
























 Химия
ХимияПохожие презентации:



")






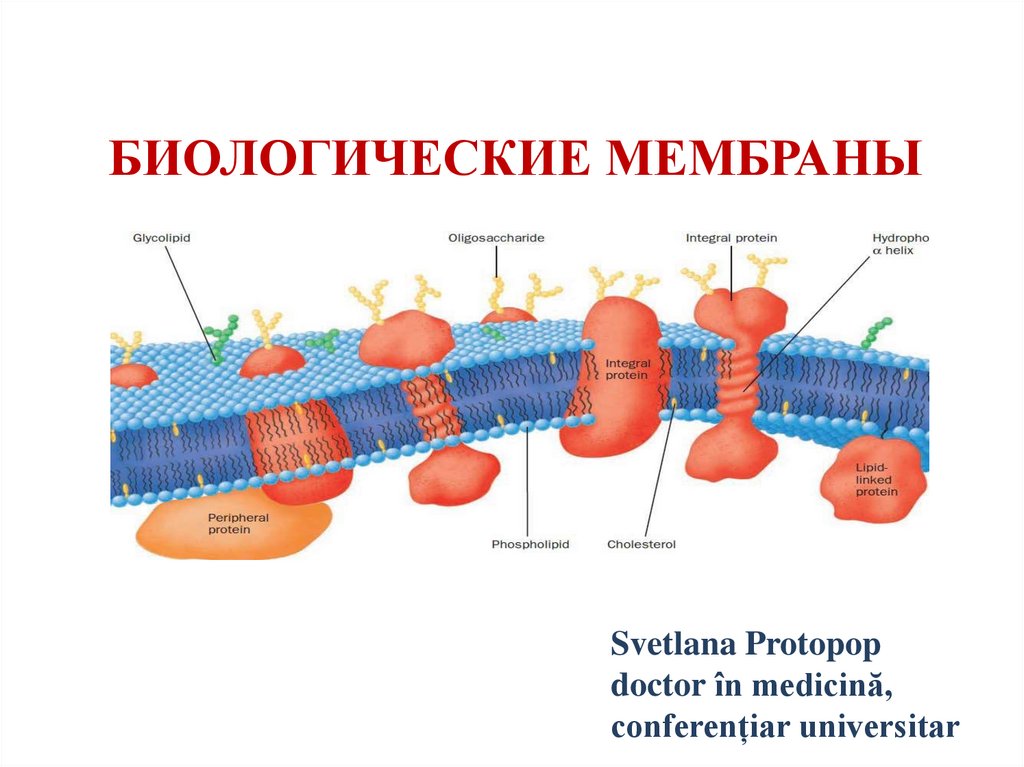
Биологические мембраны
1.
БИОЛОГИЧЕСКИЕ МЕМБРАНЫSvetlana Protopop
doctor în medicină,
conferențiar universitar
2.
Биологические мембраны• Сложные структуры, состоящие из липидов,
белков и углеводов.
• Мембраны изолируют одну клетку от другой и
обеспечивают их индивидуальность.
• Мембраны образуют также внутриклеточные
органеллы (митохондрии, ядро, лизосомы,
эндоплазматический и саркоплазматический
ретикулум, комплекс Гольджи, секреторные
гранулы.
3.
Функции биологических мембран• Ограничение и поддержание нормального состава
внутри- и внеклеточной жидкости.
• Ограничение и поддержание нормального состава
органелл (компартментализация).
• Перечисленные функции обеспечиваются избирательной
проницаемостью мембран для различных веществ.
• Метаболическая функция (в мембране протекают
химические реакции и процессы; пример – дыхательная
цепь).
• Преобразование энергии – окислительное
фосфорилирование и фотосинтез.
• Межклеточное взаимодействие.
• Рецепция и трансдукция внешних сигналов.
4.
Химический состав мембран• Липиды – 30-50%.
• Холестерол – 20% (Эритроциты – 25%).
• Белки – 30-50%.
• Углеводы – 5-15%.
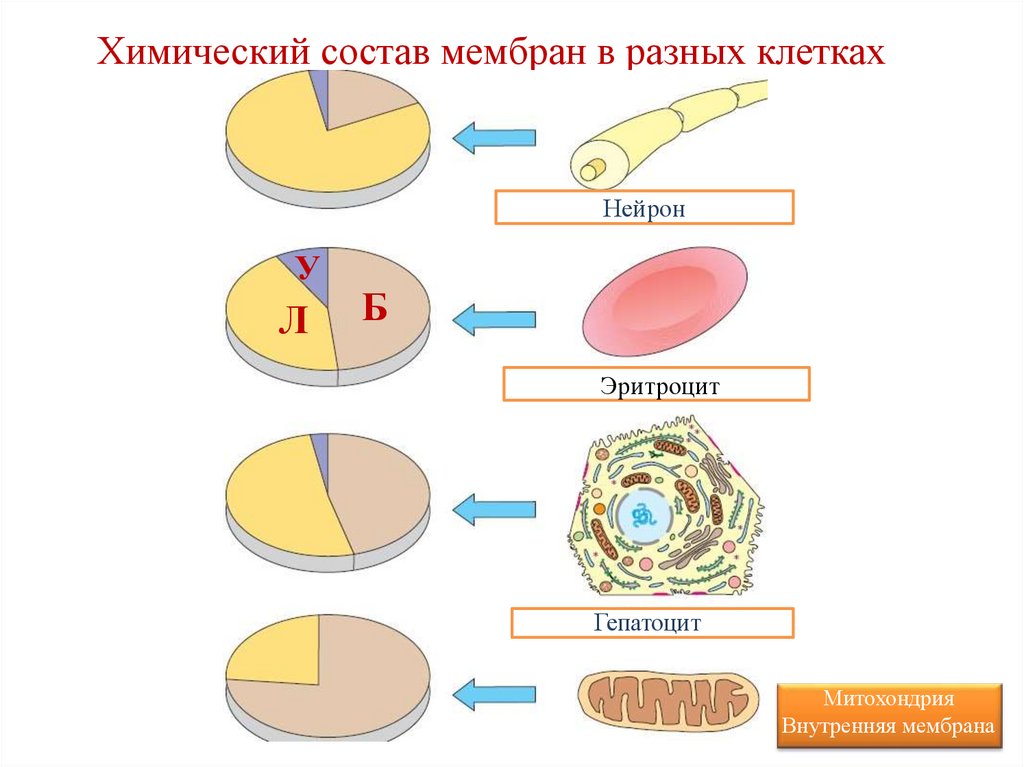
• Химический состав варьирует в
зависимости от типа мембраны.
5.
Химический состав мембран в разных клеткахНейрон
У
LЛ
БP
Эритроцит
Гепатоцит
Митохондрия
Внутренняя мембрана
6.
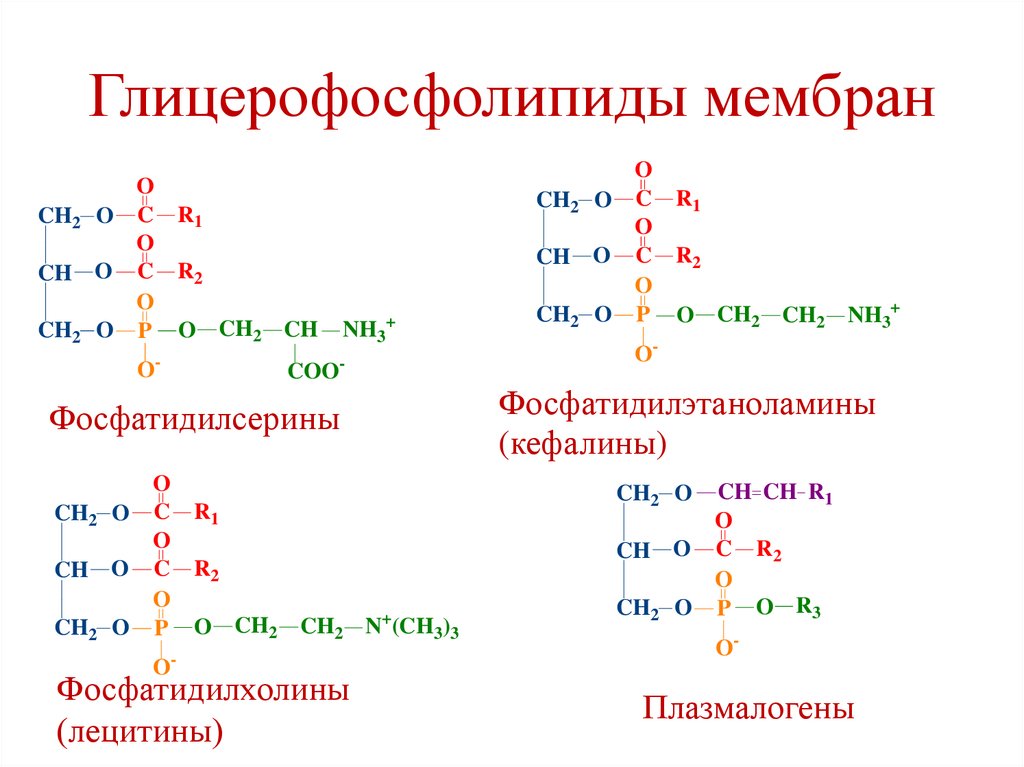
Липиды клеточных мембран• Фосфолипиды: Глицерофосфолипиды;
Сфингофосфолипиды.
• Гликолипиды:
• Холестерол.
Цереброзиды;
Ганглиозиды.
7.
Глицерофосфолипиды мембранФосфатидилхолины (лецитины)
Фосфатидилэтаноламины (кефалины)
Фосфатидилсерины
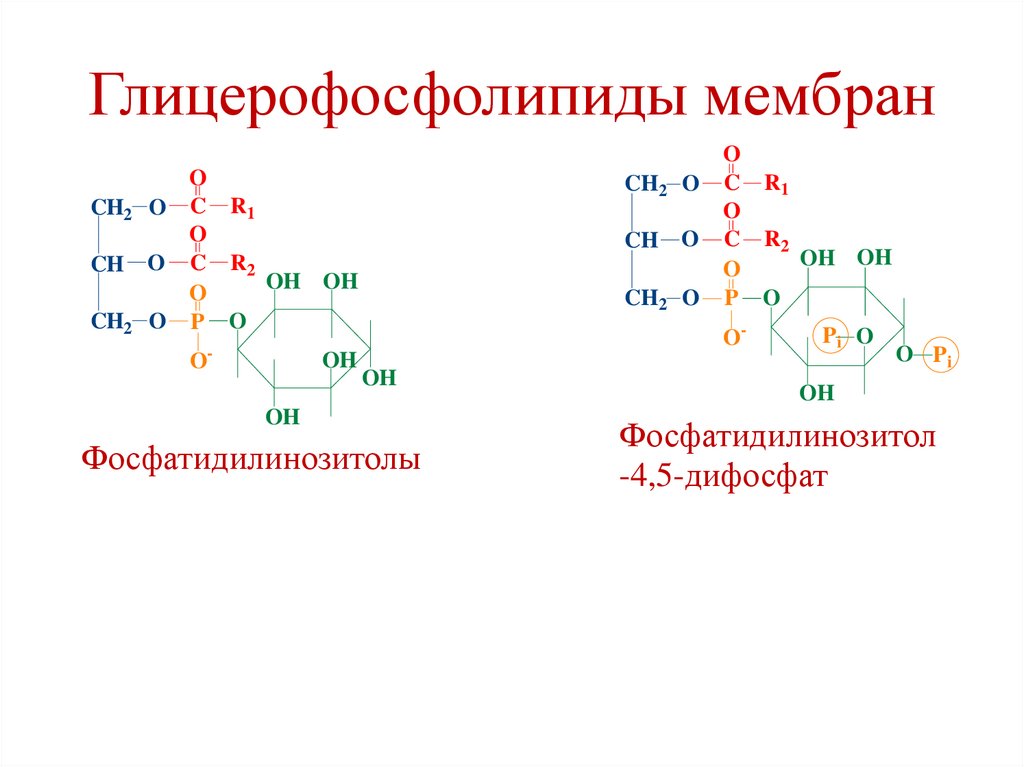
Фосфатидилинозитолы
Плазмалогены
Кардиолипины
8.
Функции глицерофосфолипидовмембран
• Образование билипидного слоя;
• Связывание с белками клеточных мембран;
• Предшественники вторичных посредников
гормонов;
• Предшественники арахидоновой кислоты для
синтеза простагландинов;
• Алостерические активаторы ферментов –
портеинкиназа С активна в комплексе с Са2+,
диацилглицеролом и 4-мя молекулами
фосфатидтлсерина.
9.
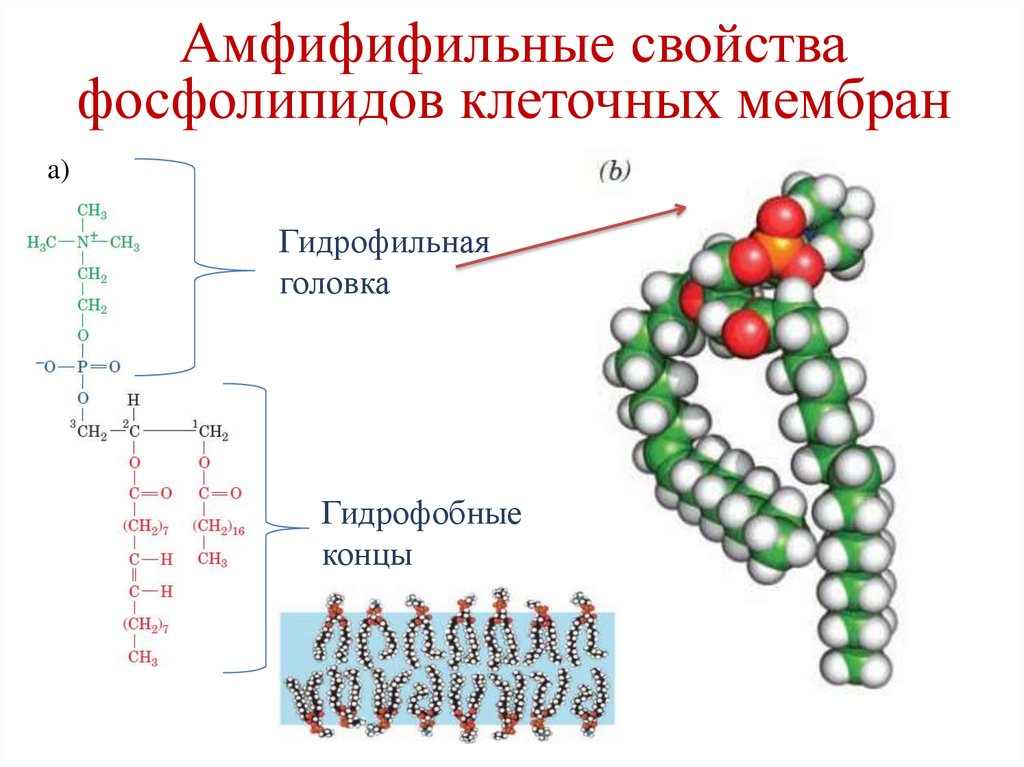
Амфифифильные свойствафосфолипидов клеточных мембран
a)
Гидрофильная
головка
Гидрофобные
концы
10.
Глицерофосфолипиды мембранCH2 O
CH O
CH2 O
O
C
O
C
O
P
CH2 O
R1
CH O
R2
NH3+
O CH2 CH
-
-
O
COO
Фосфатидилсерины
CH2 O
CH O
CH2 O
O
C
O
C
O
P
R2
O CH2 CH2 NH3+
O-
Фосфатидилэтаноламины
(кефалины)
CH O
R2
CH2 N+(CH3)3
-
O
R1
CH2 O
R1
O CH2
CH2 O
O
C
O
C
O
P
Фосфатидилхолины
(лецитины)
CH2 O
CH CH R1
O
C R2
O
P O R3
O-
Плазмалогены
11.
Глицерофосфолипиды мембранCH2 O
CH
O
CH2 O
O
C R1
O
C R2
OH OH
O
P O
-
OH
O
CH2 O
CH
O
CH2 O
O
C R1
O
C R2
OH OH
O
P O
O-
OH
OH
Фосфатидилинозитолы
Pi O
O Pi
OH
Фосфатидилинозитол
-4,5-дифосфат
12.
Сфингофосфолипиды мембранCH (CH2)12 CH3
O
NH C R
O
O P O CH2 CH2 N+(CH3)3
O-
HO CH CH
CH
CH2
Сфингомиелины
13.
Сфингофосфолипиды мембран• Сфингомиелины составляют 10-20% всех
мембранных липидов.
• Входят в состав миелиновых оболочек
нервных клеток.
• Являются предшественниками церамидов,
которые участвуют в трансдукции сигналов.
• Участвуют в апоптозе (церамиды).
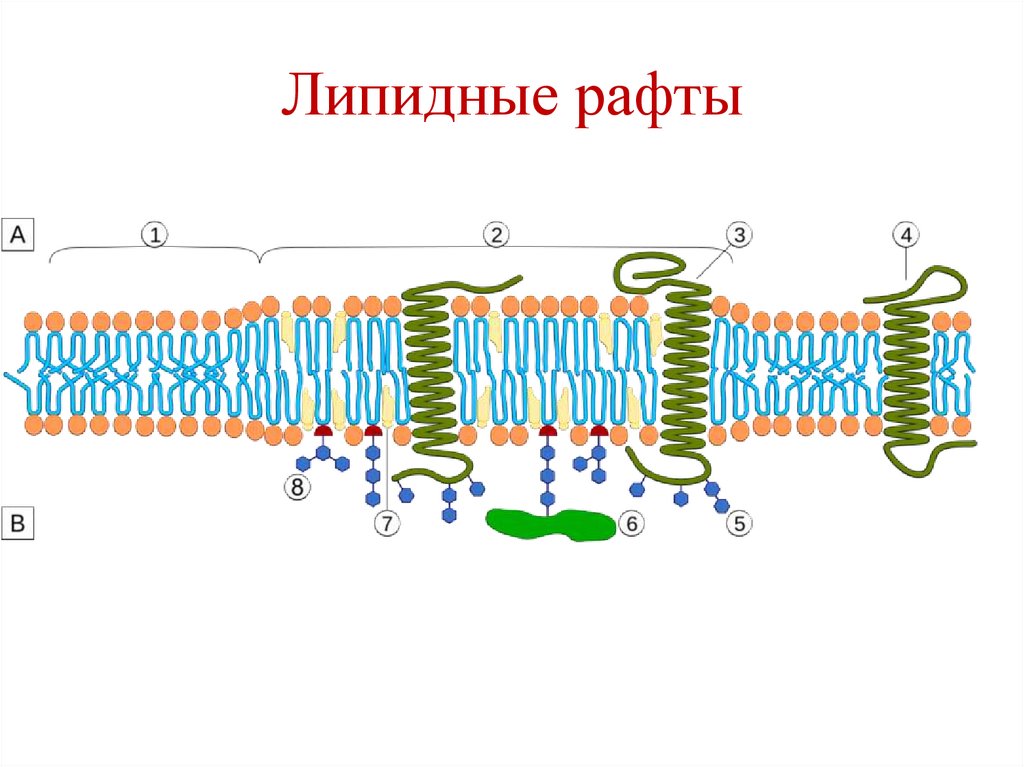
• Участвуют в образовании липидных рафтов.
14.
Липидные рафты15.
Гликолипиды мембранCH (CH2)12 CH3
O
NH C R
CH (CH2)12 CH3
O
NH C R
HO CH CH
HO CH CH
CH
CH
CH2 O
CH2OH
O
CH2 O
O
OH
CH2OH
OH
OH
OH
OH
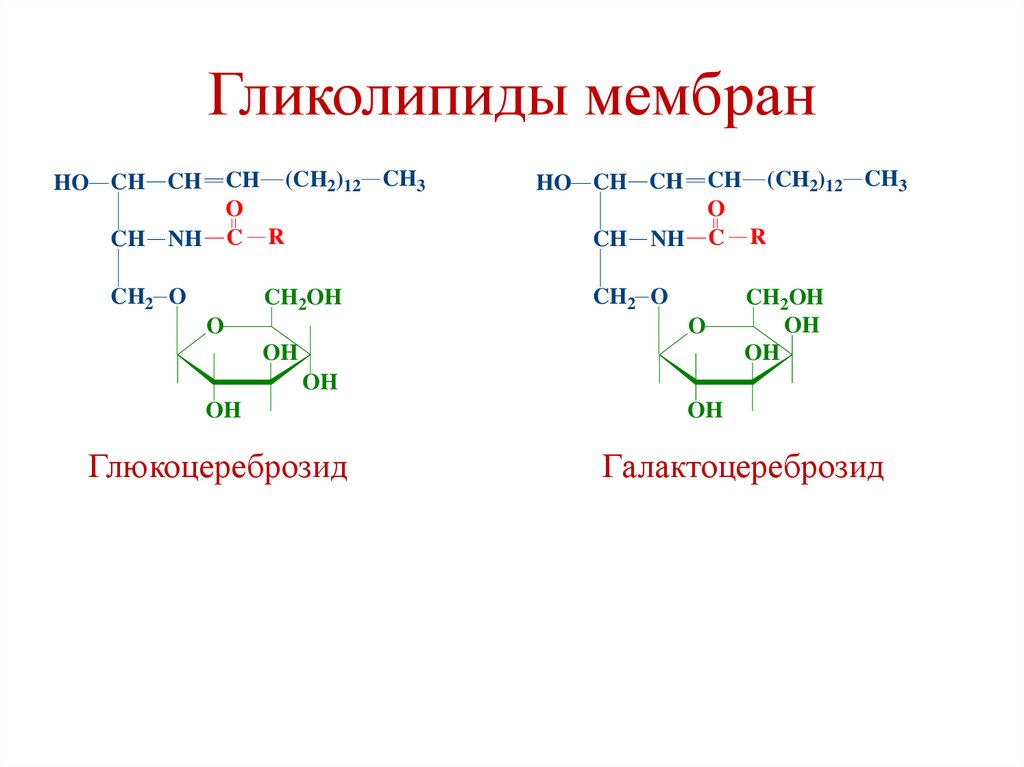
Глюкоцереброзид
OH
Галактоцереброзид
16.
Галактоцереброзиды• Основные компоненты клеточных мембран нервных
клеток, особенно головного мозга.
• 2% - серое вещество; 12% - белое вещество.
• Миелиновая оболочка; астроциты.
• Выполняют важную роль в образовании
липидных рафтов благодаря высокой
способности образовывать водородные связи.
• В комплексе с холестеролом образуют ситусы для
связывания рецепторов.
17.
Глюкоцереброзиды• Преобладают в мембранах эритроцитов,
селезенки, кожи, меньше – в мембранах
нервных клеток.
18.
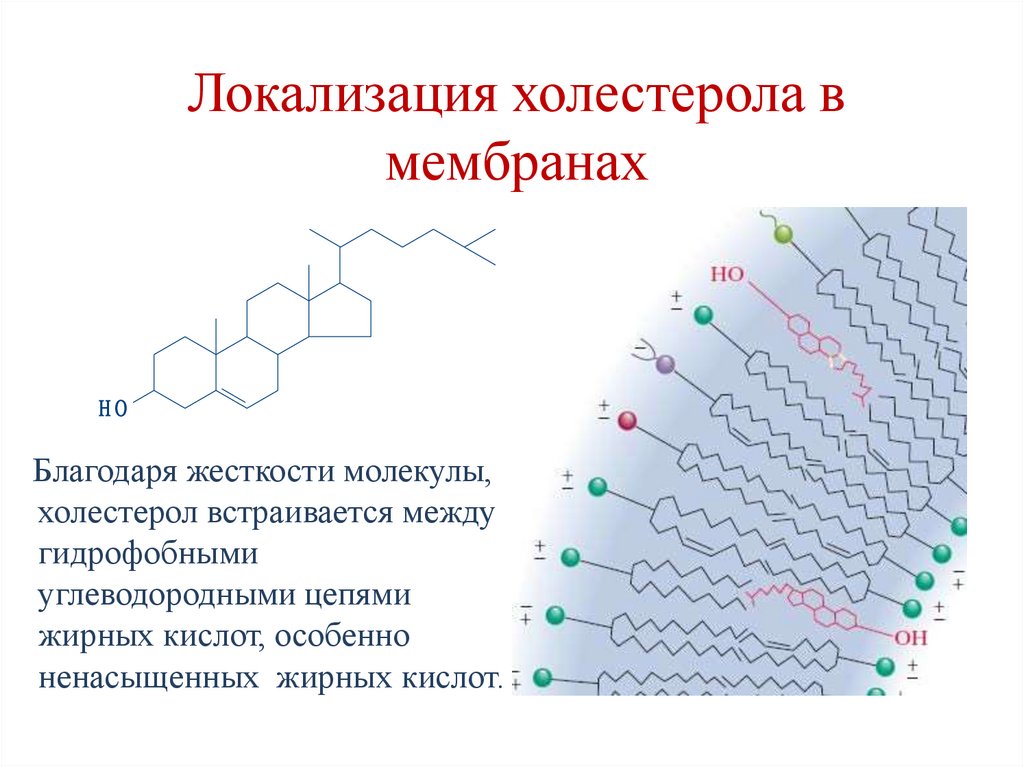
Локализация холестерола вмембранах
HO
Благодаря жесткости молекулы,
холестерол встраивается между
гидрофобными
углеводородными цепями
жирных кислот, особенно
ненасыщенных жирных кислот.
19.
Функции холестерола в мембранах• Регулирует текучесть
мембран.
• Стабилизирует
углеводородные цепи
насыщенных жирных
кислот, удерживая их в
прямом виде за счет Вандер-Ваальсовых связей. В
результате мембраны
становятся более
жесткими и
резистентными.
20.
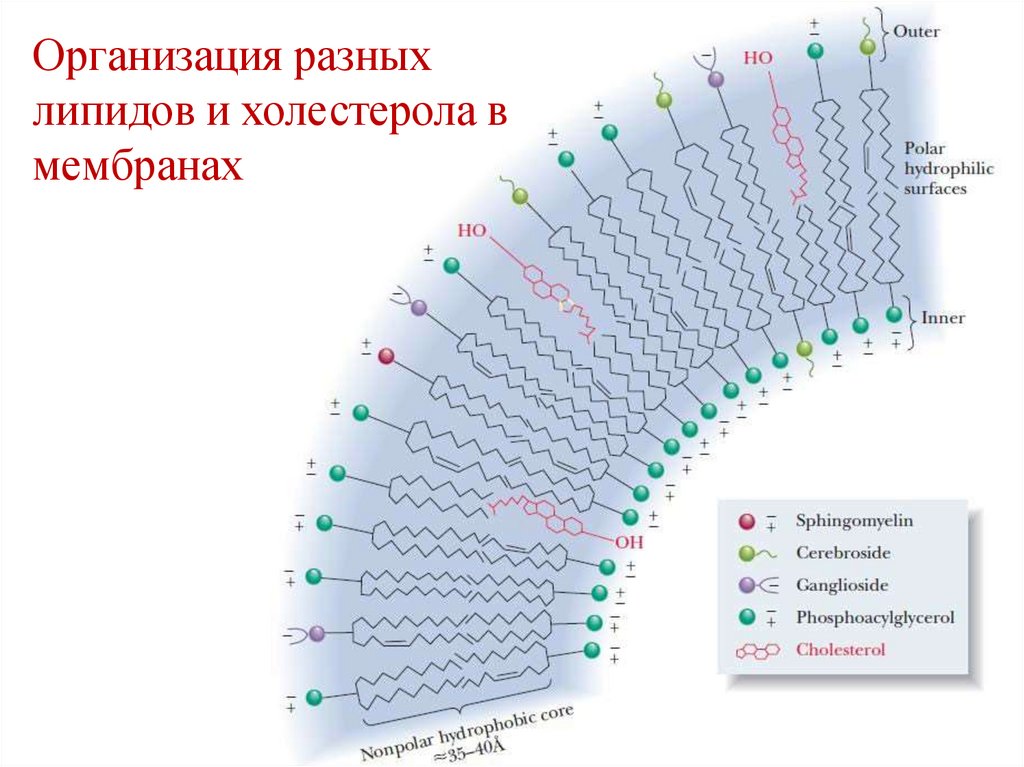
Организация разныхлипидов и холестерола в
мембранах
21.
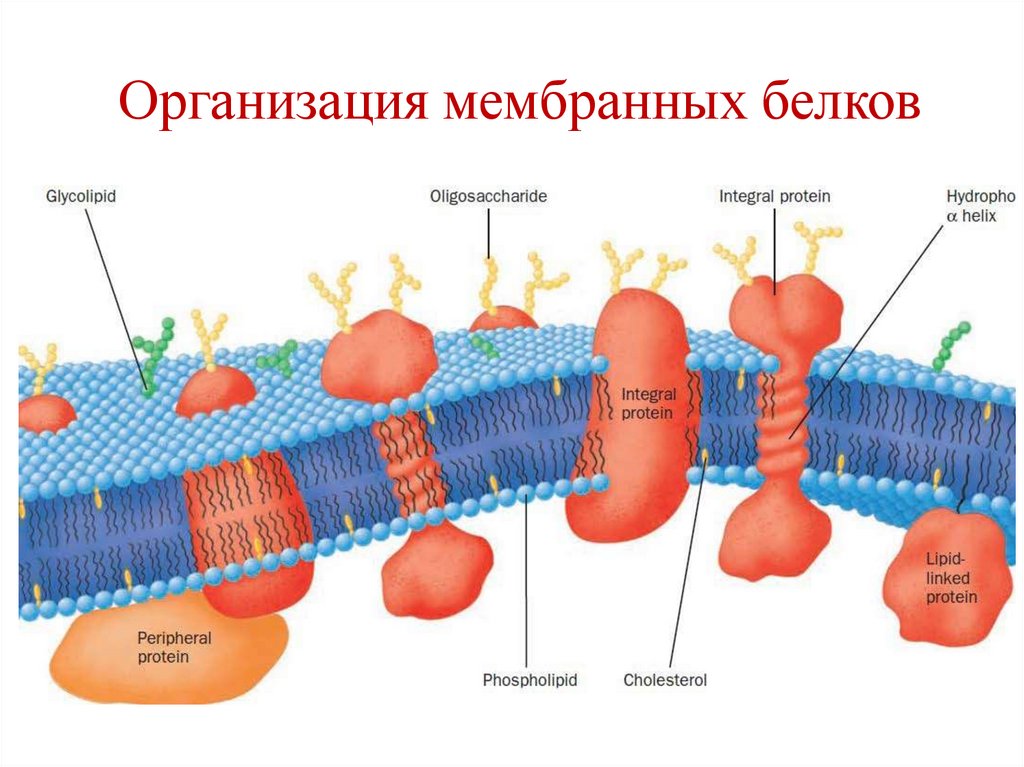
Белки мембранПо положению в мембране белки делятся на:
Интегральные;
Поверхностные.
Поверхностные белки связаны с полярными
головками липидов ионными и водородными
связями.
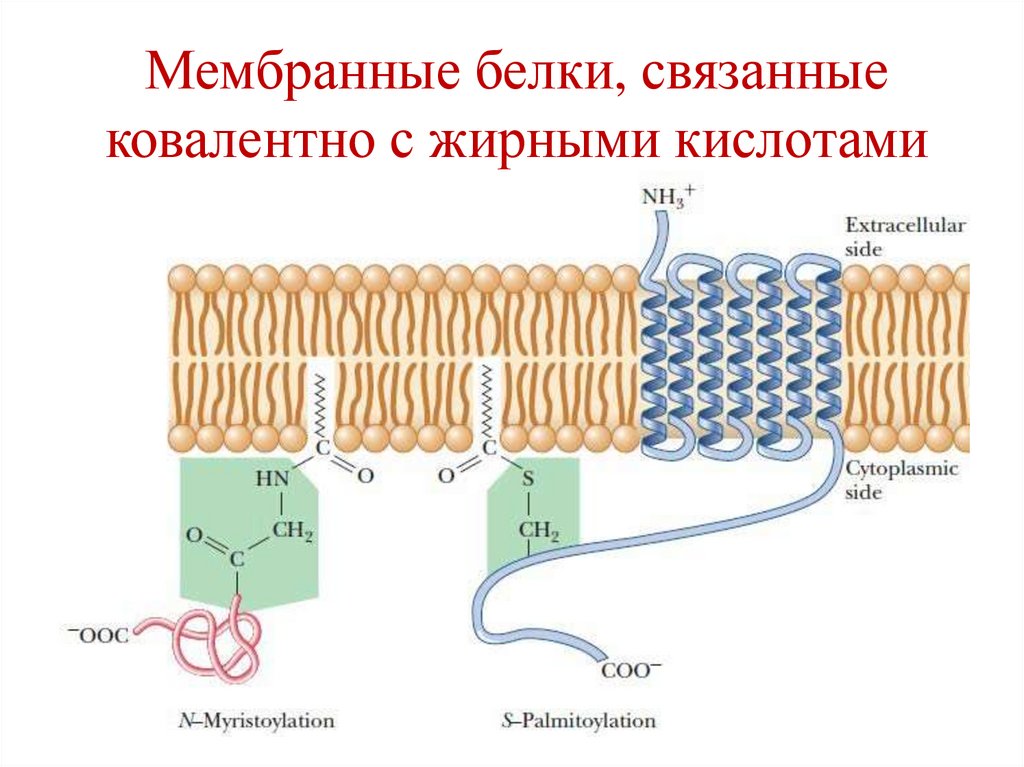
• Интегральные белки связаны более прочно с
липидами посредством многочисленных
гидрофобных взаимодействий, а также
ковалентных связей.
22.
Мембранные белкиФункции:
Транспортная.
Рецепторная.
Каталитическая.
Связывание цитоскелета с внеклеточным
матриксом.
Белки клеточных мембран обладают
латеральной диффузией.
Состав белков определяет особенности
функционирования каждой мембраны.
23.
Организация мембранных белков24.
Мембранные белки, связанныековалентно с жирными кислотами
25.
Углеводы клеточных мембран –гликокаликс
• Гликокаликс расположен на наружной
поверхности мембраны и представляет
собой молекулы олигосахаридов.
• Выполняет рецепторную и маркерную
функции, а также участвует в обеспечении
избирательности транспорта веществ,
защищают клетку от протеолиза, участвуют
в пристеночном пищеварении.
26.
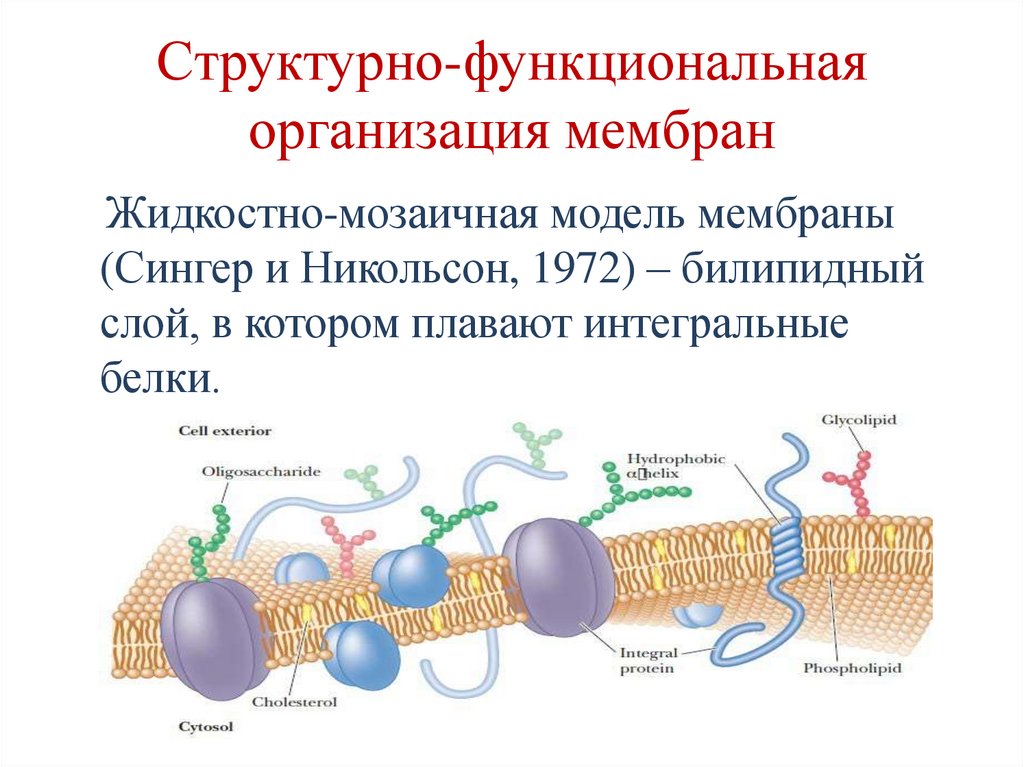
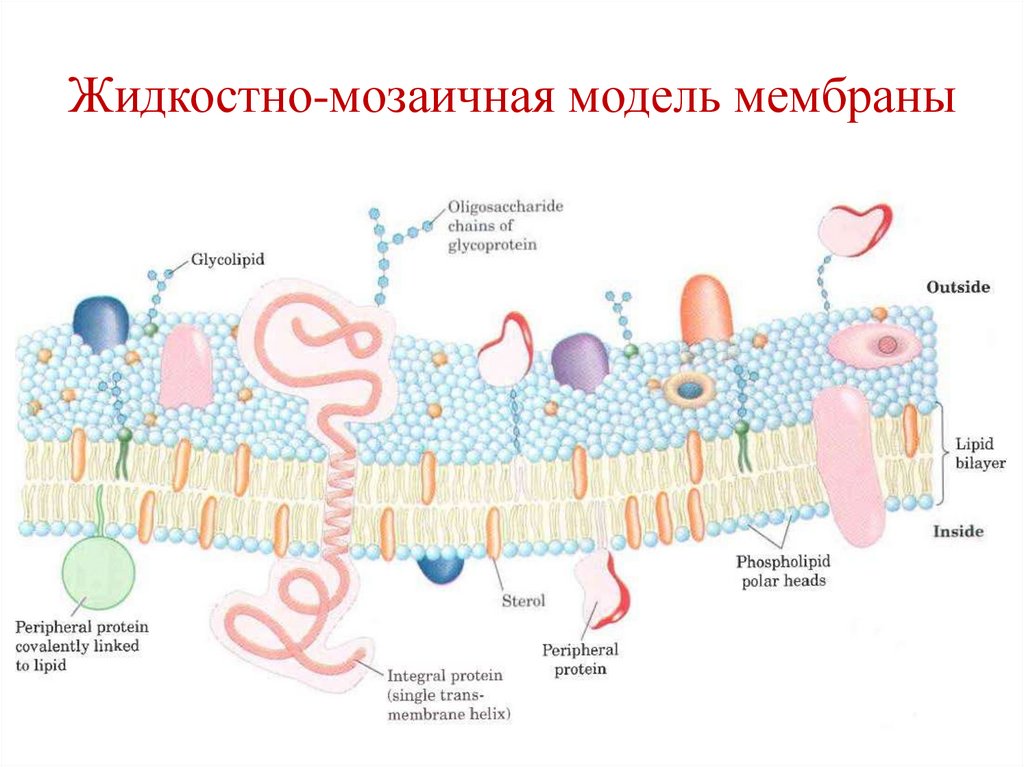
Структурно-функциональнаяорганизация мембран
Жидкостно-мозаичная модель мембраны
(Сингер и Никольсон, 1972) – билипидный
слой, в котором плавают интегральные
белки.
27.
Жидкостно-мозаичная модель мембраны28.
Свойства биологических мембранЖидкостность (текучесть).
Трансмембранная (поперечная) ассиметрия.
Подвижность липидов и белков.
Избирательная проницаемость.
Самосборка
Аutorepararea
29.
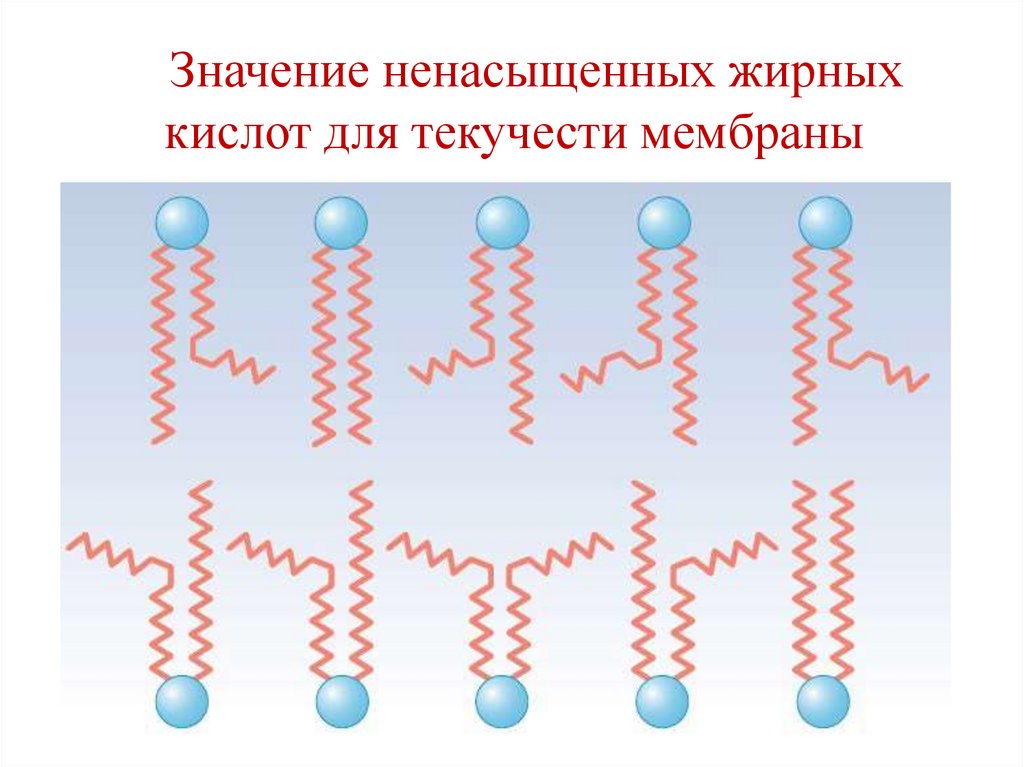
Жидкостность (текучесть) мембран –• зависит от относительного содержания
насыщенных и ненасыщенных жирных кислот
в мембранных липидах.
• Цис-конформация ненасыщенных жирных
кислот создает «изломы» в цепи,
препятствующие более плотной упаковке
молекул в мембране, делая ее более «текучей».
• На жидкостность влияет и длина цепи ЖК, с
удлинением которых мембрана становится
более «текучей».
• Холестерол регулирует текучесть мембран.
30.
Значение ненасыщенных жирныхкислот для текучести мембраны
31.
Трансмембранная (поперечная)ассиметрия
1. Фосфатидилхолины и сфингомиелины (более
объемные головки) расположены в наружном
монослое мембраны, а фосфатидилсерины и
фосфатидилэтаноламины – во внутреннем
монослое мембраны.
2. Холестерол содержится в наружном слое в
больших количествах, чем во внутреннем.
3. Белки клеточных мембран расположены
ассиметрично.
4. Углеводы содержатся на наружной
поверхности мембраны.
32.
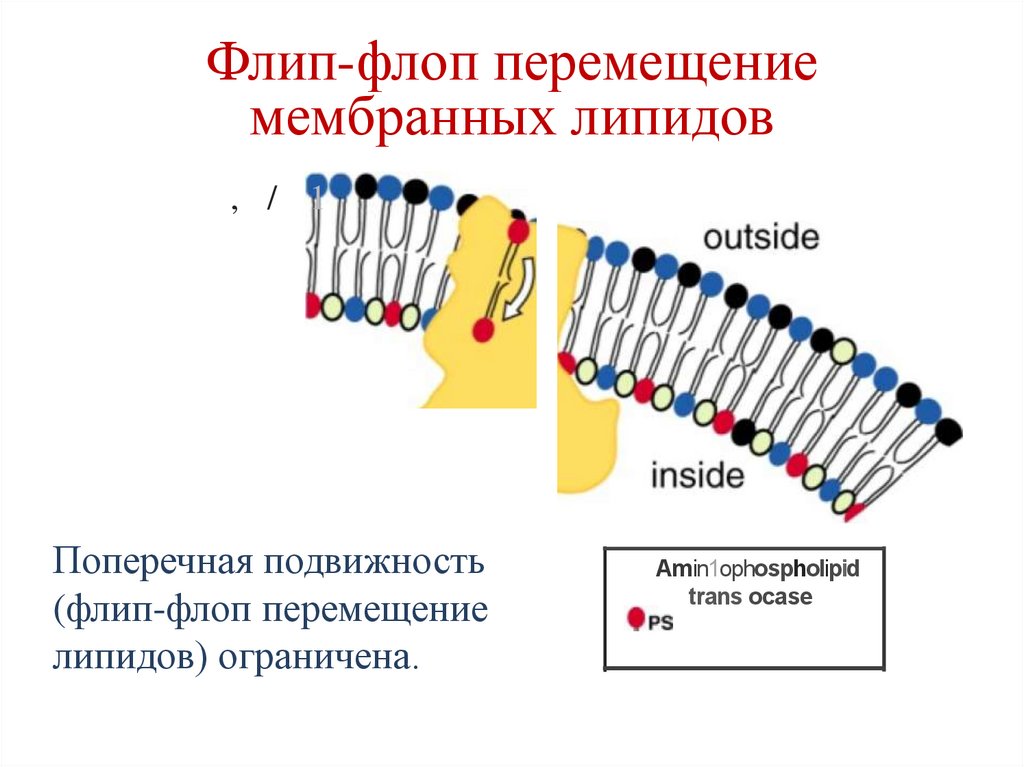
Флип-флоп перемещениемембранных липидов
, / 1
Поперечная подвижность
(флип-флоп перемещение
липидов) ограничена.
Amin1ophospholipid
trans ocase
33.
Трансмембранный транспорт1.
2.
Мембрана обладает избирательной проницаемостью
для разных веществ.
Типы трансмембранного транспорта:
Пассивный транспорт – из области большей
концентрации в область меньшей концентрации):
простая диффузия;
облегченная диффузия;
трансмембранные каналы.
Активный транспорт – против градиента
концентрации:
первичный;
вторичный.
34.
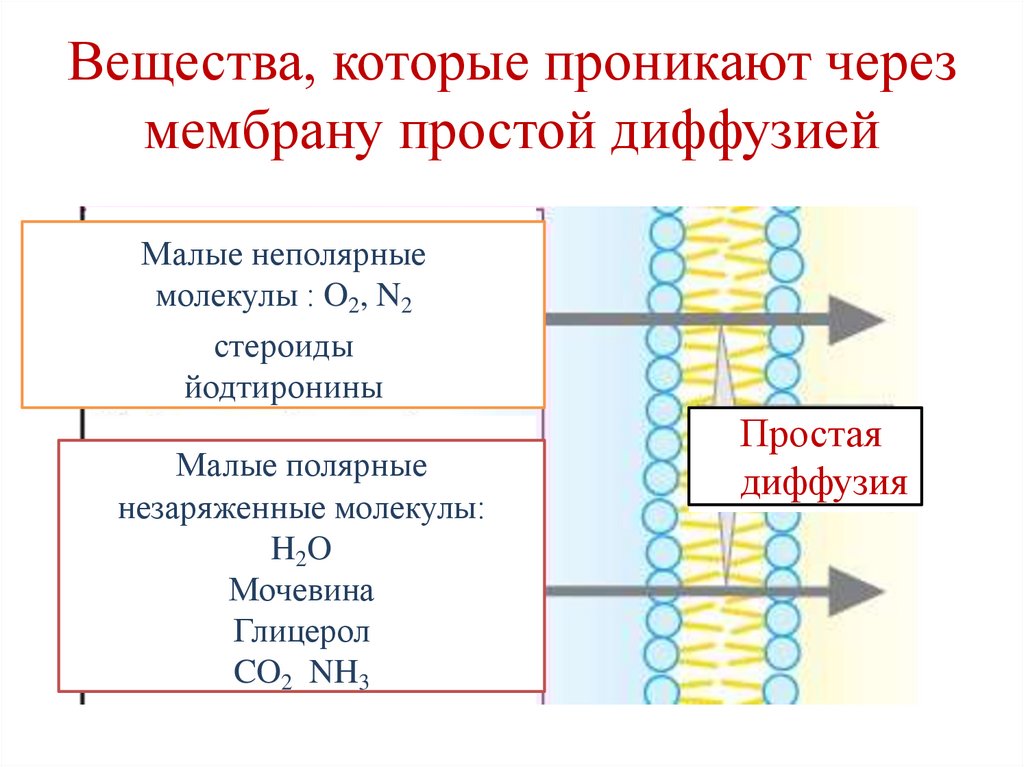
Вещества, которые проникают черезмембрану простой диффузией
Малые неполярные
молекулы : O2, N2
стероиды
йодтиронины
Малые полярные
незаряженные молекулы:
H2O
Мочевина
Глицерол
CO2 NH3
Простая
диффузия
35.
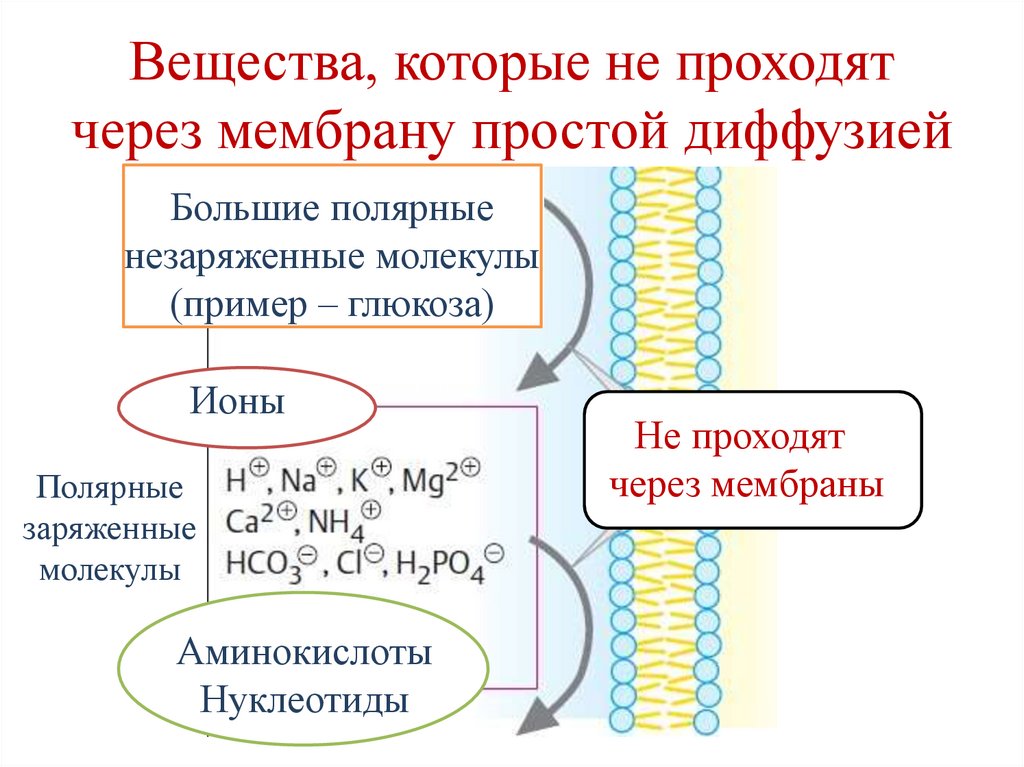
Вещества, которые не проходятчерез мембрану простой диффузией
Большие полярные
незаряженные молекулы
(пример – глюкоза)
Ионы
Полярные
заряженные
молекулы
Аминокислоты
Нуклеотиды
Не проходят
через мембраны
36.
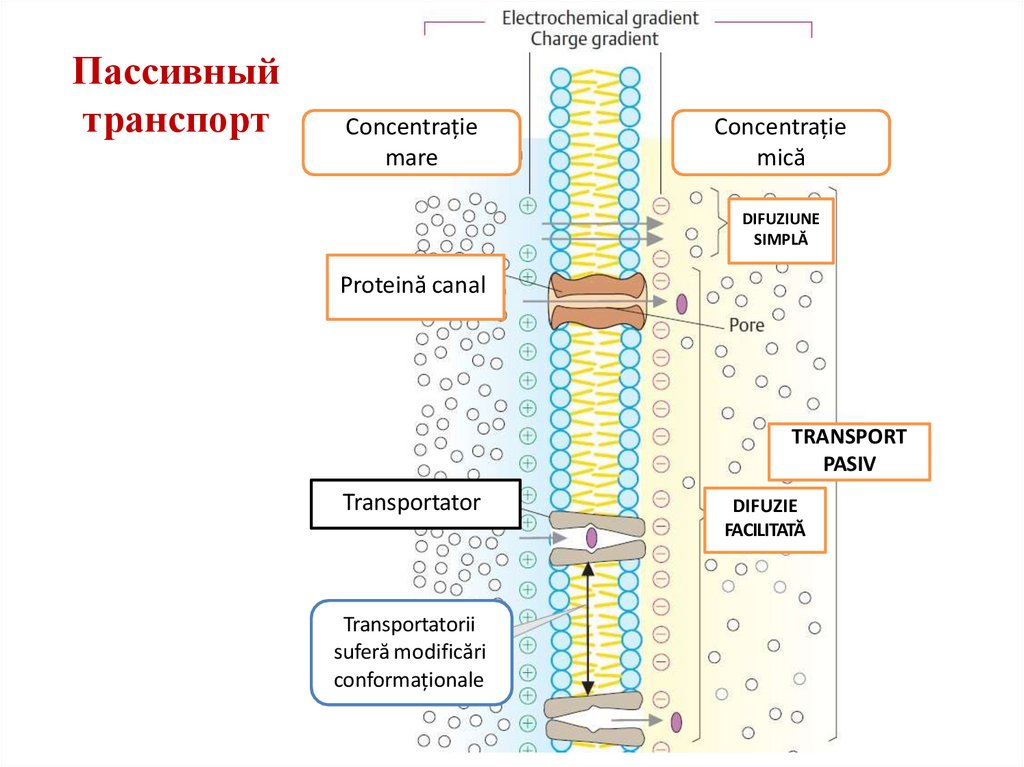
Пассивныйтранспорт
Concentrație
mare
Concentrație
mică
DIFUZIUNE
SIMPLĂ
Proteină canal
TRANSPORT
PASIV
Transportator
Transportatorii
suferă modificări
conformaționale
DIFUZIE
FACILITATĂ
37.
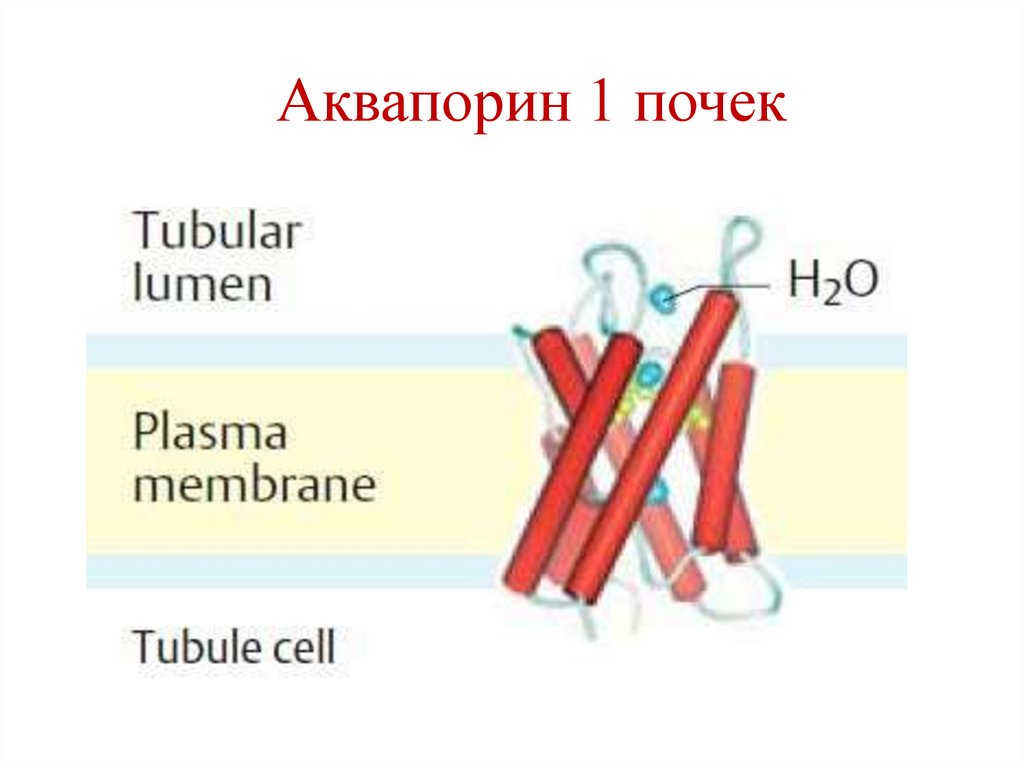
Трансмембранные каналыРазличают альфа- и бета-каналы.
Альфа-каналы являются гомо- или
гетероолигомерными белками, содержащие альфаспиральные трансмембранные участки.
Примеры: потенциал-зависимые каналы для ионов
Na, K, Ca, Cl;
Аквапорины;
Лиганд-зависимые каналы;
сАМФ-зависимые каналы.
38.
Аквапорин 1 почек39.
Трансмембранные вета-каналы(порины)
• Это белки содержащие трансмембранные
вета-структуры, образующие цилиндр
диаметром 0.6-3 nm.
• Содержатся в наружной мембране
митохондрий, являются неселективными
каналами для большого количества веществ
(от ионов до небольших белков).
40.
Облегченная диффузия• Осуществляется с участием белковтранслоказ.
• Транслоказы взаимодействуют со
специфическим лигандом, обеспечивая его
диффузию через мембрану.
• Транслоказы в процессе взаимодействия и
переноса претерпевают конформационые
изменения (в отличие от каналов).
41.
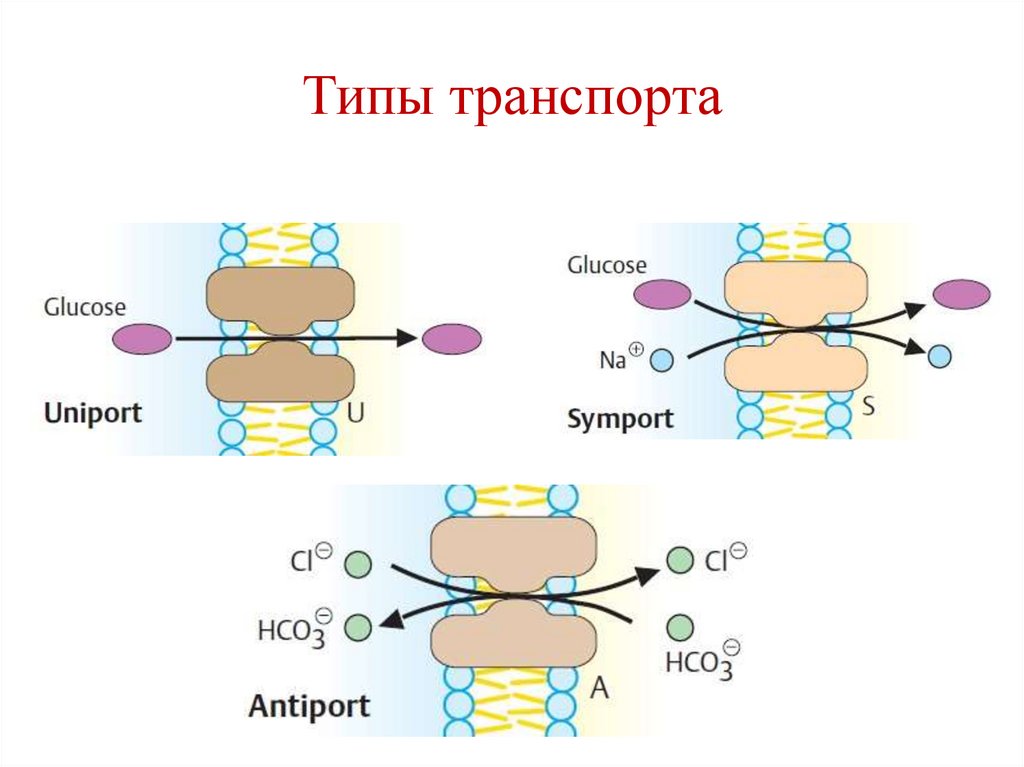
Типы транспорта42.
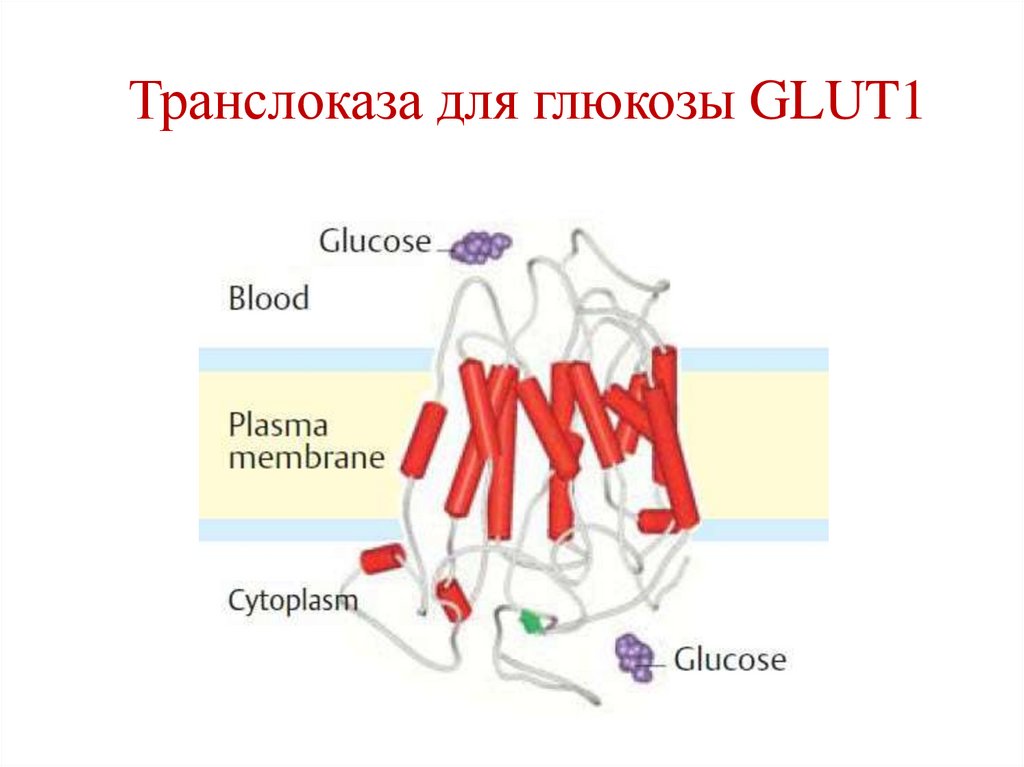
Примеры типов транспорта• Унипорт – транслоказа для глюкозы GLUT1
(эритроциты).
• Симпорт – перенос АТР и АDP через
внутреннюю мембрану митохондрий.
• Антипорт – анионный переносчик
мембраны эритроцитов (Cl- și HCO3- ).
43.
Транслоказа для глюкозы GLUT144.
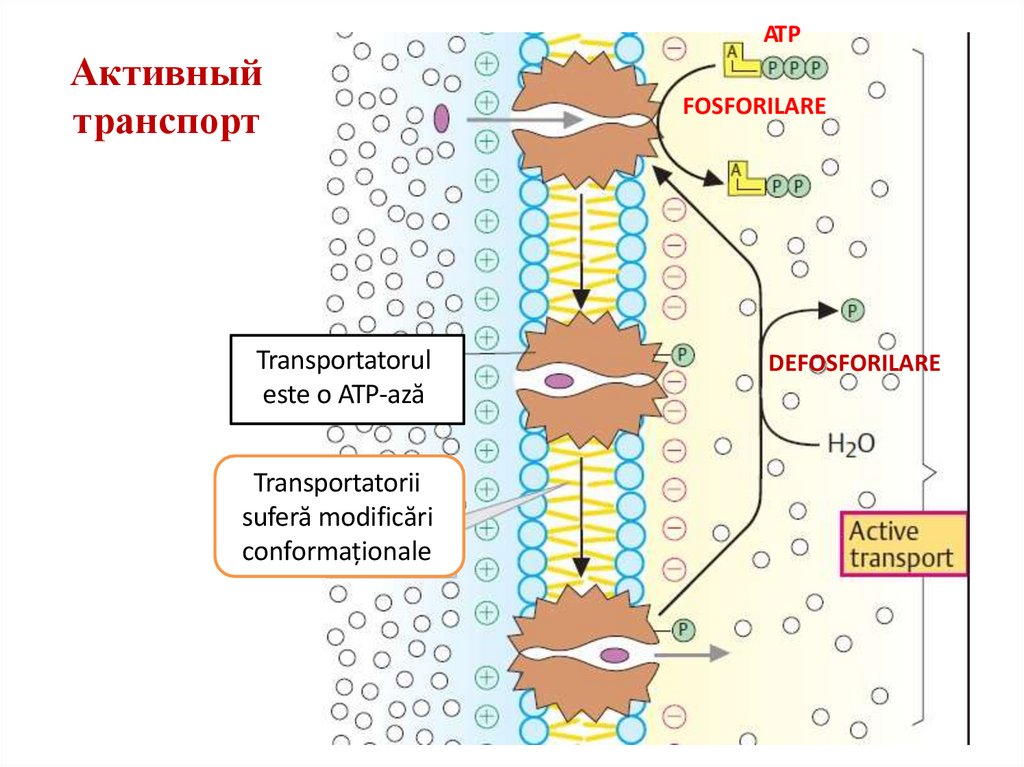
ATPАктивный
транспорт
Transportatorul
este o ATP-ază
Transportatorii
suferă modificări
conformaționale
FOSFORILARE
DEFOSFORILARE
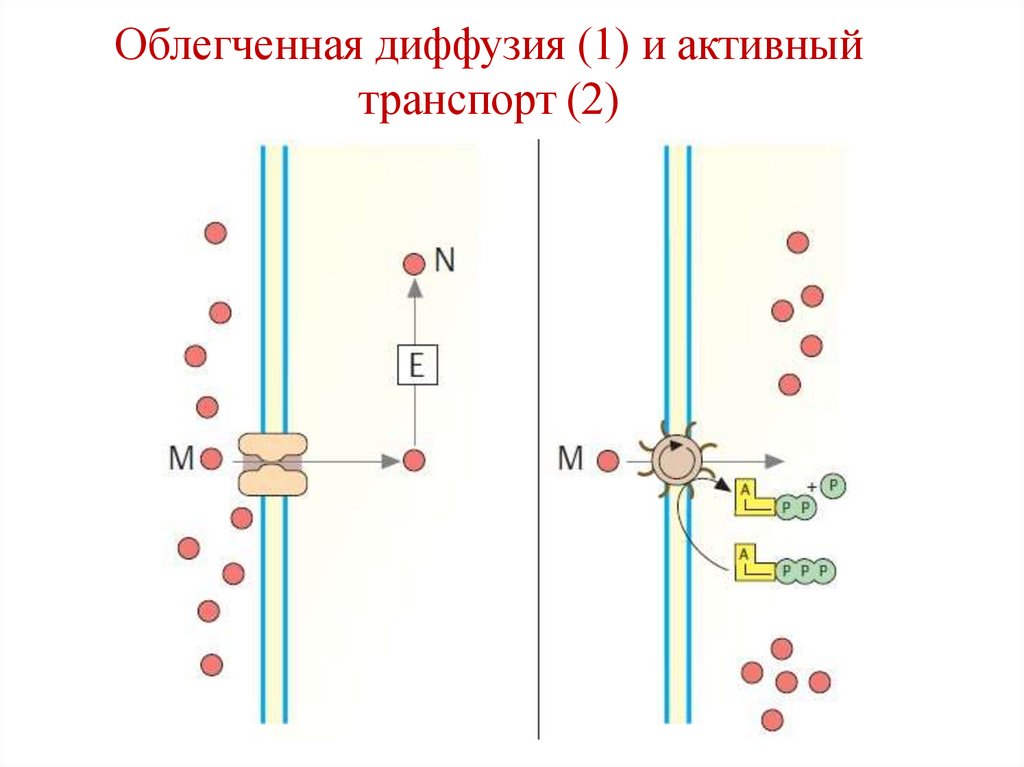
45.
Облегченная диффузия (1) и активныйтранспорт (2)
46.
Первично-активный транспорт• Перенос веществ против градиента
концентрации с использованием
транспортных АТФ-аз (ионных насосов).
АТФ-азы расщепляют АТФ, используя
фосфат для временного
аутофосфорилирования.
• Различают 3 класса транспортных АТФ-аз –
P, V и F типа.
47.
Типы транспортных АТФ-аз• Тип Р – транспортируют ионы Na, K, Ca (около 300
представителей).
• Тип V (V – от вакуоль) являются протонными насосами,
ответственными за подкисление содержимого лизосом,
эндосом, секреторных везикул и везикул аппарата Гольджи.
• Тип F присутствуют в мембранах митохондрий,
хлоропластов и бактерий. Транспортируют H+ c
использованием ATP.
• АВС-транспортеры ( ABCt – ATP-binding cassette) –
используют ATP для переноса лиганда.
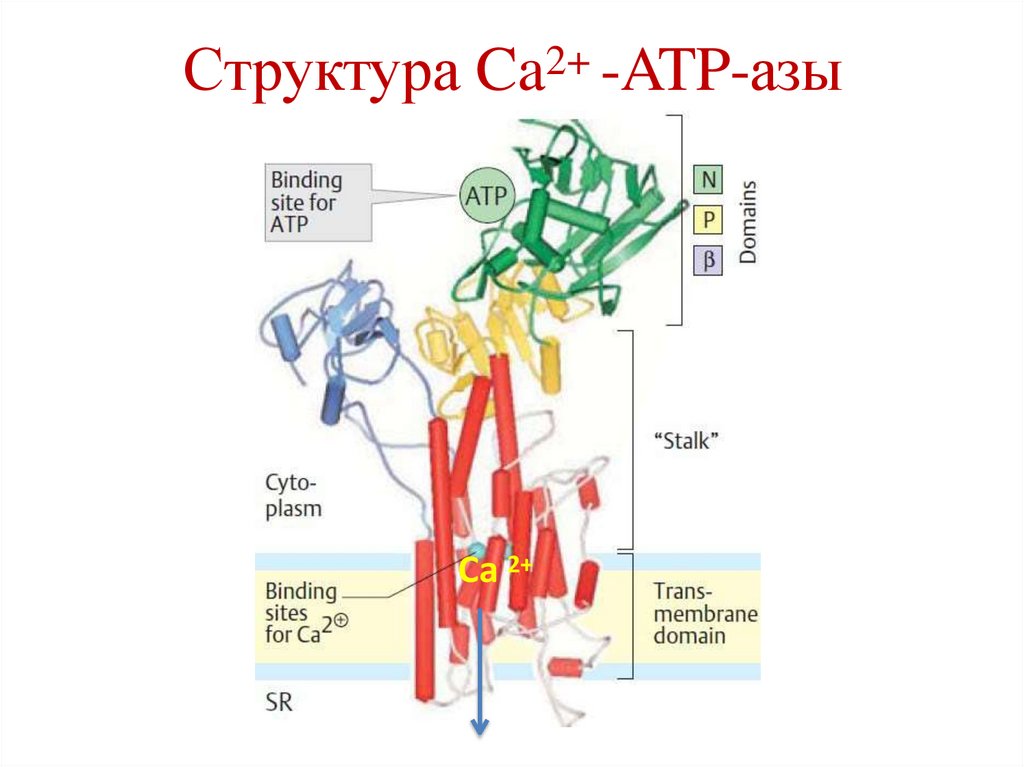
48.
Структура Ca2+ -ATP-азыCa 2+
49.
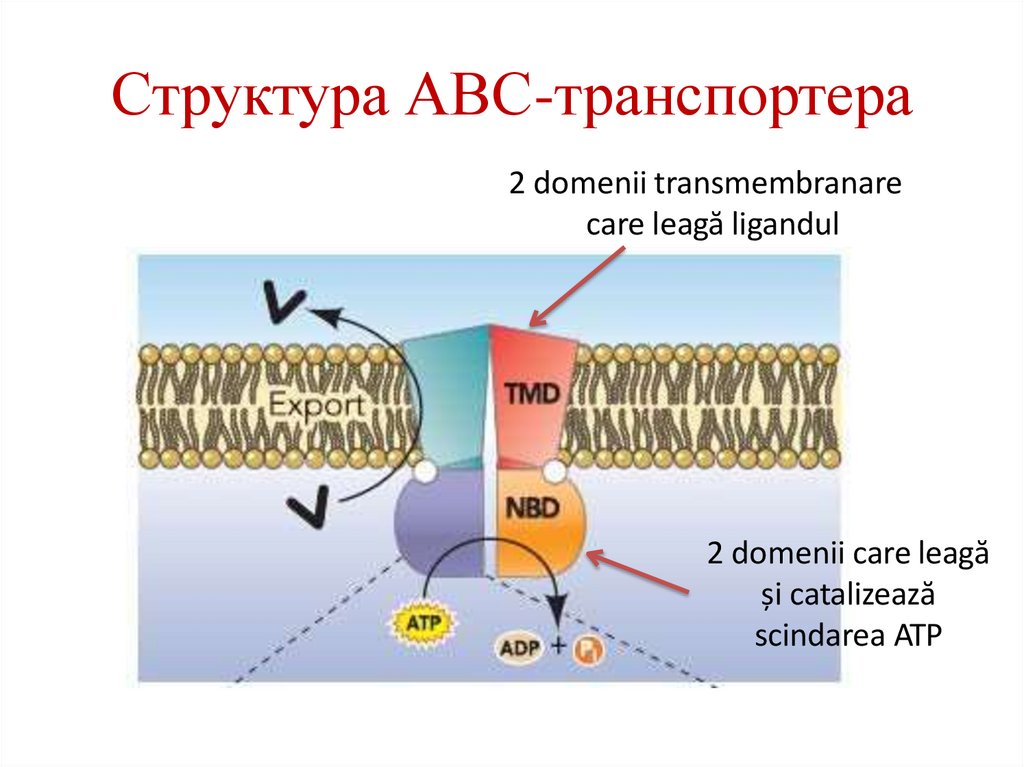
Структура АВС-транспортера2 domenii transmembranare
care leagă ligandul
2 domenii care leagă
și catalizează
scindarea ATP
50.
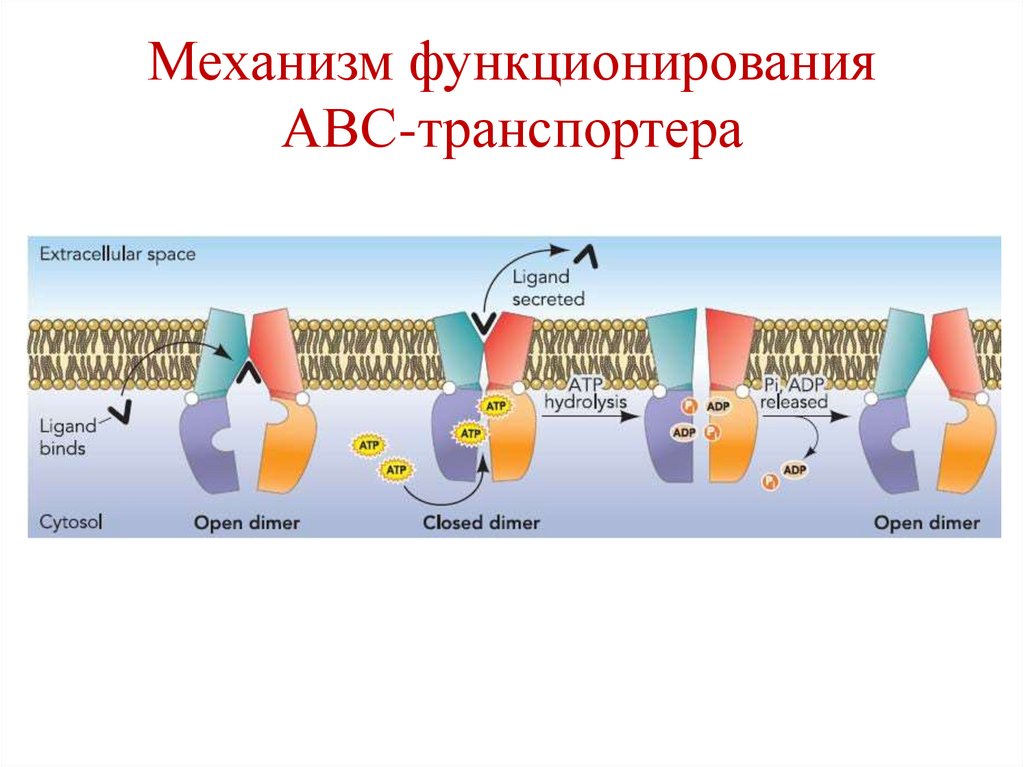
Механизм функционированияАВС-транспортера
51.
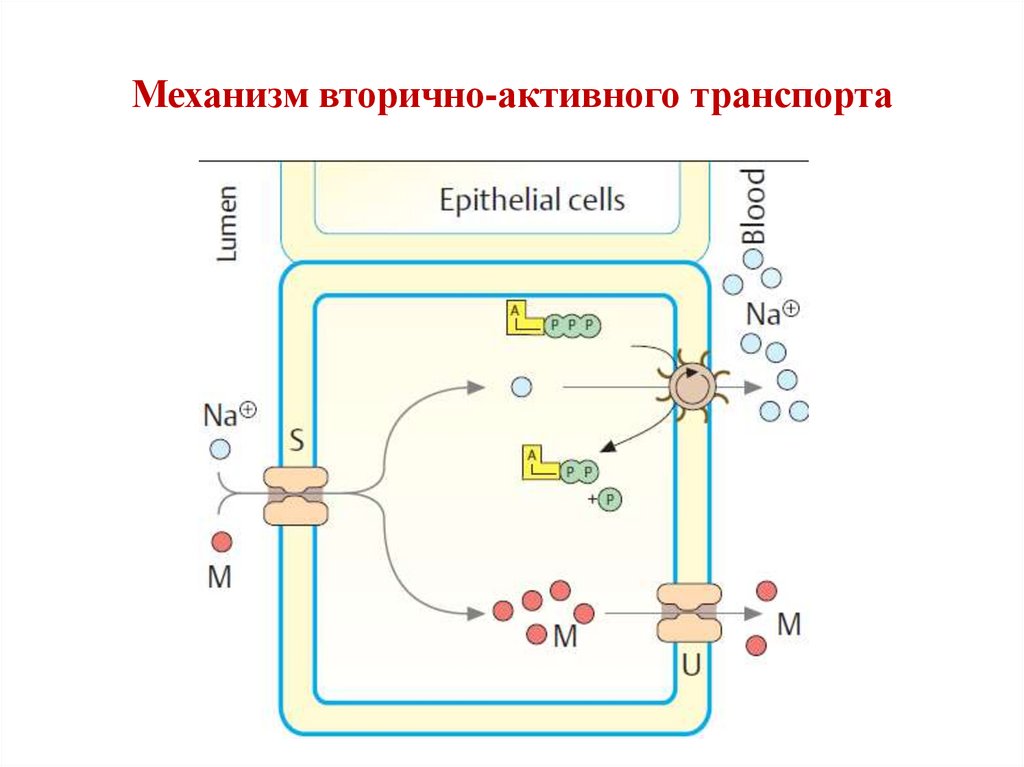
Вторично-активный транспорт• Осуществляется с участием Na+-зависимых
транспортеров (SSAT – sodium-dependent secondary active
transporters).
• Существует около 400 представителей.
• SАAT осуществляют симпорт веществ (глюкоза,
аминокислоты и др.) с Na+ , используя
электрохимический градиент Na+.
• Избыток Na+ выкачивается из клетки в обмен на K+ с
участием Na+/K+-ATP-азы. (ATP используется для
поддержания градиента ATP, а не непосредственно для
транспорта вещества).
52.
Механизм вторично-активного транспорта53.
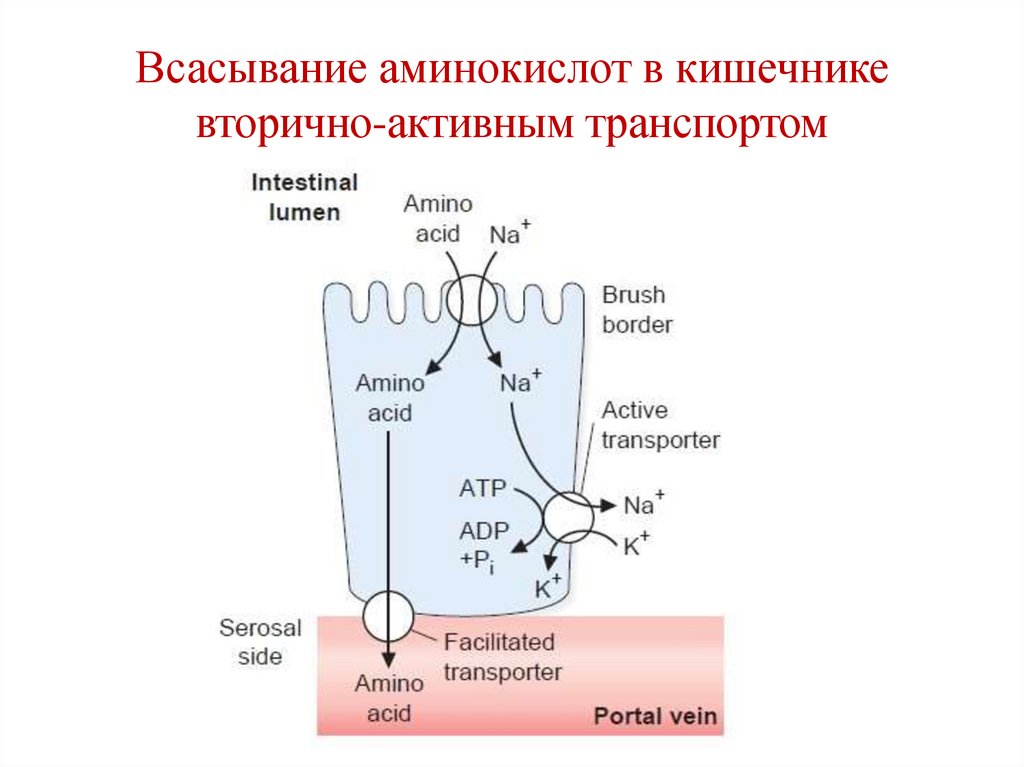
Всасывание аминокислот в кишечникевторично-активным транспортом
54.
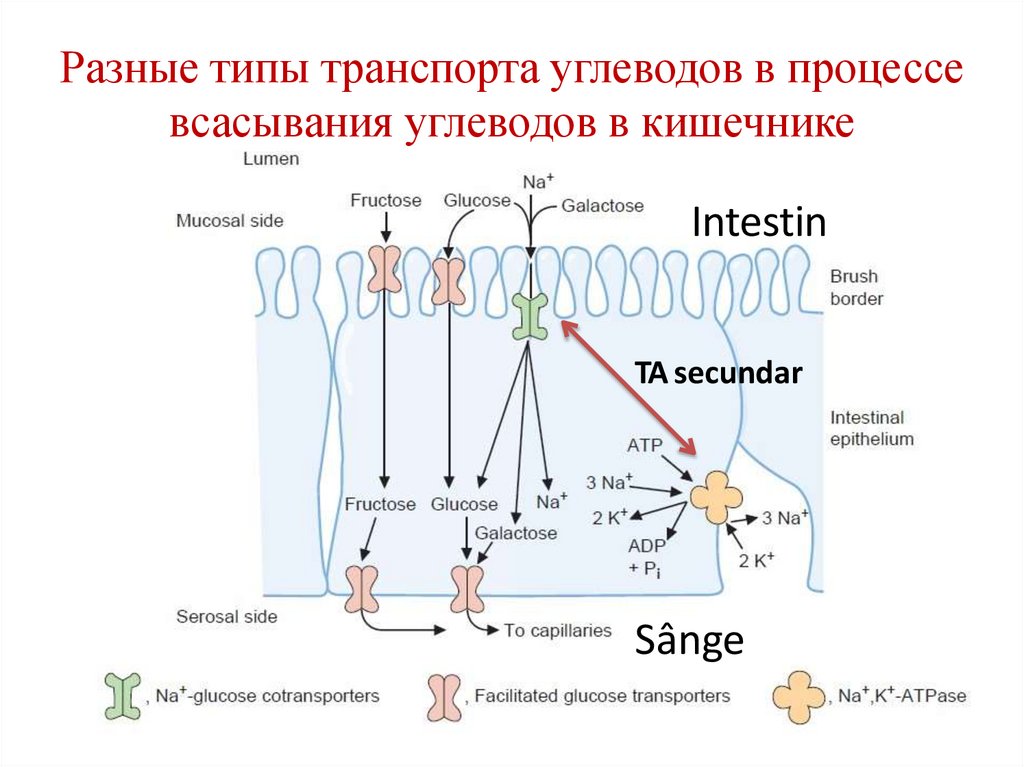
Разные типы транспорта углеводов в процессевсасывания углеводов в кишечнике
Intestin
TA secundar
Sânge