

 Биология
БиологияПохожие презентации:










Цепь Переноса Электронов
1.
ЦЕПЬ ПЕРЕНОСА ЭЛЕКТРОНОВppA + pi
pppA
ppG + pppA
(нужно затратить 31 кДж/моль)
Kp ~ 1
pppG + ppA
G ~ 0
F = nEF
Eev =
E =
F0 = RTlnK(равновесия)
RT
nF
lnK
31000
F0 = E'nF
= 0,16 вольт
2х96500 Дж
G0' = -nF E0'
G0' = -220,2 кДж = 2х96,500 [0,82-(-0,32)]=1,12 в
1 кал = 4,187 Дж
2.
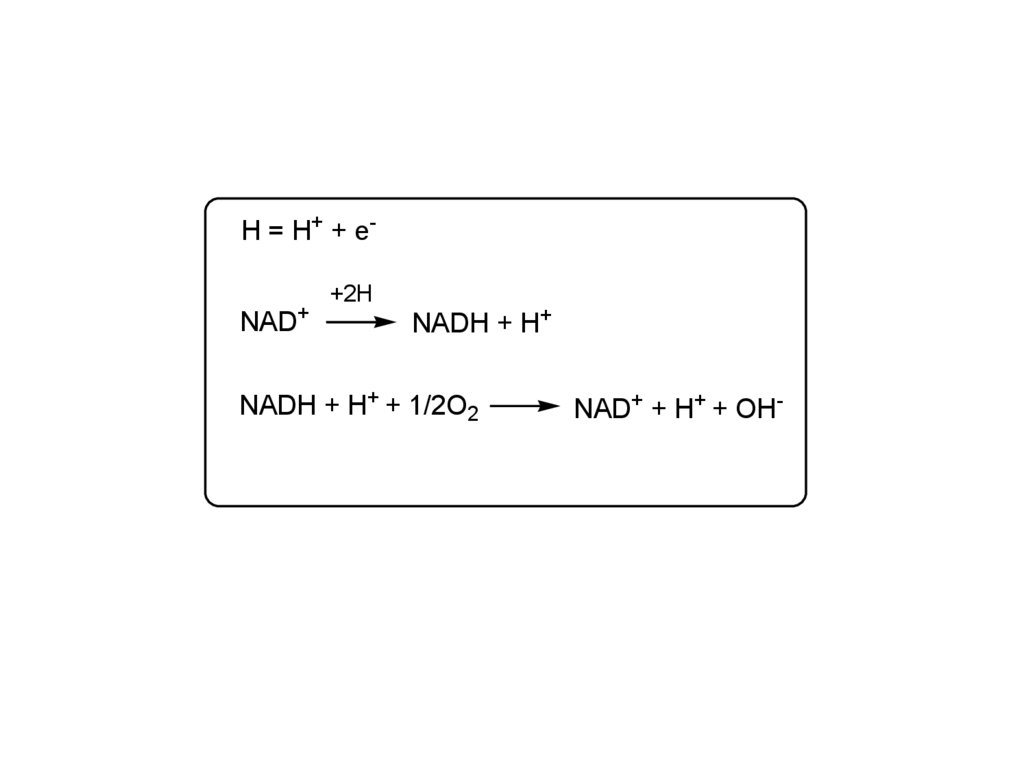
H = H+ + e+NAD
+2H
NADH + H+
NADH + H+ + 1/2O2
NAD+ + H+ + OH-
3.
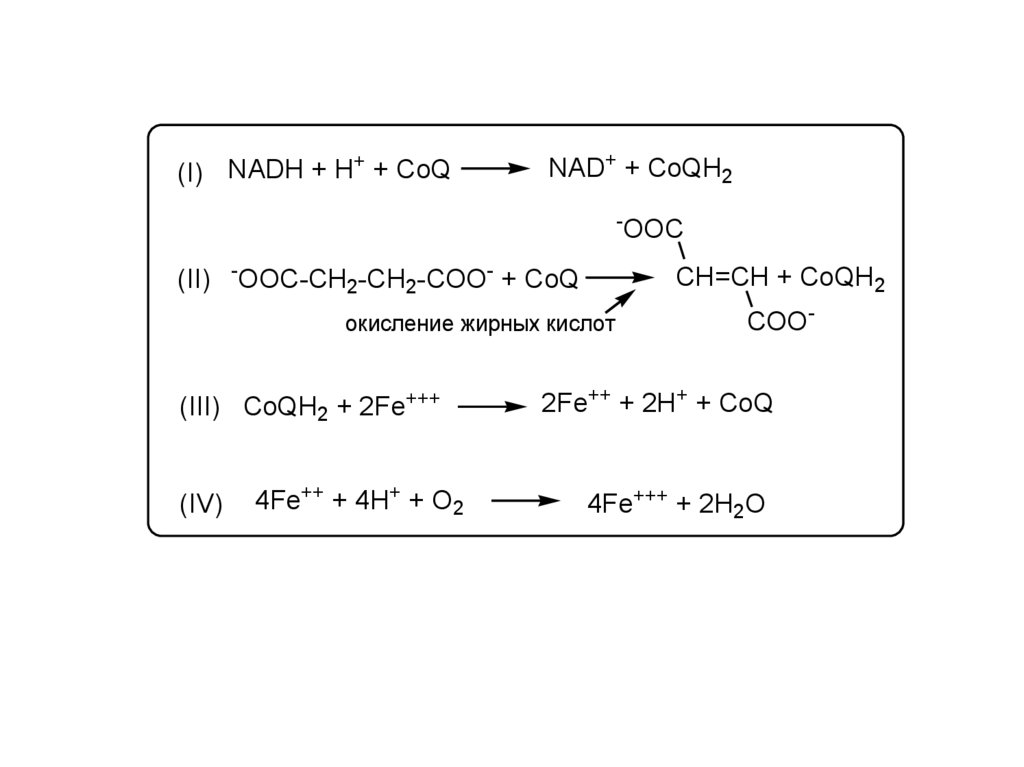
+(I) NADH + H + CoQ
NAD+ + CoQH2
-
(II)
-
OOC-CH2-CH2-COO- + CoQ
CH=CH + CoQH2
окисление жирных кислот
(III) CoQH2 + 2Fe+++
(IV)
4Fe++ + 4H+ + O2
OOC
COO-
2Fe++ + 2H+ + CoQ
4Fe+++ + 2H2O
4.
OHCH
CH 2
OH
OH CH
CH
O
CH 2
O
P
O-
ON
H3C
N
O
NH
H3C
N
O
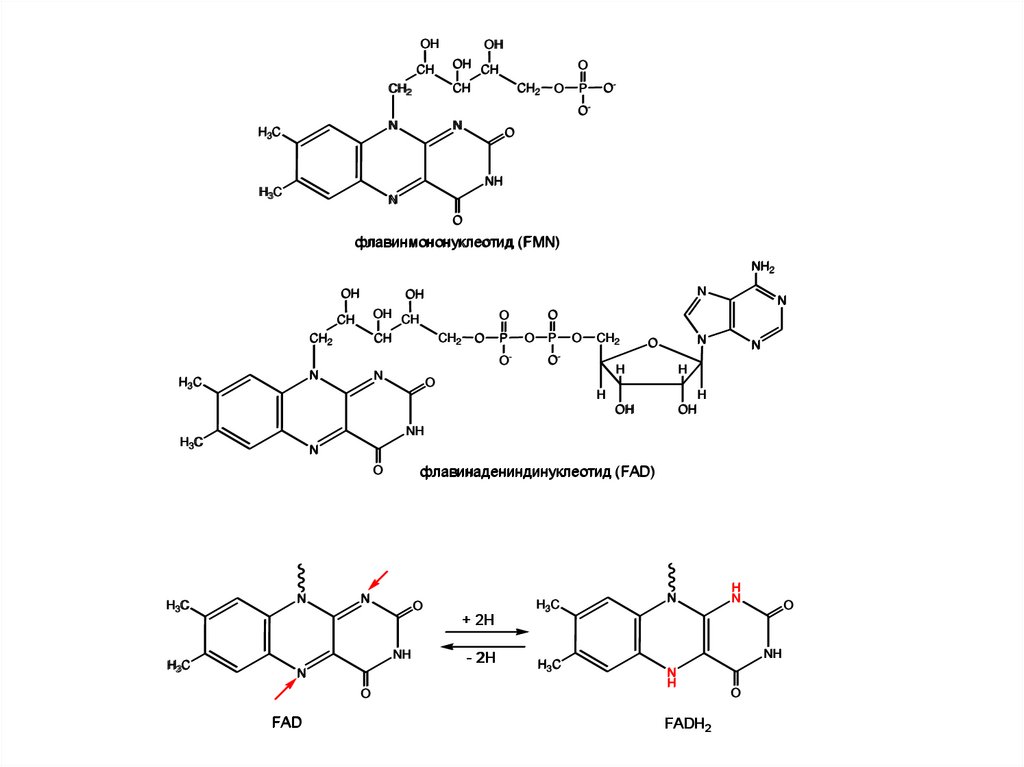
флавинмононуклеотид (FMN)
NH2
OH
N
OH
OH CH
CH
CH
CH2
CH2
O
P
O-
O
N
H3C
N
P
O
CH2
O-
O
N
O
H
H
OH
H
OH
N
O
NH
N
флавинадениндинуклеотид (FAD)
O
H3C
O
H
H3C
H3C
N
O
O
N
N
O
NH
N
O
FAD
+ 2H
- 2H
H3C
H3C
N
H
N
O
NH
N
H
FADH2
O
5.
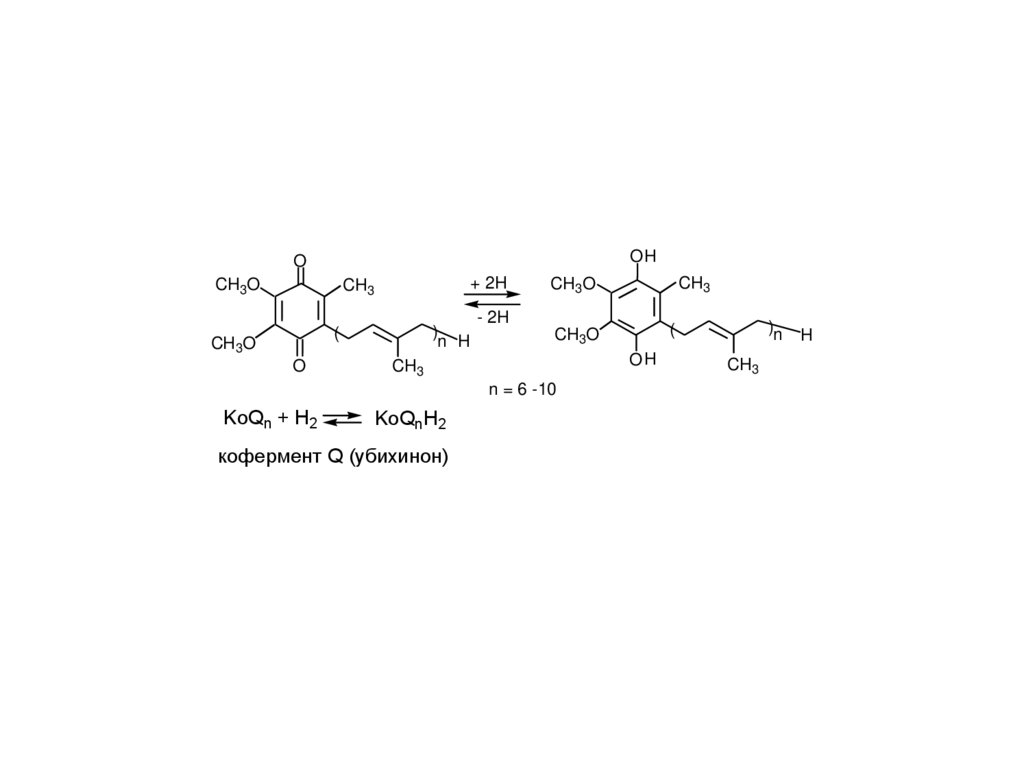
OHO
CH3O
+ 2H
CH3
)
n H
(
CH3O
O
- 2H
OH
CH3
KoQnH2
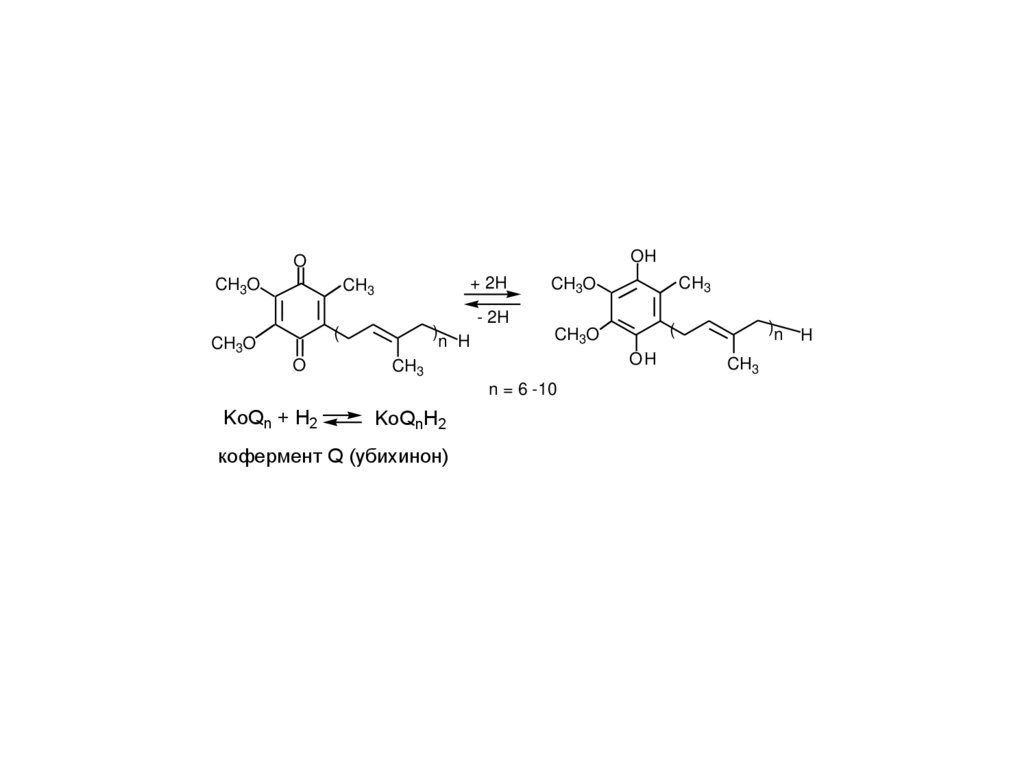
кофермент Q (убихинон)
)n
(
CH3O
n = 6 -10
KoQn + H2
CH3
CH3O
CH3
H
6.
OHO
CH3O
+ 2H
CH3
)
n H
(
CH3O
O
- 2H
OH
CH3
KoQnH2
кофермент Q (убихинон)
)n
(
CH3O
n = 6 -10
KoQn + H2
CH3
CH3O
CH3
H
7.
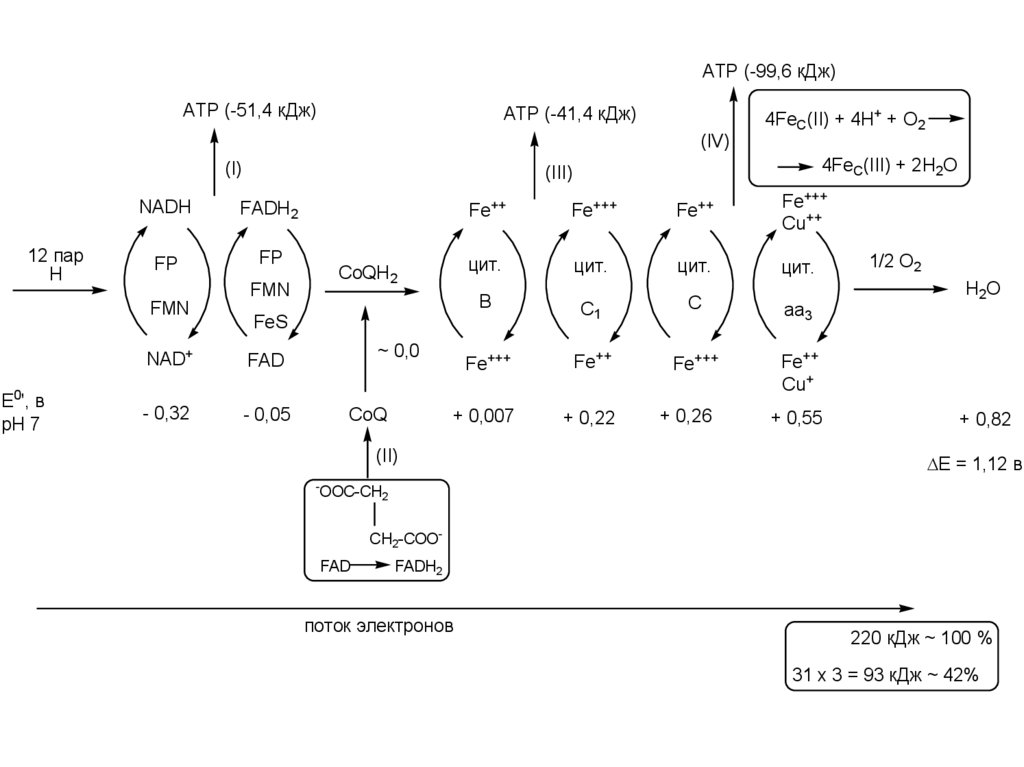
ATP (-99,6 кДж)ATP (-51,4 кДж)
ATP (-41,4 кДж)
(IV)
(I)
12 пар
Н
E0', в
pH 7
4FeC(III) + 2H2O
(III)
NADH
FADH2
FP
FP
FMN
4FeC(II) + 4H+ + O2
Fe++
Fe+++
Fe++
Fe+++
Cu++
цит.
цит.
цит.
цит.
В
C1
C
aa3
Fe+++
Fe++
Fe+++
Fe++
Cu+
+ 0,007
+ 0,22
CoQH2
FMN
FeS
NAD+
FAD
- 0,32
- 0,05
~ 0,0
CoQ
(II)
-
+ 0,26
+ 0,55
1/2 О2
Н2О
+ 0,82
E = 1,12 в
OOC-CH2
CH2-COOFAD
FADH2
поток электронов
220 кДж ~ 100 %
31 х 3 = 93 кДж ~ 42%
8.
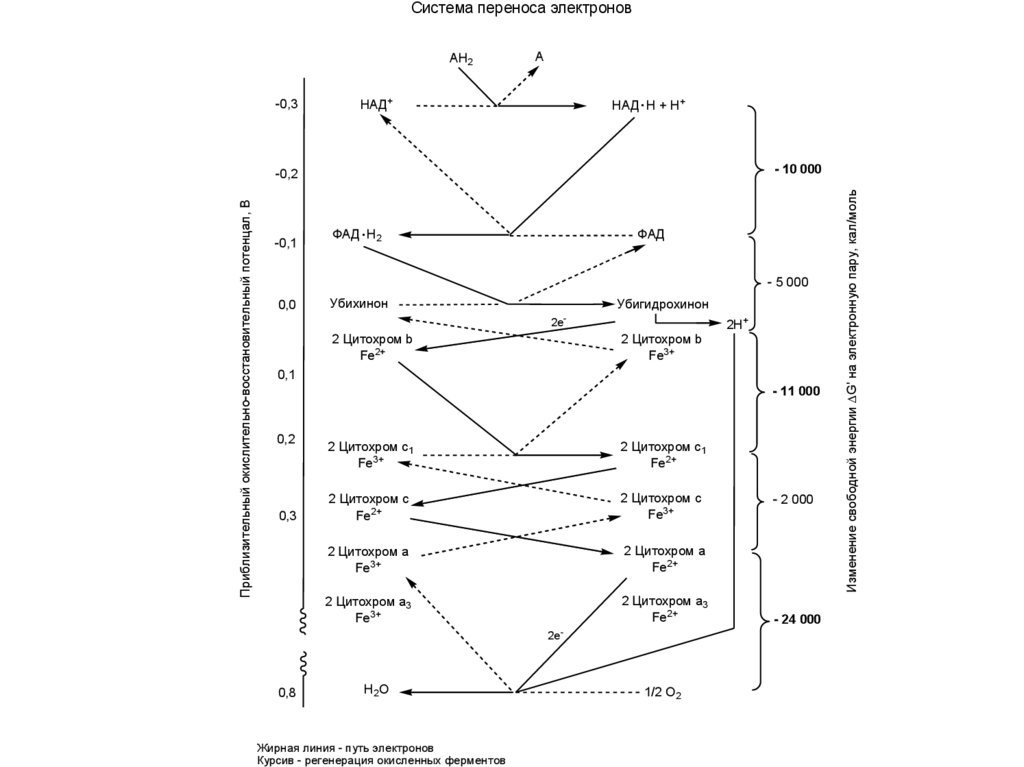
Система переноса электроновAH2
НАД. Н + Н+
НАД+
- 10 000
Приблизительный окислительно-восстановительный потенцал, В
-0,2
-0,1
ФАД . Н2
ФАД
- 5 000
0,0
Убихинон
Убигидрохинон
2е-
2 Цитохром b
Fe2+
2 Цитохром b
Fe3+
2Н+
0,1
- 11 000
0,2
0,3
2 Цитохром с1
Fe3+
2 Цитохром с1
Fe2+
2 Цитохром с
Fe2+
2 Цитохром с
Fe3+
2 Цитохром а
Fe3+
2 Цитохром а
Fe2+
2 Цитохром а3
Fe3+
2 Цитохром а3
Fe2+
2е-
0,8
Н2О
Жирная линия - путь электронов
Курсив - регенерация окисленных ферментов
1/2 О2
- 2 000
- 24 000
Изменение свободной энергии G' на электронную пару, кал/моль
-0,3
A
9.
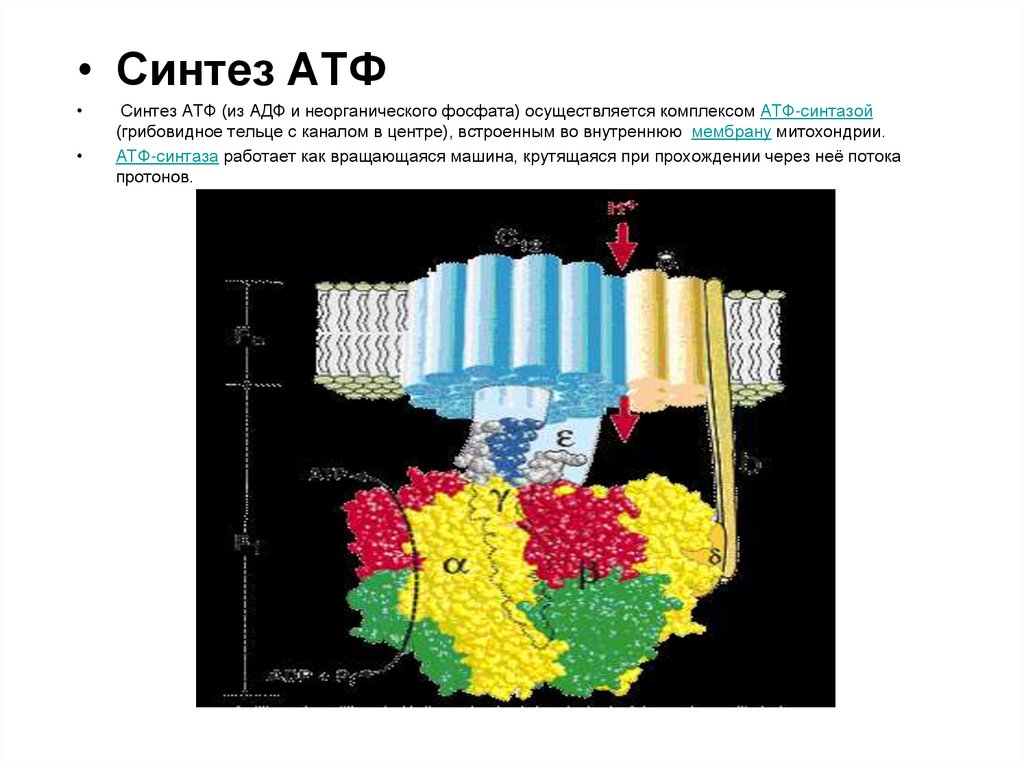
• Синтез АТФСинтез АТФ (из АДФ и неорганического фосфата) осуществляется комплексом АТФ-синтазой
(грибовидное тельце с каналом в центре), встроенным во внутреннюю мембрану митохондрии.
АТФ-синтаза работает как вращающаяся машина, крутящаяся при прохождении через неё потока
протонов.
10.
Гидрофобный комплекс F0 погружён в мембрану. Он служит основанием, которое фиксирует АТФ-синтазу
в мембране. Комплекс F0 состоит из нескольких субъединиц, образующих канал, по которому протоны
переносятся в матрикс.
Строение и механизм действия АТФ-синтазы. F0 – протонный канал (трансмембранный домен)
F1 - комплексы АТФ-синтазы находится вне мембраны, в матриксе). В состав F0входят полипептидные
цепи, которые образуют канал, пронизывающий мембрану насквозь. По этому каналу протоны
возвращаются в матрикс из межмембранного пространства;
белок F1 выступает в матрикс с внутренней стороны мембраны и содержит 9 субъединиц (Зα, 3β, γ, ε, δ),
6 из которых образуют 3 пары α и β ("головка"), прикрывающие стержневую часть, которая состоит из 3
субъединиц γ, δ и ε. γ и ε подвижны и образуют стержень, вращающийся внутри неподвижной головки и
связанный с комплексом F0. В активных центрах, образованных парами субъединиц α и β, происходит
связывание АДФ с неорганическим фосфатом (Рi) и синтезированной АТФ.
Каталитический цикл синтеза АТФ включает 3 фазы, каждая из которых проходит поочерёдно в 3
активных центрах: 1 - связывание АДФ и Н3РО4; 2 - образование фосфоангидридной связи АТФ;
3 - освобождение конечного продукта. При каждом переносе протонов через канал F0 в матрикс все
3 активных центра катализируют очередную фазу цикла. Энергия электрохимического
потенциала расходуется на поворот стержня, в результате которого циклически изменяется
конформация α- и β-субъединиц и происходит синтез АТФ.
Повышение концентрации протонов в межмембранном пространстве активирует АТФ-синтазу.
Электрохимический потенциал ΔμH+ заставляет протоны двигаться по каналу АТФ-синтазы в матрикс.
Параллельно под действием ΔμH+ происходят конформационные изменения в парах α, β-субъединиц
белка F1, в результате чего из АДФ и неорганического фосфата образуется АТФ. Электрохимический
потенциал, генерируемый в каждом из 3 пунктов сопряжения в ЦПЭ, используют для синтеза одной
молекулы АТФ.