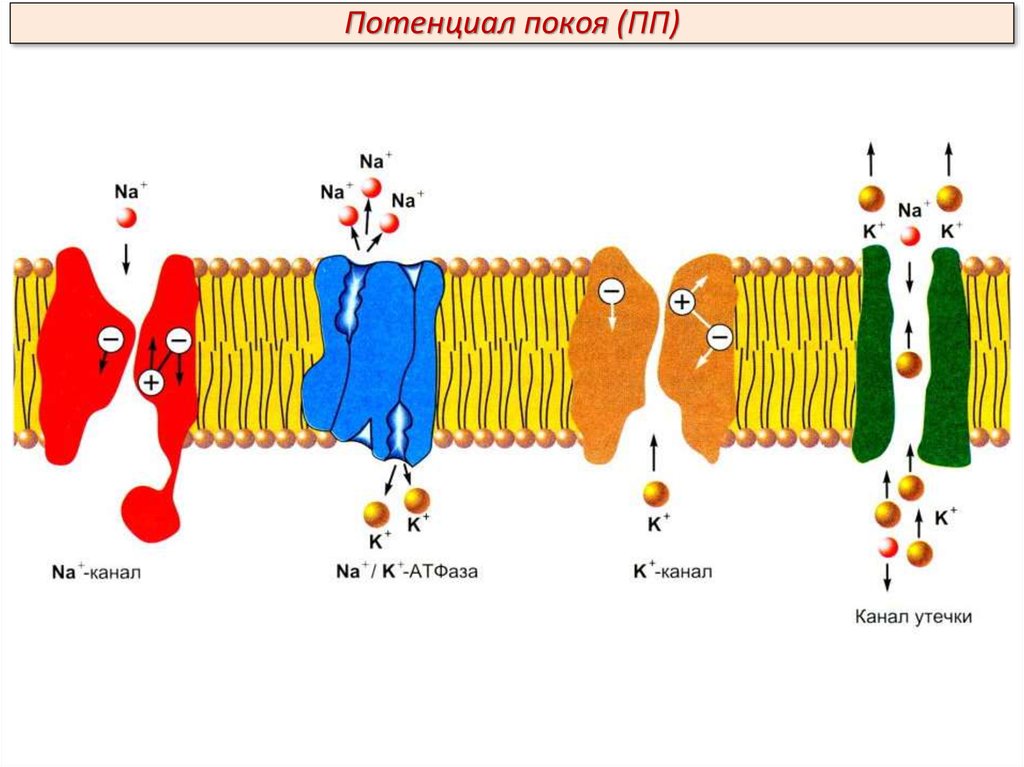
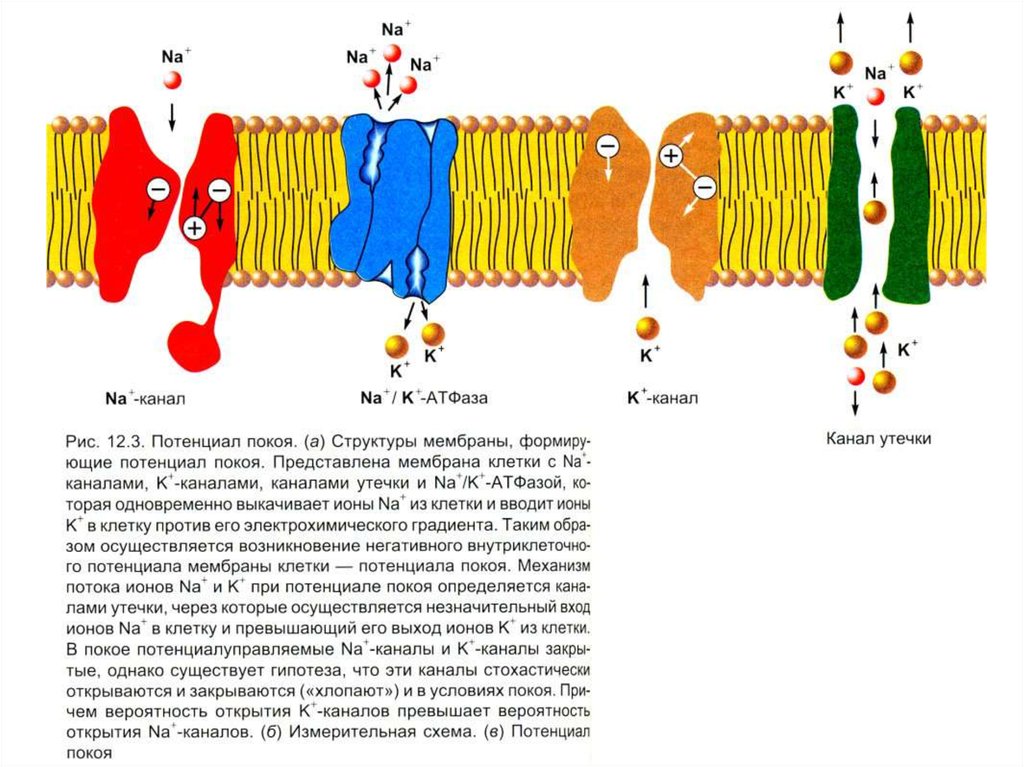
Потенциал покоя (ПП)Проницаемость клеточной мембраны – это ее способность пропускать
воду, незаряженные и заряженные частицы (ионы) согласно законам
диффузии и фильтрации. Проницаемость клеточной мембраны определяется
следующими факторами:
1. Наличием в составе мембраны различных ионных каналов – управляемых
(с воротным механизмом) и неуправляемых (каналов утечки);
2. Размерами каналов и размерами частиц;
3. Растворимостью частиц в мембране (клеточная мембрана проницаема для
растворимых в ней липидов и непроницаема для пептидов).
9.
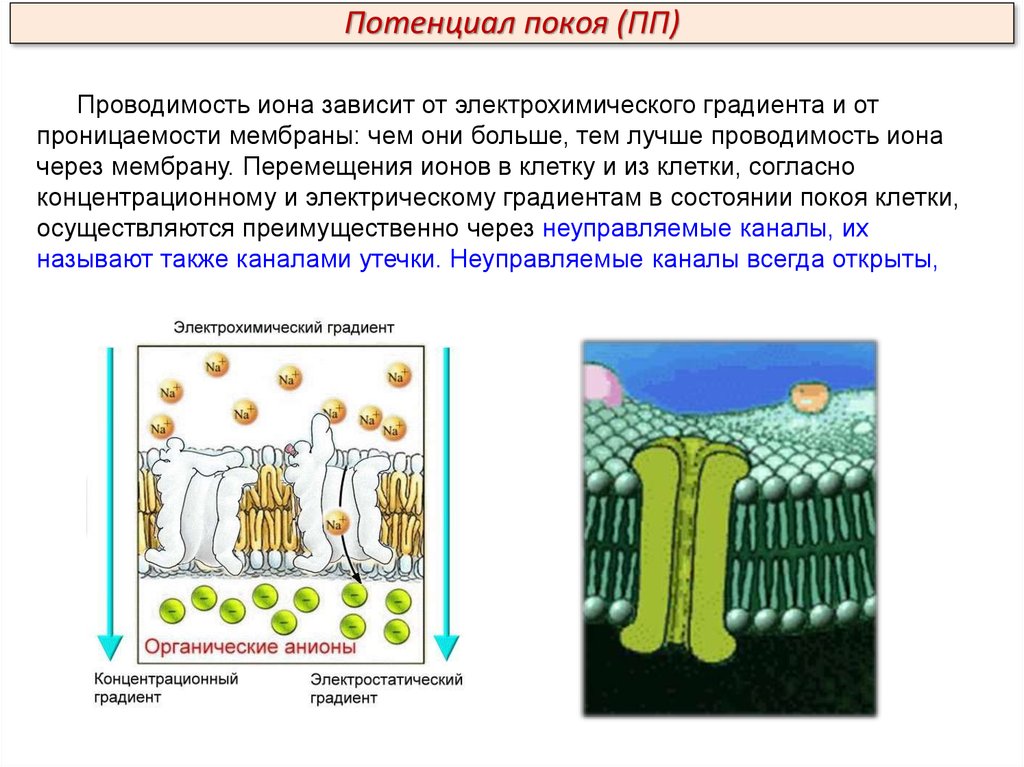
Потенциал покоя (ПП)Проводимость иона зависит от электрохимического градиента и от
проницаемости мембраны: чем они больше, тем лучше проводимость иона
через мембрану. Перемещения ионов в клетку и из клетки, согласно
концентрационному и электрическому градиентам в состоянии покоя клетки,
осуществляются преимущественно через неуправляемые каналы, их
называют также каналами утечки. Неуправляемые каналы всегда открыты,
10.
Потенциал покоя (ПП)11.
Потенциал покоя (ПП)Na+ и К+ в покоящейся клетке перемещаются через мембрану согласно
законам диффузии, при этом К+ из клетки выходит в значительно большем
количестве, чем входит Na+ в клетку, поскольку проницаемость клеточной
мембраны для К+ примерно в 25 раз больше проницаемости для Na+.
Органические анионы из-за
своих больших размеров не могут
выходить из клетки, поэтому внутри
клетки в состоянии покоя
отрицательных ионов оказывается
больше, чем положительных. По
этой причине клетка изнутри имеет
отрицательный заряд.
12.
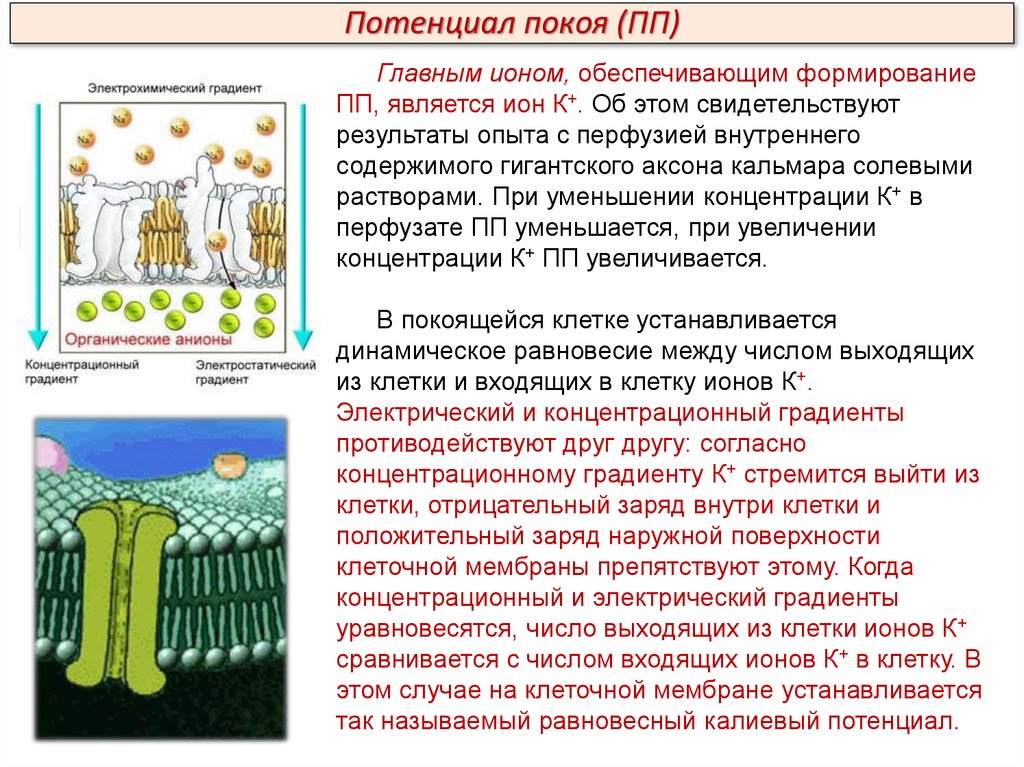
Потенциал покоя (ПП)Главным ионом, обеспечивающим формирование
ПП, является ион К+. Об этом свидетельствуют
результаты опыта с перфузией внутреннего
содержимого гигантского аксона кальмара солевыми
растворами. При уменьшении концентрации К+ в
перфузате ПП уменьшается, при увеличении
концентрации К+ ПП увеличивается.
В покоящейся клетке устанавливается
динамическое равновесие между числом выходящих
из клетки и входящих в клетку ионов К+.
Электрический и концентрационный градиенты
противодействуют друг другу: согласно
концентрационному градиенту К+ стремится выйти из
клетки, отрицательный заряд внутри клетки и
положительный заряд наружной поверхности
клеточной мембраны препятствуют этому. Когда
концентрационный и электрический градиенты
уравновесятся, число выходящих из клетки ионов К+
сравнивается с числом входящих ионов К+ в клетку. В
этом случае на клеточной мембране устанавливается
так называемый равновесный калиевый потенциал.
13.
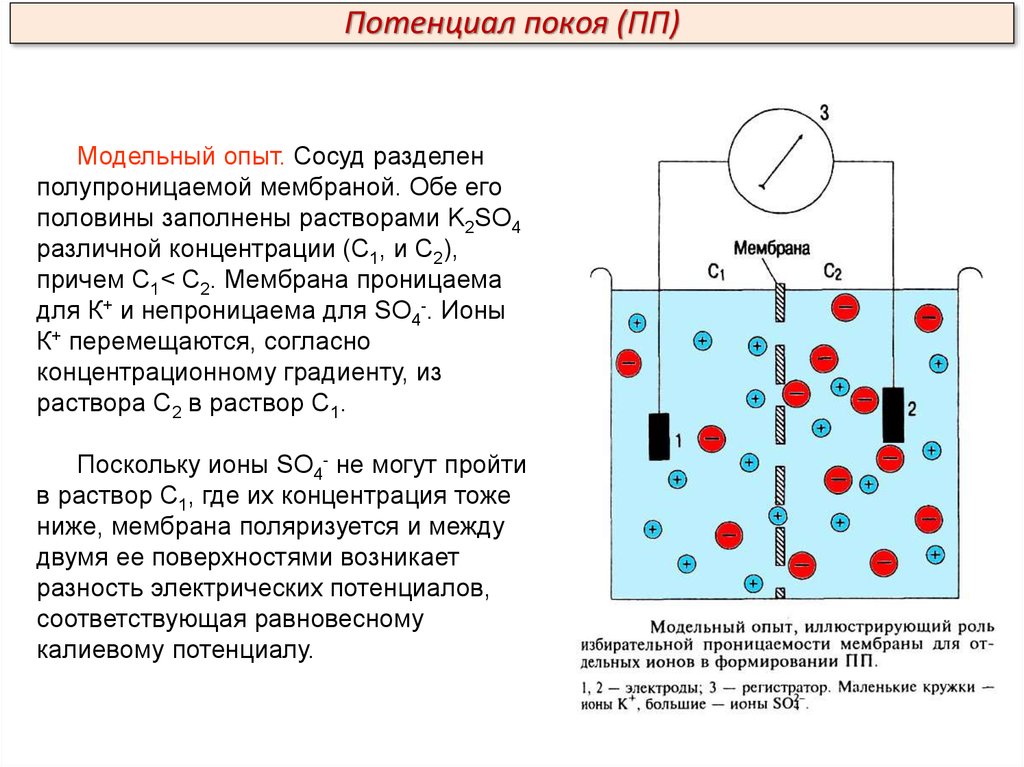
Потенциал покоя (ПП)Модельный опыт. Сосуд разделен
полупроницаемой мембраной. Обе его
половины заполнены растворами K2SO4
различной концентрации (С1, и С2),
причем С1< С2. Мембрана проницаема
для К+ и непроницаема для SO4-. Ионы
К+ перемещаются, согласно
концентрационному градиенту, из
раствора С2 в раствор С1.
Поскольку ионы SO4- не могут пройти
в раствор С1, где их концентрация тоже
ниже, мембрана поляризуется и между
двумя ее поверхностями возникает
разность электрических потенциалов,
соответствующая равновесному
калиевому потенциалу.
14.
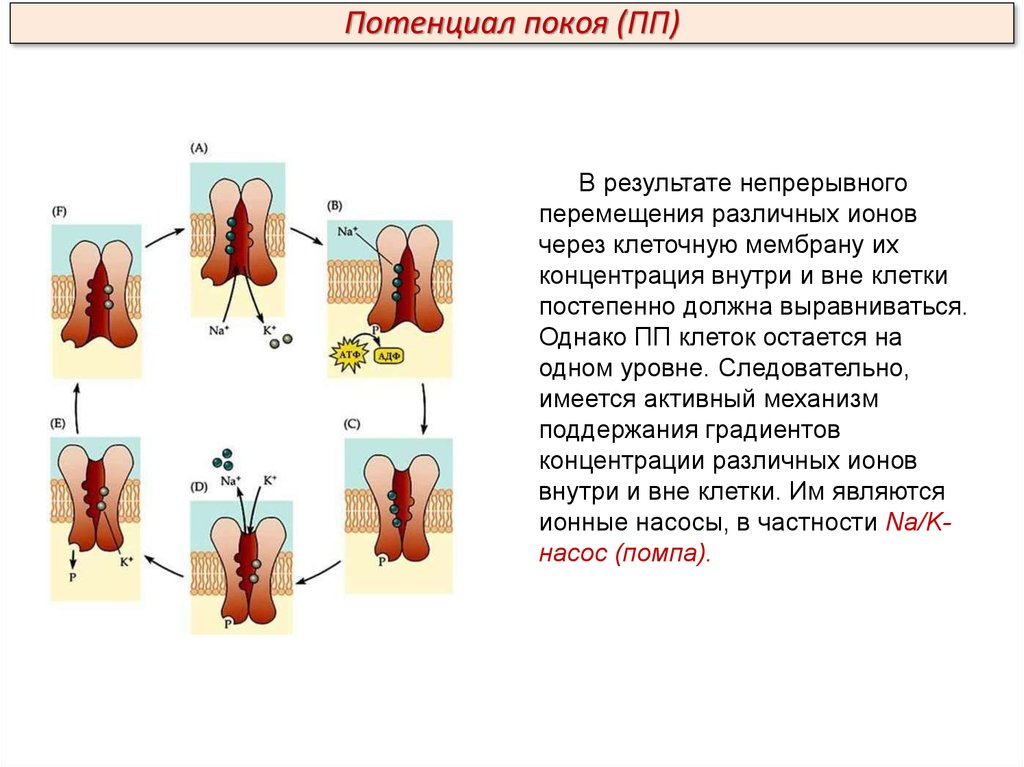
Потенциал покоя (ПП)В результате непрерывного
перемещения различных ионов
через клеточную мембрану их
концентрация внутри и вне клетки
постепенно должна выравниваться.
Однако ПП клеток остается на
одном уровне. Следовательно,
имеется активный механизм
поддержания градиентов
концентрации различных ионов
внутри и вне клетки. Им являются
ионные насосы, в частности Na/Kнасос (помпа).
15.
Потенциал покоя (ПП)Величина ПП изменяется в различных условиях деятельности клетки,
вследствие этого колеблется и ее возбудимость, например при изменении
концентрации Са2+, рН среды.
Когда концентрация Са2+ в среде повышается, клетка становится менее
возбудимой, поскольку возрастает мембранный потенциал, а когда
концентрация Са2+ снижается, возбудимость клетки возрастает, так как
мембранный потенциал уменьшается. Такое повышение возбудимости лежит
в основе синдрома тетании, связанного с дефицитом Са2+ в крови.
16.
Подведем итоги:Какие клетки относятся к возбудимым?
Нервные, мышечные, рецепторные и секреторные клетки.
Какова концентрация ионов натрия снаружи клетки и под мембраной?
Концентрация Na+ вне клетки в 10—12 раз больше, нежели в клетке.
Какова концентрация ионов калия снаружи клетки и под мембраной?
К+ внутри клетки в 30—40 раз больше, чем вне клетки.
Какова концентрация ионов хлора снаружи клетки и под мембраной?
Ионов Сl вне клетки в 30—50 раз больше, чем внутри клетки.
Какова сравнительная проницаемость мембраны для ионов К+ и Na+?
Проницаемость клеточной мембраны для К+ примерно в 25 раз больше
проницаемости для Na+.
Какой ион, в основном, обеспечивает потенциал покоя (ПП)?
Главным ионом, обеспечивающим формирование ПП, является ион К+. Об
этом свидетельствуют результаты опыта с перфузией внутреннего
содержимого гигантского аксона кальмара солевыми растворами. При
уменьшении концентрации К+ в перфузате ПП уменьшается, при
увеличении концентрации К+ ПП увеличивается.
Что характерно для натриевых каналов когда клетка находится в состоянии
покоя?
Управляемые натриевые каналы закрыты.
17.
Подведем итоги:Что характерно для калиевых каналов когда клетка находится в состоянии
покоя?
Калиевые каналы в состоянии покоя открыты.
Чему равен потенциал покоя в возбудимых клетках?
При проведении измерений потенциал окружающей клетку среды
принимают за величину, равную нулю. Относительно нулевого
потенциала внешней среды потенциал внутренней среды клетки, как
отмечалось выше, составляет величину порядка —60—90 мВ.
Имеется ли мембранный потенциал в невозбудимых клетках?
На мембранах всех остальных живых клеток тоже существует подобная
разность потенциалов, известная как мембранный потенциал, но в этих
клетках она остается постоянной, поэтому их называют невозбудимыми
клетками.
Что такое пассивный ионный транспорт и активный транспорт ионов?
Электрические свойства ионов определяются их зарядами: одноименные
заряды отталкиваются, противоположные – притягиваются. На
движение ионов влияет также их концентрация в растворе. Ион имеет
тенденцию перемещаться по градиенту концентрации, т. е. туда, где
она ниже. Результирующее движение зависит от соотношения зарядов и
концентраций по обе стороны мембраны. Это движение происходит без
затраты энергии и называется пассивным транспортом.
18.
Подведем итоги:Как калий-натриевый насос поддерживает потенциал покоя?
Для работы этого насоса необходима энергия АТФ, а действует он по
принципу антипорта: «выкачивание» трех ионов Na+ из аксона сопряжено с
«закачиванием» в него двух ионов К+ В целом траты энергии на активный
транспорт веществ огромны: лишь Na/K-насос потребляет 1/5 всей
энергии, расходуемой организмом в покое. За 1 с один Na/K-насос (одна
молекула белка) переносит 150—600 ионов Na+.
19.
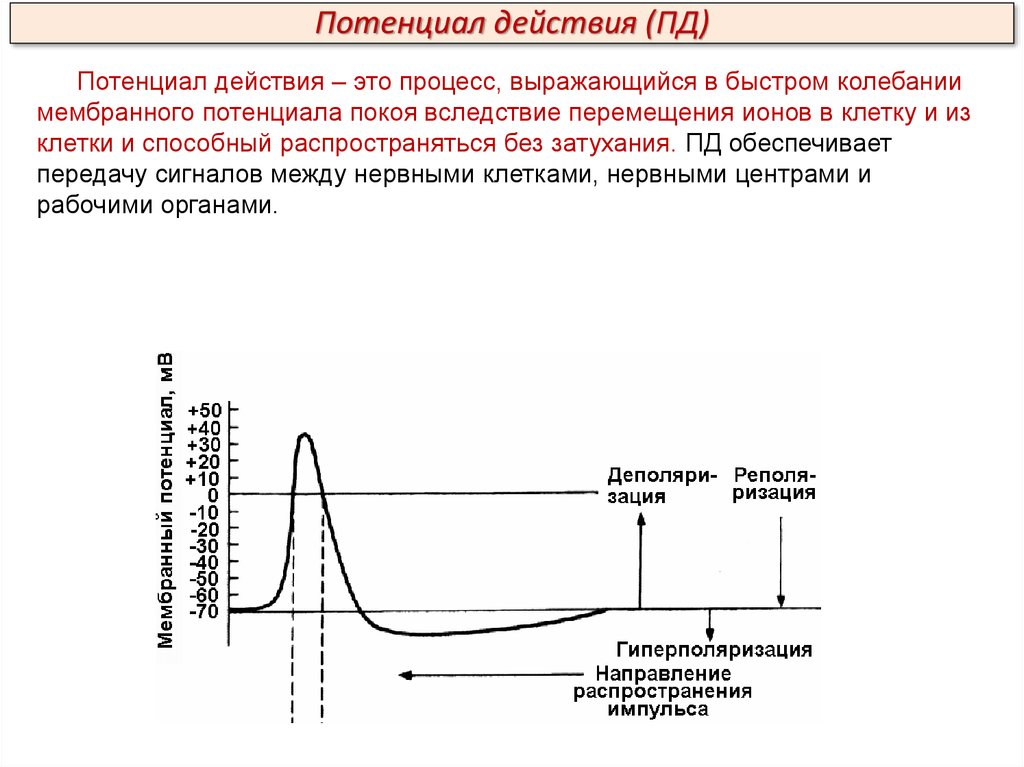
Потенциал действия (ПД)Потенциал действия – это процесс, выражающийся в быстром колебании
мембранного потенциала покоя вследствие перемещения ионов в клетку и из
клетки и способный распространяться без затухания. ПД обеспечивает
передачу сигналов между нервными клетками, нервными центрами и
рабочими органами.
20.
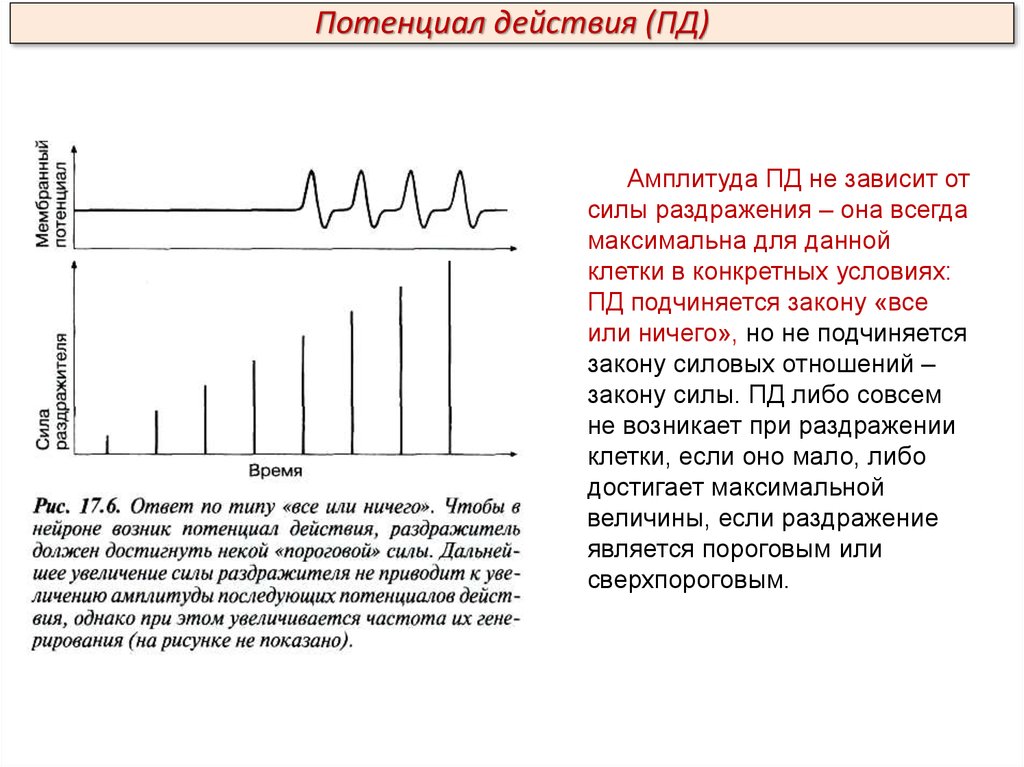
Потенциал действия (ПД)Амплитуда ПД не зависит от
силы раздражения – она всегда
максимальна для данной
клетки в конкретных условиях:
ПД подчиняется закону «все
или ничего», но не подчиняется
закону силовых отношений –
закону силы. ПД либо совсем
не возникает при раздражении
клетки, если оно мало, либо
достигает максимальной
величины, если раздражение
является пороговым или
сверхпороговым.
21.
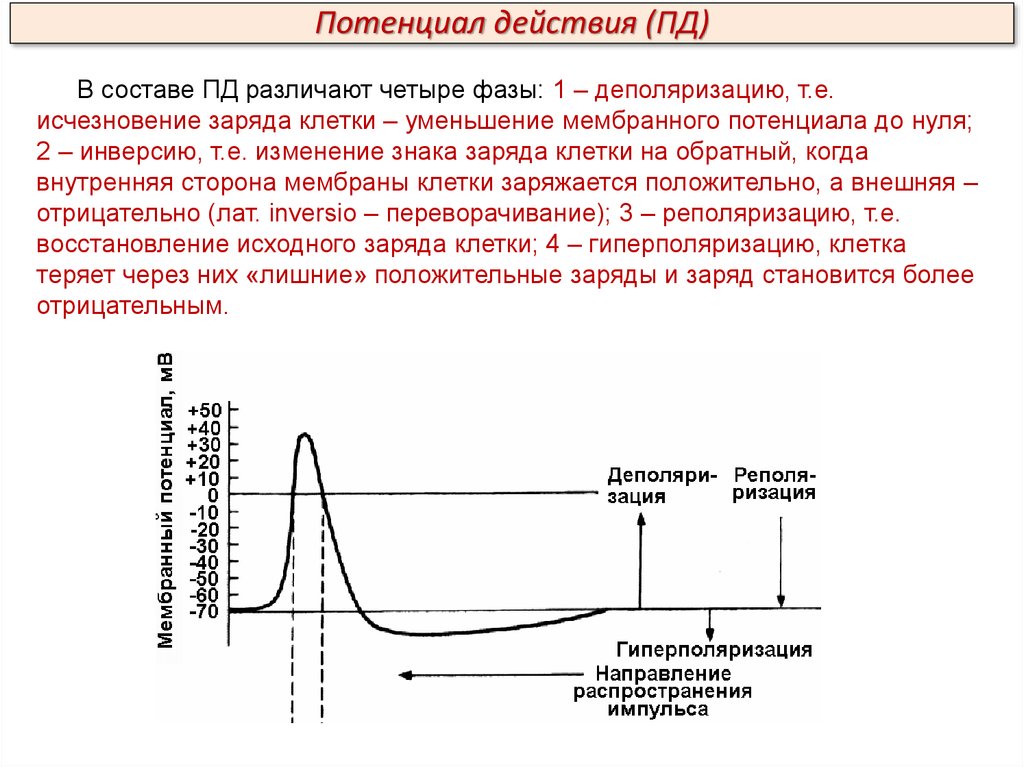
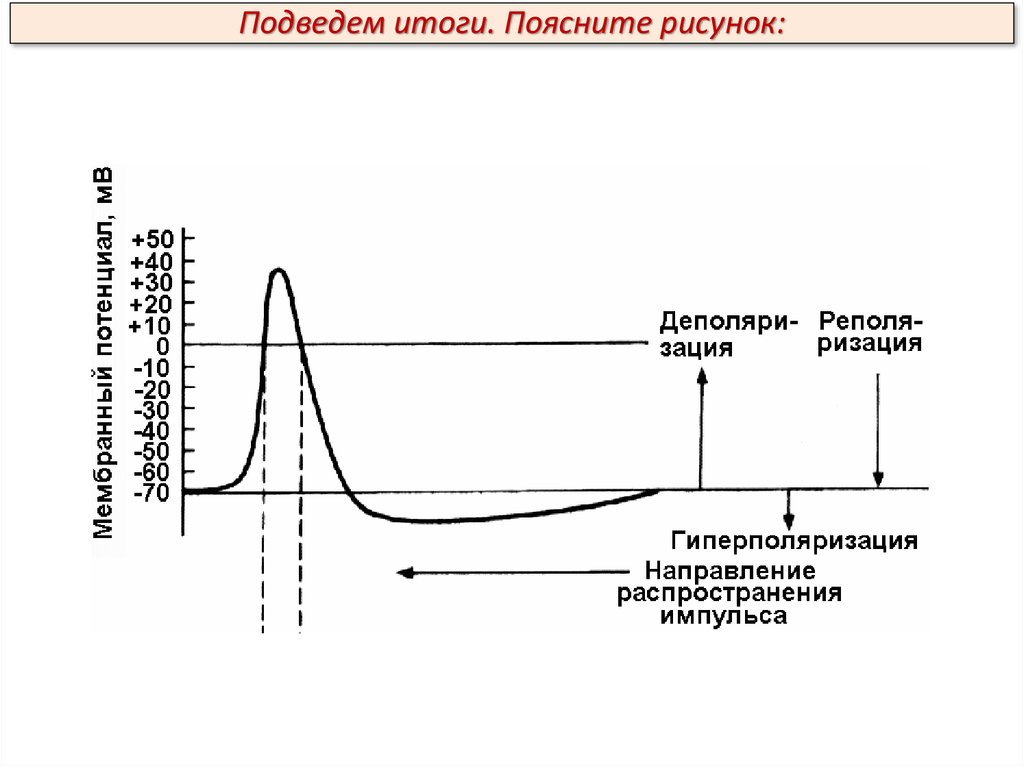
Потенциал действия (ПД)В составе ПД различают четыре фазы: 1 – деполяризацию, т.е.
исчезновение заряда клетки – уменьшение мембранного потенциала до нуля;
2 – инверсию, т.е. изменение знака заряда клетки на обратный, когда
внутренняя сторона мембраны клетки заряжается положительно, а внешняя –
отрицательно (лат. inversio – переворачивание); 3 – реполяризацию, т.е.
восстановление исходного заряда клетки; 4 – гиперполяризацию, клетка
теряет через них «лишние» положительные заряды и заряд становится более
отрицательным.
22.
Потенциал действия (ПД)Процесс развития ПД вызывают фазовые изменения проницаемости
клеточной мембраны, что обеспечивает быстрое движение Na+ в клетку. Это
наиболее часто встречаемый вариант возникновения ПД. Величина
мембранного потенциала при этом сначала уменьшается, а затем снова
восстанавливается до исходного уровня.
23.
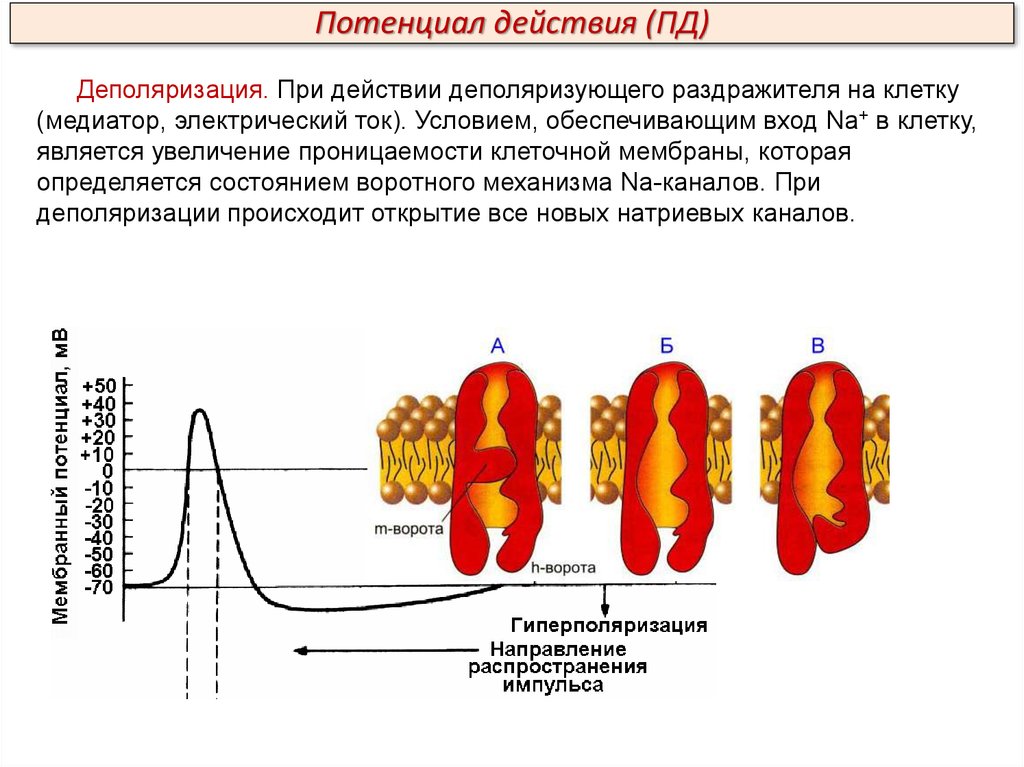
Потенциал действия (ПД)Деполяризация. При действии деполяризующего раздражителя на клетку
(медиатор, электрический ток). Условием, обеспечивающим вход Na+ в клетку,
является увеличение проницаемости клеточной мембраны, которая
определяется состоянием воротного механизма Na-каналов. При
деполяризации происходит открытие все новых натриевых каналов.
24.
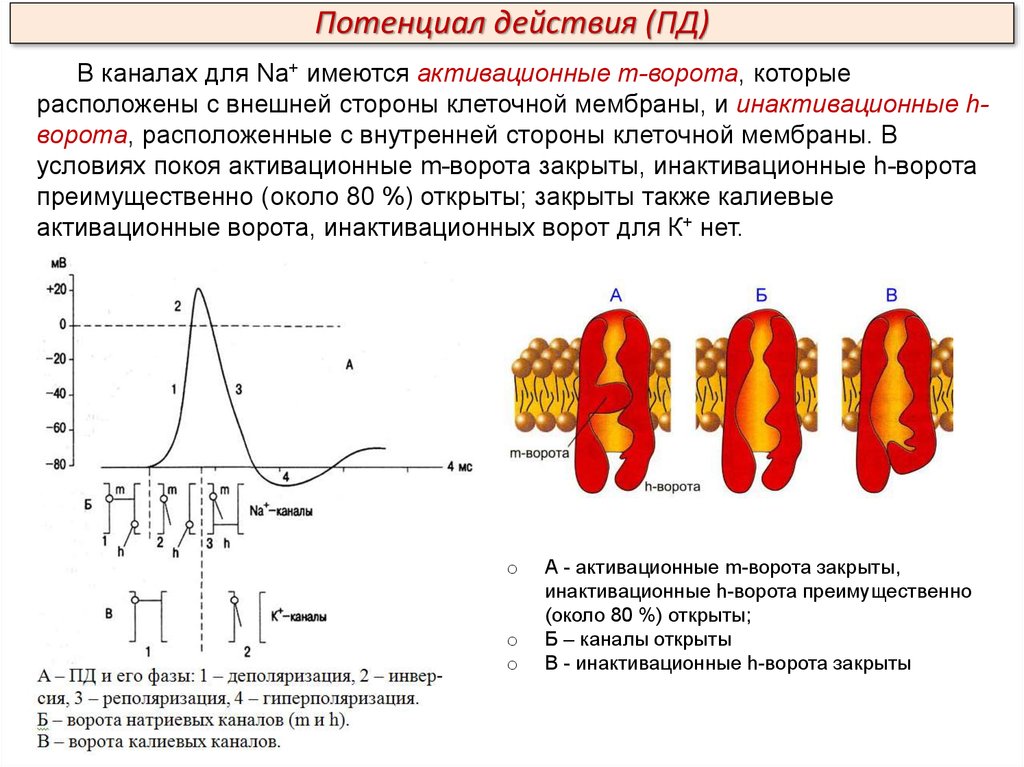
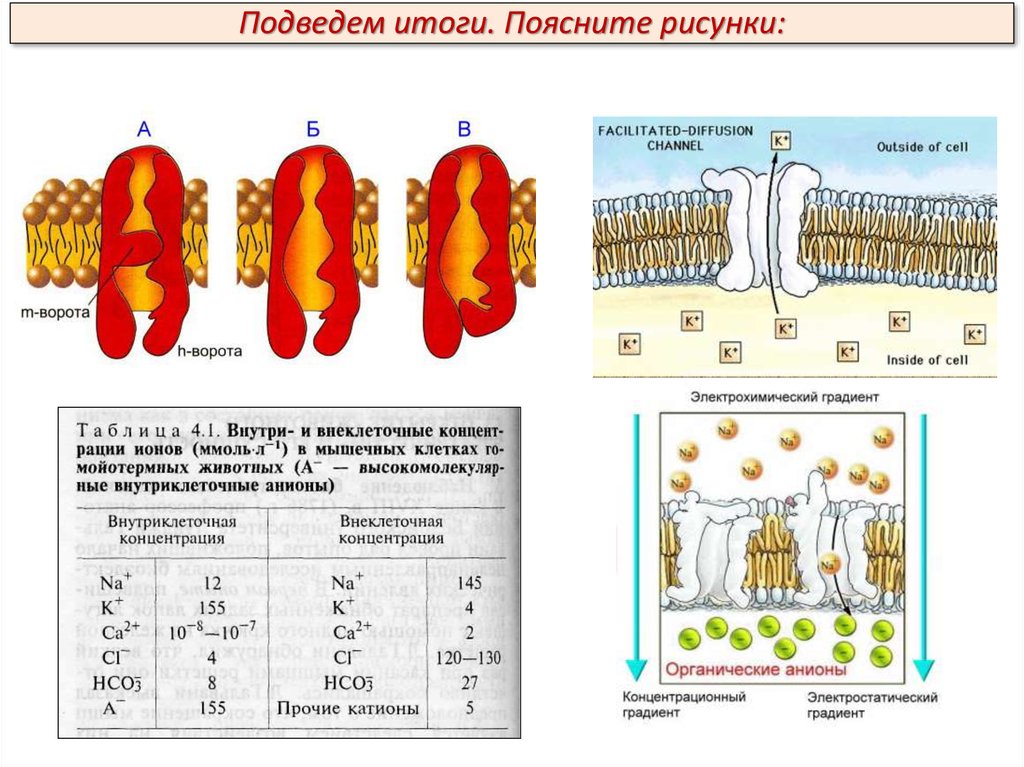
Потенциал действия (ПД)В каналах для Na+ имеются активационные m-ворота, которые
расположены с внешней стороны клеточной мембраны, и инактивационные hворота, расположенные с внутренней стороны клеточной мембраны. В
условиях покоя активационные m-ворота закрыты, инактивационные h-ворота
преимущественно (около 80 %) открыты; закрыты также калиевые
активационные ворота, инактивационных ворот для К+ нет.
o
o
o
А - активационные m-ворота закрыты,
инактивационные h-ворота преимущественно
(около 80 %) открыты;
Б – каналы открыты
В - инактивационные h-ворота закрыты
25.
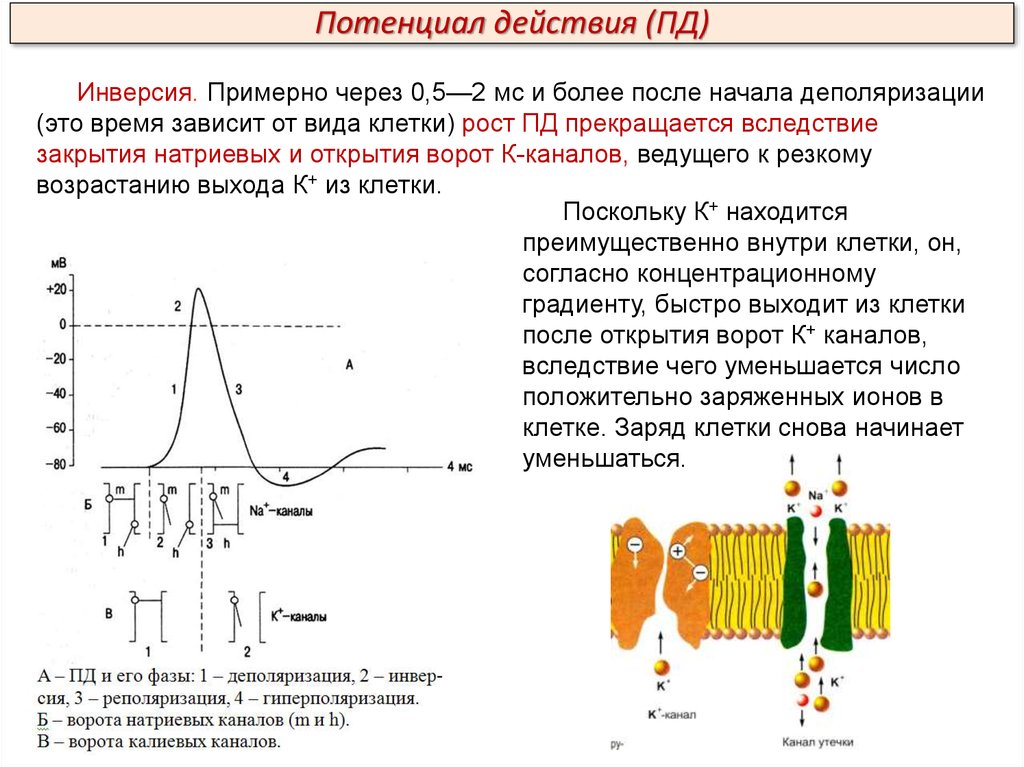
Потенциал действия (ПД)Инверсия. Примерно через 0,5—2 мс и более после начала деполяризации
(это время зависит от вида клетки) рост ПД прекращается вследствие
закрытия натриевых и открытия ворот К-каналов, ведущего к резкому
возрастанию выхода К+ из клетки.
Поскольку К+ находится
преимущественно внутри клетки, он,
согласно концентрационному
градиенту, быстро выходит из клетки
после открытия ворот К+ каналов,
вследствие чего уменьшается число
положительно заряженных ионов в
клетке. Заряд клетки снова начинает
уменьшаться.
26.
Потенциал действия (ПД)Выходу К+ из клетки способствует также и электрический градиент. К+
выталкивается положительным зарядом из клетки и притягивается
отрицательным зарядом снаружи клетки. Так продолжается до полного
исчезновения положительного заряда внутри клетки (до конца нисходящей
части фазы инверсии. Калий выходит из клетки не только по управляемым
каналам, ворота которых открыты, но и по неуправляемым – каналам утечки.
27.
Потенциал действия (ПД)Фаза реполяризации связана с тем, что проницаемость клеточной
мембраны для К+ все еще высока, ворота калиевых каналов открыты, К+
продолжает быстро выходить из клетки. Поскольку клетка теперь уже снова
внутри имеет отрицательный заряд, а снаружи – положительный,
электрический градиент препятствует выходу К+ из клетки, что снижает его
проводимость, хотя он продолжает выходить. Это объясняется тем, что
действие концентрационного градиента выражено значительно сильнее
электрического градиента. Таким образом, вся нисходящая часть пика ПД
обусловлена выходом К+ из клетки.
28.
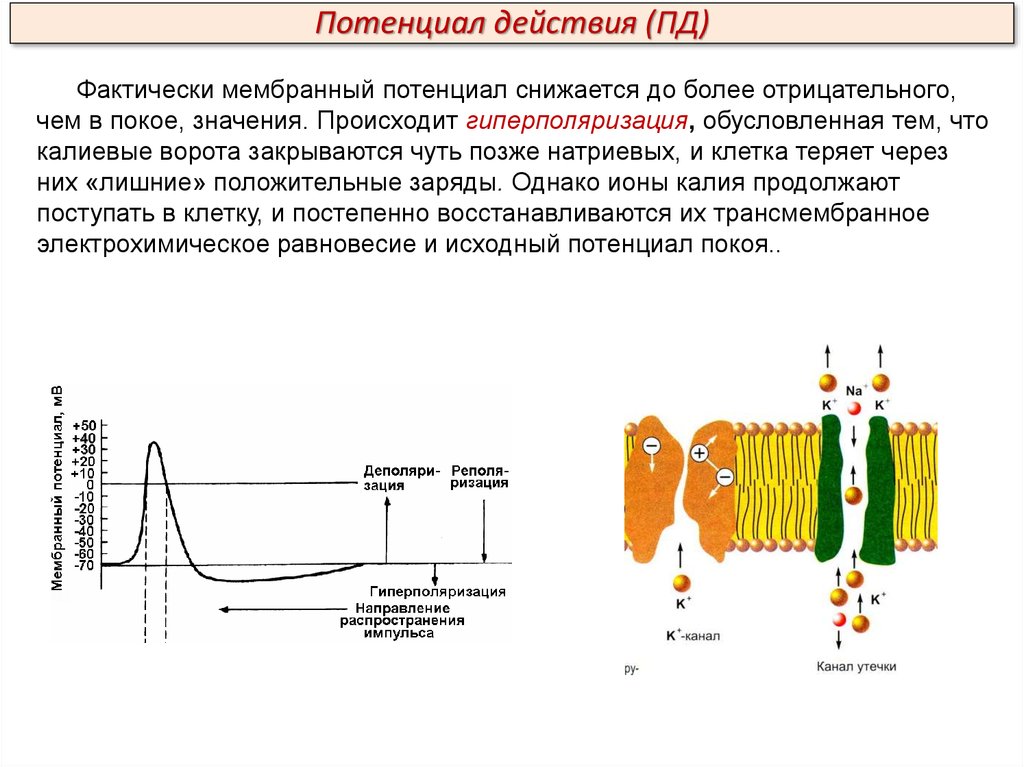
Потенциал действия (ПД)Фактически мембранный потенциал снижается до более отрицательного,
чем в покое, значения. Происходит гиперполяризация, обусловленная тем, что
калиевые ворота закрываются чуть позже натриевых, и клетка теряет через
них «лишние» положительные заряды. Однако ионы калия продолжают
поступать в клетку, и постепенно восстанавливаются их трансмембранное
электрохимическое равновесие и исходный потенциал покоя..
29.
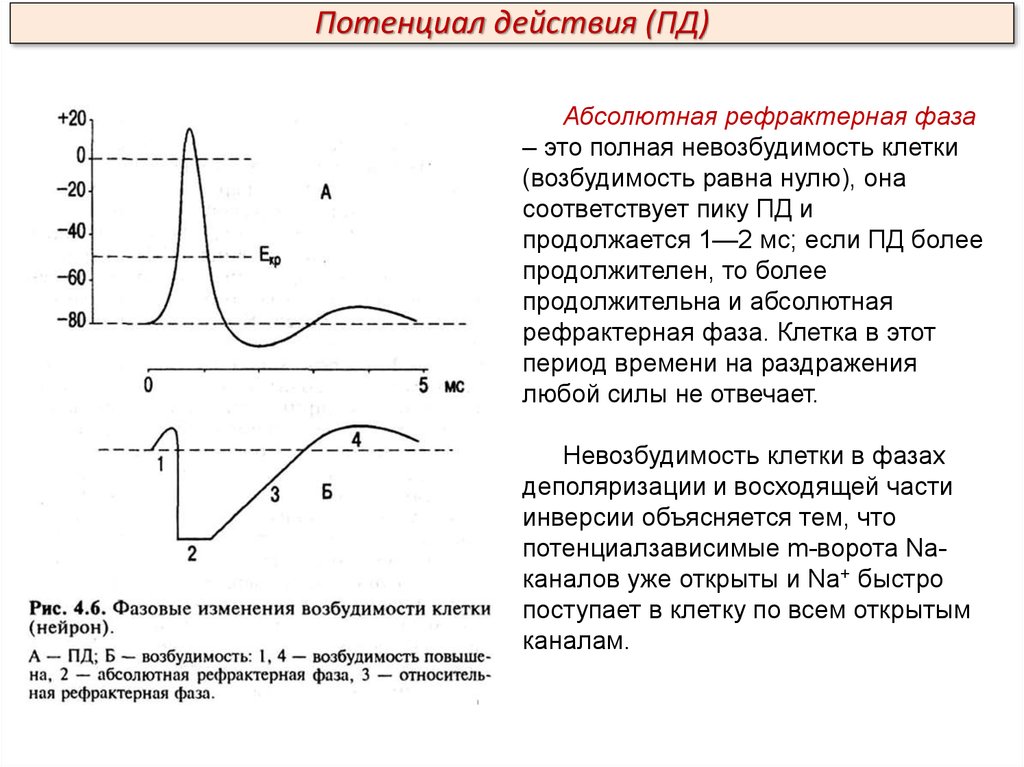
Потенциал действия (ПД)Абсолютная рефрактерная фаза
– это полная невозбудимость клетки
(возбудимость равна нулю), она
соответствует пику ПД и
продолжается 1—2 мс; если ПД более
продолжителен, то более
продолжительна и абсолютная
рефрактерная фаза. Клетка в этот
период времени на раздражения
любой силы не отвечает.
Невозбудимость клетки в фазах
деполяризации и восходящей части
инверсии объясняется тем, что
потенциалзависимые m-ворота Naканалов уже открыты и Na+ быстро
поступает в клетку по всем открытым
каналам.
30.
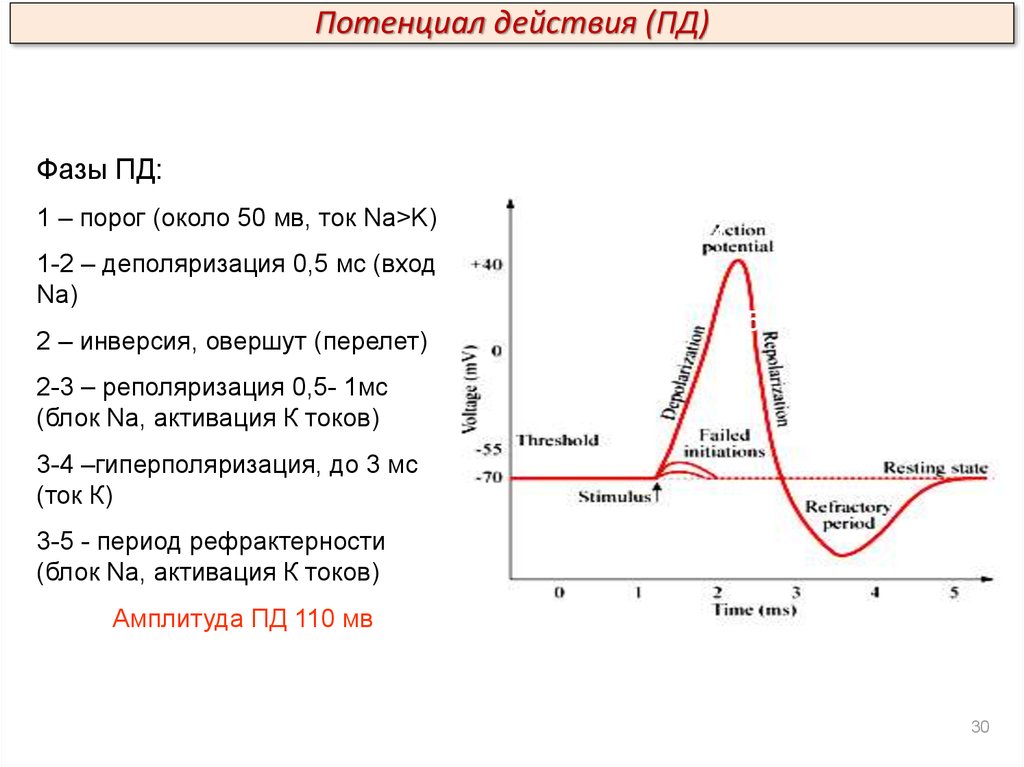
Потенциал действия (ПД)Фазы ПД:
1 – порог (около 50 мв, ток Na>K)
3
2
4
1-2 – деполяризация 0,5 мс (вход
Na)
2 – инверсия, овершут (перелет)
5
1
2-3 – реполяризация 0,5- 1мс
(блок Na, активация К токов)
3-4 –гиперполяризация, до 3 мс
(ток К)
3-5 - период рефрактерности
(блок Na, активация К токов)
Амплитуда ПД 110 мв
30
31.
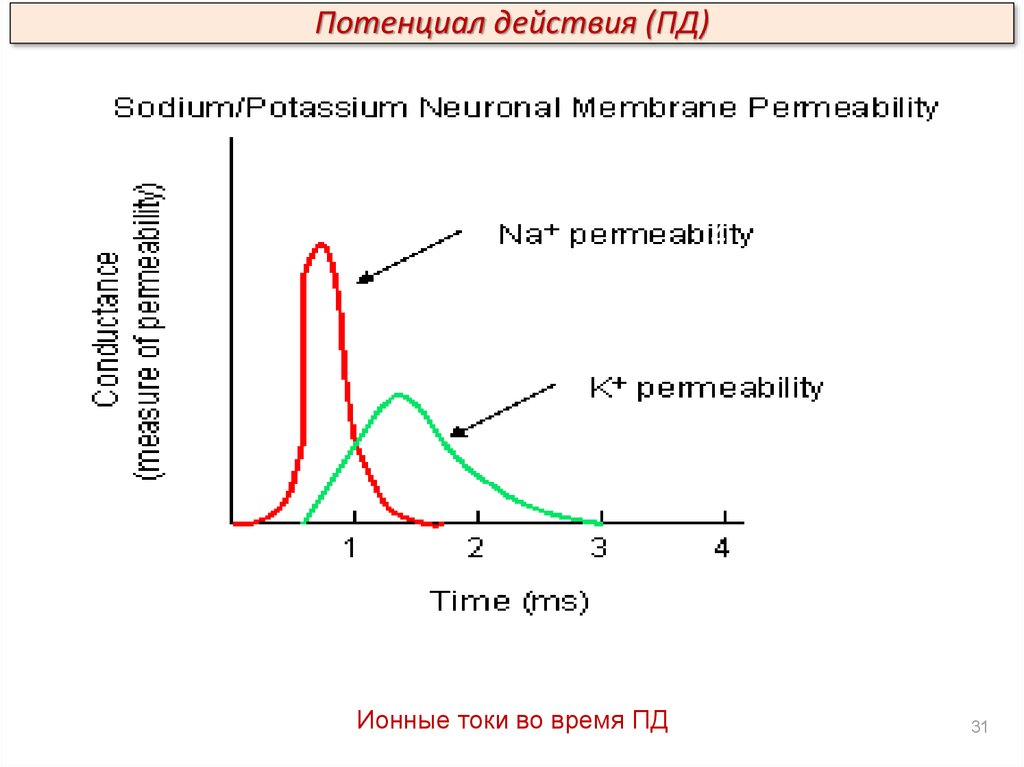
Потенциал действия (ПД)3
2
4
5
1
Ионные токи во время ПД
31
32.
33.
Подведем итоги:Какому закону подчиняется потенциал действия?
ПД подчиняется закону «все или ничего». ПД либо совсем не возникает при
раздражении клетки, если оно мало, либо достигает максимальной
величины, если раздражение является пороговым или сверхпороговым.
За счет чего происходит деполяризация мембраны во время ПД?
Потенциал действия возникает в результате внезапного
кратковременного открывания натриевых ворот. Это происходит в ответ
на сигнал, приводящий к пороговой деполяризации мембраны аксона.
За счет чего происходит реполяризация мембраны?
Вскоре после открывания натриевых ворот деполяризация мембраны
заставляет открываться и калиевые ворота. В результате усиливается
диффузия ионов калия из клетки, а поскольку у них такой же заряд, как у
ионов натрия, их выход делает внутреннюю среду аксона менее
положительной – начинается процесс реполяризации.
Чем обусловлена гиперполяризация мембраны?
Фактически мембранный потенциал снижается до более отрицательного,
чем в покое, значения. Происходит гиперполяризация, обусловленная тем,
что калиевые ворота закрываются чуть позже натриевых, и клетка
теряет через них «лишние» положительные заряды.
Какие ионы, в основном, обеспечивает потенциал действия (ПД)?
Ионы натрия.
34.
Подведем итоги:Что происходит с внешними (m-воротами) и
внутренними (h-воротами) натриевых
каналов во время покоя и при
деполяризации?
В условиях покоя активационные m-ворота
закрыты, инактивационные h-ворота
преимущественно (около 80 %) открыты;
при деполяризации открываются mворота.
Что происходит с внешними (m-воротами) и
внутренними (h-воротами) натриевых
каналов во время инверсии и
реполяризации?
h-ворота закрываются в фазе инверсии,
когда заряд внутри клетки становится
положительным, что и является причиной
их закрытия.
Что происходит с воротами калиевых каналов во время деполяризации и
реполяризации?
Во время деполяризации К-каналы закрыты, во время реполяризации
открываются, что ведет к резкому возрастанию выхода К+ из клетки.
35.
Подведем итоги. Что обозначено цифрами 1-4:1. Возбудимость повышена.
2. Абсолютная рефрактерная
фаза.
3. Относительная
рефрактерная фаза.
4. Возбудимость повышена.
36.
Проведение нервного импульсаВ немиелинизированных аксонах, типичных
для беспозвоночных, скорость распространения
потенциалов действия зависит от сопротивления
аксоплазмы. Это сопротивление в свою очередь
зависит от диаметра аксона – чем меньше
диаметр, тем больше сопротивление. В тонких
аксонах (<0,1 мм) высокое сопротивление
аксоплазмы влияет на проведение тока и
снижает длину местных цепей, так что в них
включаются только те участки, которые
расположены непосредственно впереди
потенциала действия. В результате скорость
распространения импульсов в этих аксонах низка
– всего около 0,5 м/с. Диаметр гигантских
аксонов, свойственных многим кольчатым
червям, членистоногим и моллюскам, равен
примерно 1 мм, а скорость проведения по ним
импульсов достигает 100 м/с. Этого вполне
достаточно для передачи жизненно важной
информации.
36
37.
Проведение нервного импульсаУ позвоночных подавляющая часть
нервных волокон, особенно в
спинномозговых и черепных нервах,
окружены миелиновой оболочкой,
образованной шванновскими
клетками. Миелин – это материал
белково-липидной природы,
обладающий высоким электрическим
сопротивлением и действующий как
изолятор, подобно резиновому или
пластиковому покрытию
электрического провода.
Там, где в миелиновой оболочке имеются разрывы, называемые
перехватами Ранвье, замыкаются местные цепи, и именно здесь через
мембрану аксона проходит ток, генерирующий следующий потенциал
действия. В результате импульс перескакивает от одного перехвата Ранвье к
другому и пробегает по миелинизированному аксону быстрее, чем серия
меньших по величине местных токов в немиелинизированном нервном
волокне. Такой способ распространения потенциала действия, называемый
сальтаторным (от лат. saltare – прыгать), может обеспечивать проведение
37
импульса со скоростью 120 м/с.
38.
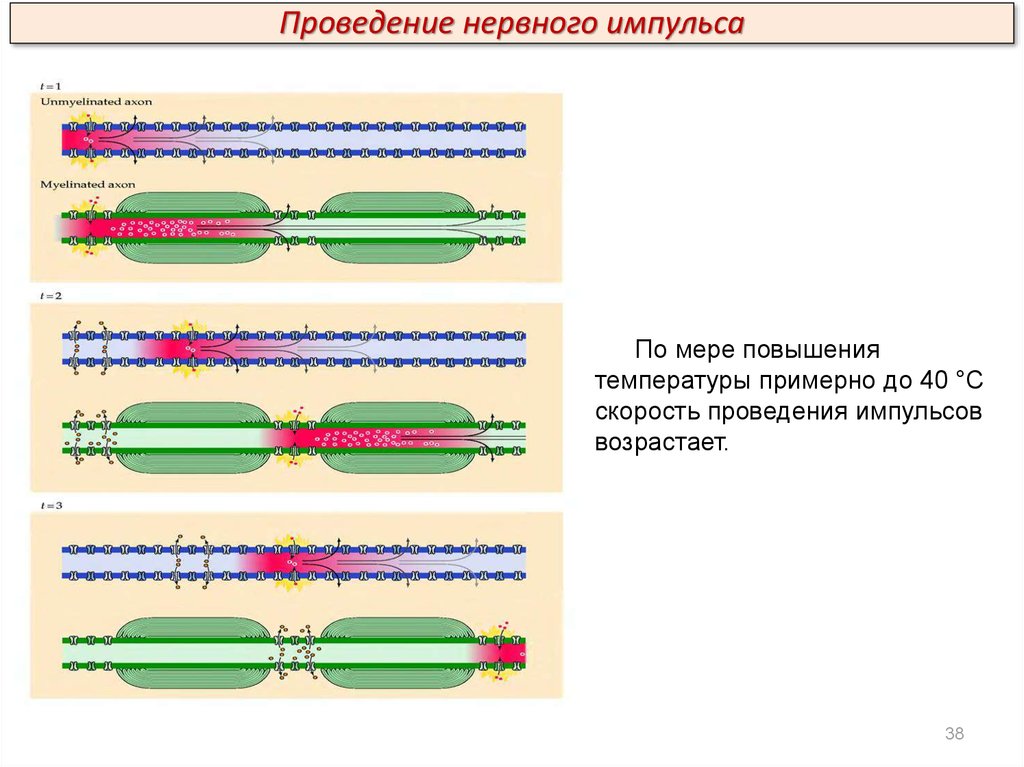
Проведение нервного импульсаПо мере повышения
температуры примерно до 40 °С
скорость проведения импульсов
возрастает.
38
39.
40.
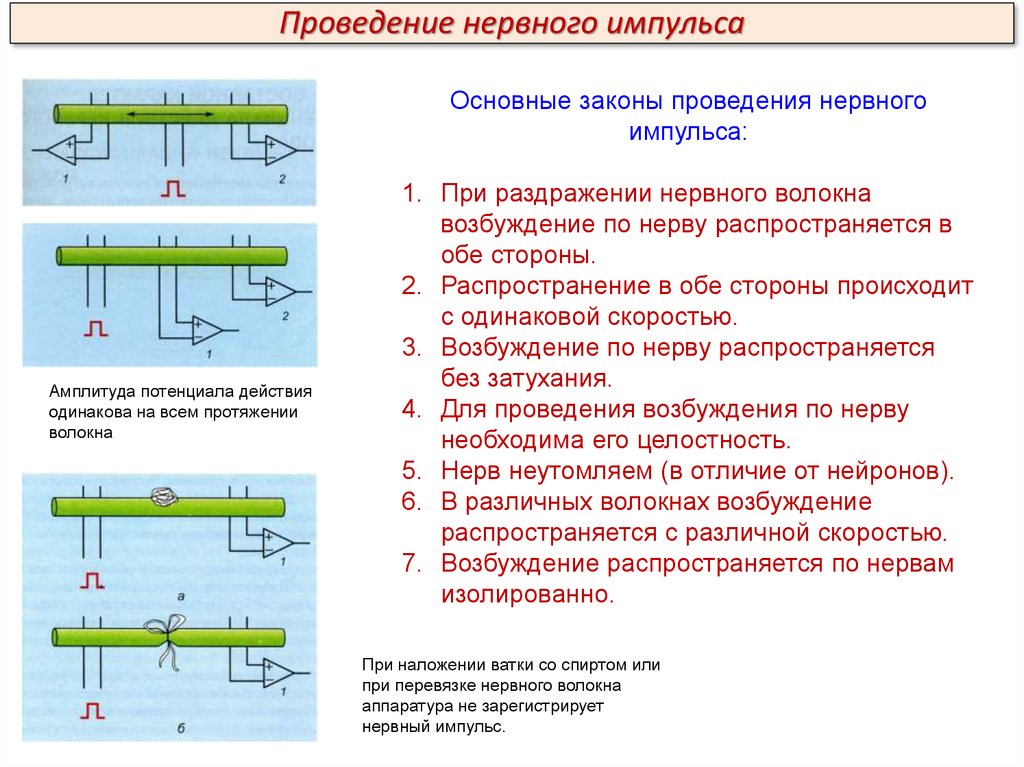
Проведение нервного импульсаОсновные законы проведения нервного
импульса:
Амплитуда потенциала действия
одинакова на всем протяжении
волокна
1. При раздражении нервного волокна
возбуждение по нерву распространяется в
обе стороны.
2. Распространение в обе стороны происходит
с одинаковой скоростью.
3. Возбуждение по нерву распространяется
без затухания.
4. Для проведения возбуждения по нерву
необходима его целостность.
5. Нерв неутомляем (в отличие от нейронов).
6. В различных волокнах возбуждение
распространяется с различной скоростью.
7. Возбуждение распространяется по нервам
изолированно.
При наложении ватки со спиртом или
при перевязке нервного волокна
аппаратура не зарегистрирует
нервный импульс.
41.
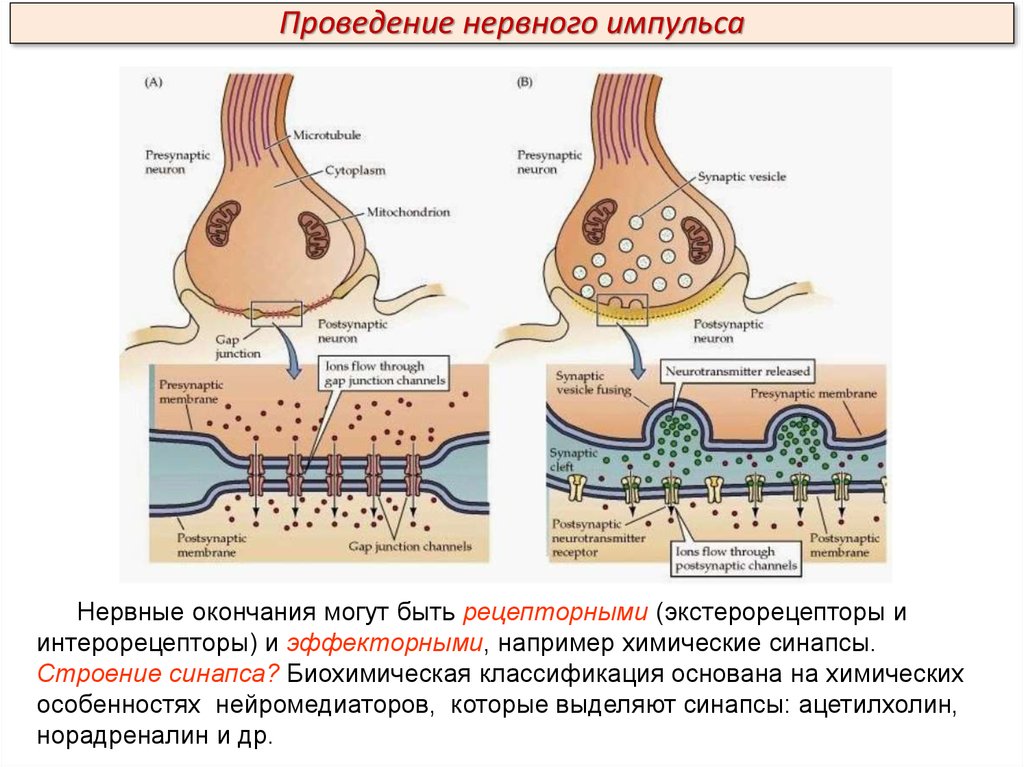
Проведение нервного импульсаНервные окончания могут быть рецепторными (экстерорецепторы и
интерорецепторы) и эффекторными, например химические синапсы.
Строение синапса? Биохимическая классификация основана на химических
особенностях нейромедиаторов, которые выделяют синапсы: ацетилхолин,
норадреналин и др.
42.
43.
44.
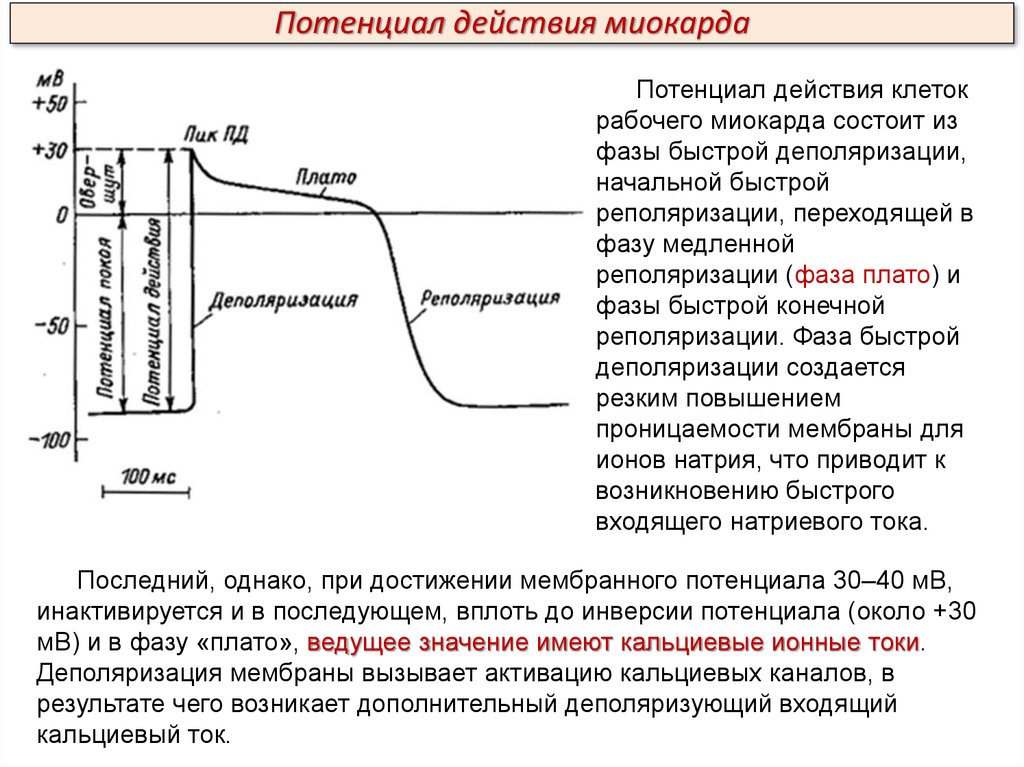
Потенциал действия миокардаПотенциал действия клеток
рабочего миокарда состоит из
фазы быстрой деполяризации,
начальной быстрой
реполяризации, переходящей в
фазу медленной
реполяризации (фаза плато) и
фазы быстрой конечной
реполяризации. Фаза быстрой
деполяризации создается
резким повышением
проницаемости мембраны для
ионов натрия, что приводит к
возникновению быстрого
входящего натриевого тока.
Последний, однако, при достижении мембранного потенциала 30–40 мВ,
инактивируется и в последующем, вплоть до инверсии потенциала (около +30
мВ) и в фазу «плато», ведущее значение имеют кальциевые ионные токи.
Деполяризация мембраны вызывает активацию кальциевых каналов, в
результате чего возникает дополнительный деполяризующий входящий
кальциевый ток.
45.
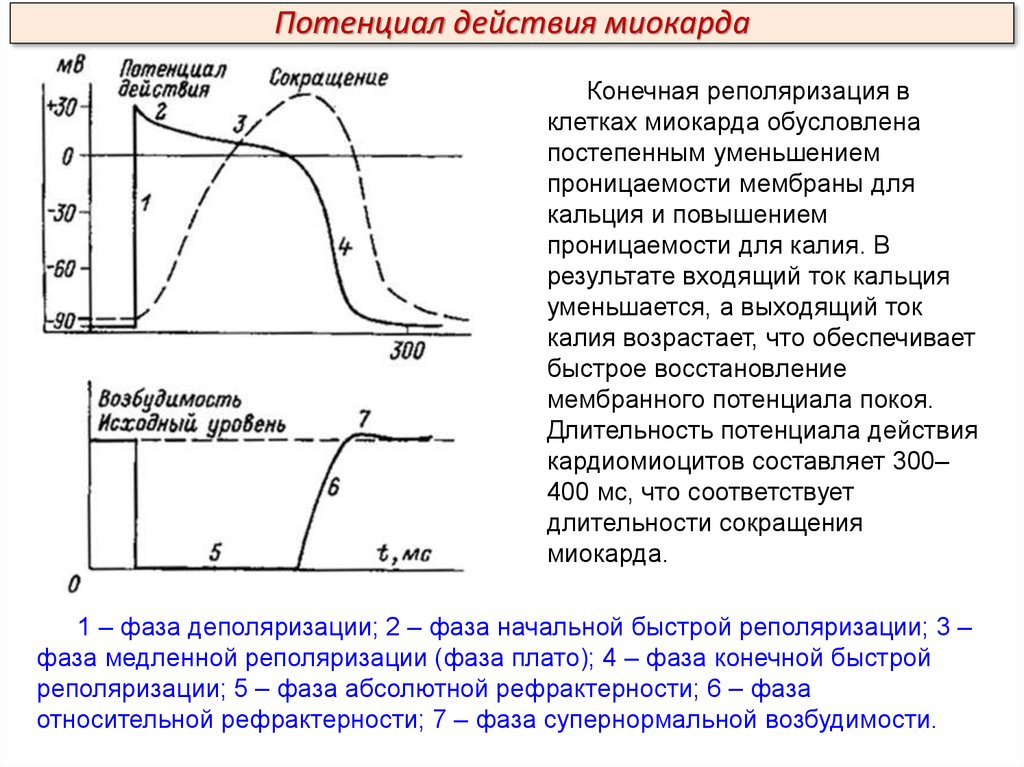
Потенциал действия миокардаКонечная реполяризация в
клетках миокарда обусловлена
постепенным уменьшением
проницаемости мембраны для
кальция и повышением
проницаемости для калия. В
результате входящий ток кальция
уменьшается, а выходящий ток
калия возрастает, что обеспечивает
быстрое восстановление
мембранного потенциала покоя.
Длительность потенциала действия
кардиомиоцитов составляет 300–
400 мс, что соответствует
длительности сокращения
миокарда.
1 – фаза деполяризации; 2 – фаза начальной быстрой реполяризации; 3 –
фаза медленной реполяризации (фаза плато); 4 – фаза конечной быстрой
реполяризации; 5 – фаза абсолютной рефрактерности; 6 – фаза
относительной рефрактерности; 7 – фаза супернормальной возбудимости.
46.
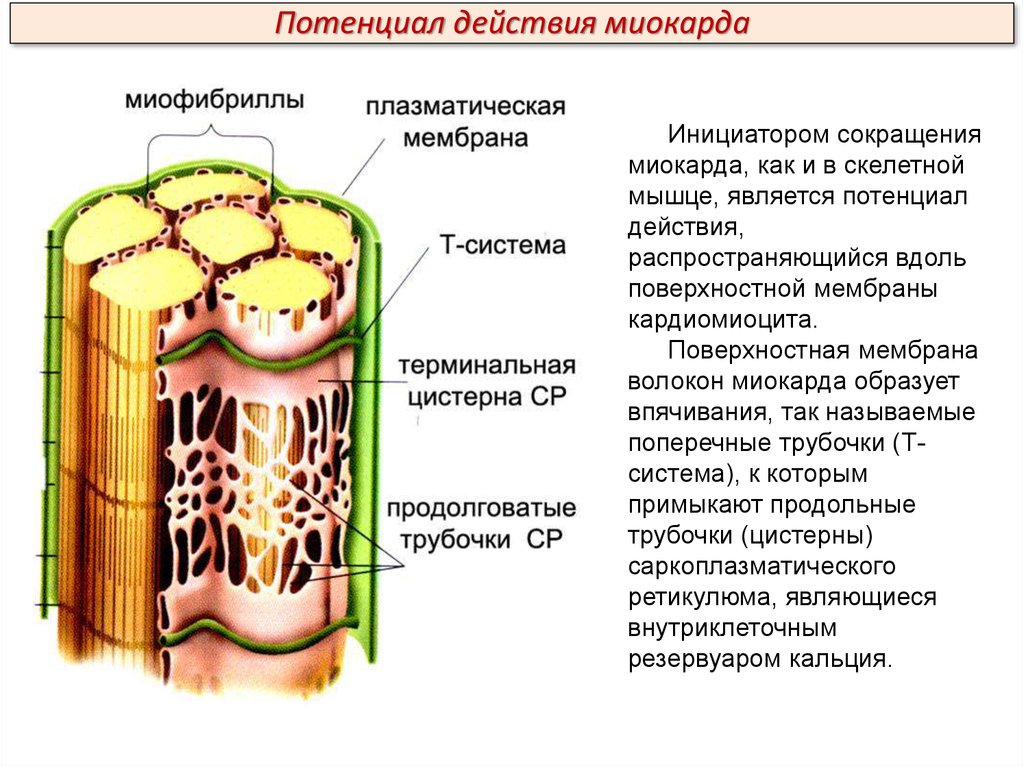
Потенциал действия миокардаИнициатором сокращения
миокарда, как и в скелетной
мышце, является потенциал
действия,
распространяющийся вдоль
поверхностной мембраны
кардиомиоцита.
Поверхностная мембрана
волокон миокарда образует
впячивания, так называемые
поперечные трубочки (Тсистема), к которым
примыкают продольные
трубочки (цистерны)
саркоплазматического
ретикулюма, являющиеся
внутриклеточным
резервуаром кальция.
47.
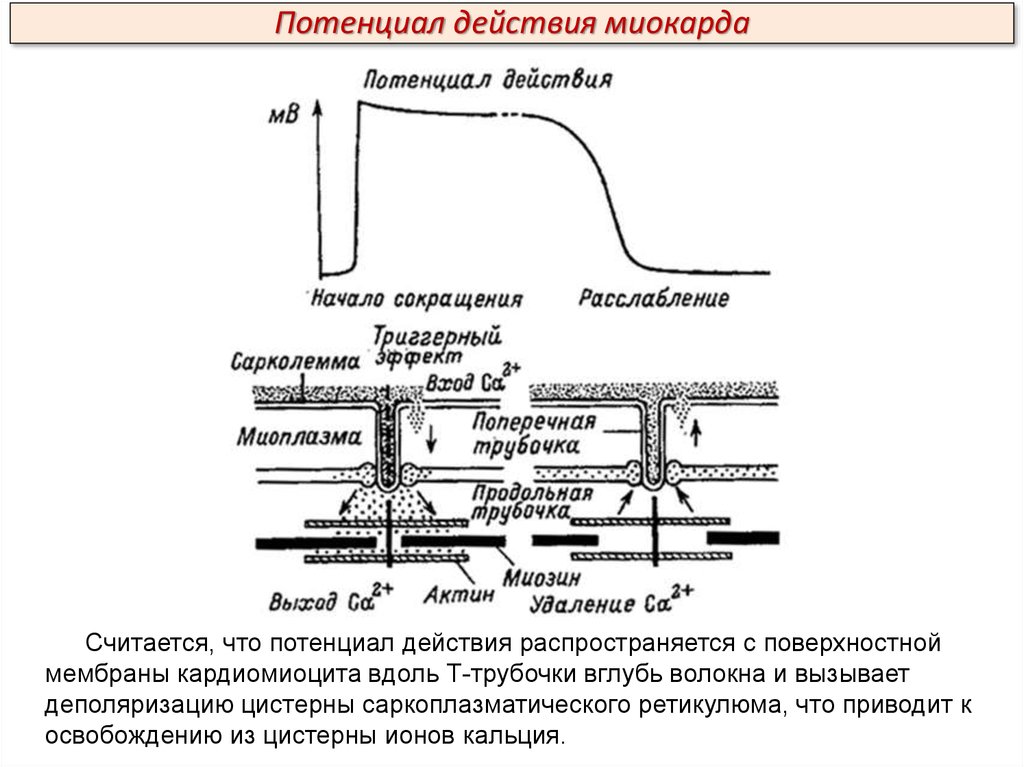
Потенциал действия миокардаСчитается, что потенциал действия распространяется с поверхностной
мембраны кардиомиоцита вдоль Т-трубочки вглубь волокна и вызывает
деполяризацию цистерны саркоплазматического ретикулюма, что приводит к
освобождению из цистерны ионов кальция.
48.
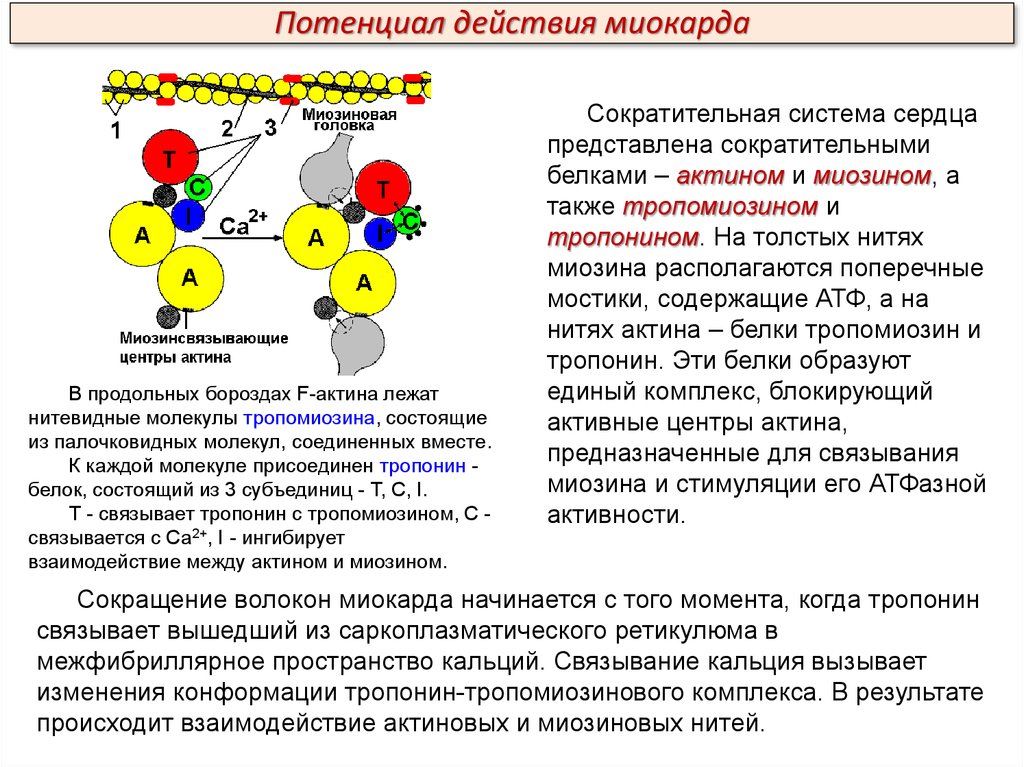
Потенциал действия миокардаВ продольных бороздах F-актина лежат
нитевидные молекулы тропомиозина, состоящие
из палочковидных молекул, соединенных вместе.
К каждой молекуле присоединен тропонин белок, состоящий из 3 субъединиц - Т, С, I.
Т - связывает тропонин с тропомиозином, С связывается с Са2+, I - ингибирует
взаимодействие между актином и миозином.
Сократительная система сердца
представлена сократительными
белками – актином и миозином, а
также тропомиозином и
тропонином. На толстых нитях
миозина располагаются поперечные
мостики, содержащие АТФ, а на
нитях актина – белки тропомиозин и
тропонин. Эти белки образуют
единый комплекс, блокирующий
активные центры актина,
предназначенные для связывания
миозина и стимуляции его АТФазной
активности.
Сокращение волокон миокарда начинается с того момента, когда тропонин
связывает вышедший из саркоплазматического ретикулюма в
межфибриллярное пространство кальций. Связывание кальция вызывает
изменения конформации тропонин-тропомиозинового комплекса. В результате
происходит взаимодействие актиновых и миозиновых нитей.
49.
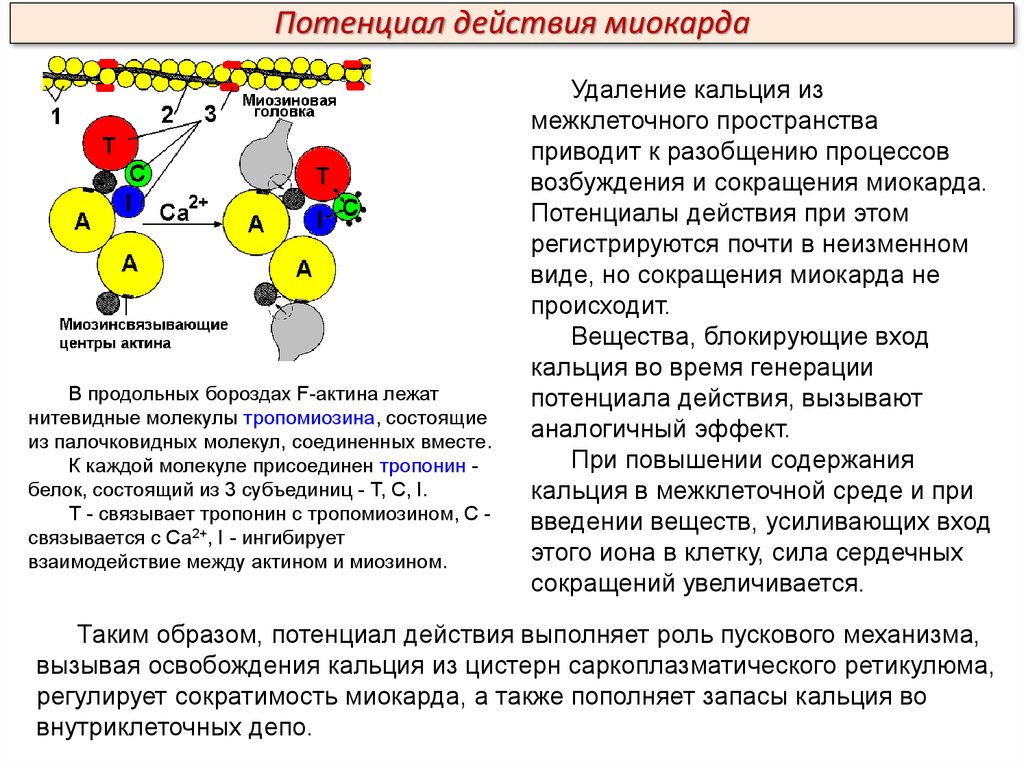
Потенциал действия миокардаВ продольных бороздах F-актина лежат
нитевидные молекулы тропомиозина, состоящие
из палочковидных молекул, соединенных вместе.
К каждой молекуле присоединен тропонин белок, состоящий из 3 субъединиц - Т, С, I.
Т - связывает тропонин с тропомиозином, С связывается с Са2+, I - ингибирует
взаимодействие между актином и миозином.
Удаление кальция из
межклеточного пространства
приводит к разобщению процессов
возбуждения и сокращения миокарда.
Потенциалы действия при этом
регистрируются почти в неизменном
виде, но сокращения миокарда не
происходит.
Вещества, блокирующие вход
кальция во время генерации
потенциала действия, вызывают
аналогичный эффект.
При повышении содержания
кальция в межклеточной среде и при
введении веществ, усиливающих вход
этого иона в клетку, сила сердечных
сокращений увеличивается.
Таким образом, потенциал действия выполняет роль пускового механизма,
вызывая освобождения кальция из цистерн саркоплазматического ретикулюма,
регулирует сократимость миокарда, а также пополняет запасы кальция во
внутриклеточных депо.
50.
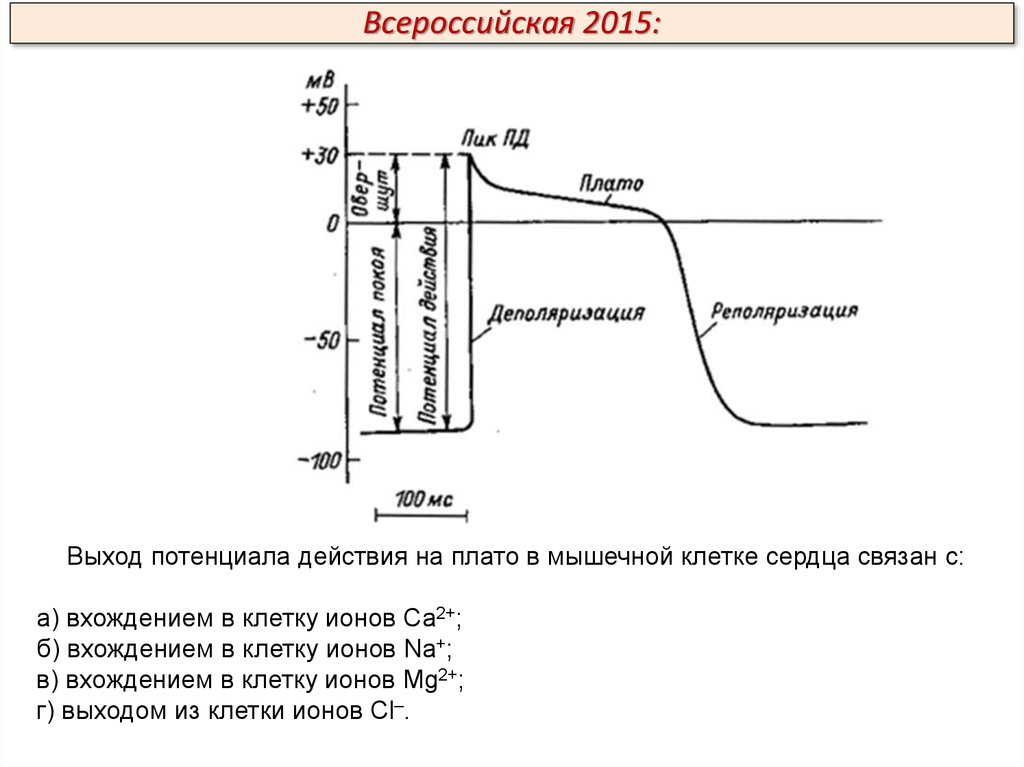
Всероссийская 2015:Выход потенциала действия на плато в мышечной клетке сердца связан с:
а) вхождением в клетку ионов Са2+;
б) вхождением в клетку ионов Na+;
в) вхождением в клетку ионов Mg2+;
г) выходом из клетки ионов Cl–.
51.
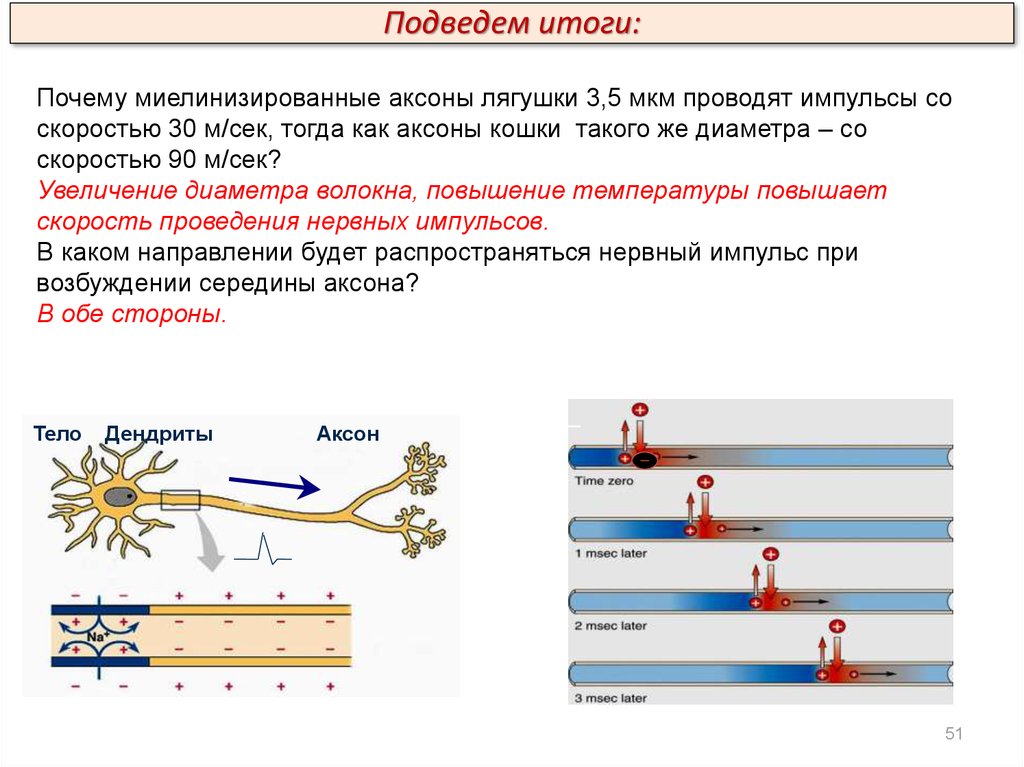
Подведем итоги:Почему миелинизированные аксоны лягушки 3,5 мкм проводят импульсы со
скоростью 30 м/сек, тогда как аксоны кошки такого же диаметра – со
скоростью 90 м/сек?
Увеличение диаметра волокна, повышение температуры повышает
скорость проведения нервных импульсов.
В каком направлении будет распространяться нервный импульс при
возбуждении середины аксона?
В обе стороны.
Тело
Дендриты
Аксон
51