













 Биология
БиологияПохожие презентации:










Память
1.
ПамятьПсихофизиология
Джебраилова Т.Д.
2.
Память— процесс, характеризующийся приобретением
(запоминанием), хранением и воспроизведением информации, а
также ее забывание.
Энграмма —след памяти, сформированный в результате обучения.
3.
Основные процессы памяти:Запоминание — это процесс памяти, посредством которого происходит запечатление
следов, ввод новых элементов ощущений, восприятие, мышления или переживания в
систему ассоциативных связей. Основу запоминания составляет связь материала со
смыслом в одно целое.
Хранение — процесс накопления материала в структуре памяти, включающий его
переработку и усвоение. Сохранение опыта дает возможность для обучения
человека, развития его перцептивных процессов, мышления и речи.
Воспроизведение и узнавание — процесс актуализации элементов прошлого опыта
(образов, мыслей, чувств, движений). Простой формой воспроизведения является
узнавание — опознание воспринимаемого объекта или явления как уже известного по
прошлому опыту, установлением сходств между объектом и его образом в памяти.
Забывание — потеря возможности воспроизведения, а иногда даже и в узнавании
раннее запомненного. Наиболее часто забывается то, что не значимо, не
актуализируется.
4.
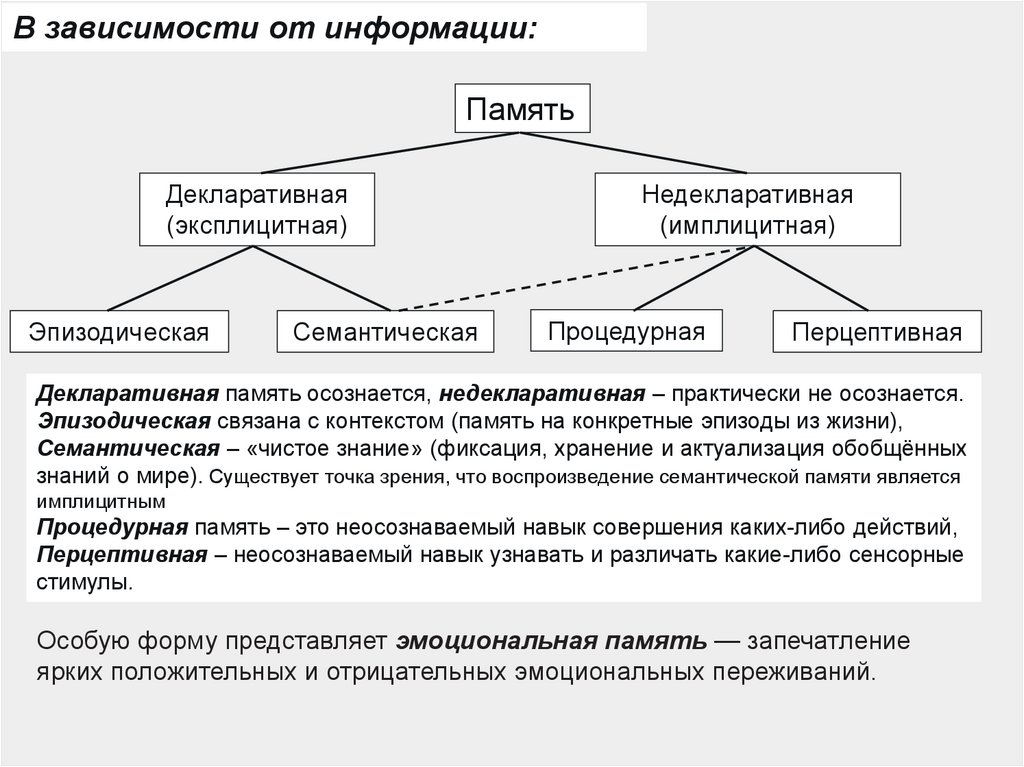
В зависимости от информации:Память
Декларативная
(эксплицитная)
Эпизодическая
Семантическая
Недекларативная
(имплицитная)
Процедурная
Перцептивная
Декларативная память осознается, недекларативная – практически не осознается.
Эпизодическая связана с контекстом (память на конкретные эпизоды из жизни),
Cемантическая – «чистое знание» (фиксация, хранение и актуализация обобщённых
знаний о мире). Существует точка зрения, что воспроизведение семантической памяти является
имплицитным
Процедурная память – это неосознаваемый навык совершения каких-либо действий,
Перцептивная – неосознаваемый навык узнавать и различать какие-либо сенсорные
стимулы.
Особую форму представляет эмоциональная память — запечатление
ярких положительных и отрицательных эмоциональных переживаний.
5.
Концепция временной организации памятиОсновное
- понятие о кратковременной и долговременной памяти.
- представление о разных нейрофизиологических, молекулярных,
биохимических и морфофункциональных основах энграммы на разных
стадиях ее жизни
- происходит смена одной формы существования энграммы на другую.
консолидация — процесс, приводящий к физическому закреплению
энграммы, переходу из кратковременной памяти в долговременную
Длительность консолидации — интервал времени, необходимый для
перехода следа памяти из кратковременного хранения, в котором он
находится в виде реверберирующей импульсной активности, в
долговременное, обеспечивающее длительное существование энграммы.
6.
Сенсорная (образная, иконическая ) память. Сенсорный образ, мгновенный отпечатоккартины внешнего мира, удерживается в памяти в пределах 50–500 мс. Зрительный
образ сохраняется при мигании. Этот вид памяти лежит в основе слитного восприятия
мелькающих на кино- и телеэкране изображений.
Кратковременная память (от нескольких секунд до нескольких минут) определяет
значимость поступающей информации для организма. Если эта информация важна для
организма, особенно для удовлетворения его ведущих потребностей, она
обрабатывается в промежуточной памяти и переходит в долговременную память. В
противном случае она быстро забывается.
Промежуточная память определяет сохранение полученной организмом информации в
течение нескольких минут или часов (формирование мыслей при разговоре,
запоминание телефонных разговоров, выполняемых заданий).
Долговременная память сохраняется в течение времени, сопоставимого с
продолжительностью жизни человека.
Важные для субъекта, особенно эмоционально окрашенные, события запечатляются в
долговременной памяти достаточно ярко.
Рабочая память ассоциирована, главным образом, с кратковременной памятью,
Используется в ходе решения определенных задач, ее использование завершается по мере
разрешения проблемы.
7.
Реверберация - основа кратковременной памятиЦиркуляция возбуждения по замкнутому
нейронному кругу определяет механизм
кратковременной памяти (А).
Циркуляция возбуждений прекращается, когда
в одном из нейронов цепи развивается
тормозной постсинаптическй потенциал под
влиянием тормозного нейрона (Б)
Реверберация возбуждений по большому
лимбическому кругу Пейпца
Стрелками показано:
1 — поясная извилина; 2 — гиппокамп;
3 — миндалина; 4 — гипоталамус
Прерывание или предотвращение реверберации должно приводить к
физическому разрушению следа памяти и его исчезновению ретроградной амнезии.
Однако….
Нормальная физиология: учебник /под ред. К.В.Судакова
М.: ГЭОТАР-Медиа, 2012.
8.
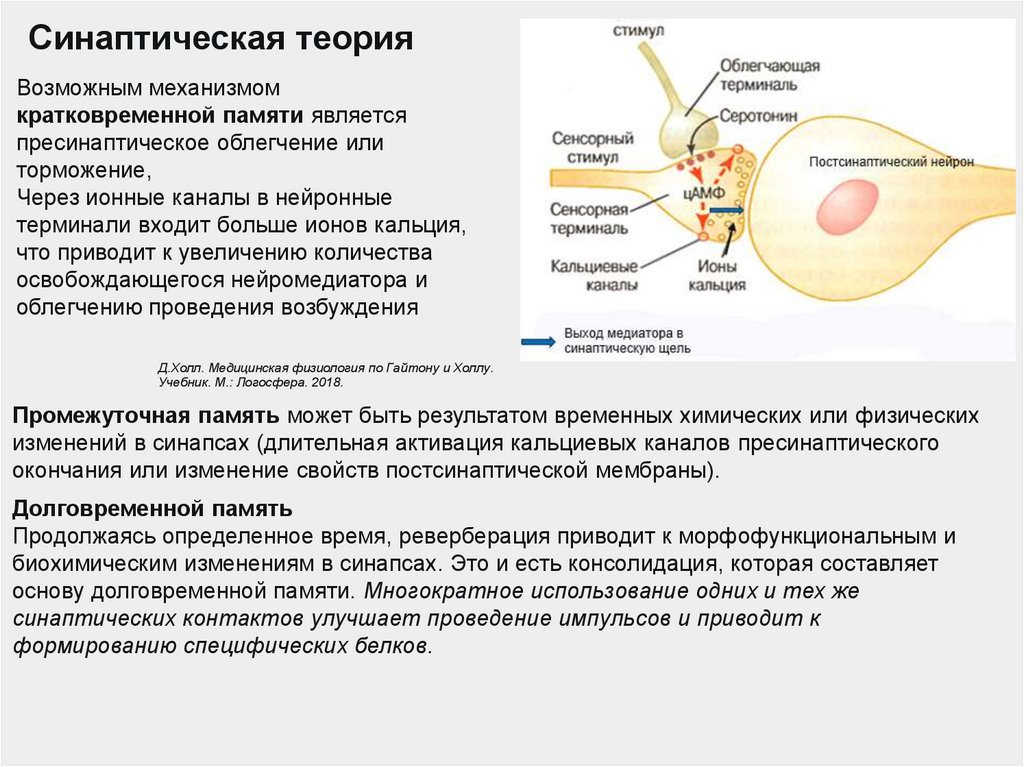
Синаптическая теорияВозможным механизмом
кратковременной памяти является
пресинаптическое облегчение или
торможение,
Через ионные каналы в нейронные
терминали входит больше ионов кальция,
что приводит к увеличению количества
освобождающегося нейромедиатора и
облегчению проведения возбуждения
Д.Холл. Медицинская физиология по Гайтону и Холлу.
Учебник. М.: Логосфера. 2018.
Промежуточная память может быть результатом временных химических или физических
изменений в синапсах (длительная активация кальциевых каналов пресинаптического
окончания или изменение свойств постсинаптической мембраны).
Долговременной память
Продолжаясь определенное время, реверберация приводит к морфофункциональным и
биохимическим изменениям в синапсах. Это и есть консолидация, которая составляет
основу долговременной памяти. Многократное использование одних и тех же
синаптических контактов улучшает проведение импульсов и приводит к
формированию специфических белков.
9.
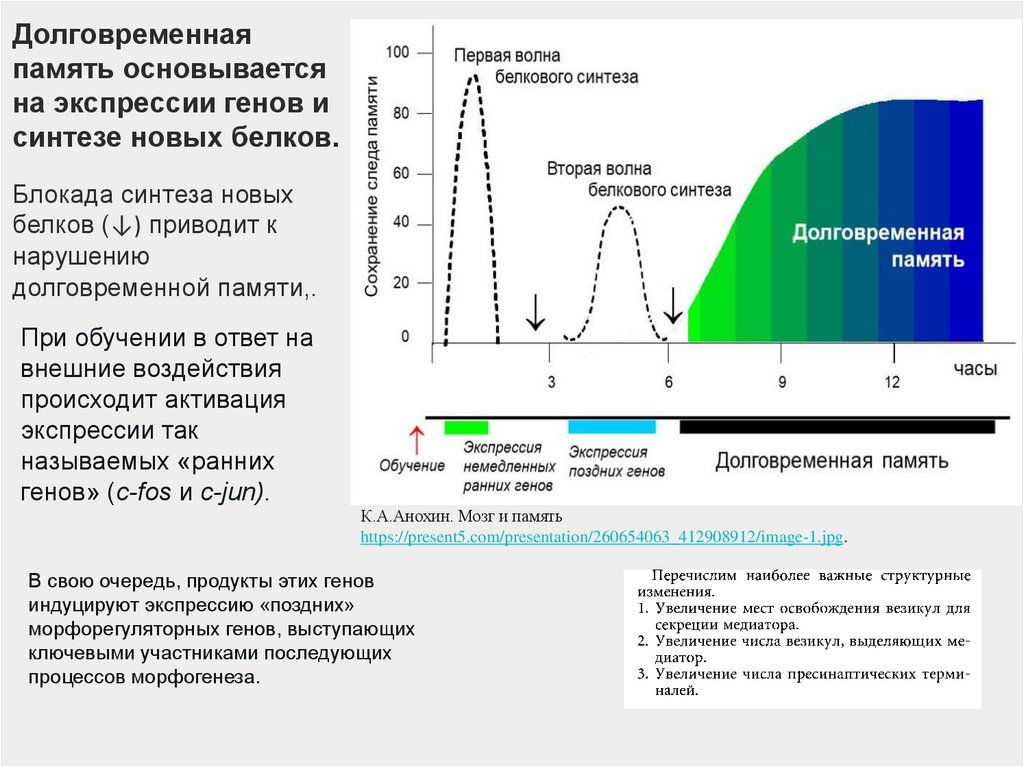
Долговременнаяпамять основывается
на экспрессии генов и
синтезе новых белков.
Блокада синтеза новых
белков (↓) приводит к
нарушению
долговременной памяти,.
При обучении в ответ на
внешние воздействия
происходит активация
экспрессии так
называемых «ранних
генов» (с-fos и c-jun).
К.А.Анохин. Мозг и память
https://present5.com/presentation/260654063_412908912/image-1.jpg.
В свою очередь, продукты этих генов
индуцируют экспрессию «поздних»
морфорегуляторных генов, выступающих
ключевыми участниками последующих
процессов морфогенеза.
10.
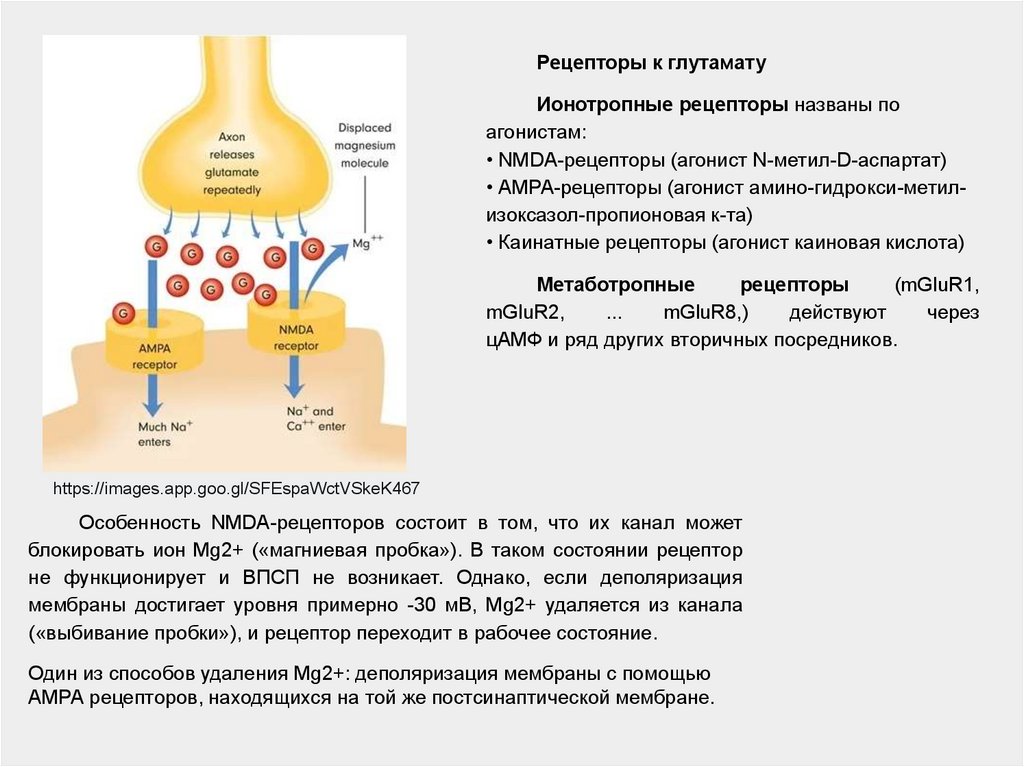
Рецепторы к глутаматуИонотропные рецепторы названы по
агонистам:
• NMDA-рецепторы (агонист N-метил-D-аспартат)
• АМРА-рецепторы (агонист амино-гидрокси-метилизоксазол-пропионовая к-та)
• Каинатные рецепторы (агонист каиновая кислота)
Метаботропные
рецепторы
(mGluR1,
mGluR2,
...
mGluR8,)
действуют
через
цАМФ и ряд других вторичных посредников.
https://images.app.goo.gl/SFEspaWctVSkeK467
Особенность NMDA-рецепторов состоит в том, что их канал может
блокировать ион Mg2+ («магниевая пробка»). В таком состоянии рецептор
не функционирует и ВПСП не возникает. Однако, если деполяризация
мембраны достигает уровня примерно -30 мВ, Mg2+ удаляется из канала
(«выбивание пробки»), и рецептор переходит в рабочее состояние.
Один из способов удаления Mg2+: деполяризация мембраны с помощью
АМРА рецепторов, находящихся на той же постсинаптической мембране.
11.
глутамат в механизмах памятиНа мембране пирамидных клеток гиппокампа (а также
в других частях мозга) находится две разновидности
глутаматных рецепторов: AMPA- и NMDA-рецепторы.
• пресинаптический нейрон, контактирующий с пирамидной
клеткой, выделяет глутамат;
• глутамат, связываясь с AMPA-рецепторами, изменяет
их конформацию таким образом, что ионный канал начинает
пропускать внутрь постсинаптической клетки ионы натрия;
• поступление ионов натрия вызывает локальную
деполяризацию,
• локальная деполяризация приводит к тому, что ион магния,
закрывающий ионный канал NMDA-рецепторов, покидает
своё место;
• глутамат связывается с NMDA-рецепторами,
в результате чего их свободные от ионов магния каналы
начинают пропускать внутрь пирамидной клетки ионы
кальция.
• Входящий в клетку кальций запускает цепь реакций,
активирующих кальций-зависимые киназы (СаМКII) –
ферменты, фосфорилирующие белки.
• Фосфорилирование дополнительных AMPAрецепторов и встраивание их в мембрану.
• Усиление сигнала, получаемого постсинаптическим
нейроном за счёт увеличения числа AMPAрецепторов.
Поступающие внутрь постсинаптического нейрона ионы кальция вызывают в ней изменения, которые приводят к ещё большему
увеличению проводимости синапса за счёт увеличения количества AMPA-рецепторов.
Измененное состояние синапса сохраняется в течение нескольких минут или часов (кратковременная и
промежуточная память)
12.
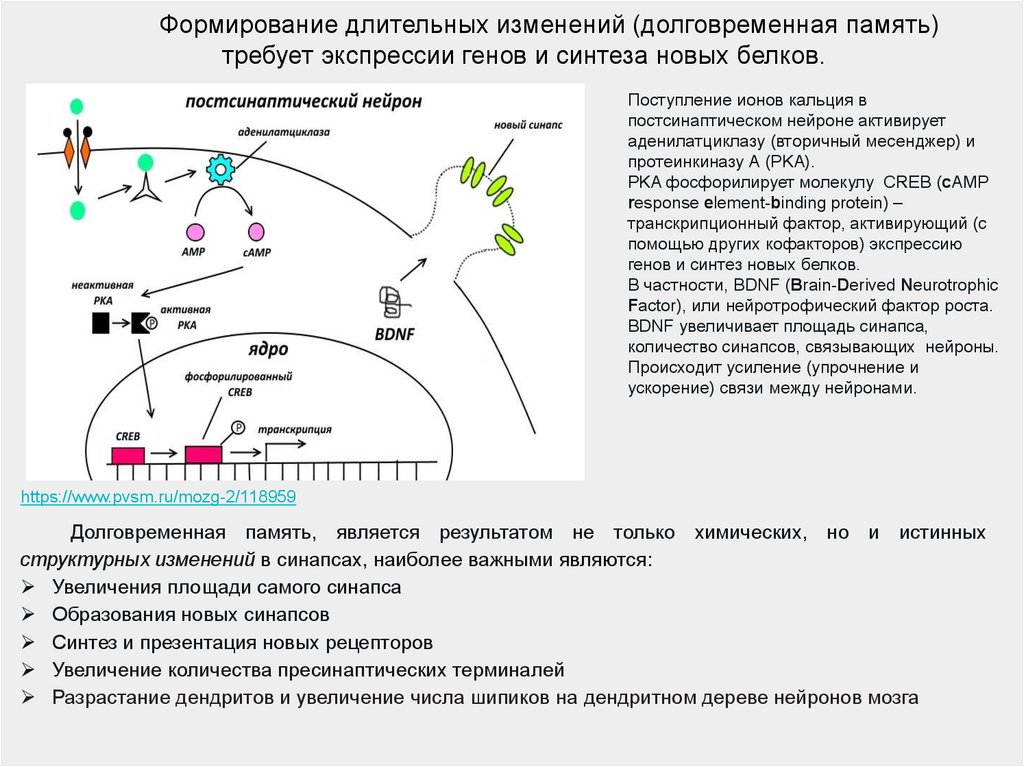
Формирование длительных изменений (долговременная память)требует экспрессии генов и синтеза новых белков.
Поступление ионов кальция в
постсинаптическом нейроне активирует
аденилатциклазу (вторичный месенджер) и
протеинкиназу А (PKA).
PKA фосфорилирует молекулу CREB (cAMP
response element-binding protein) –
транскрипционный фактор, активирующий (с
помощью других кофакторов) экспрессию
генов и синтез новых белков.
В частности, BDNF (Brain-Derived Neurotrophic
Factor), или нейротрофический фактор роста.
BDNF увеличивает площадь синапса,
количество синапсов, связывающих нейроны.
Происходит усиление (упрочнение и
ускорение) связи между нейронами.
https://www.pvsm.ru/mozg-2/118959
Долговременная память, является результатом не только химических, но и истинных
структурных изменений в синапсах, наиболее важными являются:
Увеличения площади самого синапса
Образования новых синапсов
Синтез и презентация новых рецепторов
Увеличение количества пресинаптических терминалей
Разрастание дендритов и увеличение числа шипиков на дендритном дереве нейронов мозга
13.
Важнейшую роль в механизмах памяти играют глутаматергические синапсы.К глутамату известно несколько типов рецепторов. В частности, такие ионотропные рецепторы, как:
• NMDA-рецепторы (агонист N-меил-D-аспартат)
• АМРА-рецепторы (агонист амино-гидрокси-метил-изоксазол-пропионовая к-та)
Уникальная особенность NMDA-рецепторов состоит в том, что их канал может блокировать ион Mg2+ («магниевая
пробка»). В такой ситуации рецептор выключен, и ВПСП не возникает. Однако, если заряд в нейроне оказывается выше
уровня -30 мВ, Mg2+ удаляется из канала («выбивание пробки»), и рецептор переходит в рабочее состояние.
Этот механизм – один из важнейших способов резко усилить эффективность работы синапса, создать новый путь для
передачи информации. Подобные изменения лежат в основе процессов обучения и формирования памяти.
Один из способ удаления Mg2+: деполяризация мембраны с помощью АМРА рецепторов, находящихся на той же
постсинаптической мембране.
Подобного рода синапсы, способные практически мгновенно увеличить эффективность работы, характерны для коры
больших полушарий и, особенно, гиппокампа, избирательно связанного с кратковременной памятью.
Взаимодействие глутамата с рецепторами приводит к повышенью проницаемости постсинаптической мембраны для
ионов кальция. Входящий в клетку Са2+ активирует каскад химических реакций, фосфорилирование дополнительных
рецепторов и их встраивание в мембрану. Измененное состояние синапса сохраняется в течение нескольких минутчасов (кратковременная и промежуточная память).
Более длительные изменения обеспечиваются передачей сигнала посредством ионов Са2+ на ядерную ДНК,
активацией генов, синтезом дополнительных белков-рецепторов и их встраиванием в постсинаптическую мембрану.
Подобная реакция требует значительного времени (часы-сутки), но зато измененное состояние синапса сохраняется
неограниченно долго («долговременная память»).
Кратковременная память требует фосфорилирования уже присутствующих в клетке белков.
Долговременная память основывается на экспрессии генов и синтезе новых белков.
14.
Концепция распределенной памятиПамять возникает при взаимодействии многих структур мозга
Опыты с локальными раздражениями мозга показали, что след памяти через
разное время реализуется разными мозговыми структурами или их частями. след
как бы распределен по нервным клеткам, принадлежащим различным
мозговым образованиям
Принцип распределенности энграммы — основа организации памяти
след памяти не имеет строгой локализации, а считывается
с нейронов разных структур мозга в зависимости от обстоятельств
Американский нейропсихолог К. Лешли обучал крыс инструментальным
оборонительным задачам и удалял у них различные участки коры головного мозга в
поисках локализации энграммы памяти.
Оказалось, что нарушения памяти не зависели от локализации повреждения, а
определялись только объемом повреждения мозга. Это легло в основу
представления об эквипотенциальности коры. К.Лешли пришел к выводу о том, что
«памяти нигде нет, но в то же время она всюду».
Удаление области коры, при стимуляции которой у людей развивались
определенные воспоминания, не отражается на возможности вызывать те же самые
воспоминания раздражением другой области коры
15.
Гиппокампальнаясистема
может
быть
местом
сохранения
кратковременной памяти, потому что синапсы гиппокампа способны
быстро изменяться.
Неокортикальные синапсы изменяются медленнее. С течением времени и в
результате повторных активаций память полностью переходит в
неокортекс.
Консолидация происходит, когда гиппокампальная система повторно
реактивирует представительство в неокортексе, что постепенно приводит к
усилению внутренних связей кортикальных областей, которые могут
поддерживать память независимо от гиппокампальной системы.
16.
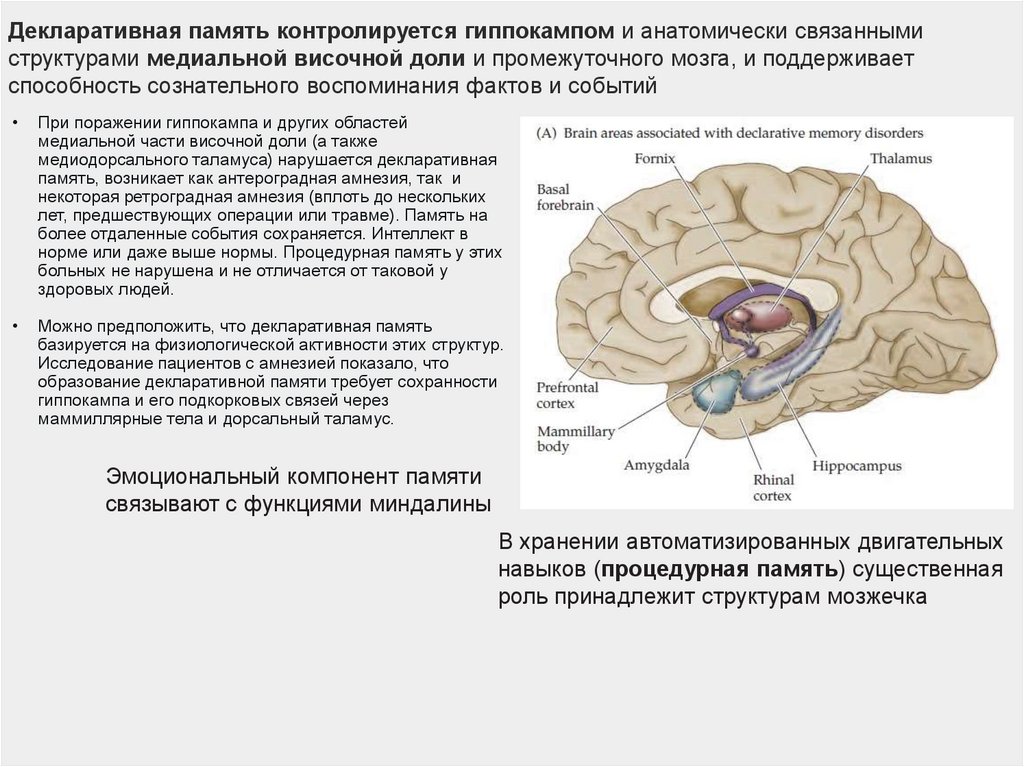
Декларативная память контролируется гиппокампом и анатомически связаннымиструктурами медиальной височной доли и промежуточного мозга, и поддерживает
способность сознательного воспоминания фактов и событий
При поражении гиппокампа и других областей
медиальной части височной доли (а также
медиодорсального таламуса) нарушается декларативная
память, возникает как антероградная амнезия, так и
некоторая ретроградная амнезия (вплоть до нескольких
лет, предшествующих операции или травме). Память на
более отдаленные события сохраняется. Интеллект в
норме или даже выше нормы. Процедурная память у этих
больных не нарушена и не отличается от таковой у
здоровых людей.
Можно предположить, что декларативная память
базируется на физиологической активности этих структур.
Исследование пациентов с амнезией показало, что
образование декларативной памяти требует сохранности
гиппокампа и его подкорковых связей через
маммиллярные тела и дорсальный таламус.
Эмоциональный компонент памяти
связывают с функциями миндалины
В хранении автоматизированных двигательных
навыков (процедурная память) существенная
роль принадлежит структурам мозжечка
17.
Роль височной коры в процессе воспоминанияКанадский нейрохирург У. Пенфилд впервые показал, что стимуляция медиальных
отделов височных долей коры головного мозга у пациентов во время
нейрохирургических операций вызывала детальные воспоминания о событиях,
имевших место в отдаленном прошлом в адекватной временной последовательности
Следует иметь в виду, что память обусловлена деятельностью большого количества
объединенных в структурно-функциональные ансамбли нейронов
головного мозга, расположенных как в коре, так и в подкорковых
образованиях, особенно в лимбических структурах головного мозга.
Стимуляция височной коры только активирует эту корково-подкорковую
энграмму памяти.
18.
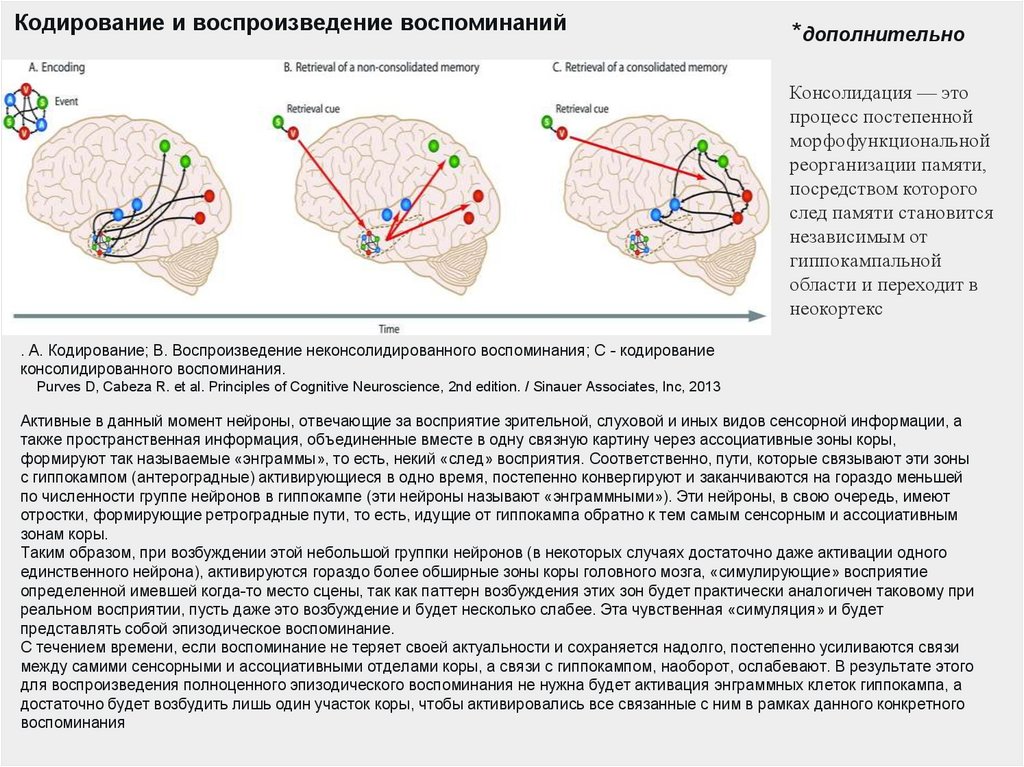
Кодирование и воспроизведение воспоминаний*дополнительно
Консолидация — это
процесс постепенной
морфофункциональной
реорганизации памяти,
посредством которого
след памяти становится
независимым от
гиппокампальной
области и переходит в
неокортекс
. А. Кодирование; В. Воспроизведение неконсолидированного воспоминания; С - кодирование
консолидированного воспоминания.
Purves D, Cabeza R. et al. Principles of Cognitive Neuroscience, 2nd edition. / Sinauer Associates, Inc, 2013
Активные в данный момент нейроны, отвечающие за восприятие зрительной, слуховой и иных видов сенсорной информации, а
также пространственная информация, объединенные вместе в одну связную картину через ассоциативные зоны коры,
формируют так называемые «энграммы», то есть, некий «след» восприятия. Соответственно, пути, которые связывают эти зоны
с гиппокампом (антероградные) активирующиеся в одно время, постепенно конвергируют и заканчиваются на гораздо меньшей
по численности группе нейронов в гиппокампе (эти нейроны называют «энграммными»). Эти нейроны, в свою очередь, имеют
отростки, формирующие ретроградные пути, то есть, идущие от гиппокампа обратно к тем самым сенсорным и ассоциативным
зонам коры.
Таким образом, при возбуждении этой небольшой группки нейронов (в некоторых случаях достаточно даже активации одного
единственного нейрона), активируются гораздо более обширные зоны коры головного мозга, «симулирующие» восприятие
определенной имевшей когда-то место сцены, так как паттерн возбуждения этих зон будет практически аналогичен таковому при
реальном восприятии, пусть даже это возбуждение и будет несколько слабее. Эта чувственная «симуляция» и будет
представлять собой эпизодическое воспоминание.
С течением времени, если воспоминание не теряет своей актуальности и сохраняется надолго, постепенно усиливаются связи
между самими сенсорными и ассоциативными отделами коры, а связи с гиппокампом, наоборот, ослабевают. В результате этого
для воспроизведения полноценного эпизодического воспоминания не нужна будет активация энграммных клеток гиппокампа, а
достаточно будет возбудить лишь один участок коры, чтобы активировались все связанные с ним в рамках данного конкретного
воспоминания
19.
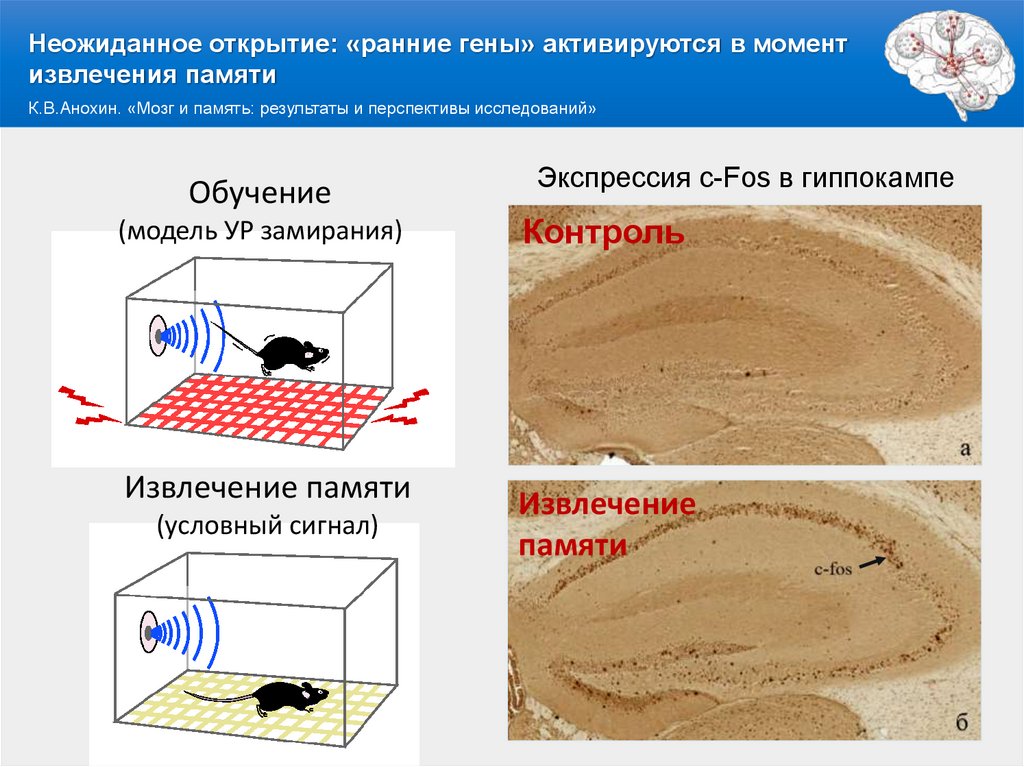
Неожиданное открытие: «ранние гены» активируются в моментизвлечения памяти
К.В.Анохин. «Мозг и память: результаты и перспективы исследований»
Обучение
(модель УР замирания)
Извлечение памяти
(условный сигнал)
Экспрессия c-Fos в гиппокампе
Контроль
Извлечение
памяти
20.
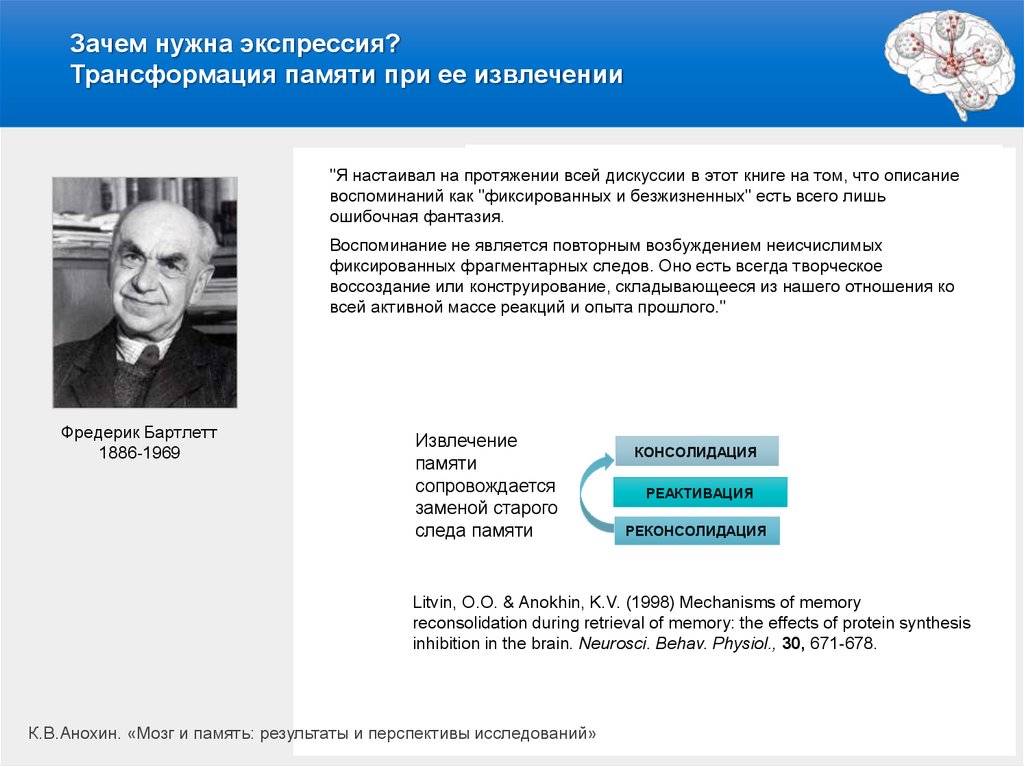
Зачем нужна экспрессия?Трансформация памяти при ее извлечении
"Я настаивал на протяжении всей дискуссии в этот книге на том, что описание
воспоминаний как "фиксированных и безжизненных" есть всего лишь
ошибочная фантазия.
Воспоминание не является повторным возбуждением неисчислимых
фиксированных фрагментарных следов. Оно есть всегда творческое
воссоздание или конструирование, складывающееся из нашего отношения ко
всей активной массе реакций и опыта прошлого."
Фредерик Бартлетт
1886-1969
Извлечение
памяти
сопровождается
заменой старого
следа памяти
КОНСОЛИДАЦИЯ
РЕАКТИВАЦИЯ
РЕКОНСОЛИДАЦИЯ
Litvin, O.O. & Anokhin, K.V. (1998) Mechanisms of memory
reconsolidation during retrieval of memory: the effects of protein synthesis
inhibition in the brain. Neurosci. Behav. Physiol., 30, 671-678.
К.В.Анохин. «Мозг и память: результаты и перспективы исследований»
21.
ЗабываниеЗабывание - это невозможность вспомнить в данный момент то, что было
известно раньше.
Две основные формы определения забывания:
• как полная потеря («стирание») следа памяти, воспроизвести который невозможно
независимо от применяемых средств;
• как невозможность «извлечения памяти», при сохранении ее материальной основы.
(оба определения относятся только к случаям «здорового» забывания, при котором у испытуемых нет
патологических нарушений).
Забывание рассматривается как одно из проявлений процесса формирования
структуры индивидуального опыта: оно зависит от содержания материала, задачи
запоминания, требует синтеза белков, является, по-видимому, необходимой частью
модификации опыта. Забывание является частью реорганизации, или динамики,
памяти (Созинов А. А., Александров Ю. И., 2022)
22.
• Амнезия - нарушение процессов памяти (запоминания,хранения, воспроизведения).
• Ретроградная амнезия - это утрата памяти на события,
предшествующие действию амнестического агента (травма,
интоксикация, электрошок и т.п.)
• Антероградная амнезия - нарушение способности к
передаче информации из кратковременно в долговременную
память. Может проявляться в утрате памяти на недавние
события при сохранении памяти на отдаленные события, т.е.
долговременная память может быть сохранна.
23.
В системных процессах извлечения следов памяти ведущая роль принадлежитдоминирующей мотивации. Мотивация:
обусловливает быстрое извлечение накопленного опыта из памяти
способствует его удержанию при наличии препятствий к достижению цели
выступает как ведущий фактор фиксации опыта в памяти
наибольшая эффективность запоминания соответствует оптимальному уровню
мотивации
В системной организации целенаправленных поведенческих актов память является
одним из компонентов стадии афферентного синтеза, участвуя во взаимодействии с
мотивацией и обстановочной афферентацией в создании так называемой предпусковой
интеграции