















 НАДН - КоQ-редуктаза ( НАДН-дегилрогеназа); 2) сукцинат – КоQ-редуктаза (сукцинатдегидрогеназа); 3) КоQ-ци")










 Биология
БиологияПохожие презентации:









")
Биоэнергетика. Тканевое дыхание и окислительное фосфорилирование
1. Биоэнергетика Тканевое дыхание и окислительное фосфорилирование
• Лекция 3Биоэнергетика
Тканевое дыхание и окислительное фосфорилирование
2. Разнообразие метаболизма
По источнику энергии:• Фототрофы
–
воспринимают
энергию Солнца и накапливают
ее
в
виде
химических
соединений
• ( белков, жиров, углеводов).
• Хемотрофы
–
энергию
путем
химических
фототрофов
извлекают
окисления
соединений
3. 2-ой закон Термодинамики
системы стремятся к неупорядоченности- при необратимых процессах энтропия системы
и окружающей среды увеличивается
ЭНТРОПИЯ (S) от греч. entropia -- поворот, превращение
4. Энтропия
мера беспорядка (случайности) в системе (S)Энтропия
растет!
5. ВЗАИМОСВЯЗЬ ОБМЕНА ВЕЩЕСТВ И ЭНЕРГИИ.
Пищевые веществаМетаболиты
Катаболизм
Энергия
Образование конечных продуктов
обмена (углекислого газа и воды,
мочевина)
Анаболизм
Синтез структурно-функциональных компонентов клетки
Функциональная
активность
Энергия ?????
6. АТФ
Karl Lohmann (1898-1978) –Fritz Albert Lipmann (1899-1986)
“ATP is energy-transfering molecule
in the cell” (1941)
the discoverer of ATP (1929)
7. АТФ-энергетическая «валюта» клетки
Макроэргическиесвязи
АТФ → АДФ + Ф + 30,5 кДЖ/моль
8.
Взрослый человекпотребляет примерно
60 кг АТФ в день.
Во взрослом организме
содержится около 50 г АТФ
???
Цикл АТФ - АДФ
АТФ ↔ АДФ + Ф
Реакция фосфорилирования
9. АТФ образуется в клетке двумя путями: - окислительное фосфорилирование: синтез АТФ с использованием энергии выделяемой при окислении S:
Q – это энергия окисления субстрата- субстратное фосфорилирование: синтез АТФ
с использованием энергии S ( макроэрга)
Макроэргические соединения имеют
макроэргические связи, т. е. связи, при гидролитическом
расщеплении которых высвобождается более 30 кДж/моль
(или 7 ккал/моль) энергии.
10. Макроэрги
- нуклеозидтрифосфаты,- нуклеозиддифосфаты,
- креатинфосфат,
- 1,3-дифосфоглицерат,
- ацетил-КоА
- фосфоенолпируват и
др.
11.
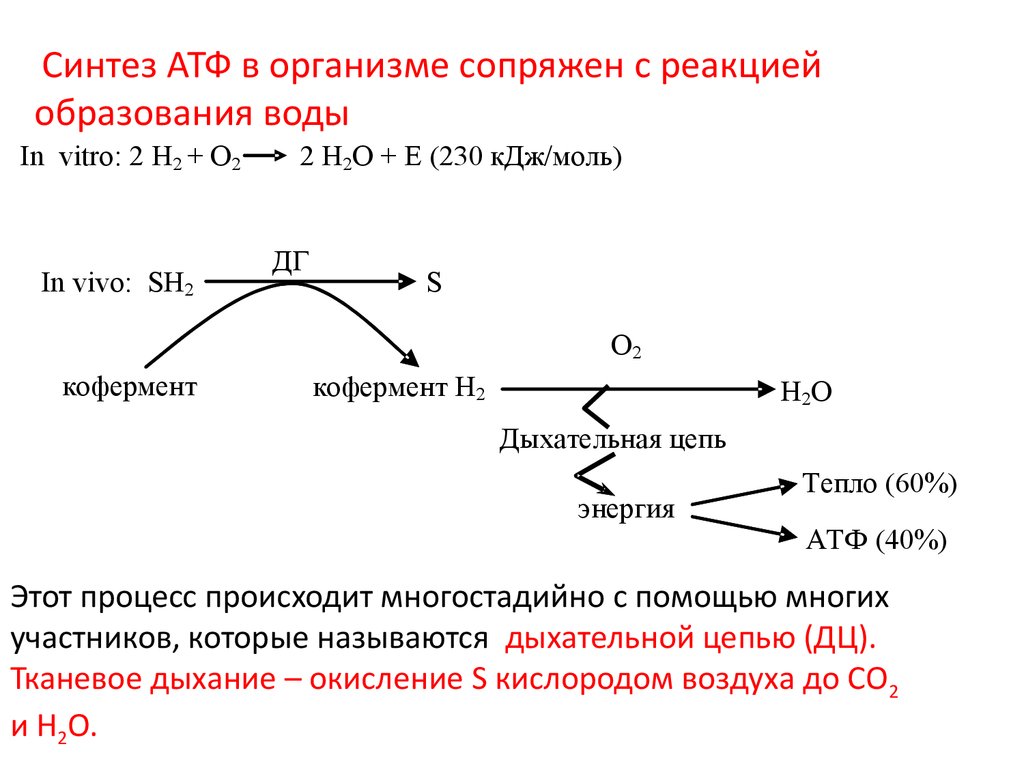
Синтез АТФ в организме сопряжен с реакциейобразования воды
In vitro: 2 Н2 + O2
In vivo: SH2
2 H2О + Е (230 кДж/моль)
ДГ
S
О2
кофермент
кофермент Н2
Н2О
Дыхательная цепь
энергия
Тепло (60%)
АТФ (40%)
Этот процесс происходит многостадийно с помощью многих
участников, которые называются дыхательной цепью (ДЦ).
Тканевое дыхание – окисление S кислородом воздуха до СО2
и Н2О.
12. Общий путь катаболизма
13.
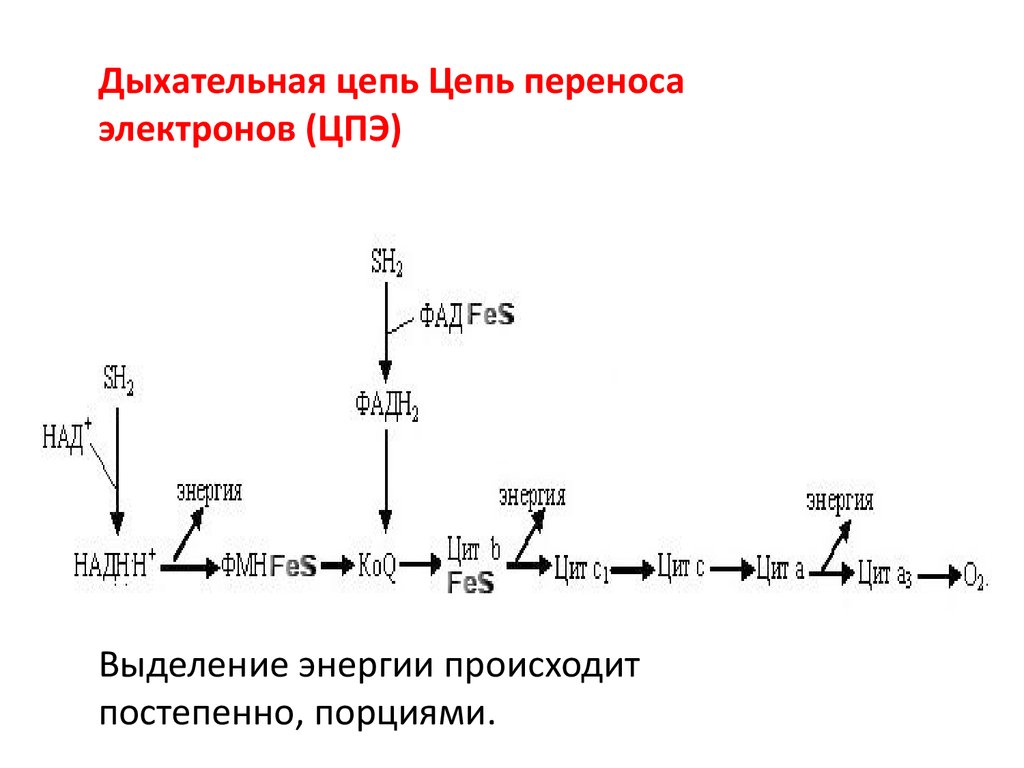
• Дыхательная цепь (цепь переноса электронов,ЦПЭ) – последовательность
оксидоредуктаз,транспортирующих Н+ и е- от
S на О2.
• ЭТЦ позволяет запасти энергию,
выделяющуюся в ходе окисления НАДН2 и
ФАДН2.
• Высвобожденная энергия используется на
синтез макроэргических связей АТФ или тепло.
14. Ферменты ДЦ локализованы на внутренней мембране митохондриях
15. Компоненты ДЦ Пиридиновые ДГ
16. Флавиновые ДГ
17. Компоненты ДЦ
• Коензим Q (убихинон)R
R
• Цитохромы b, c1 , c , a, a3
Цит. (Fe3+ ) +
е →
Цит. ( Fe2+ )
•железосерные белки (FeS-белки); цитохромы и железосерные
белки переносят только электроны. Железосерные белки
ассоциированы с ФМН, ФАД и цитохромом b.
18.
Дыхательная цепь Цепь переносаэлектронов (ЦПЭ)
Выделение энергии происходит
постепенно, порциями.
19. ДЦ состоит из 5 ферментных комплесов: 1) НАДН - КоQ-редуктаза ( НАДН-дегилрогеназа); 2) сукцинат – КоQ-редуктаза (сукцинатдегидрогеназа); 3) КоQ-ци
ДЦ состоит из 5 ферментных комплесов:1) НАДН - КоQ-редуктаза ( НАДН-дегилрогеназа);
2) сукцинат – КоQ-редуктаза (сукцинатдегидрогеназа);
3) КоQ-цитохром c-редуктаза ( QH2-дегидрогеназа) ;
4) цитохром а –оксидаза;
5) АТФ-синтаза (или протонная АТФ-аза (Н +- АТФ-аза)
и 2 подвижных переносчиков: убихинона и цит.С
20.
Принцип работы дыхательной цепи: разделение потоковпротонов и электронов, поступающих из матрикса.
Электроны передаются на конечный акцептор — кислород;
протоны
выбрасываются
в
митохондриальное
межмембранное пространство (ММП).
21. Дыхательные ферменты расположены в порядке ↑ величины редокс - потенциала
Место того или иного переносчика в ДЦопределяется величиной редокс-потенциала
Дыхательные ферменты расположены в порядке
↑ величины редокс - потенциала
22.
Редокс-потенциал (Ео )численно равен электрондвижущей силе в Вольтах, возникающей
между растворами окислителя и восстановителя (концентрации 1М,
рН = 7,0, температура 25 С).
Чем отрицательнее Ео системы, тем выше ее
способность отдавать электроны (восстановители).
Чем положительнее редокс-потенциал, тем выше
способность вещества присоединять электроны
(окислители).
23.
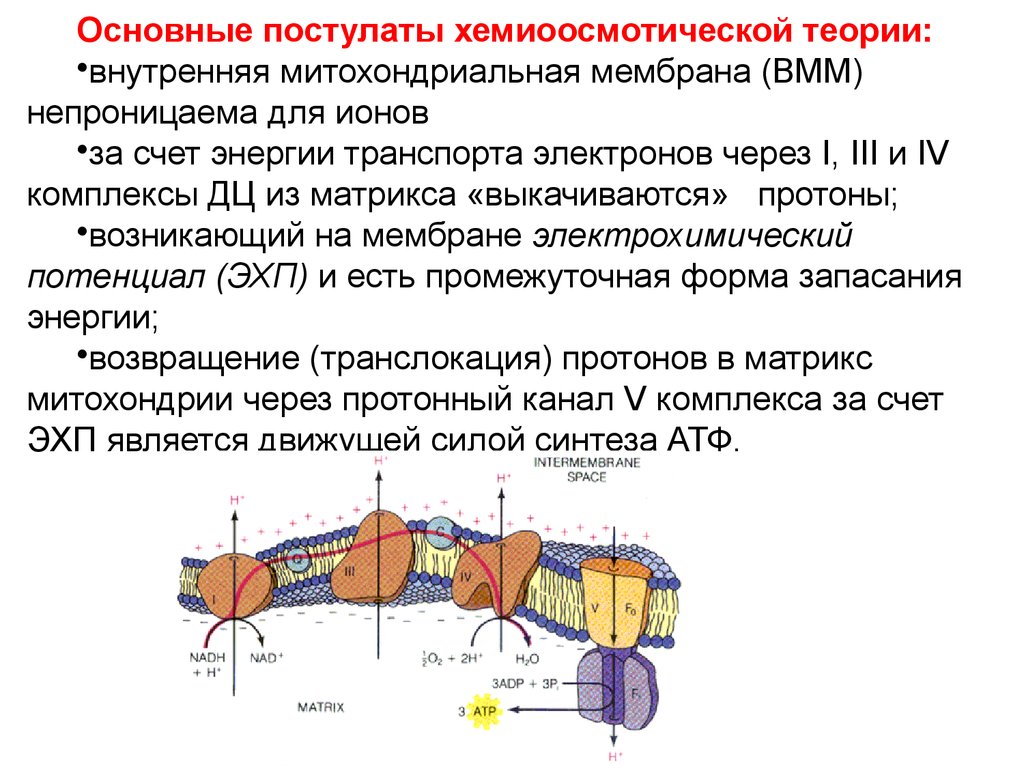
Хемиоосмотическая теория окислительногофосфорилирования (Нобелевская премия 1978 г.).
24.
Основные постулаты хемиоосмотической теории:•внутренняя митохондриальная мембрана (ВММ)
непроницаема для ионов
•за счет энергии транспорта электронов через I, III и IV
комплексы ДЦ из матрикса «выкачиваются» протоны;
•возникающий на мембране электрохимический
потенциал (ЭХП) и есть промежуточная форма запасания
энергии;
•возвращение (транслокация) протонов в матрикс
митохондрии через протонный канал V комплекса за счет
ЭХП является движущей силой синтеза АТФ.
25. Связь между транспортом электронов в ДЦ и синтезом АТФ: протонная АТФ -синтаза
Дальнейшие исследования (Дж.Уокер, П. Бойер, Нобелевская
премия 1997 г.) подтвердили
предположения Митчелла.
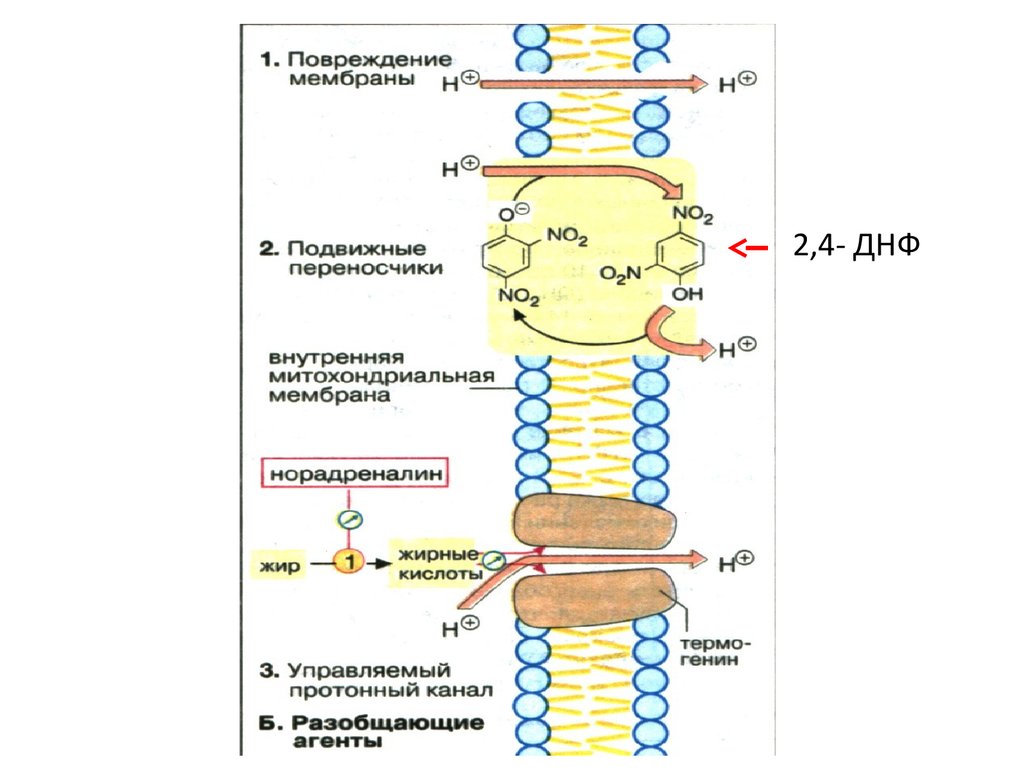
26. Энергетический эффект биологического окисления
Пункты сопряжения тканевого дыхания иокислительного фосфорилирования.
Синтез 1 моль АТФ ≈ 32 кДж/моль .
Ео не менее 0,26 В.
I
III
IV
27. Дыхательный контроль Регулируется скорость работы дыхательной цепи энергетическим зарядом клетки, т. е. соотношением АТФ/АДФ.
• Эффективность окислительногофосфорилирования оценивают по
коэффициенту фосфорилирования Р/О:
количество молекул Ф, использованных для
синтеза АТФ, в расчете на один атом
поглощенного кислорода.
Р/О ≤ 3
Для НАД-зависимых S - P/O =3;
Для ФАД зависимых S - Р/О = 2
Дыхательный контроль
Регулируется скорость работы дыхательной
цепи энергетическим зарядом клетки, т. е.
соотношением АТФ/АДФ.