






")

")


")



")

")















 Биология
БиологияПохожие презентации:










Цепь тканевого дыхания
1. ЦЕПЬ ТКАНЕВОГО ДЫХАНИЯ
2. ВОПРОСЫ ЛЕКЦИИ
• Пути использования кислорода в клетке• Компоненты и организация
дыхательной цепи.
• Цепь переноса электронов
• Окислительное фосфорилирование
• Разобщители и ингибиторы
дыхательной цепи
• Нарушения работы дыхательной цепи
3. Пути использования кислорода в клетке
О2ЦТД
(90%)
СРО активные формы
СВОБОДНОЕ ОКИСЛЕНИЕ
оксидазы оксигеназы
НАДФН + Н +
МАО
(ФАД)
диоксигеназы
монооксигеназы
Цитохром Р450 (редуктаза)
МСО
(гепатоциты)
ПОЛ
R+O2=RO2
Цитохром Р450
(гидроксилирование АрАК)
4. Пути использования кислорода в клетке
Существует следующие пути использования кислородав клетке, которые характеризуются следующими
процессами:
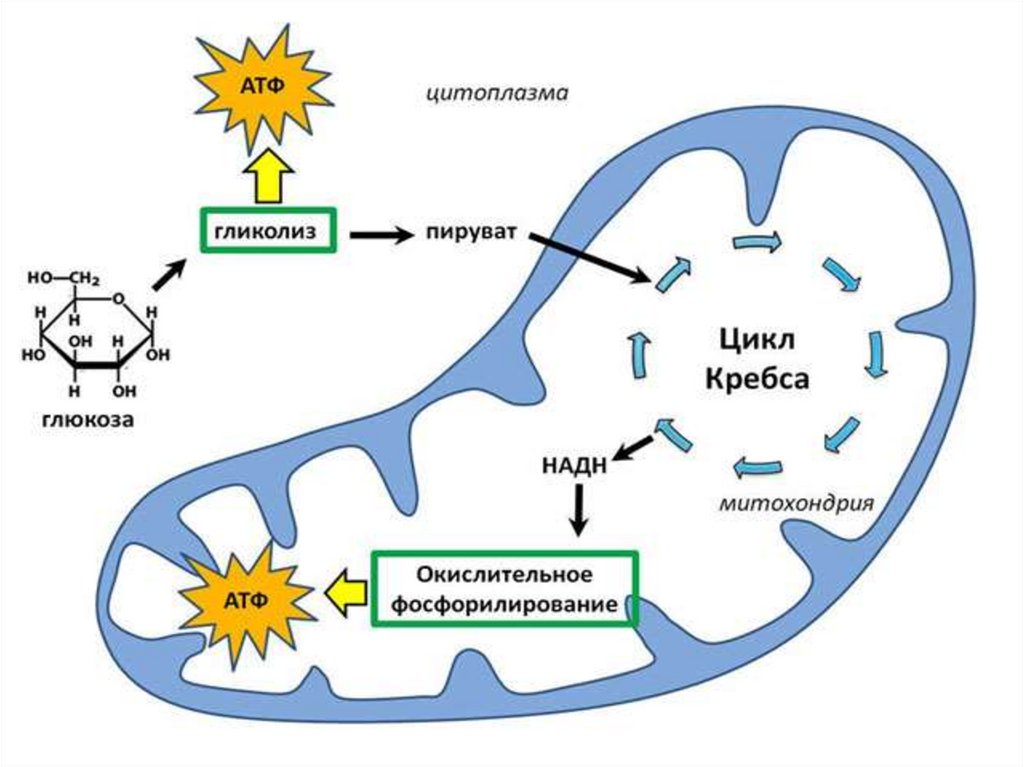
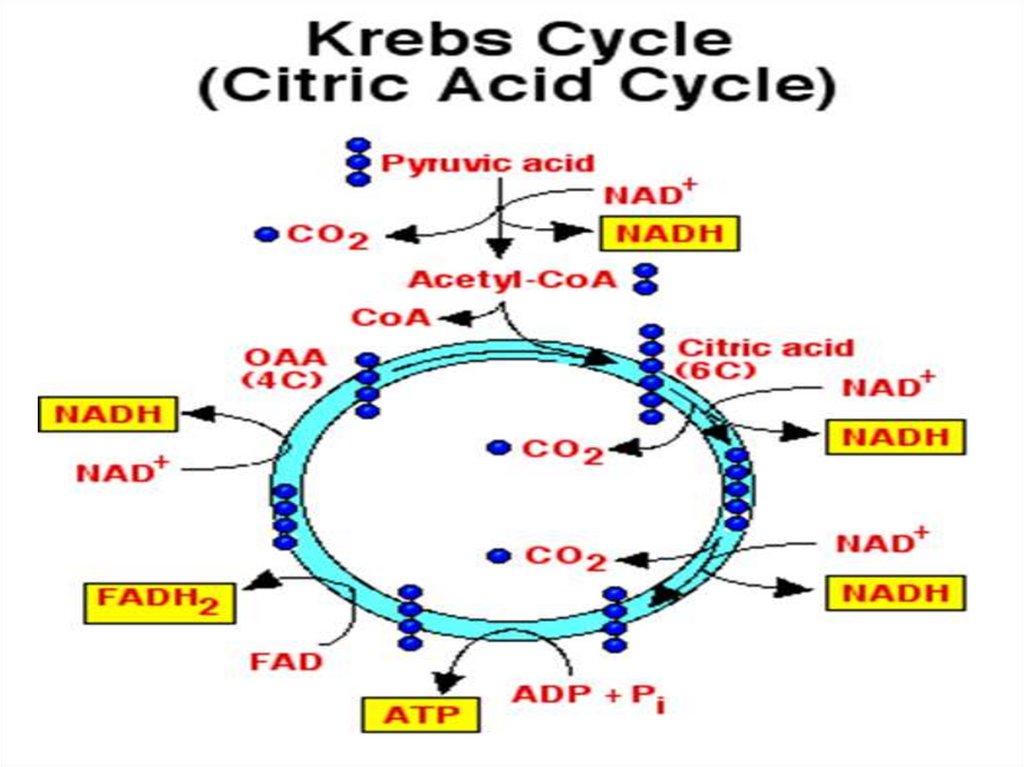
1. Тканевое дыхание
(90% поступившего кислорода восстанавливается до
Н2О при участии фермента цитохромоксидазы) с
образованием АТФ (окислительное фосфорилирование)
02 +4е+4Н+ → 2Н2О
+ АТФ
Тканевое дыхание (цепь переноса электронов) окислительное фосфорилирование. Является основным
источником АТФ в аэробных тканях.
Потребляет 90% кислорода
5. ДЫХАТЕЛЬНАЯ ЦЕПЬ МИТОХОНДРИЙ
Или Цепь Тканевого Дыхания (или цепь переносаэлектронов) - совокупность окислительновосстановительных процессов в клетках, органах и
тканях, протекающих с участием молекулярного
кислорода и сопровождающихся запасанием
энергии в фосфорильной связи молекул АТФ.
Тканевое дыхание является важнейшей частью
обмена веществ и энергии в организме. В
результате тканевого дыхания при участии
специфических ферментов происходит
окислительный распад крупных органических
молекул — субстратов дыхания — до более
простых и в конечном счете до СО2 и Н2О с
высвобождением энергии АТФ.
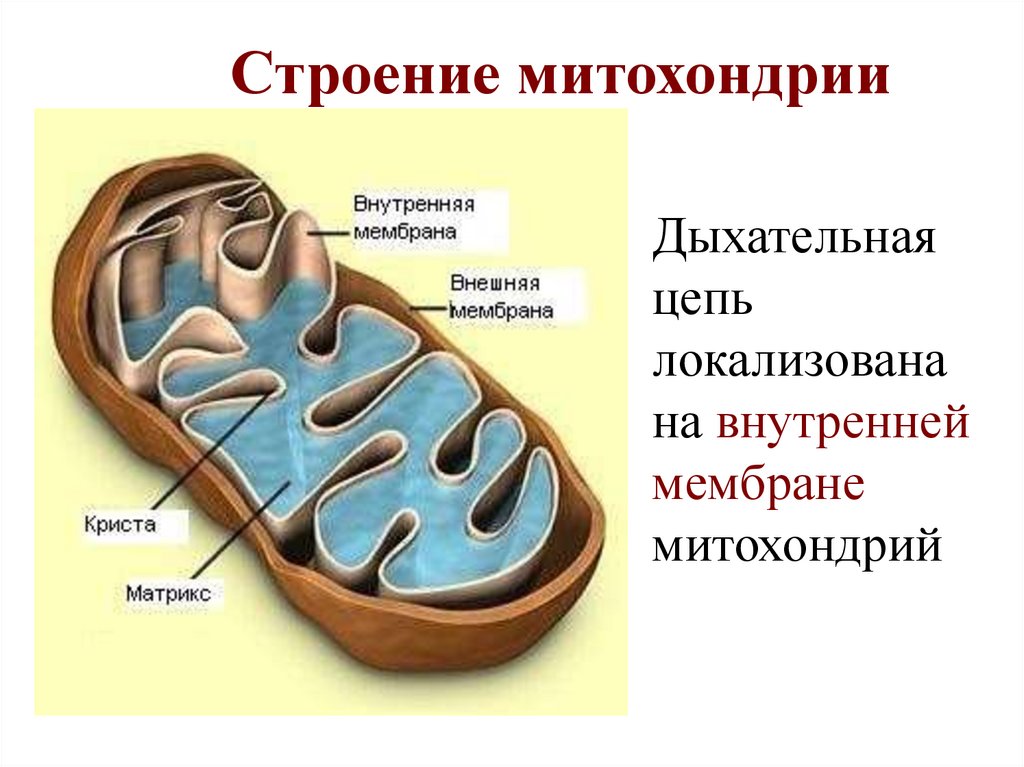
6.
Строение митохондрииДыхательная
цепь
локализована
на внутренней
мембране
митохондрий
7.
8.
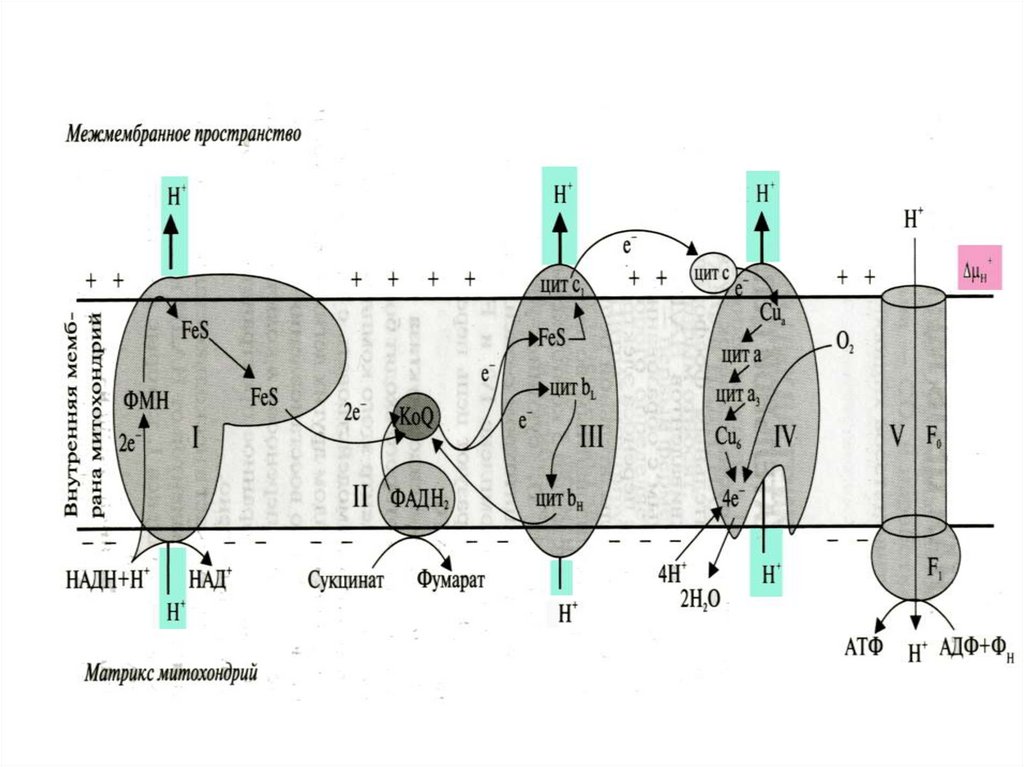
9. СХЕМА СТРОЕНИЯ ДЫХАТЕЛЬНОЙ ЦЕПИ МИТОХОНДРИЙ
10. ДЫХАТЕЛЬНАЯ ЦЕПЬ МИТОХОНДРИЙ
I. НАДН-KoQ-редуктаза (содержит промежуточныеакцепторы водорода: флавинмононуклеотид и
железосерные белки).
II. Сукцинат-KoQ-редуктаза
(содержит промежуточные акцепторы водорода: ФАД
и железосерные белки).
III. KoQН2 -цитохром с-редуктаза
(содержит акцепторы электронов: цитохромы b и с1
гемное железо).
IV. Цитохром с-оксидаза (содержит акцепторы
электронов: цитохромы а и а3, ионы меди Cu2+).
В качестве промежуточных переносчиков электронов
выступают убихинон (коэнзим Q) и цитохром с.
11. КОМПЛЕКС I (NADH: УБИХИНОН-РЕДУКТАЗА)
12.
13. КОМПЛЕКС I
Сложный флавопротеиновый комплекс, в составкоторого входит 25 полипептидных цепей. НАДНсвязывающий центр ориентирован в матрикс, это
позволяет ему взаимодействовать с НАДН,
восстановленным дегидрогеназами ЦТК и
другими дегидрогеназами.
Электроны движутся по атомам железа (негемное
железо) внутри железо-серных белков.
Комплекс катализирует реакцию восстановления
убихинона (KoQ), сопряженную с
трансмембранным переносом 4-х протонов из
матрикса митохондрий в межмембранное
пространство
14. КОМПЛЕКС I (NADH:УБИХИНОН-РЕДУКТАЗА)
КОМПЛЕКС I (NADH:УБИХИНОНРЕДУКТАЗА)15. СТРОЕНИЕ ЖЕЛЕЗО-СЕРНЫХ КЛАСТЕРОВ
16. Комплекс II
Сукцинат-убихинон-редуктаза:осуществляет перенос восстановительных
эквивалентов от ФАДН2 (образованный в
ходе реакции окисления сукцината в ЦТК)
на убихинон (KoQ) через Fe-S центры.
Биологическая роль коэнзима Q – перенос
электронов в дыхательной цепи от
флавопротеинов (комплексы I и II) к
цитохромам (комплекс III).
17. КОМПЛЕКС II (СУКЦИНАТ:УБИХИНОН-РЕДУКТАЗА)
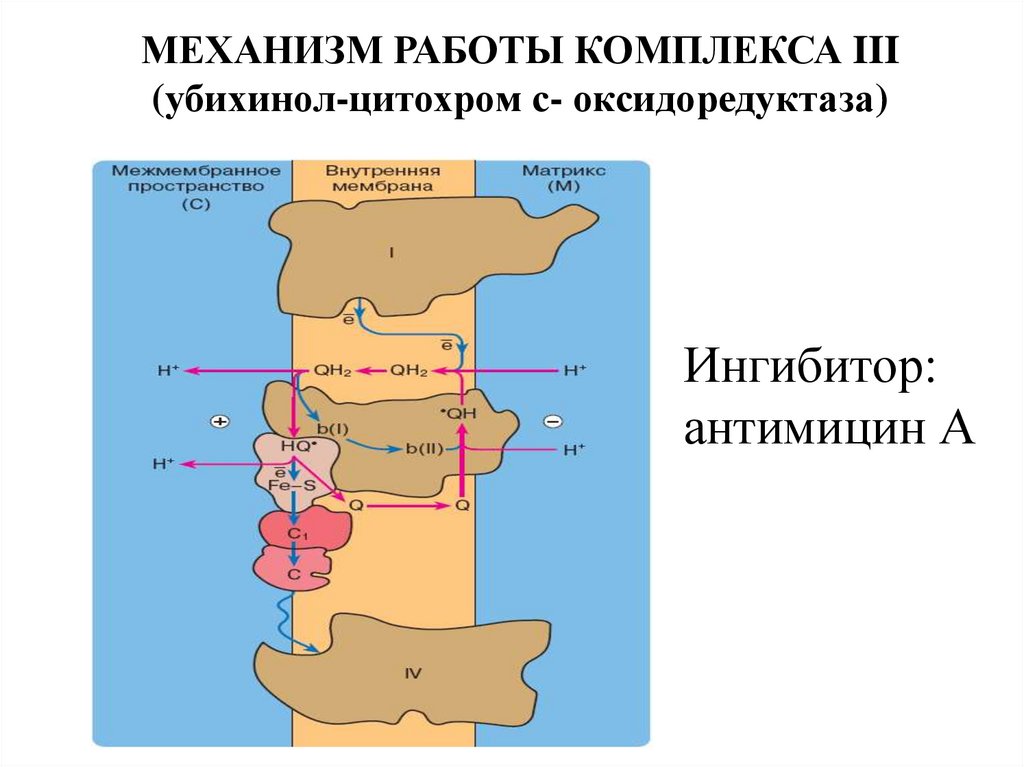
18. Комплекс III: убихинон-цитохром с-редуктаза
Это комплекс цитохромов bc1 осуществляетперенос электронов от QН2 на цитохром c.
Цитохромы – это гемопротеины. Их 5 типов:
b, c, c1 , a и a3.
Цитохром с – сложный белок, хромопротеин,
простетическая группа которого – гем –
содержит железо с переменной валентностью
(Fe3+ в окисленной форме и Fe2+ в
восстановленной форме). Цитохром с является
водорастворимым соединением и располагается
на периферии внутренней митохондриальной
мембраны в гидрофильной фазе.
19. КОМПЛЕКС III
Биологическая роль цитохрома с –перенос электронов в дыхательной
цепи от комплекса III к комплексу IV.
Здесь также осуществляется
трансмембранный перенос 4-х
протонов из матрикса митохондрий в
межмембранное пространство
20.
МЕХАНИЗМ РАБОТЫ КОМПЛЕКСА III(убихинол-цитохром с- оксидоредуктаза)
Ингибитор:
антимицин А
21. КОМПЛЕКС IV
Катализирует реакцию окислениявосстановленного цитохрома с молекулярным
кислородом, сопряженную с трансмембранным
переносом 2-х протонов в межмембранное
пространство.
Переход электронов здесь осуществляется от
атома железа в цитохроме на атом меди.
На этом комплексе происходит присоединение
4-х электронов к молекуле кислорода, что
приводит к полному его восстановлению с
образованием эндогенной воды
22. КОМПЛЕКС IV (ЦИТОХРОМОКСИДАЗА)
23. Комплекс V – F0-F1-АТФ-синтаза
Это трансмембранный ферментныйкомплекс, состящий из двух
компонентов – факторы F0 –F1:
1-й отвечает за транспорт протонов
из межмембранного пространства
внутрь,
2-й – за синтез АТФ (окислительное
фосфорилирование)
24. КОМПЛЕКС V (FoF1-ATP СИНТАЗА)
25. ТРАНСПОРТ СУБСТРАТОВ ДЛЯ СИНТЕЗА АТР
26. Электрохимический потенциал
компоненты дыхательной цепи, расположенные вовнутренней митохондриальной мембране, в ходе
переноса электронов могут «захватывать» протоны
из матрикса митохондрий и передавать их в
межмембранное пространство. При этом наружная
поверхность внутренней мембраны приобретает
положительный заряд, а внутренняя –
отрицательный, т.е. создаётся градиент
концентрации протонов с более кислым значением
рН снаружи. Так возникает трансмембранный
потенциал (ΔµН+). Существует три участка
дыхательной цепи, на которых он образуется. Эти
участки соответствуют I, III и IV комплексам цепи
переноса электронов
27.
Электрохимический потенциал протонов ∆μН+∆ψ-электрический
потенциал
∆рН - химический
(концентрационный)
градиент
Энергия
Синтез АТФ Теплопродукция
Транспорт
веществ через
мембрану
митохондрий
28. Коэффициент окислительного фосфорилирования
Степень сопряжённости окисления ифосфорилирования в митохондриях
характеризует коэффициент
фосфорилирования (Р/О). Это
количественная оценка синтеза АТФ.
Он равен отношению количества молекул
неорганического фосфата (Н3РО4),
перешедшего в АТФ, к количеству атомов
потреблённого кислорода (О2).
29.
Мера эффективности работы дыхательной цепи- коэффициент Р/О
Р - количество потребленного Н3РО4,
(эквивалентно количеству синтезированного
АТФ)
О - количество потребленного кислорода
Для НАДН - Р/О максимально 3
Для ФАДН2 - Р/О максимально 2
Для аскорбата - Р/О максимально 1
30. Основные пути использования энергии АТФ:
1) синтез сложных химических веществиз более простых (реакции анаболизма);
2) сокращение мышц (механическая
работа);
3) образование трансмембранных
биопотенциалов;
4) активный транспорт веществ через
биологические мембраны.
31. ИНГИБИТОРЫ ДЫХАТЕЛЬНОЙ ЦЕПИ
32.
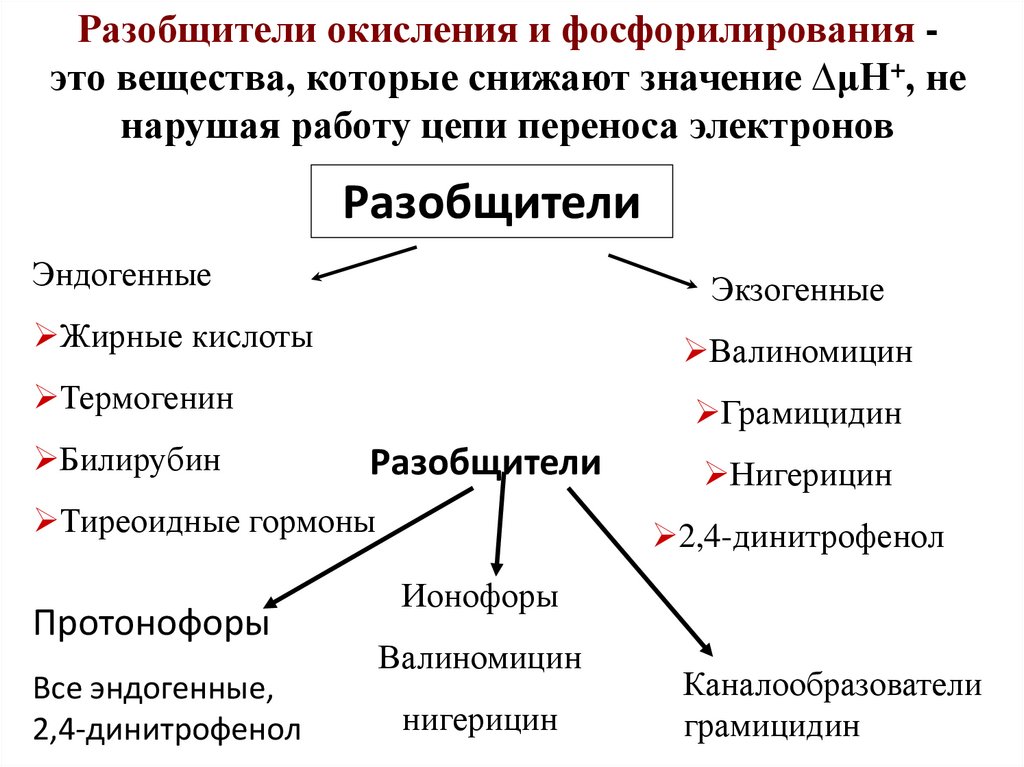
Разобщители окисления и фосфорилирования это вещества, которые снижают значение ∆μН+, ненарушая работу цепи переноса электронов
Разобщители
Эндогенные
Экзогенные
Жирные кислоты
Валиномицин
Термогенин
Билирубин
Грамицидин
Разобщители
Тиреоидные гормоны
Протонофоры
Все эндогенные,
2,4-динитрофенол
Нигерицин
2,4-динитрофенол
Ионофоры
Валиномицин
нигерицин
Каналообразователи
грамицидин
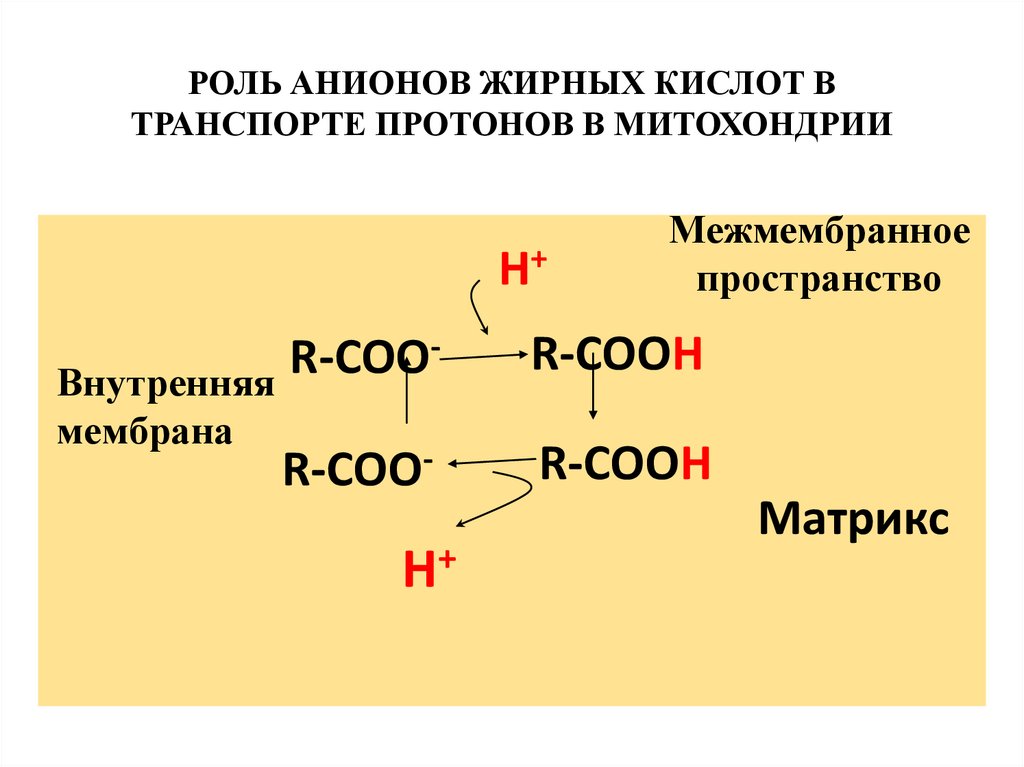
33. Протонофоры
Протонофоры представляют собой слабыегидрофобные органические кислоты,
которые в форме аниона (R-COO-)
связывают протоны в межмембранном
пространстве, диффундируют через
мембрану и диссоциируют в матриксе с
образованием протонов. К этой группе
относятся, например, свободные жирные
кислоты, гормоны щитовидной железы,
салицилаты, дикумарол, 2,4-динитрофенол
34.
РОЛЬ АНИОНОВ ЖИРНЫХ КИСЛОТ ВТРАНСПОРТЕ ПРОТОНОВ В МИТОХОНДРИИ
H+
R-COO
Внутренняя
мембрана
R-COO-
H+
Межмембранное
пространство
R-COOH
R-COOH
Матрикс
35. Термогенин
разобщающий белок, обнаруженныйв митохондриях адипоцитов бурой жировой
ткани.
Используется в качестве основного
механизма продукции тепла у новорожденных и
у млекопитающих, впавших в спячку.
Они увеличивают проницаемость внутренней
митохондриальной мембраны, позволяя
протонам, перенесенным в межмембранное
пространство, возвращаться в
митохондриальный матрикс.
Производство тепла при помощи термогенина в
бурой жировой ткани происходит с
разобщением клеточного дыхания
36. СХЕМА ДЕЙСТВИЯ ТЕРМОГЕНИНА
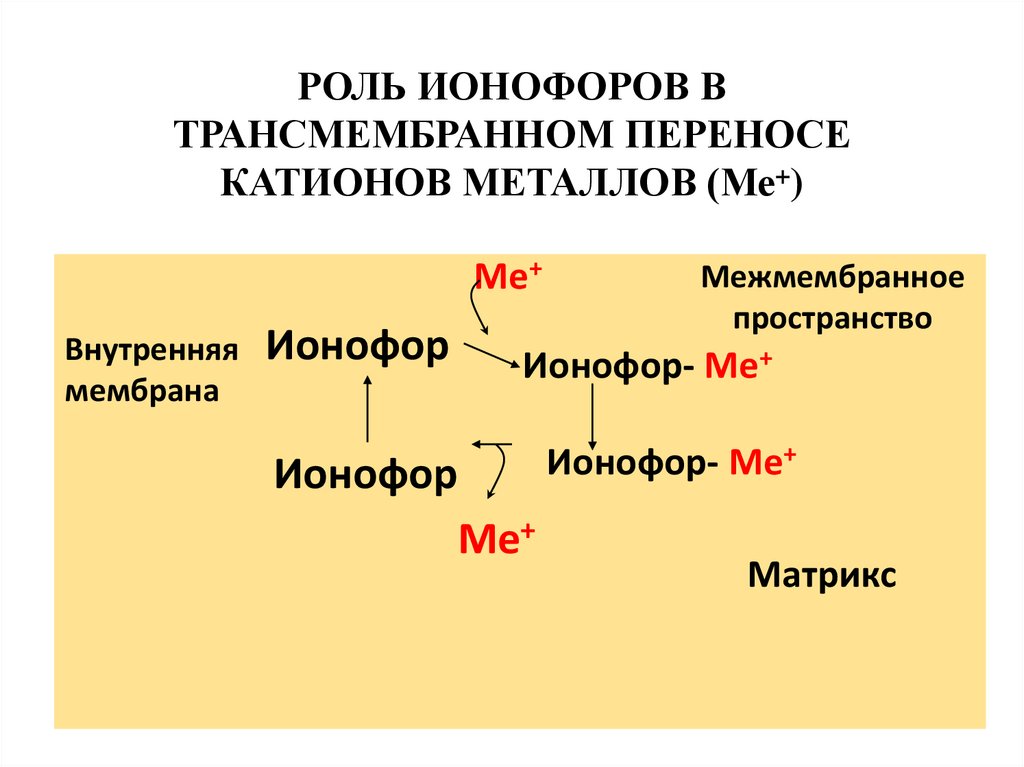
37. Ионофоры
Ионофоры (валиномицин, нигерицин,грамицидин) способны встраиваться в
мембрану, образуя канал, по которому
могут перемещаться протоны и другие
одновалентные катионы – Na+ или K+.
В результате снижается протонный
потенциал, что приводит к уменьшению
электрохимического потенциала и
нарушается синтез АТФ.
38.
РОЛЬ ИОНОФОРОВ ВТРАНСМЕМБРАННОМ ПЕРЕНОСЕ
КАТИОНОВ МЕТАЛЛОВ (Ме+)
Ме+
Внутренняя
мембрана
Ионофор
Межмембранное
пространство
Ионофор- Ме+
Ионофор- Ме+
Ионофор
Ме+
Матрикс
39. ПРИНЦИП ДЕЙСТВИЯ ИОНОФОРОВ-КАНАЛООБРАЗОВАТЕЛЕЙ
ПРИНЦИП ДЕЙСТВИЯ ИОНОФОРОВКАНАЛООБРАЗОВАТЕЛЕЙН+, К+, Na+
Мембрана
Н+, К+, Na+
40. Врожденные нарушения дыхательной цепи
• Дефицит цитохрома с : подостраянекротическая энцефалопатия (синдром Ли).
Развиваются симметричные некрозы в
головном и спинном мозге. Это ведет к
атаксии, судорогам, задержке умственного
развития и гипотензии.
• Дефицит цитохромоксидазы – проявляется
миопатией (слабость мышц, утомляемость,
судороги, дыхательная недостаточность)
41. Приобретенные нарушения дыхательной цепи
Индуцированный паркинсонизм –снижение общей двигательной активности,
тремор рук, нарушение координации
движений. Патология возможна при
применении наркотика меперидина,
который ингибирует комплекс I, подобно
ротенону. Нарушается окислительное
фосфорилирование, снижается уровень
АТФ в клетках черной субстанции мозга
(развивается паркинсонизм)
42.
При клеточных поврежденияхтормозится поступление кислорода в
клетку – возникает гипоксия.
Тормозится процесс ЦТК и
поступление восстановленных
коферментов в ЦТД, нарушается
синтез АТФ.
Подобный эффект возможен при
авитаминозе витаминов В2, РР,
дефиците железа и меди.