



")















")










































 Биология
БиологияПохожие презентации:










Биологическое окисление. Окислительное фосфорилирование
1.
Лекция: Биологическоеокисление. Окислительное
фосфорилирование.
2. ФИЗИОЛОГИЯ ДЫХАНИЯ
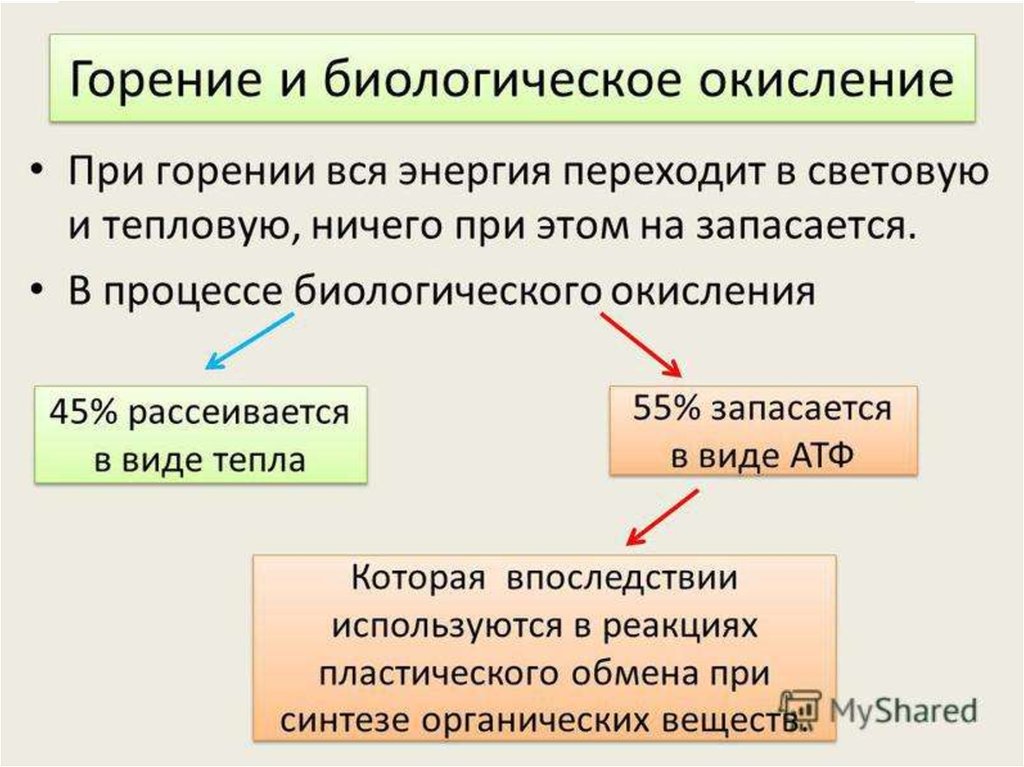
3. Биологическое окисление
- Этосовокупность
окислительновосстановительных реакций,
сопровождающихся передачей
протонов и электронов.
4. ОСНОВНАЯ РОЛЬ:
Обеспечитьорганизм
энергией в доступной для
использования форме
(АТФ).
5. Аденозинтрифосфорная кислота (АТФ)
6.
7.
8.
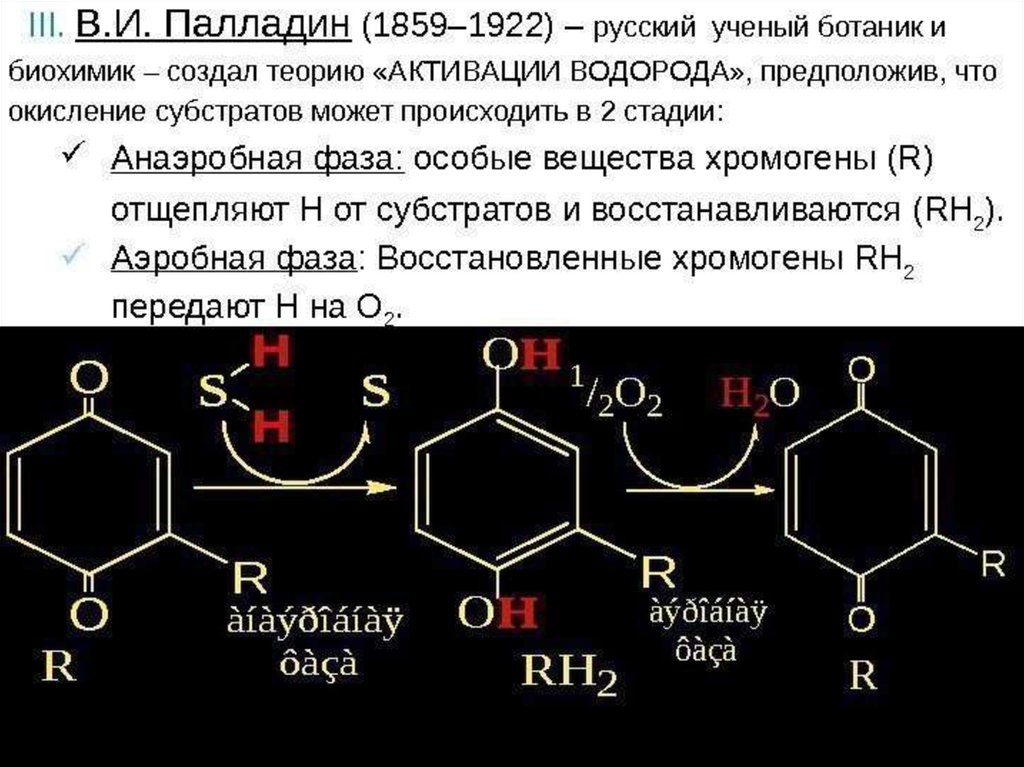
9. Тканевое дыхание
ВдыхаемыйО2
используется для синтеза
Н2О за счет Н2
окисляемых S:
SH2 + ½ O2 ==> S + H2O
10.
11.
12.
Переносчики электронов ипротонов образуют цепь
переноса электронов (ЦПЭ)
или дыхательную цепь
(ДЦ).
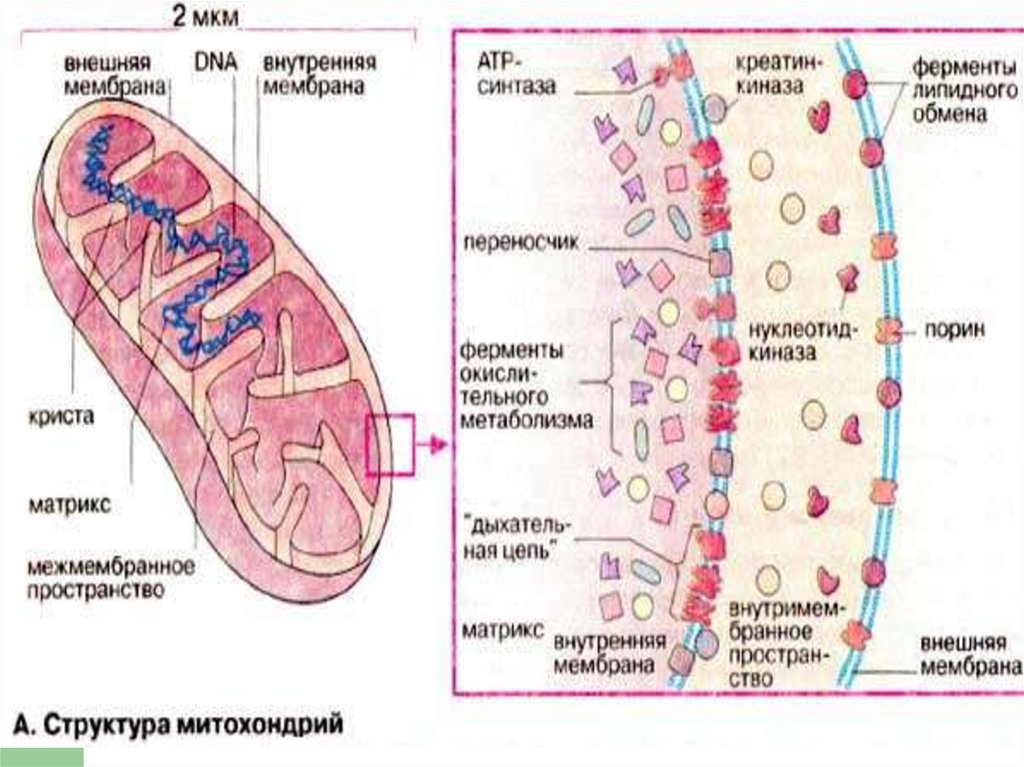
13. ЦПЭ и МИТОХОНДРИИ
Всеферменты ЦПЭ связаны с
МХ.
1 гепатоцит содержит около
1000 МХ.
14.
15. ФЕРМЕНТЫ НАРУЖНОЙ МЕМБРАНЫ
МоноаминооксидазаСистема
удлинения цепи
жирных кислот
Фосфолипаза А
16. ФЕРМЕНТЫ ВНУТР.МЕМБРАНЫ
НАДН-дегидрогеназаСукцинатдегидрогеназа
Цитохромы
в,с1, с, а, а3
Н+-АТФ-синтаза
АДФ-АТФ-транслоказа
17. ФЕРМЕНТЫ МАТРИКСА
ФерментыЦТК (кроме
сукцинатдегидрогеназы)
Ферменты бета-окисления
ЖК
Глутаматдегидрогеназа
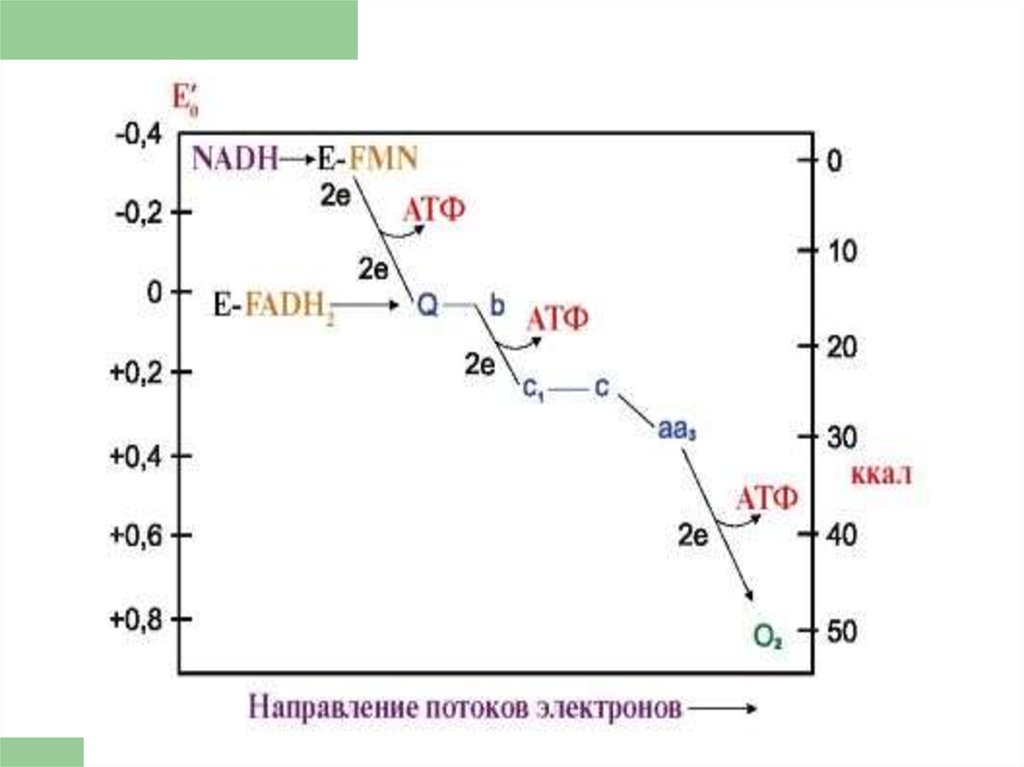
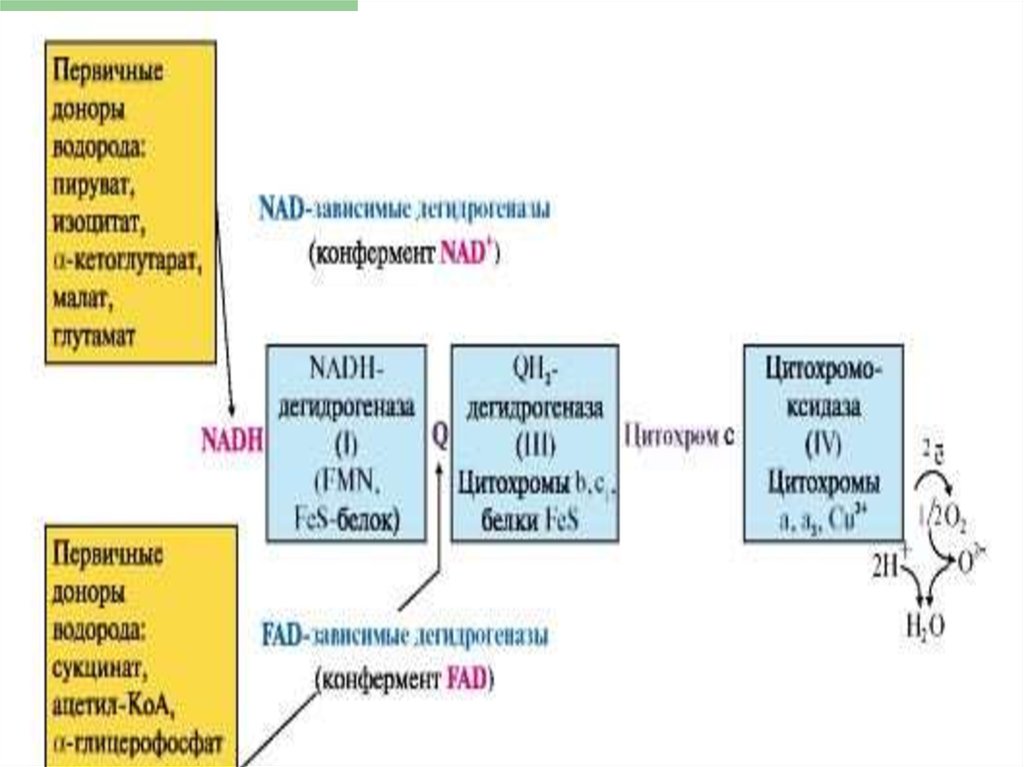
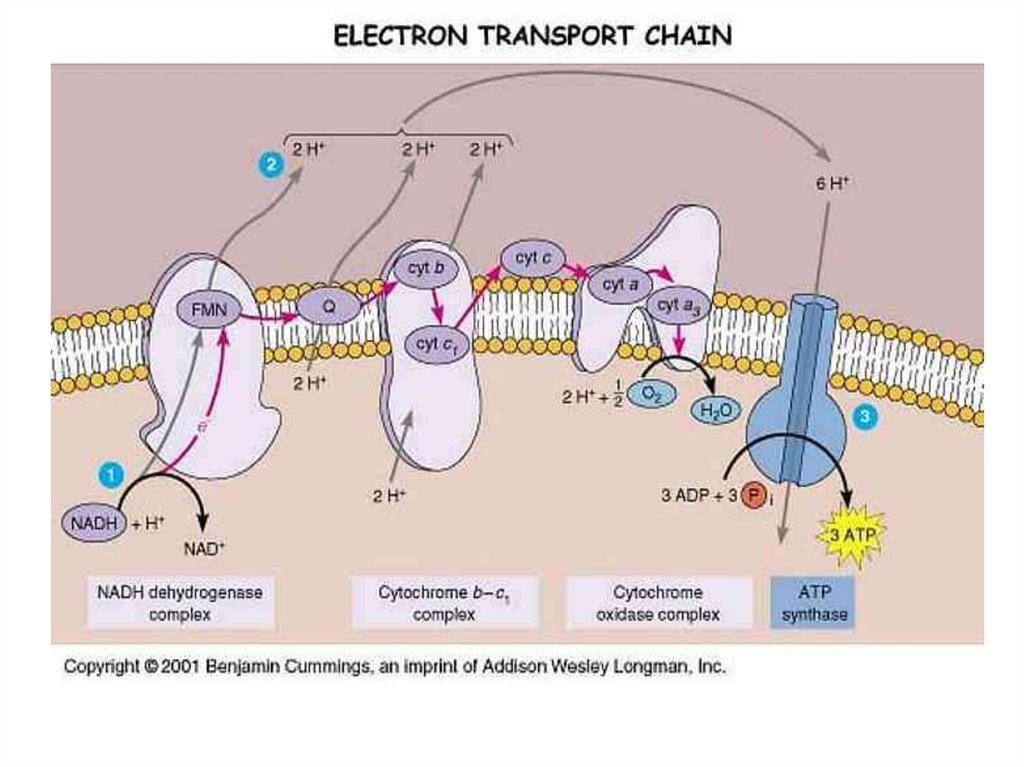
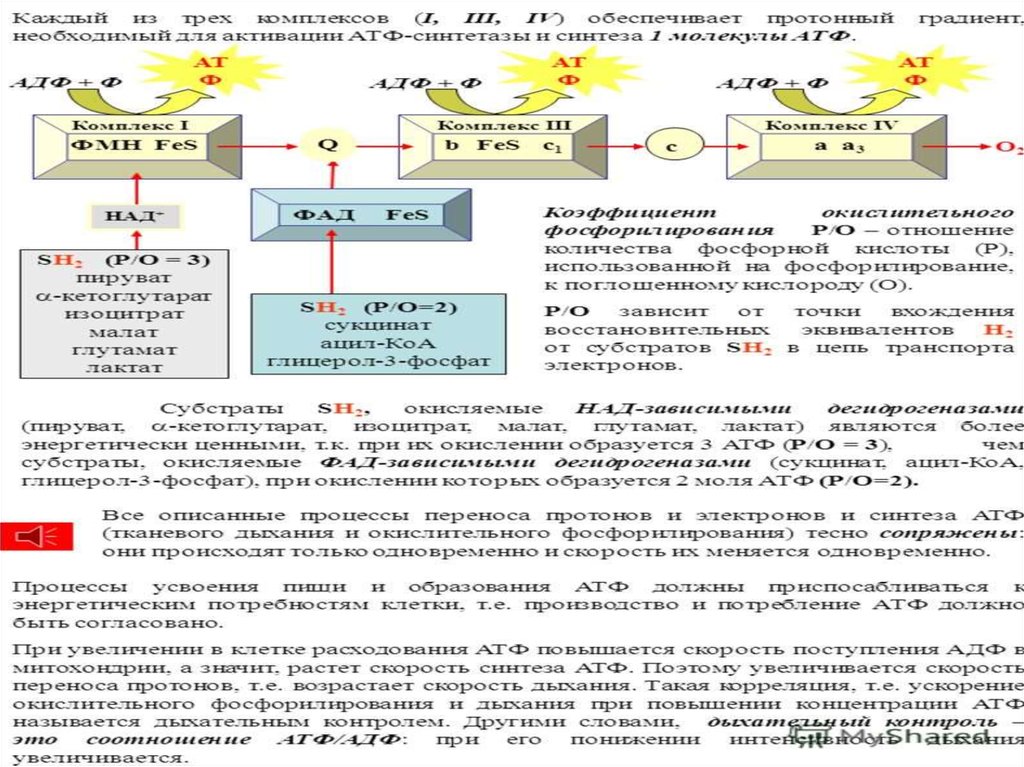
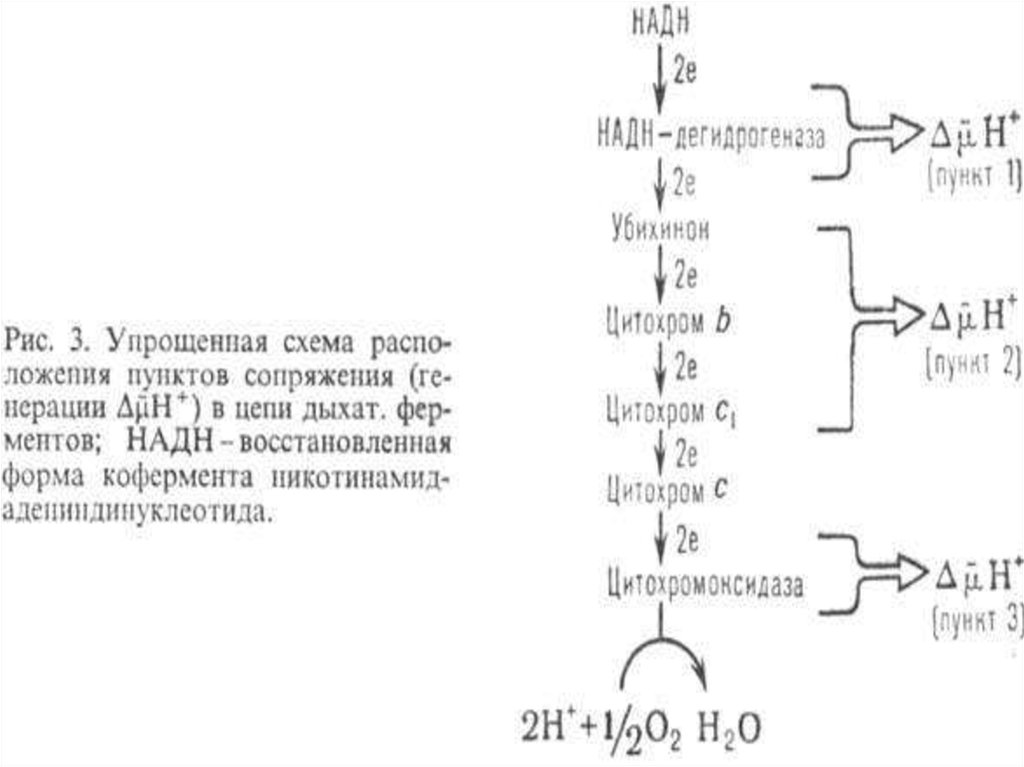
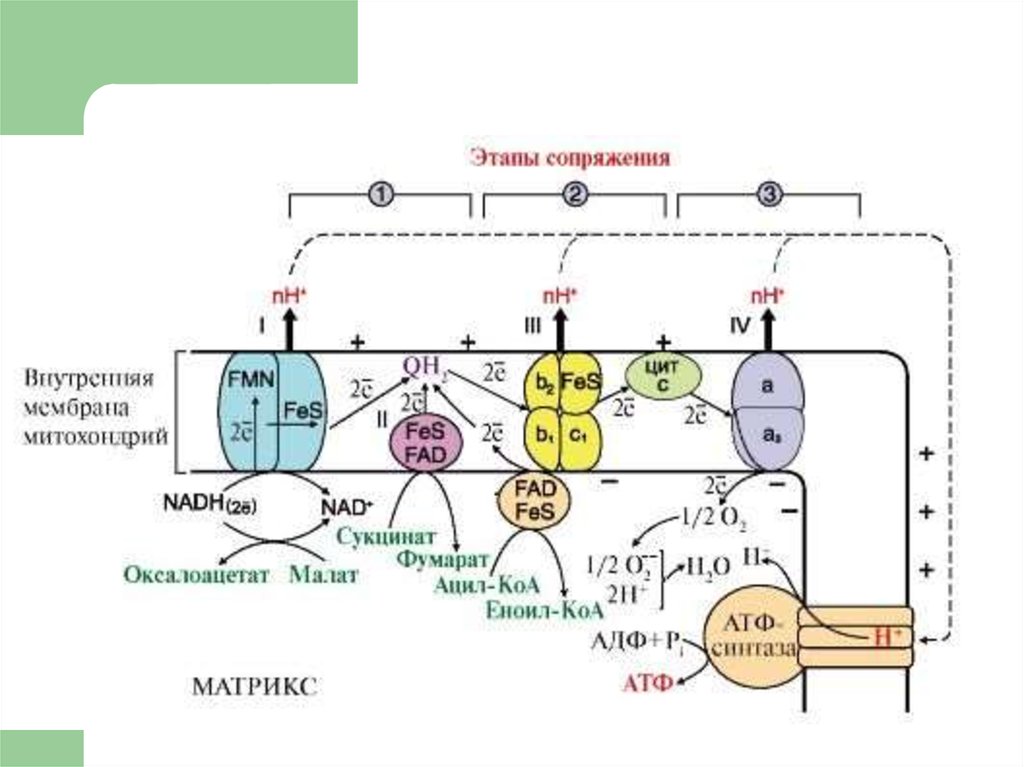
18. ЦПЭ
* НАД-зависимых ДЕГИДРОГЕНАЗ,ФАД-зависимых ДЕГИДРОГЕНАЗ,
УБИХИНОНА,
ЦИТОХРОМОВ.
Последовательность
расположения определяется
их редокс-потенциалами.
19.
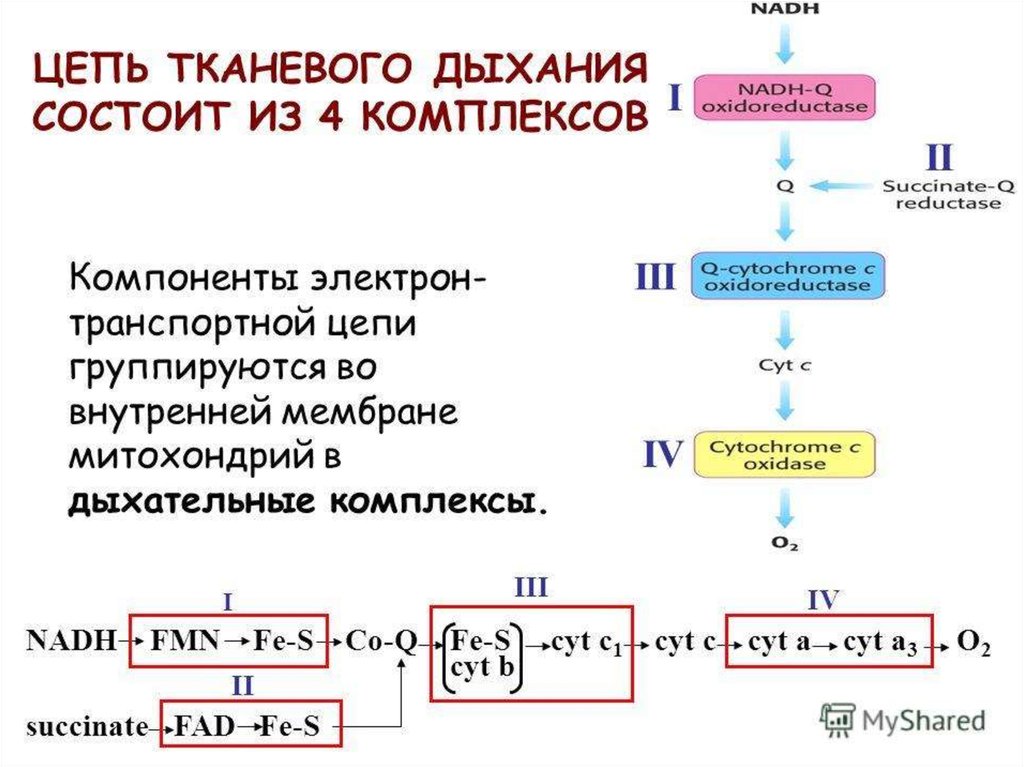
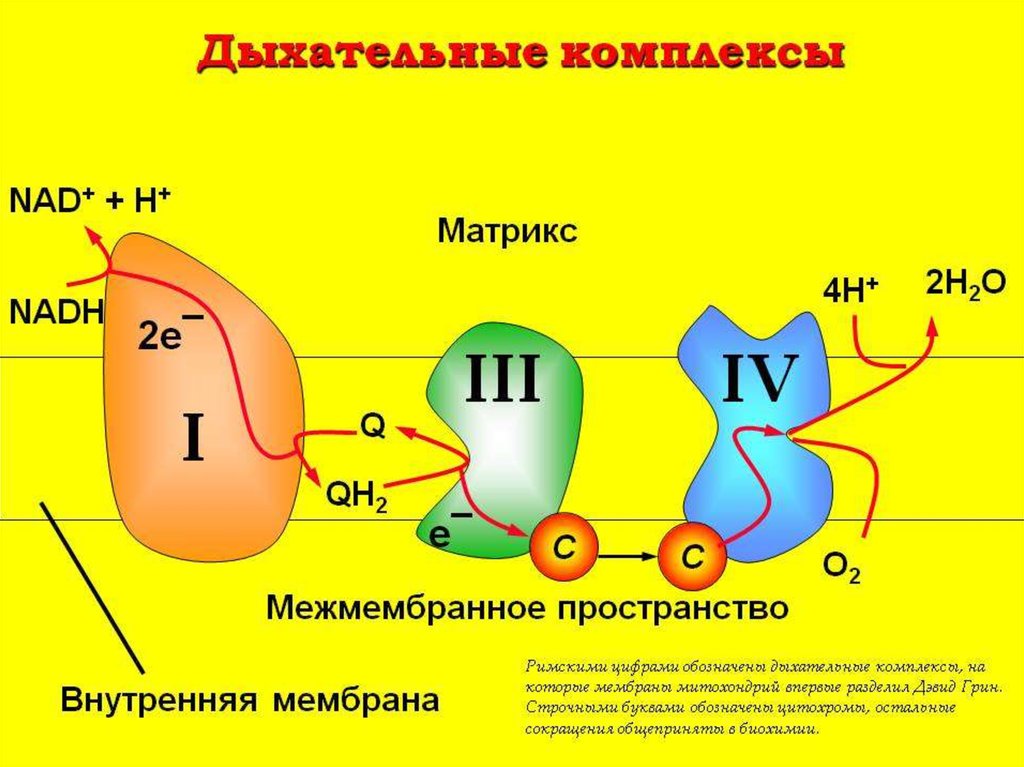
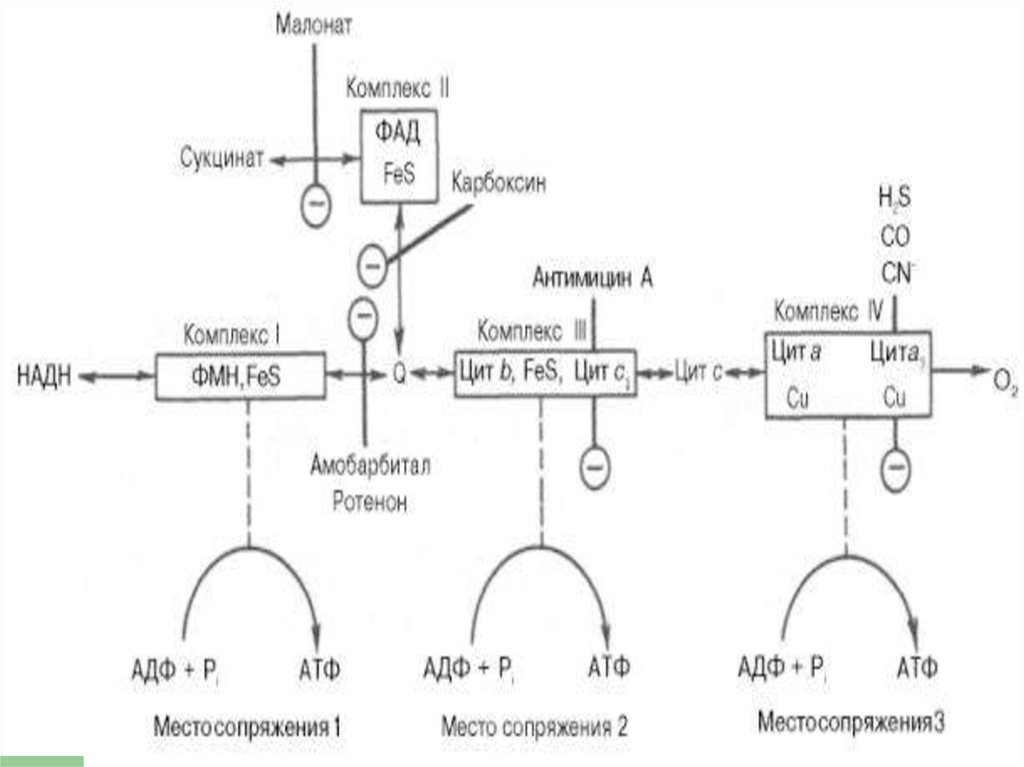
20. Комплексы цепи переноса электронов
ЦПЭсостоит из
комплексов, каждый из них
осуществляет
определенную стадию ЦПЭ.
21.
22.
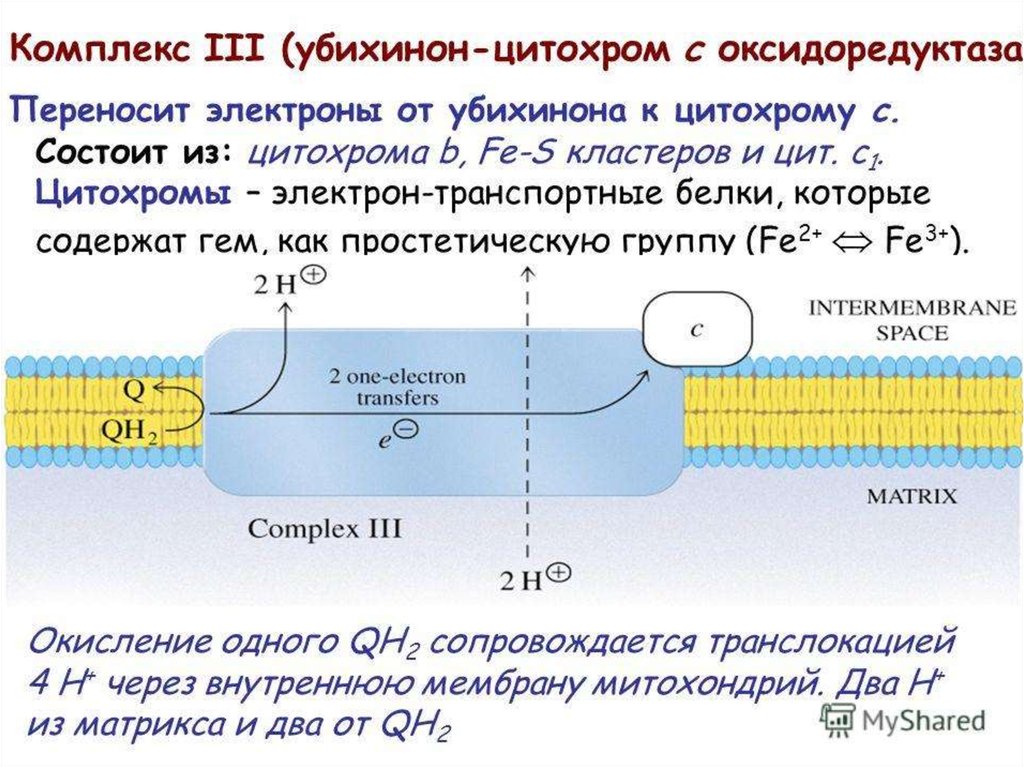
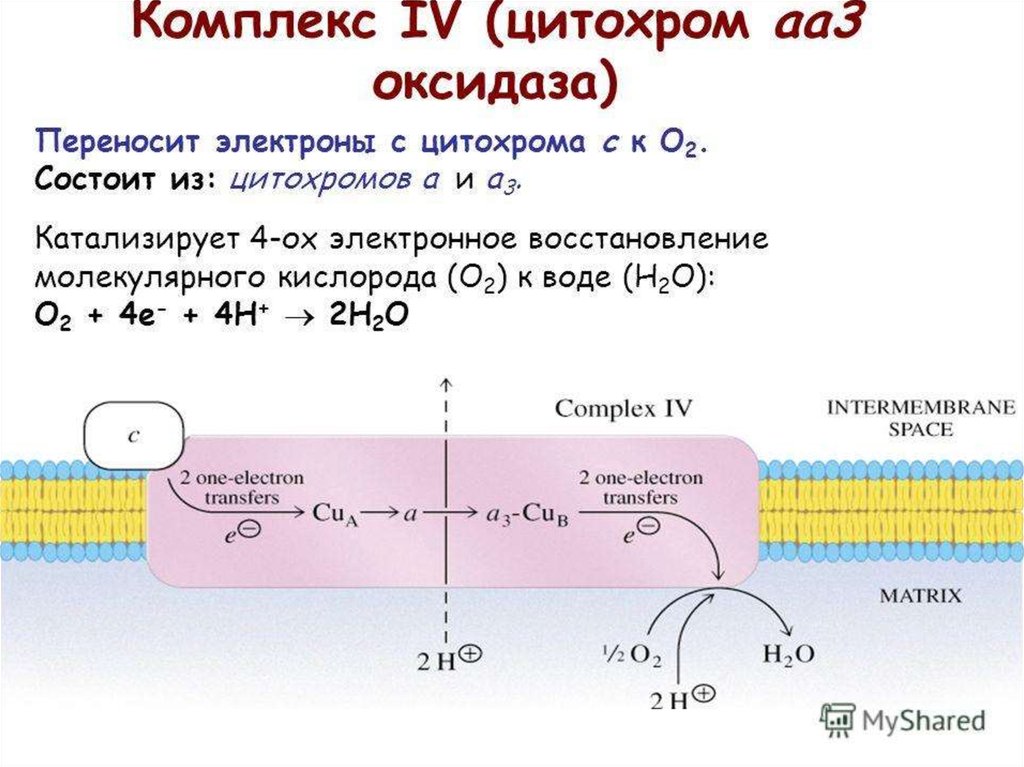
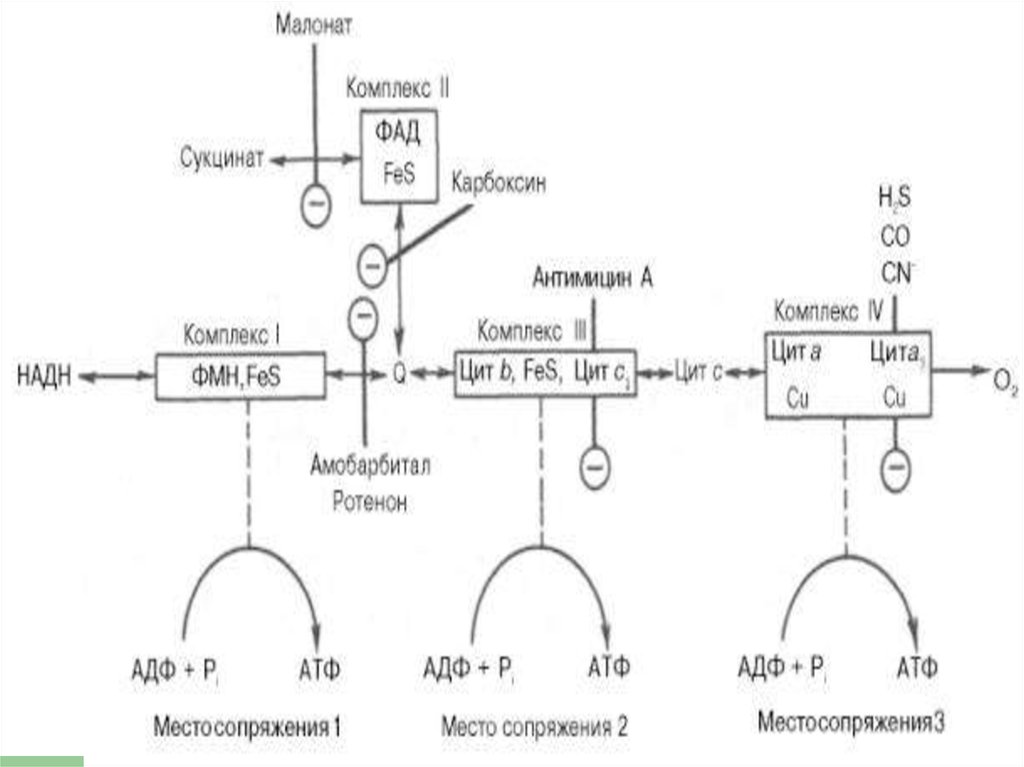
23. Комплексы
Обозначают комплексы I, II, III и IV.I - НАДН-дегидрогеназа
II - Сукцинатдегидрогеназа
III - Убихинол-цитохром средуктаза
IV - Цитохромоксидаза
24.
25.
26.
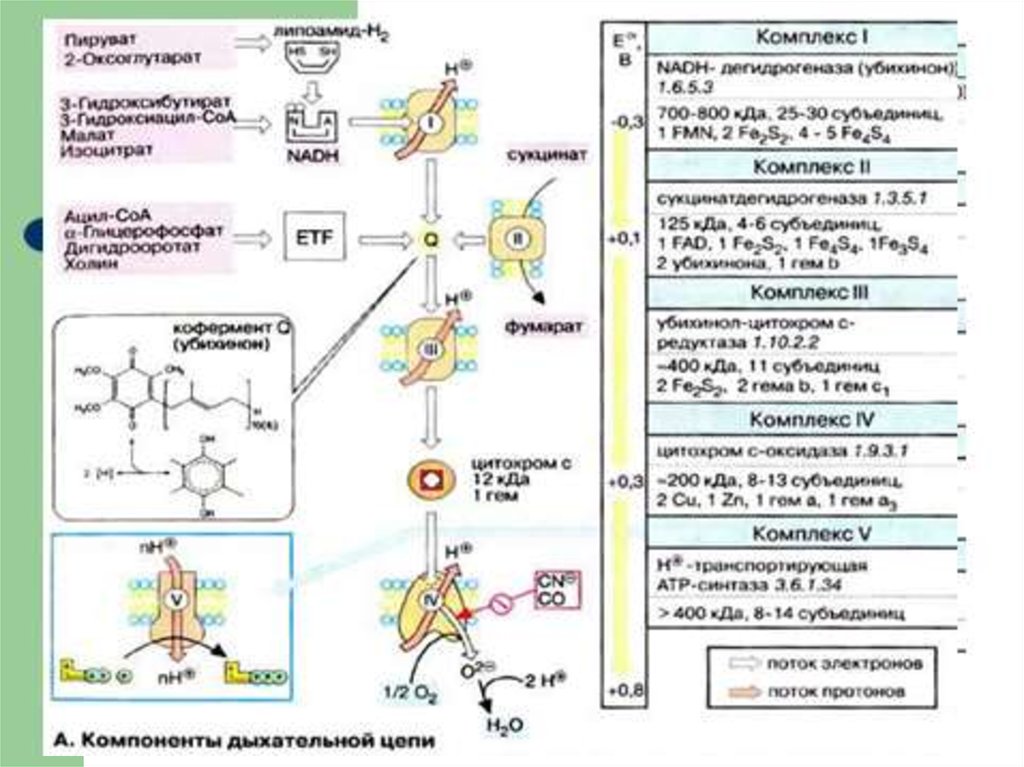
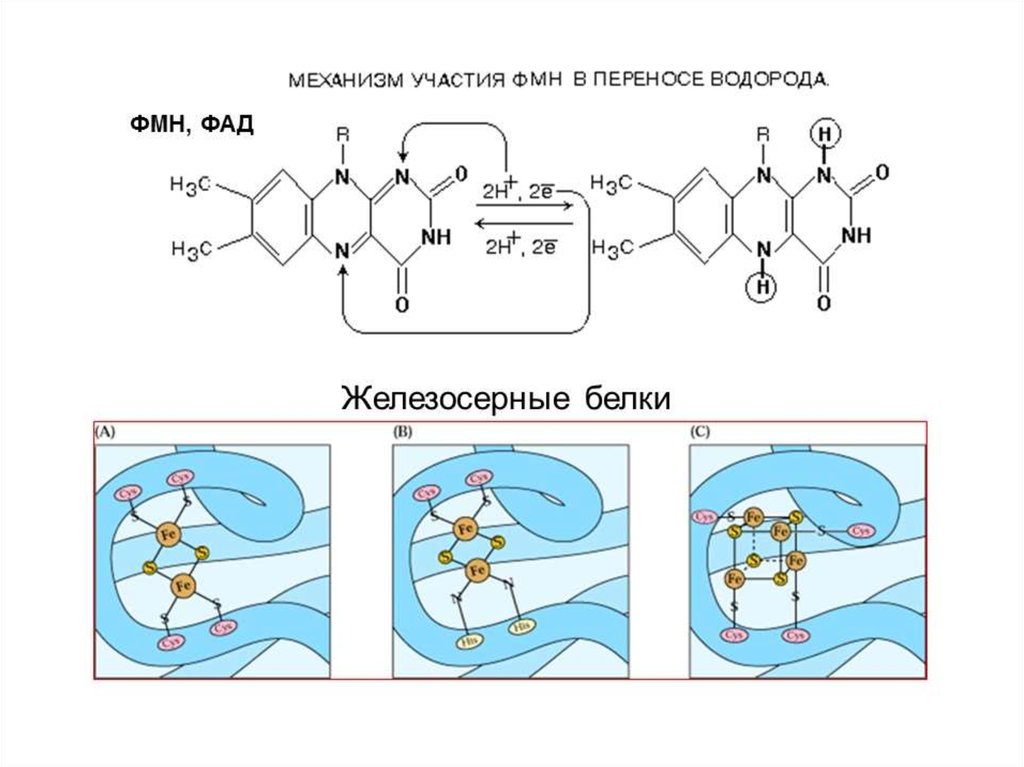
Комплексы ДЦ построены из белков иокислительно-восстановительных
коферментов ( ФМН и ФАД, в
комплексах I и II), железо-серные центры
(в I, II и III) и группы гема (в II, III и IV).
27.
28.
Электроны поступают в ДЦразличными путями.
При окислении НАДН + Н+
комплекс I переносит электроны
через ФМН и Fe/S-центры на
убихинон.
29.
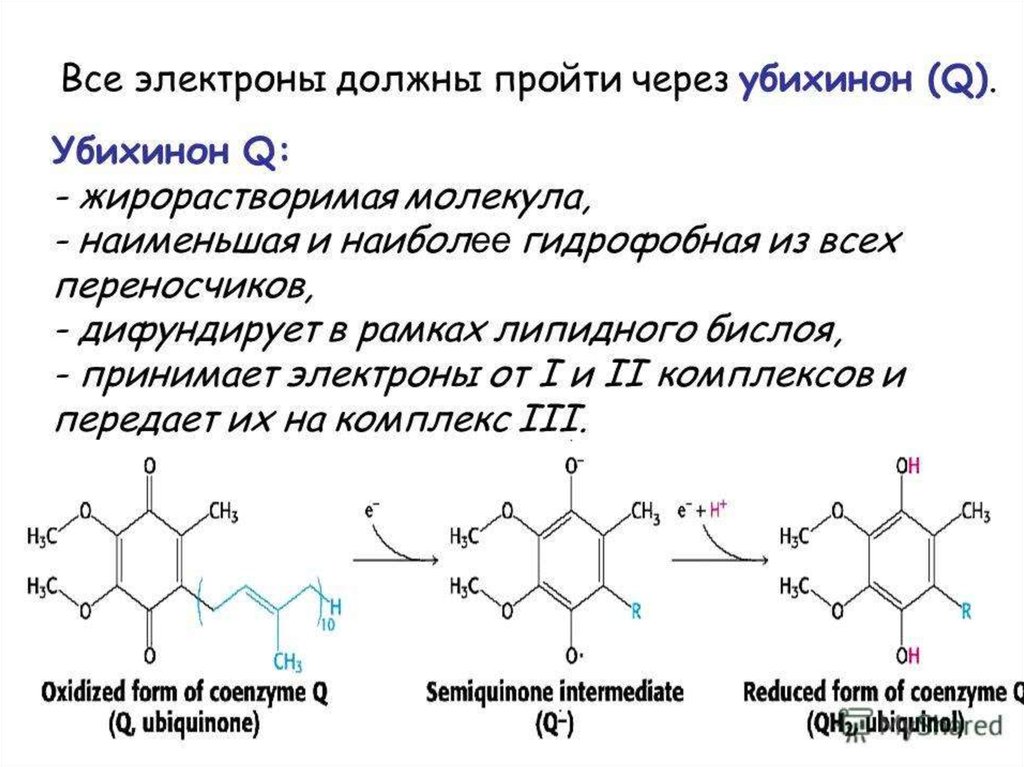
Образующиеся при окислениисукцината, ацил-КоА электроны
переносятся на убихинон комплексом II
через ФАДН2.
При этом окисленная форма кофермента
Q восстанавливается в QH2.
30.
31. Строение убихинона (коэнзима Q)
OCH3O
OH
CH3
CH3O
CH3
CH3
CH3O
(CH2–CH=C–CH2)10H
O
Хиноидная форма
CH3
CH3O
(CH2–CH=C–CH2)10H
OH
Гидрохинонная форма
32. Восстановленный коэнзим Q
переносит электроны вкомплекс III, который
поставляет на цитохром
с. Последний переносит
электроны к комплексу IV,
цитохромоксидазе.
QH2
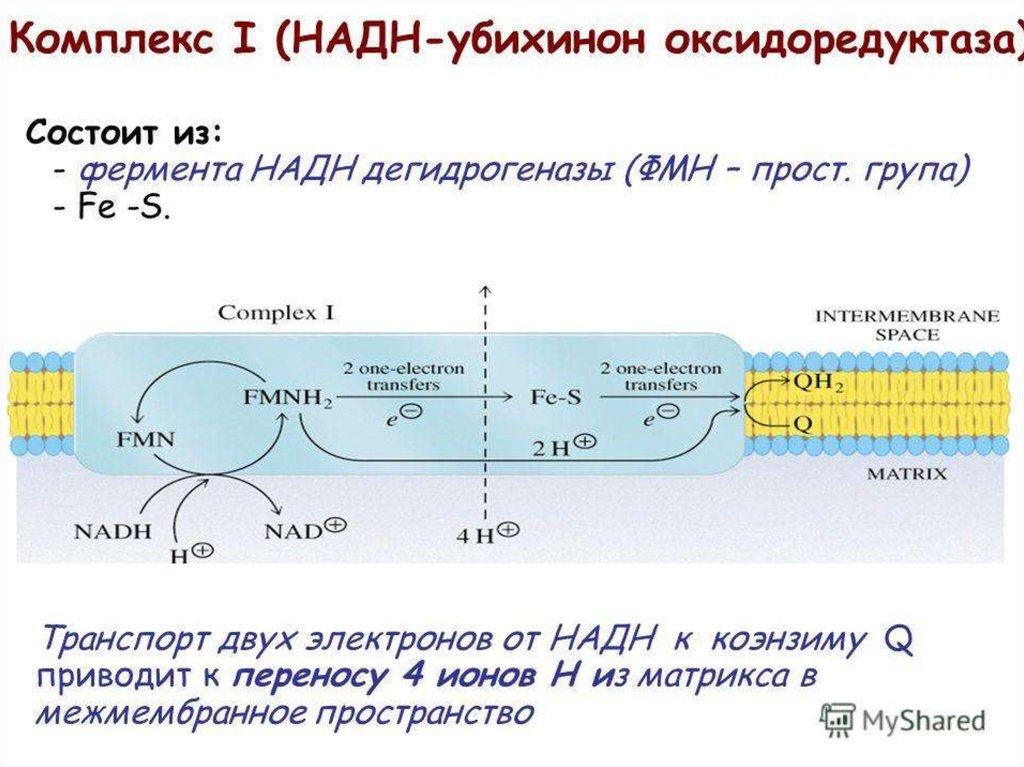
33. Комплекс I
КомплексI катализирует
окисление NADH
переносчиком электронов —
убихиноном (коферментом Q).
34.
35. НАД
+НАД
36. НАДН.Н
+НАДН.Н
37. Убихинон
существует в окисленной ивосстановленной формах CoQ и CoQH2:
38. Восстановленный коэнзим Q
переносит электроны вкомплекс III, который
поставляет на цитохром
с. Последний переносит
электроны к комплексу IV,
цитохромоксидазе.
QH2
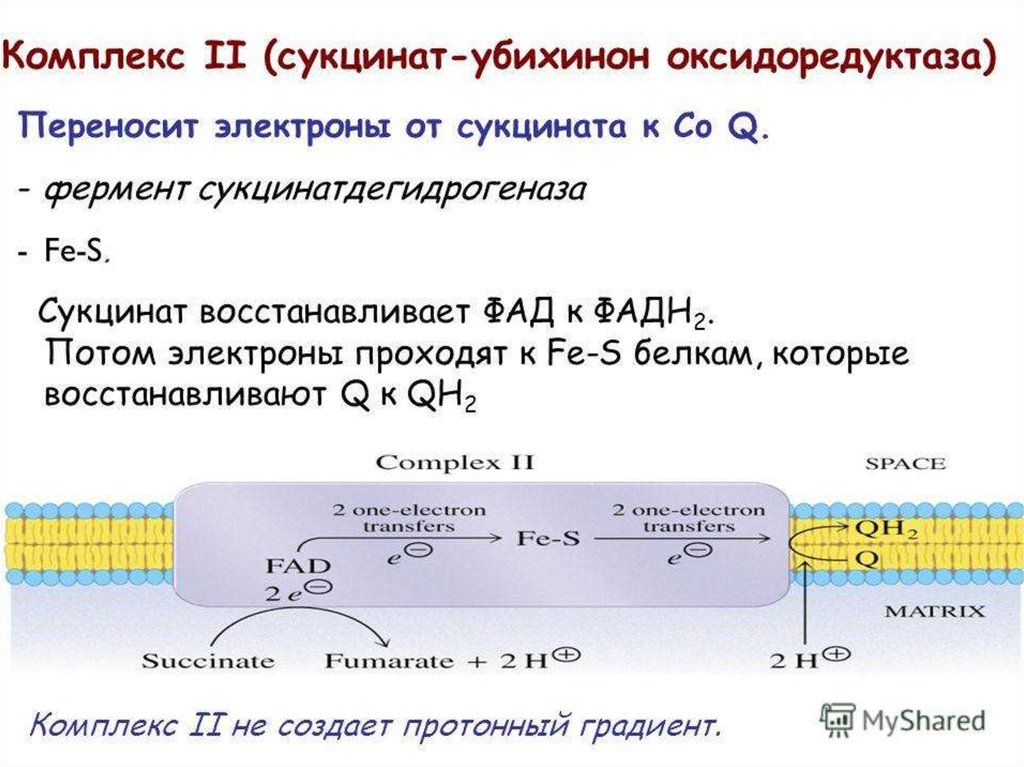
39. Комплекс II
• Суммарный процесс, катализ. комплексом INADH
+
H
+
CoQ
NAD
+
CoQH2
• Комплекс II катализирует окисление
сукцината в фумарат по реакции:
OOC–CH2–CH2–COO
Cукцинат
+
OOC
CoQ
H
C C
H
Фумарат
COO
+
CoQH2
40.
41. ФАД
42.
43. Комплекс III
Комплекс III катализирует окислениеQН2 цитохромом с.
Уравнение реакции:
CoQH2
2e
+
2Fec(III)
2Fec(II)
+
2H
+
CoQ
44.
45.
46.
47. Комплекс IV
Комплекс IV - цитохромоксидаза,катализирует перенос электронов
от цитохрома с непосредственно
на кислород по реакции:
2e
2Fec(II)
+
1/2O2
+
2H
2Fec(III)
+
H2O
48.
При восстановлении О2образуется сильный
основной анион О2-,
который связывает два
протона и переходит в
воду.
49.
50. Дыхательная цепь
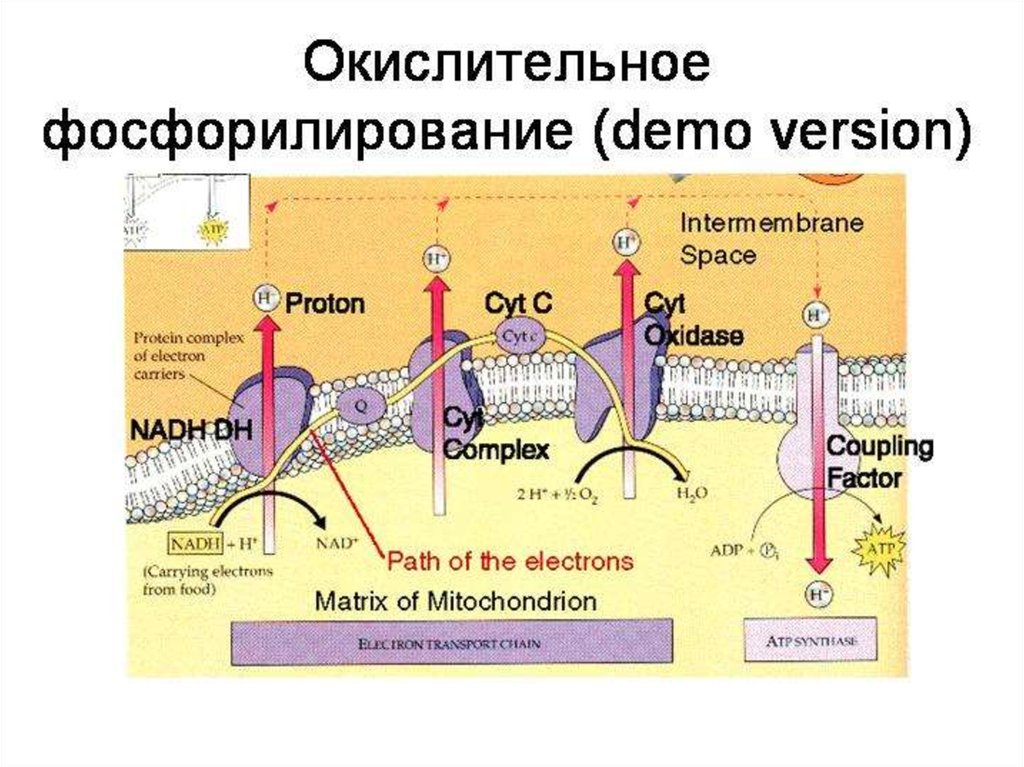
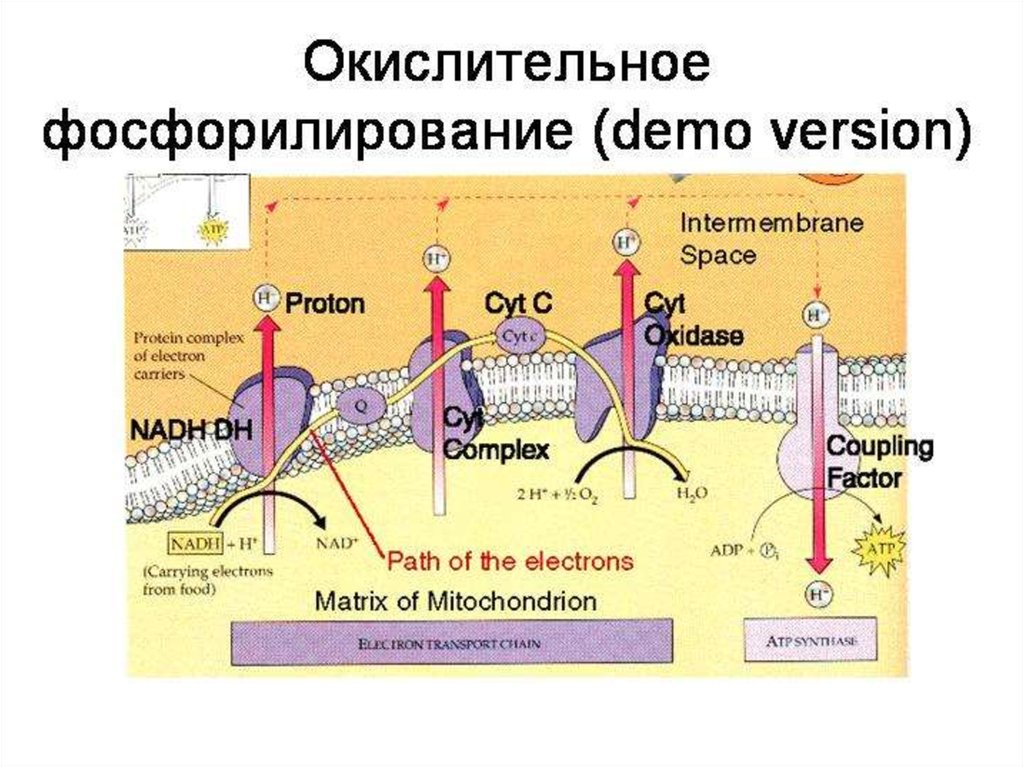
51.
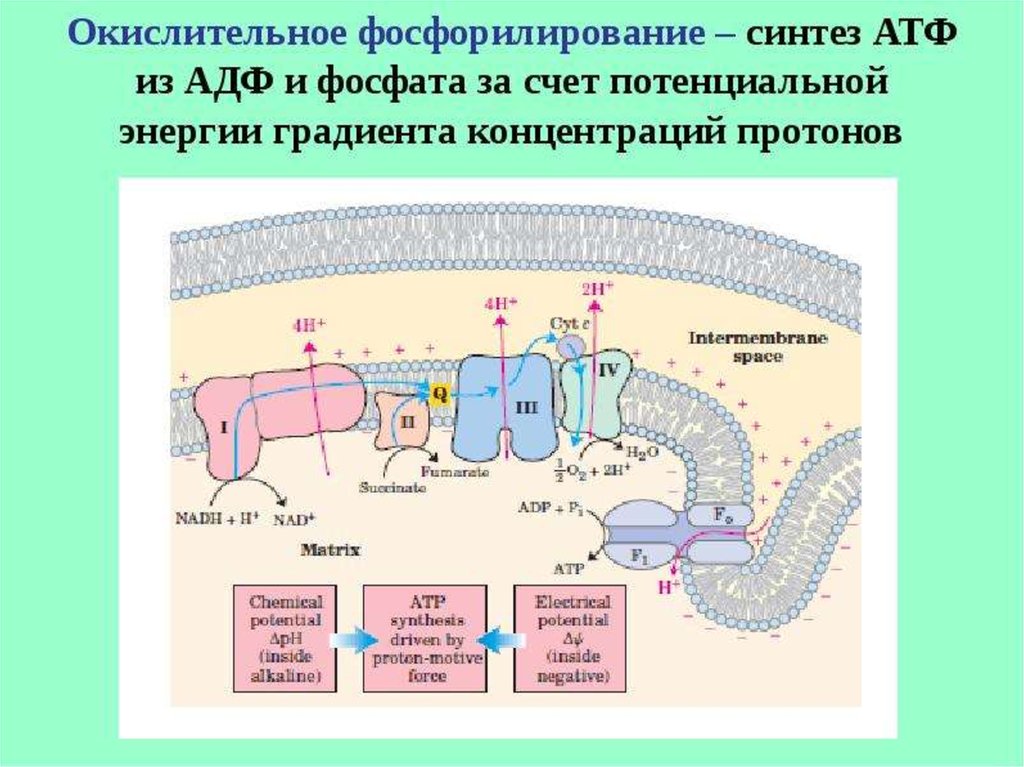
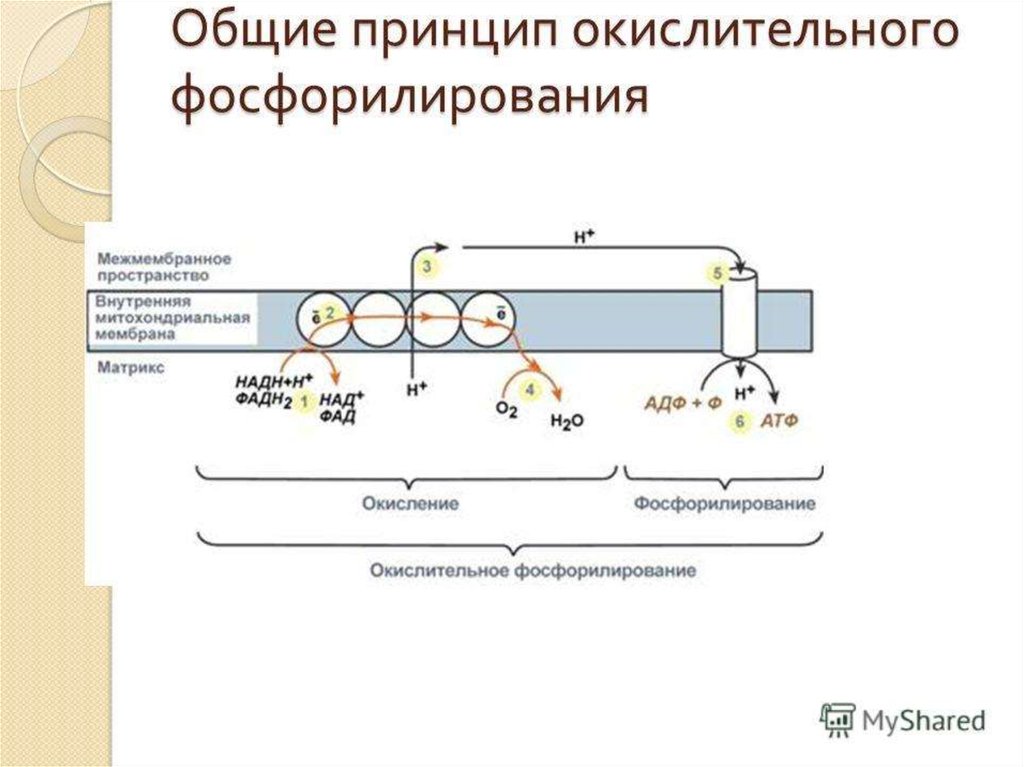
ОКИСЛИТЕЛЬНОЕФОСФОРИЛИРОВАНИЕ
52.
53.
54. Окислительное фосфорилирование
55.
56. Окислительное фосфорилирование
СинтезАТФ из АДФ и Н3РО4 за
счет энергии окисления
веществ, связанный с
переносом электронов по
ЦПЭ наз. ОКИСЛИТЕЛЬНЫМ
ФОСФОРИЛИРОВАНИЕМ.
57.
58.
59.
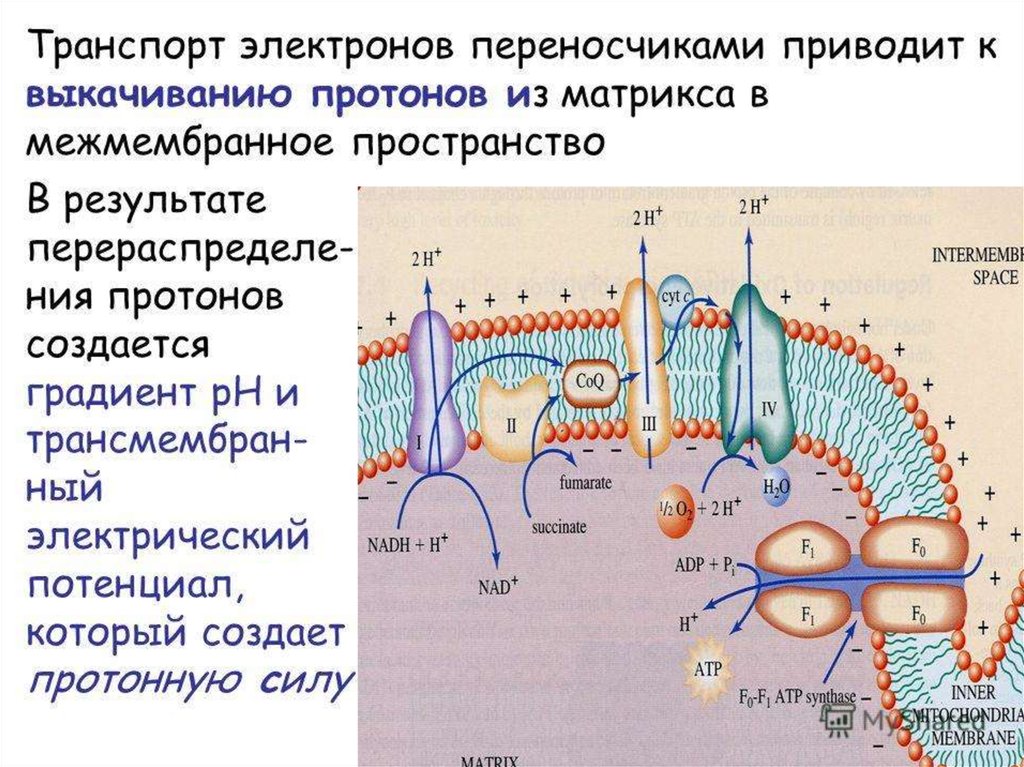
Поток электронов сопряжен собразованием комплексами I,
III и IV протонного градиента.
60.
Большая частьвыделяющейся энергии
используется для создания
градиента протонов и,
наконец, для образования АТФ
с помощью АТФ-синтазы.
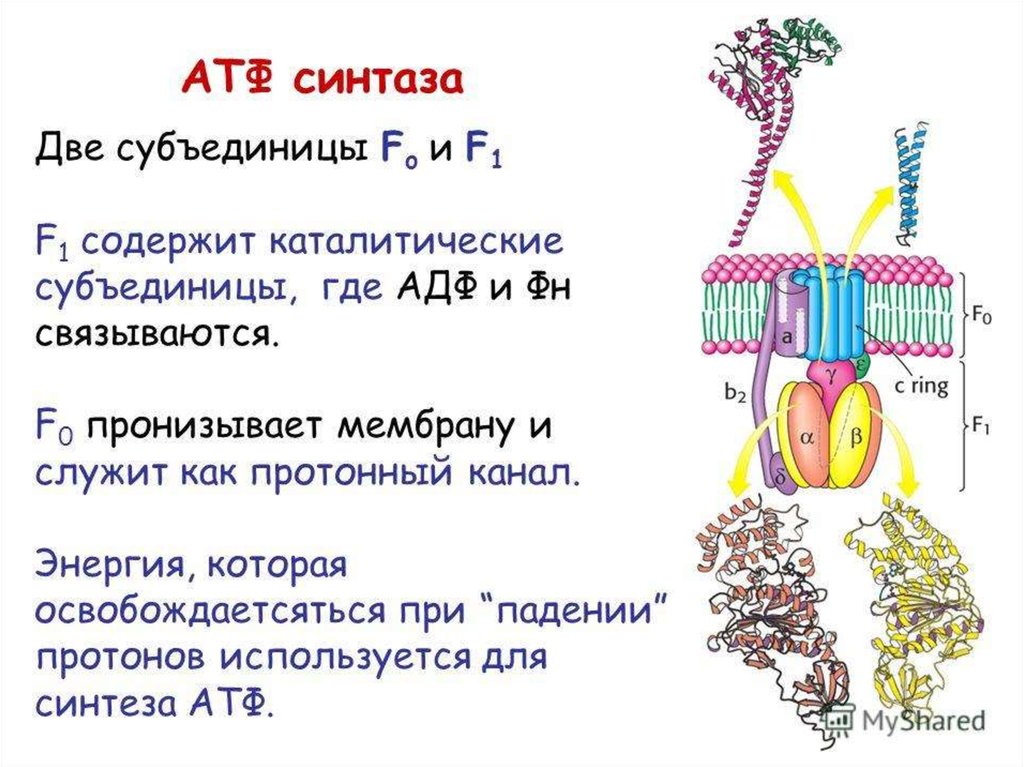
61. АТФсинтаза
Главная функция ферментазаключается в
фосфорилировании AДФ
ортофосфатом с
образованием АТФ.
62. Механизм сопряжения фосфорилирования
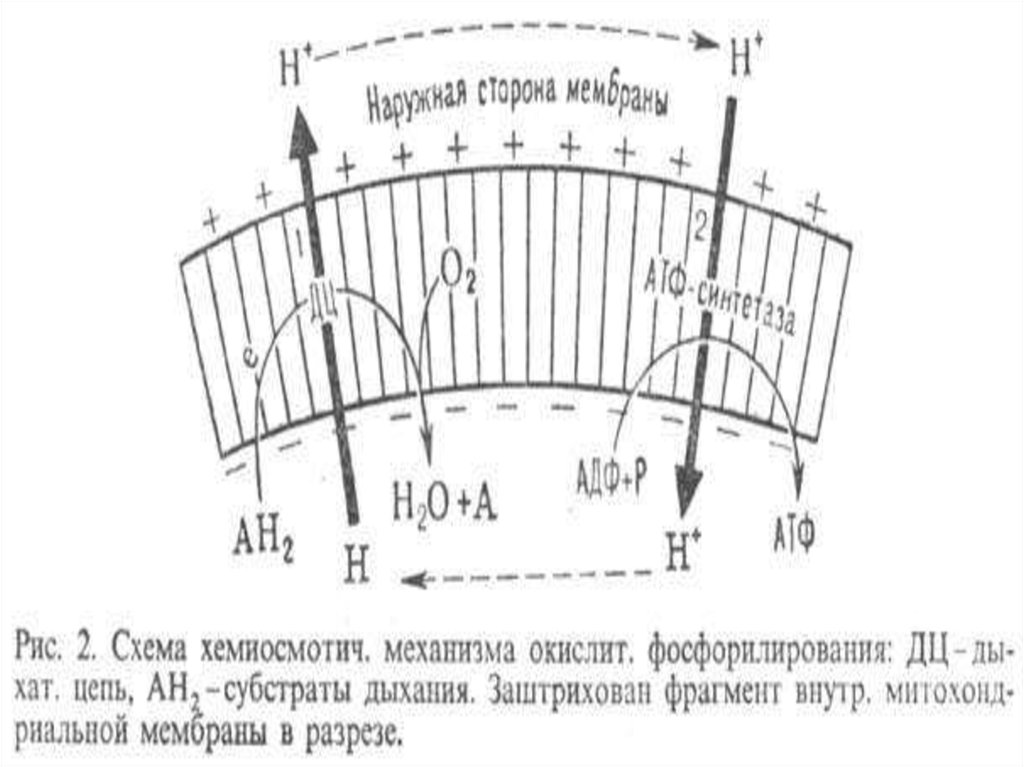
Объясняетхемиосмотическая гипотеза
Митчелла.
63.
64. Гипотеза Митчелла
Реакции,сопровождающиеся
образованием протонов,
протекают на вн. МХ мембране
таким образом, что протоны
переносятся с внутренней
стороны мембраны на
внешнюю.
65.
66.
67.
Энергияпереноса электронов
и протонов вдоль ДЦ
сосредотачивается в виде
протонного потенциала или
электрохимического градиента
Н+.
68.
Диффузияпротонов
обратно через мембрану
сопряжена с
фосфорилированием,
которое осуществляет
АТФсинтаза.
69. Дыхательная цепь
70. ЦПЭ, ДЫХАТЕЛЬНАЯ ЦЕПЬ
71.
Трансмембраннаяразность
электрических потенциалов
(протонный потенциал).
Трансмембранная разность
концентраций ионов Н+
72.
Н+- АТФ-синтаза состоит из:протонного канала (F0) и
каталитической субъединицы
(F1), образованной тремя α- и
тремя β-субъединицами, между
которыми расположены γ-, δ- и εсубъединицы.
73. АТФ-синтаза
74. АТФ-синтаза
75.
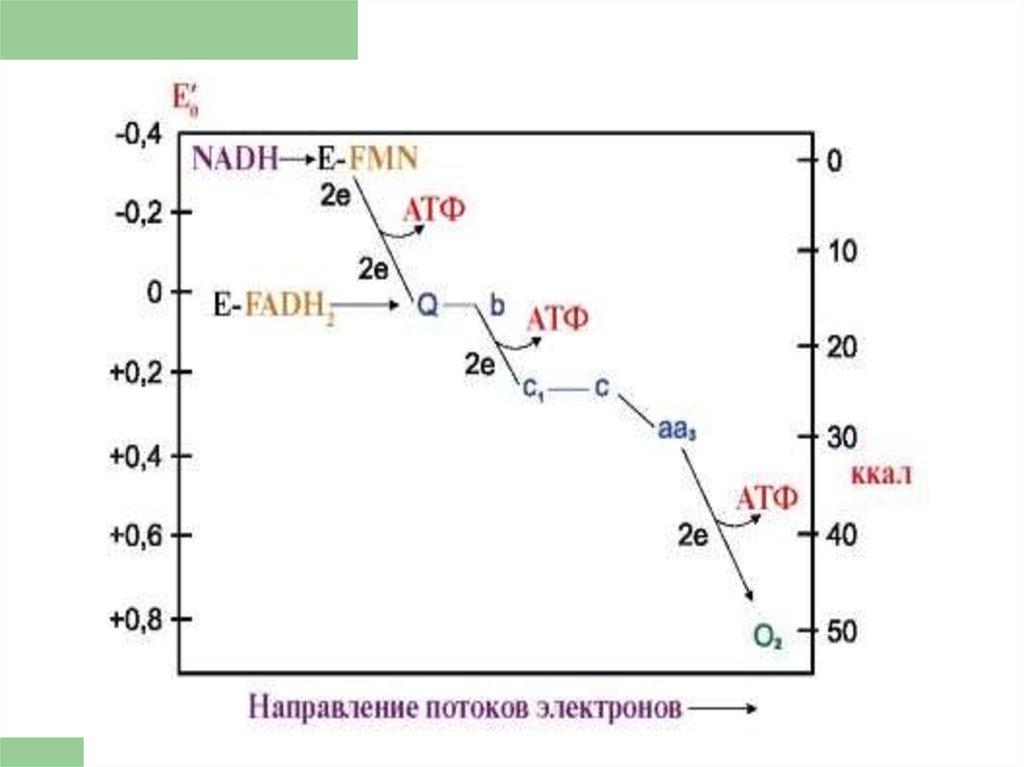
76. Коэффициент Р/О
При окислении каждой молекулыНАДН образуется 3 молекулы АТФ
(по одной в комплексе I, III и IV), а
при окислении одной молекулы
ФАДН2 — 2 молекулы АТФ (в
комплексе III и IV).
77.
78.
79.
80. Точки сопряжения
81.
82.
83.
84.
Механизм регуляции образования ипотребления АТФ наз. дыхательным
контролем.
Скорость О.Ф. зависит от
содержания АДФ.
85.
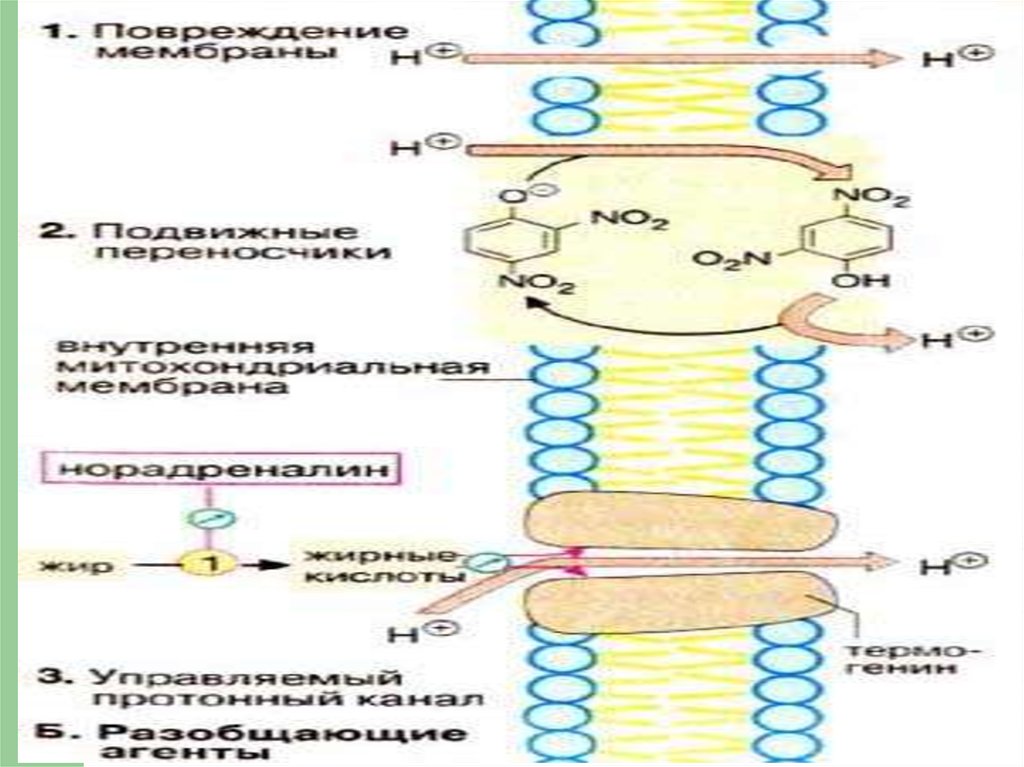
Разобщающие агентыВещества, которые функционально
разделяют между собой окисление и
фосфорилирование, наз.
разобщающими агентами.
Они содействуют переносу протонов
из ММП в матрикс без участия
АТФсинтазы.
86.
87. Разобщение
в результате- механического повреждения
внутренней мембраны;
- действия протонофоров –
(2,4-динитрофенол, тироксин).
88.
89.
90. Динитрофенол как разобщитель
91. Гипертермия
Если создание протонногоградиента подавлено, процессы
окисления субстрата и переноса
электронов протекают
значительно быстрее, однако
вместо синтеза АТФ выделяется
тепло (гипертермия).