
























")
")





")






:")






















")













 Медицина
МедицинаПохожие презентации:
")


")






Высшая нервная деятельность
1. ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ
Мухина Ирина ВасильевнаЛечебный, педиатрический факультеты
2022
2. Содержание
1. Высшая и низшая нервная деятельность по Павлову И.П.2. Функциональная организация условного рефлекса
3. Классификация условных и безусловных рефлексов
4. Торможение условных рефлексов
5. Динамические стереотипы
6. Типы ВНД
7. Методы исследования деятельности головного мозга
8. Физиология памяти
9. Физиология мотиваций
10. Физиология эмоций
11. Роль ретикулярной формации (РФ) в интегративной
деятельности мозга
12. Физиология сна
2
3. По Павлову И.П.
• Низшая нервная деятельность представляет собойсовокупность безусловных рефлексов, обеспечивающую
координированную деятельность всех его систем и
направленную на поддержание гомеостаза (показателей
кровяного, осмотического и онкотического давления, активной
реакции среды, температуры, парциального напряжения О2 и
СО2 в крови, содержание в ней сахара и т.д.).
• Высшая нервная деятельность - интегративная деятельность
высших отделов ЦНС, обеспечивающая индивидуальное
поведенческое приспособление человека и животных к
изменяющимся условиям внешней и внутренней среды.
Термин “ВНД” введен Павловым как эквивалент понятия
“психическая деятельность”.
3
4.
НИЗШАЯ НЕРВНАЯДЕЯТЕЛЬНОСТЬ
1.
2.
3.
4.
5.
ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ
БЕЗУСЛОВНЫЕ РЕФЛЕКСЫ:
постоянны;
1.
стереотипны;
2.
наследственно предопределены;
являются видовыми,
3.
возникают на адекватные
биологически значимые раздражения 4.
соответствующих рефлексогенных
зон.
5.
УСЛОВНЫЕ РЕФЛЕКСЫ:
динамичны;
разнообразны, изменчивы,
непостоянны;
индивидуальны и не
наследуются;
формируются при
определенных условиях;
могут проявляться на
раздражения любых
рецептивных полей.
Инстинкт – сложнейший безусловный
рефлекс, представляющий собой ряд
(цепочку) рефлекторных реакций,
Динамический стереотип каждая из которых является толчком для система условных и безусловных
последующей реакции.
рефлексов,
представляющая
собою единый функциональный
комплекс (навыки)
4
5. УСЛОВИЯ ВЫРАБОТКИ УСЛОВНЫХ РЕФЛЕКСОВ
УСЛОВИЯ ВЫРАБОТКИ УСЛОВНЫХРЕФЛЕКСОВ
УС
БП
• 1. Сочетание индифферентного УС с БП.
• 2. Опережающее действие УС.
• 3. Многократное повторение сочетаний УС с БП.
• 4. Большая биологическая значимость БП по
сравнению с УС.
• 5. Нормальная работоспособность ЦНС
• 6.Отсутствие посторонних раздражителей.
5
6. ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ УСЛОВНОГО РЕФЛЕКСА. УРОВНИ ЗАМЫКАНИЯ ВРЕМЕННЫХ СВЯЗЕЙ
67.
КЛАССИФИКАЦИЯ БЕЗУСЛОВНЫХРЕФЛЕКСОВ
На
основе
биологической
значимости:
пищевые,
оборонительные,
половые,
родительские,
игровые,
ориентировочный и т.д.
На
основе
анатомического
принципа:
простые (спинномозговые),
усложнённые (бульбарные),
сложные (мезенцефальные)
сложнейшие
(субкортикальнокортикальные).
КЛАССИФИКАЦИЯ УСЛОВНЫХ
РЕФЛЕКСОВ
Классификации, основанные на
особенностях БП:
По наличию или отсутствию БП
По биологическому значению БП
По характеру деятельности
организма, вызванной БП
По характеру подкрепления
Классификации, основанные на
особенностях УС:
По характеру рецепторов, на
которые действуют УС
По природе УС
По структуре УС
Классификации, основанные
на временных соотношениях между
УС и БП:
Наличные
Следовые
7
8. Классификации условных рефлексов, основанные на особенностях безусловного подкрепления:
1. По наличию или отсутствию БП:• подкрепляемые (возбудительные, положительные) ,
• неподкрепляемые (тормозные, отрицательные).
2. По биологическому значению БП:
• витальные (пищевые, оборонительные, регуляции сна и
бодрствования),
• зоосоциальные (половые, родительские, территориальные),
• саморазвития (исследовательские, имитационные, игровые).
3. По характеру деятельности организма, вызванной БП:
• вегетативные: секреторные (слюно-, желче-,
слезоотделительные, желудочносекреторные и др.), органные
(сердечные, сосудистые, дыхательные и др).
• двигательные: неинструментальные,
инструментальные (оперантные),
4. По характеру подкрепления:
1 порядка – один условный сигнал подкрепляется БП (звонок +пища).
2 порядка – сложный условный сигнал подкрепляется БП
((свет+звонок)+пища)
8
9. Классификации условных рефлексов, основанные на особенностях УС:
• По характеру рецепторов, на которые действуют УС:• экстероцептивные (зрительные, слуховые, обонятельные,
вкусовые, тактильные, температурные и др),
• интероцептивные (химические, механические,
осмотические и др.)
• 2. По природе УС:
• натуральные
• искусственные.
• 3. По структуре УС:
• Простые (мономодальные)
• Комплексные (мультимодальные): одновременные и
последовательные.
9
10. Классификации условных рефлексов, основанные на временных соотношениях между УС и БП:
• 1.Наличные:– совпадающие
(БП не позднее 5” после
начала действия УС),
– отставленные (БП спустя 5-30”),
– запаздывающие (БП спустя 30” и более).
• 2. Следовые - БП отделено от окончания
действия УС > или < временем.
10
11. ТОРМОЖЕНИЕ УСЛОВНЫХ РЕФЛЕКСОВ
1112. Роль торможения условных рефлексов:
• а) координирующая,• б) охранительная,
• в) совершенствование условных
рефлексов
12
13. Формы торможения условных рефлексов:
• 1. Безусловное или врожденноеторможение;
• 2. Условное или приобретенное,
торможение.
13
14. БЕЗУСЛОВНОЕ ТОРМОЖЕНИЕ
• Безусловное торможение не нужновырабатывать, оно изначально свойственно
всем элементам ЦНС и связано с
врожденными свойствами нервной системы.
Различают два вида безусловного торможения:
• 1. Внешнее;
• 2. Запредельное.
14
15. Внешнее торможение
• Внешнее торможение возникает под влиянием внешнихпосторонних для данного условного рефлекса раздражений.
• Причина – врожденный безусловный рефлекс на новизну (по
И.П. Павлову «Что такое?) – ориентировочный рефлекс,
который тормозит условную реакцию.
По характеру действия на условнорефлекторную деятельность
раздражители делятся на две группы:
• 1. Гаснущий тормоз – посторонний раздражитель, но который с
повторением его действия теряет свое тормозящее влияние,
так как не имеет существенного значения для организма.
• 2. Постоянный тормоз - посторонний раздражитель, который с
повторением его действия не теряет свое тормозящее
влияние, так как имеет существенное значение для организма.
15
16. Запредельное торможение
• Запредельное торможение возникает подвлиянием сильных или длительных, но
характерных для данного условного
рефлекса раздражений.
• Запредельное торможение развивается не
только при чрезмерной силе условного
раздражителя, но и при низком
функциональном состоянии нервной
системы, например у людей пожилого
возраста, на фоне нервного истощения
16
17. УСЛОВНОЕ ТОРМОЖЕНИЕ
• Условное торможение вырабатывается в процессежизнедеятельности и не возникает при первом же
применении раздражителя.
• Согласно И.П. Павлову различают четыре
разновидности условного торможения, объединенные
одним непременным обстоятельством – временным
или постоянным отсутствием подкрепления
безусловным раздражителем:
• 1. Угасательное
• 2. Дифференцировочное
• 3. Условный тормоз
• 4. Запаздывательное торможение
17
18.
• 1. Угасательное торможение - развивается приотмене безусловного подкрепления условного
сигнала.
• 2. Дифференцировочное торможение развивается
при неподкреплении раздражителей, похожих на
условный сигнал. Оно позволяет
дифференцировать (различать) раздражители
внешней среды.
• 3. Условнотормозное (условный тормоз) –
развивается в ситуации, когда условный сигнал,
отдельно подкрепляемый, не подкрепляется, если
применяется в комбинации с другим,
дополнительным раздражителем.
• 4.Запаздывательное торможение - развивается
при отставлении подкрепления на 1-3 мин
относительно начала действия условного сигнала.
18
19. ВЗАИМОДЕЙСТВИЕ РАЗЛИЧНЫХ ВИДОВ ТОРМОЖЕНИЯ
• Растормаживание выражается в том, что одинвид торможения частично или полностью
подавляет другой. Главным условием
растормаживания является неожиданное
вмешательство безусловного торможения в
момент реализации условного торможения.
• Суммация торможения может проявиться в
более быстром угашении следовых условных
рефлексов (Запаздывательное
торможение+Угасательное торможение) по
сравнению с угашением наличных рефлексов.
19
20. Динамические стереотипы
Динамический стереотип – это система условных ибезусловных рефлексов, представляющая собою
единый функциональный комплекс
Динамические стереотипы
характеризуются тем, что:
• они легко выполнимы.
• позволяют адекватно реагировать на изменения
обстановки,
• с возрастом крепнут,
• переделка динамических стереотипов возможна,
но всегда трудна.
20
21. ТИПЫ ВНД
• Тип ВНД – это совокупность врожденныхи приобретенных свойств нервной
системы, определяющих характер
взаимодействия организма с
окружающей средой и находящих свое
отражение во всех функциях организма.
21
22.
Параметры мозговой деятельности человека по Павлову И.П.:Сила нервных процессов оценивается по:
• скорости выработки условного рефлекса,
• реакции на увеличение силы условного сигнала,
• реакции на факторы, повышающие возбудимость (голодание, кофеин и
др).
Уравновешенность нервных процессов оценивается по:
• сравнительной скорости выработки «+» и «-» УР,
• соотношению «+» и «-» УР в ходе их формирования.
Подвижность нервных процессов оценивается по:
• скорости переделок дифференцировок и динамических стереотипов,
• реакции на многократную смену «+» и «-» УР с малыми временными
интервалами.
Параметры мозговой деятельности человека по Г. Айзенку:
•Экстравертивность и интравертивность
•Эмоциональная стабильность и нестабильность (нейротизм)
•Психотизм (следование социальным нормам)
22
23. По Павлову:
1 тип – «живой» - сильный,
уравновешенный, подвижный.
Для этого типа характерны
одинаково сильно выраженные
процессы В и Т, легко
приходящие на смену друг другу.
2 тип – «спокойный» сильный, уравновешенный,
инертный. В и Т выражены
также хорошо, как и у 1 типа, но
протекают значительно
медленнее.
3 тип – «безудержный» сильный, неуравновешенный.
Характеризуется сильным
процессом В, преобладающим
над Т.
4 тип – «слабый» характеризуется слабостью
процессов В и Т.
По Айзенку:
1. Стабильный экстраверт –
сильный, уравновешенный,
подвижный (сангвиник)
2. Стабильный интраверт
сильный, уравновешенный,
инертный (флегматик)
3. Нестабильный экстраверт –
сильный, неуравновешенный,
подвижный (холерик)
4. Нестабильный интроверт –
слабый тип (меланхолик).
23
24. МЕТОДЫ ИССЛЕДОВАНИЯ ДЕЯТЕЛЬНОСТИ ГОЛОВНОГО МОЗГА
2425.
• 1) разрушение различных участков головного мозга, вт.ч. удаления (экстирпации) тех или иных зон коры
больших полушарий,
• 2) электрическое или химическое раздражение
структур головного мозга,
• 3) временное выключение их функции (холодовое),
• 4) исследование условно-рефлекторной деятельности,
• 5) регистрация электрических проявлений в структурах
головного мозга в тех или иных условиях
(электрофизиологические методики – ЭЭГ, ВП),
• 6) Магнитоэнцефалография (МЭГ)
• 7) Измерение мозгового кровотока
• 8) Рентгенологическая компьютерная томография (КТ)
• 9) Позитронно-эмиссионная томография (ПЭТ)
• 10) Магнитно-резонансная томография (МРТ, фМРТ,
спектрМРТ)
• 11) Клинические наблюдения.
25
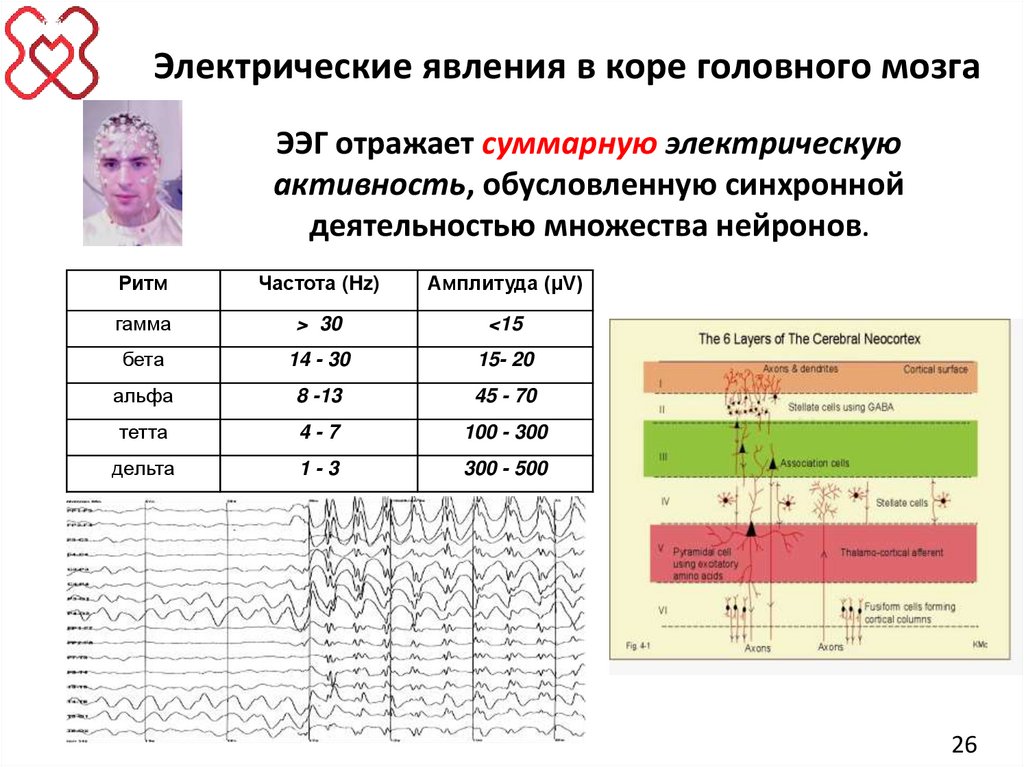
26.
Электрические явления в коре головного мозгаЭЭГ отражает суммарную электрическую
активность, обусловленную синхронной
деятельностью множества нейронов.
Ритм
Частота (Hz)
Амплитуда (µV)
гамма
> 30
<15
бета
14 - 30
15- 20
альфа
8 -13
45 - 70
тетта
4-7
100 - 300
дельта
1-3
300 - 500
26
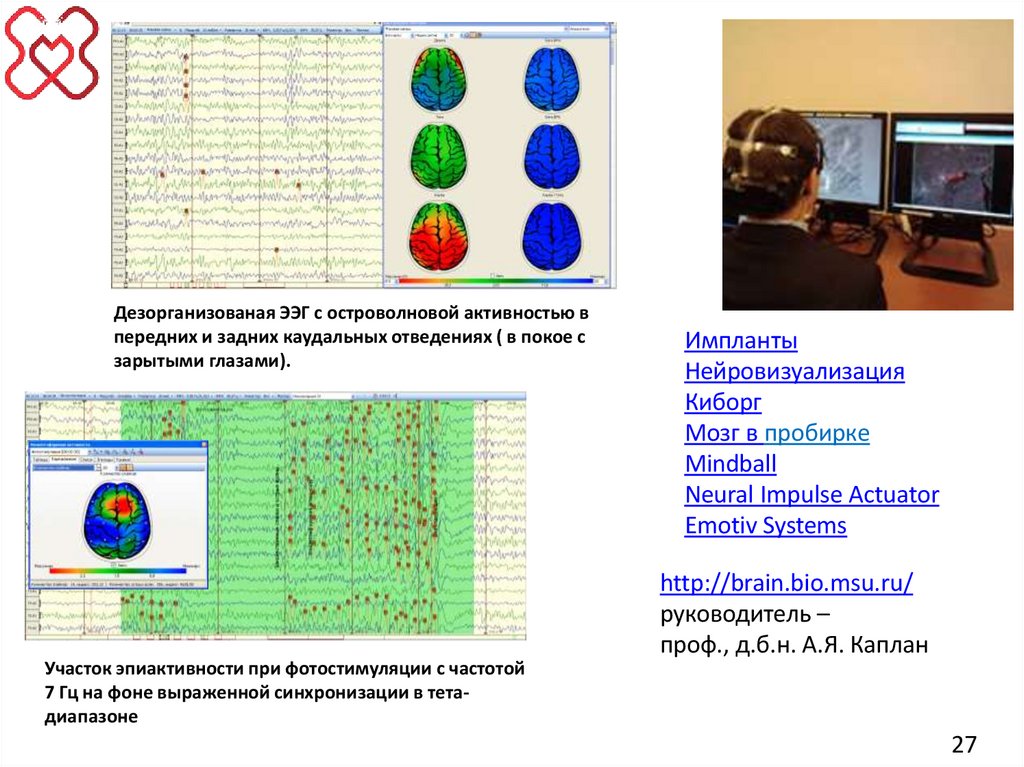
27.
Дезорганизованая ЭЭГ с островолновой активностью впередних и задних каудальных отведениях ( в покое с
зарытыми глазами).
Импланты
Нейровизуализация
Киборг
Мозг в пробирке
Mindball
Neural Impulse Actuator
Emotiv Systems
http://brain.bio.msu.ru/
руководитель –
проф., д.б.н. А.Я. Каплан
Участок эпиактивности при фотостимуляции с частотой
7 Гц на фоне выраженной синхронизации в тетадиапазоне
27
28. Функциональная магнитно-резонансная томография (фМРТ)
Функциональная магнитнорезонансная томография (фМРТ)• ФМРТ основана на парамагнитных
свойствах оксигенированого и
дезоксигенированого гемоглобина и
дает возможность увидеть изменения
кровообращения головного мозга в
зависимости от его активности. Такие
изображения показывают, какие
участки мозга активированы при
исполнении определённых заданий.
• Их изображение накладывается на
обычную МРТ мозга.
Разрешение ФМРТ 2—3 мм, ограниченное
кровоснабжением, влияющим на
нейрональную активность
Срез аксиальной МРТ на
уровне базальных ганглиев,
изображающий изменения
сигнала ФМРТ в красных
(увеличение уровня
оксигенации крови) и
голубых (его уменьшение)
тонах
28
29. Позитронно-эмиссионная томография (ПЭТ)
ПЭТ измеряет выброс радиоактивно меченныхметаболически активных химических веществ,
введённых в кровеносное русло.
• Информация обрабатывается
компьютером в 2-х или 3-мерные
изображения распределения этих
химических веществ в головном мозге.
• Обширные массивы лигандов для
создания карты разных аспектов
активности нейромедиаторов, для чего
чаще всего используют ПЭТ-индикатор
меченный формой глюкозы
(Фтордезоксиглюкоза (ФДГ)).
• Временное разрешение – 30 с
ПЭТ-скан здорового мозга в
возрасте 20 лет
29
30. ФИЗИОЛОГИЯ ПАМЯТИ
3031. Биологическая память – это фундаментальное свойство живой матери приобретать, сохранять и воспроизводить информацию
ПАМЯТЬГенетическая
Кратковременная
Нервная
Иммунная
Долговременная
Сенсорная
Первичная
Промежуточная
Вторичная
Третичная
Оперативная
образная
моторная
эмоциональна
я
смысловая
31
32.
• Генетическая - эволюционно наиболееранняя форма памяти. Материальным
носителем является геном - генетический
аппарат клетки, ДНК которой содержит
генетический код, обеспечивающий передачу
наследственных признаков.
• Иммунная - реализуется клетками иммунной
системы, лимфоцитами;
• Нервная - приобретается каждым организмом
в процессе его жизни
32
33. НЕРВНАЯ ПАМЯТЬ
• Нервная память - совокупность процессовфиксации, хранения и извлечения
информации, получаемой организмом в
течение жизни.
• Память, как результат обучения, заключается в
комплексе структурно-функциональных
изменений в ЦНС, называемых памятными
следами или энграммами, которые
сохраняются в течение некоторого времени
(секунды - года).
33
34. ВРЕМЕННАЯ ОРГАНИЗАЦИЯ ПАМЯТИ
• Концепция временной организации памятипринадлежит канадскому психологу
Д. Хеббу (D.Hebb).
• Память принято делить на
кратковременную и долговременную,
каждая из которых также имеет
разновидности.
34
35. Классификация видов долговременной памяти, предложенная И.С. Бериташвили (1968)
• образная (декларативная) - сохранение и репродукцияоднажды воспринятого важного объекта.
• моторная(процедурная, условно-рефлекторная) воспроизведение привычных заученных движений - игра на
музыкальном инструменте, работа с компьютером,
плавание, рисование, письмо и т. д.
• эмоциональная- воспроизведение ранее пережитого
эмоционального состояния - сдача экзамена, ссора с другом и
т. д.
• словесно - логическую (смысловая) - на словесные сигналы,
обозначающие как внешние объекты, так и внутренние
переживания и поведенческие реакции. Каждое знакомое
слово становится логической единицей информации.
35
36.
Фиксация образов, явлений происходит в три этапа:1. формирование энграммы,
2. сортировка и выделение новой информации,
3. долговременное хранение значимой для
организма информации.
Процесс запоминания может быть:
• произвольным
• непроизвольным.
36
37. МЕХАНИЗМЫ ПАМЯТИ
• Кратковременная, и долговременнаяпамяти имеют разную природу и
регулируются разными механизмами,
• Механизмы кратковременной, и
долговременной памяти связаны между
собой и являются последовательными
этапами единого процесса.
37
38. МЕХАНИЗМЫ КРАТКОВРЕМЕННОЙ ПАМЯТИ
• По современным представлениям в основекратковременной памяти лежат два
взаимно дополняющих друг друга
механизма:
1. реверберационный
(циркуляционный)
2. синаптический
38
39. Механизм кратковременной синаптической пластичности
- изменение амплитуды постсинаптического потенциала за счет либо
увеличения либо уменьшения величины секреции медиатора
(пресинаптический механизм):
Фасилитация (мс);
Усиление (с);
Посттетаническая потенциация (мин);
Депрессия (мс - мин).
Основной причиной является изменение концентрации
внутриклеточного кальция в пресинаптической клетке.
Т.о., механизмы кратковременной памяти представляют собой
возникновение непродолжительных обратимых изменений физикохимических свойств мембран и динамики медиаторов в синапсах,
временно перестраивающих деятельность нейронных цепей.
39
40. МЕХАНИЗМЫ ДОЛГОВРЕМЕННОЙ ПАМЯТИ
Механизм долговременнойсинаптической пластичности – изменение
амплитуды постсинаптического потенциала
за счет постсинаптических механизмов:
1. Долговременная потенциация (LTP);
2. Долговременная депрессия (LTD).
40
41.
• Основной причиной является изменениеконцентрации внутриклеточного кальция в
постсинаптической клетке:
1. Через NMDAзависимые каналы. Вход Са2+ зависит от
величины деполяризации, достаточной для снятия
Mg2+ блока в NMDA –зависимых каналах;
2. Через внутриклеточные депо кальция (ЭПР);
3. Через потенциалзависимые кальциевые каналы.
• Активация кальцием внутриклеточных протеинкиназ
(кальций-кальмодулинзависимые киназы – СаМКII,
цАМФ-зависимая протеин-киназа и др.) вызывает
фосфорилирование определенных белков: ионных
каналов (вход положительных ионов → ВПСП),
регуляторов активности ДНК (увеличение количества
постсинаптических рецепторов АМРА) → включение
молчащих синапсов
41
42. Модель формирования долговременной памяти (Анохин К.В., 1997):
1.2.
3.
4.
Активация кальцием внутриклеточных протеинкиназ (кальцийкальмодулинзависимые киназы – СаМКII, цАМФ-зависимая
протеин-киназа и др.) вызывает фосфорилирование белковрегуляторов экспрессии генов ДНК - ранних генов (c-fos и с-jun);
Активация генов приводит к синтезу белков-регуляторов FOS и JUN;
Экспрессия поздних генов – синтез морфорегуляторных белков;
Рост и изменение клеточных связей в мозге (дендритные шипики,
дополнительные синапсы и т.п.) – формирование долговременных
энграмм.
• Экспрессия ранних генов возникает на ранних этапах обучения в
ответ на новизну и с автоматизацией исчезает. Чем труднее идет
обучение, тем сильнее выражена их экспрессия («Генные
потенциалы»).
• При воспоминании экспрессия ранних генов возобновляется!!!. Если
заблокировать синтез белка в этот период, то можно стереть то, что
было в долговременной памяти.
42
43. ЛОКАЛИЗАЦИЯ СЛЕДОВ ПАМЯТИ
• Большинство нейрофизиологов полагает,что запечатление и извлечение энграмм
идет при участии тысяч нейронов КБП
(лобная доля, височная кора),
лимбической системы, таламуса и
гипоталамуса. По их мнению, следы
памяти распределены диффузно, но особую
роль в этих процессах играют два отдела –
гиппокамп и височная кора.
43
44. ФИЗИОЛОГИЯ МОТИВАЦИЙ
4445.
• Мотивация - стремление кудовлетворению потребностей (влечения,
побуждения, драйвы), имеющая
эмоциональную окраску.
• Различают:
1. низшие (биологические)
2. высшие (социальные).
45
46. ПЕРИФЕРИЧЕСКИЕ И ЦЕНТРАЛЬНЫЕ МЕХАНИЗМЫ МОТИВАЦИЙ
• Причиной мотиваций являютсяпреимущественно раздражители внутренней
среды – это изменения тех или иных гомео
• Афферентная импульсация от внутренних
органов и гуморальные факторы адресуются к
специализированным структурам головного
мозга, которые располагаются в основном в
области гипоталамуса, синего пятна и РФ
среднего мозга.
• Ведущая роль принадлежит
гипоталамусустатических показателей.
46
47. Мотивационные центры:
• 1. Центр голода - вентромедиальное ядро и латеральный ГПТ– нейроны которых избирательно чувствительны к изменению
содержания глюкозы в крови, а их электростимуляция вызывает
пищевую мотивацию.
• 2. Центр жажды – супраоптическое и паравентрикулярное
ядра ГПТ – реагируют на изменения осмотического давления
крови. Их электростимуляция вызывает питьевую мотивацию.
• 3. Центр страха – задний ГПТ. Нейроны этого отдела
откликаются на изменение содержания адреналина в крови.
Электростимуляция вызывает мотивацию страха.
• 4. Половой центр – передний ГПТ. Его нейроны контролируют
содержание половых гормонов.
Возбуждение, возникающее в гипоталамусе, по обширным связям
распространяется на лимбические структуры – перегородку,
гиппокамп, миндалины, а также на РФ, КБП (особенно на
передние ее отделы) и другие структуры головного мозга.
47
48.
• Мотивации – это особое интегрированноесостояние головного мозга, в которое каждая его
структура вносит свой специфический вклад.
Особая роль в этом состоянии отводится
гипоталамусу. Анохин и Судаков (1968)
сформулировали концепцию о «пейсмекерной»
роли гипоталамуса, подобной роли САУ в сердце.
Концепция базируется на том, что
• а) возбуждения в гипоталамусе возникают также
периодически (триггерно),
• б) нейроны гипоталамуса обладают повышенной
чувствительностью к гуморальным факторам,
• в) мотивационные центры гипоталамуса
подчиняют себе другие структуры головного мозга.
48
49. ФИЗИОЛОГИЯ ЭМОЦИЙ
4950.
• Эмоция (волнение, возбуждение) –субъективное состояние человека и
животных, которое возникает в ответ на
внешние и внутренние раздражения и
выражается в форме переживаний
позитивного «+» или негативного «-»
характера.
• «+» эмоции - радость, чувство комфорта,
удовольствие, наслаждение, любовь.
• «-» эмоции – страх, ужас, чувство
дискомфорта, неудовольствие, тоска,
негодование, гнев, ярость, ненависть.
50
51. Функции эмоций:
• Эмоции являются одним из главныхмеханизмов регуляции психической
деятельности, направленной на
удовлетворение потребностей.
• Эмоции лежат в основе обучения. Выполняя
функцию «+» или «-» подкрепления, они
способствуют выработке, закреплению и
сохранению биологически и социально
целесообразных форм поведения.
• Существуют аффекты, эмоции, чувства,
настроение.
51
52. ТЕОРИИ ВОЗНИКНОВЕНИЯ ЭМОЦИЙ
• 1. Периферическая теория (Джеймс – Ланге, 1885).Эмоции рассматриваются как следствие изменений
в деятельности внутренних органов и скелетной
мускулатуры. «…мы грустим, потому, что плачем,
сердимся, потому, что наносим удар, боимся,
потому что дрожим».
• 2. Центральная таламическая теория (Кеннон Бард, 1928). Нервные импульсы первоначально
поступают в таламус, откуда идут в кору
(субъективное переживание) и гипоталамус
(вегетативный компанент). Таламус
рассматривается как центр формирования эмоций.
52
53.
• 3. Центральная лимбическая теория Пейпс (Papez), 1937.Ведущая роль в формировании эмоций отдается лимбическим
структурам.
• Гиппокамп как место возникновения эмоционального
возбуждения →
• Мамиллярные ядра гипоталамуса →
• Передневентральное ядро таламуса →
• Поясная извилина – особая роль (субстрат осознанных эмоций)
→
• Гиппокамп.
• «Круг Пейпеца» (J. Papez) рассматривается как рецептивное
поле эмоций.
• Позднее МакЛин (McLean) в 1952 году предложил назвать
поясную извилину и связанные с ней структуры мозга
лимбической системой (limbus – край).
• В настоящее время к структурам, ответственным за
формирование эмоций, дополняют лобную и височную кору и
миндалину.
53
54.
• 4. Активационная теория (Линдсли, 1958). Подчеркивает роль РФ вформировании эмоций разного знака. Вызывают эмоции нейроны РФ,
импульсы от которой идут к таламусу, гипоталамусу, коре. Согласуется
с представлением о роли НА, дофаминергических,
серотонинергических и опиоидергических нейронов в развитии
эмоций.
• 5. Биологическая теория (Анохин,1949). Э рассматриваются как
средство оценки потребностей и их удовлетворения. Если
получаемый эффект совпадает с свойствами акцептора результата
действия (моделью будущего результата), то возникают «+»Э, при
несовпадении достигнутого с задуманным - «-»Э.
• 6. Потребностно-информационная теория (Симонов, 1966). Эмоции
рассматриваются как результат достаточности или дефицита
средств к удовлетворению потребностей. К средствам
удовлетворения потребностей относятся информация (умение, опыт),
энергия и время. Если уже существующие меньше необходимых,
возникает эмоциональное напряжение, и чем больше их дефицит, тем
сильнее состояние напряжения (СН).
54
55.
5556.
• Симонов различает четыре состоянияэмоционального напряжения (СН):
I СН
II СН –
стеническая
внимание,
мобильность,
активность
негодование,
гнев,
ярость
«вегетативная буря»
III СН –
тоска, ужас,
астеническая
хаос»
невроз
IV СНневротическая
«вегетативный
56
57. Периферические компоненты эмоций представлены:
1. произвольно управляемымипроявлениями, такими, как движения,
мимика, речь, дыхание, и
2. неуправляемыми (вегетативными):
частота и сила сердечных сокращений, АД,
деятельность желез, моторика ЖКТ и др.
57
58. Функций эмоций
• отражательная (оценочная),• побуждающая,
• подкрепляющая,
• переключательная,
• коммуникативная.
58
59.
РОЛЬ РЕТИКУЛЯРНОЙ ФОРМАЦИИ (РФ) ВИНТЕГРАТИВНОЙ ДЕЯТЕЛЬНОСТИ МОЗГА
59
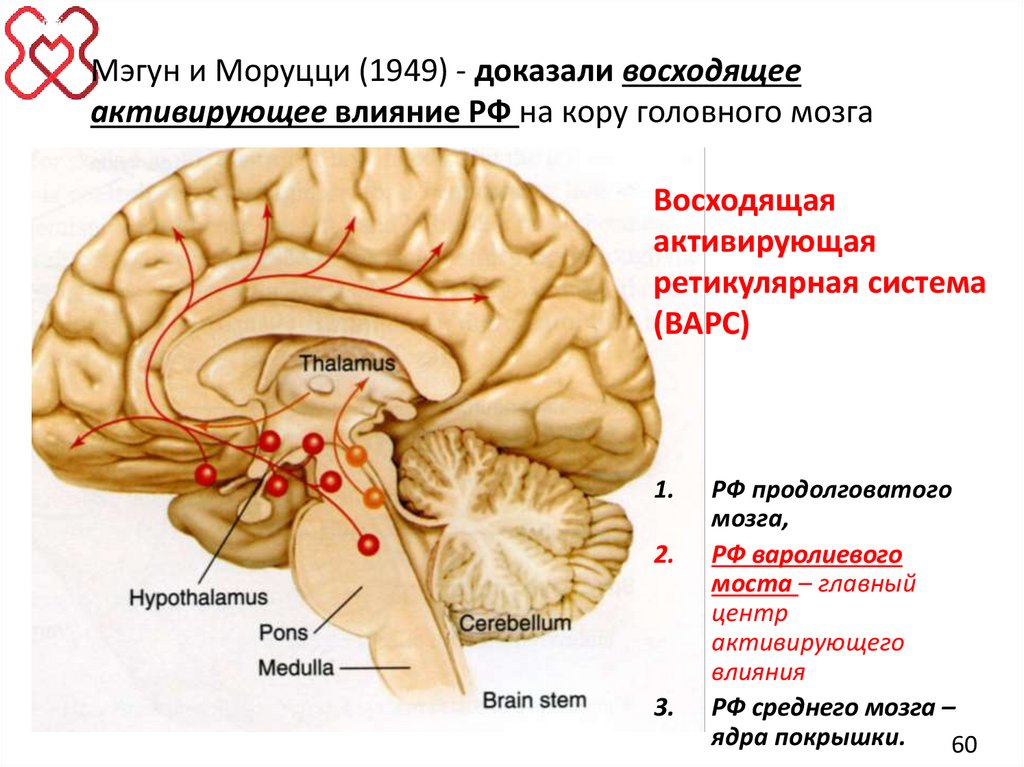
60.
Мэгун и Моруцци (1949) - доказали восходящееактивирующее влияние РФ на кору головного мозга
Восходящая
активирующая
ретикулярная система
(ВАРС)
1.
2.
3.
РФ продолговатого
мозга,
РФ варолиевого
моста – главный
центр
активирующего
влияния
РФ среднего мозга –
ядра покрышки.
60
61. Неспецифическая проекционная таламо-кортикальная система
Группа медиальныхядер таламуса,
составляет
неспецифическую
проекционную
таламокортикальную
систему (НПТКС)
(Мэгун и Моруцци ).
61
62. ФИЗИОЛОГИЯ СНА
• Благослови, господь, изобретшего сон.М. Сервантес де Сааведра. "Дон Кихот"
• Сон — это чудо матери природы, вкуснейшее
из блюд в земном пиру.
В. Шекспир
• Кто познает тайну сна, познает тайну мозга.
М. Жуве
62
63.
• Сон - активное физиологическое состояние,которое характеризуется потерей активных
психических связей субъекта с окружающим его
миром.
1.Рефлекторные реакции во время сна снижены, порог
раздражения рефлексов возрастает, а латентный период
удлиняется.
2.Сознание выключается, снижается мышечный тонус и все
виды чувствительности.
3.Вегетативные показатели (дыхание урежается, обмен веществ
и температура тела, частота сердечных сокращений, артериальное
давление и диурез) снижаются.
4.Мозговой кровоток повышается в ряде глубинных структур
головного мозга почти вдвое, что указывает на повышение
метаболизма.
63
64. Выделяют несколько видов сна:
1. Физиологический (периодическийежесуточный сон)
2. Не физиологический сон:
- наркотический сон, вызываемый
различными химическими или
физическими агентами,
- гипнотический сон;
- патологический сон.
64
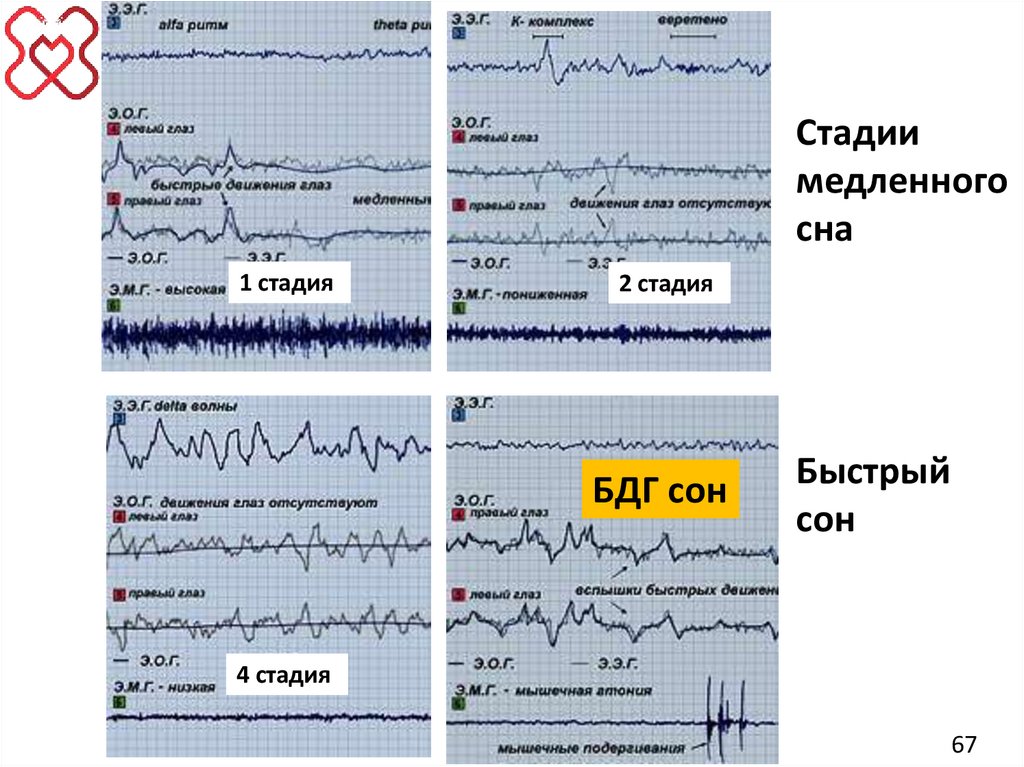
65. СТРУКТУРА СНА
"Медленный" сон:1. Стадия тета-волн.
2. Стадия «сонных веретен» и «К-комплексов».
3. Стадия дельта-волн и «сонных веретен».
4. Стадия дельта-волн.
"Быстрый" сон (БДГ- сон).
65
66. "Медленный" сон:
"Медленный" сон:1.
2.
3.
4.
Очень поверхностный сон. Высокоамплитудные медленные
тета-волны (3-7 Гц).
Поверхностный сон. Веретенообразные всплески бета-ритма
(сонные веретена 12-14 Гц) и К-комплексы. Пробуждение в эту
стадию затруднено, она сопровождается рядом изменений
вегетативных показателей: уменьшается частота сердечных
сокращений, снижается кровяное давление, температура тела
и др.
Умеренно глубокий сон. Высокоамплитудные медленные
дельта-волны с частотой 3,0-3,5 Гц.
Глубокий сон или дельта-сон. Сверхмедленные колебания
(0,7-1,2 Гц). Частота сердечных сокращений, артериальное
давление, температура тела в эту фазу достигают
минимальных значений.
66
67.
Стадиимедленного
сна
1 стадия
2 стадия
БДГ сон
Быстрый
сон
4 стадия
67
68. "Быстрый" сон (БДГ- сон)
"Быстрый" сон (БДГ- сон)• Основной отличительной особенностью "быстрого"
сна является наличие быстрых движений глазных
яблок (фаза быстрых движений глаз – БДГ).
• Кроме того, этот тип сна характеризуется:
– высоким порогом пробуждения,
– высокой активностью мозга,
– генерацией низкоамплитудных волн различных частот,
– полным подавлением тонуса скелетных мышц,
– сериями быстрых перемещений глазных яблок и
мелкими случайными подергиваниями мышц лица или
пальцев.
68
69. ТЕОРИИ СНА
Переход от бодрствования ко снупредполагает два возможных пути:
1. Пассивный, в результате утомления ЦНС
2. Активный, т.е. торможение механизмов
бодрствования
69
70. 1. Гипотеза гипнотоксинов или гуморальная теория
• Лежандр и Пьерон (1910)• Сон обусловлен накоплением в крови
особых продуктов метаболизма –
факторов сна, тогда как во время сна за
счет удаления или обменных процессов
восстанавливаются исходные
концентрации этих веществ, характерные
для бодрствования.
70
71.
Традиционные обсуждаемые нейрохимическиеагенты, имеющие значение в организации цикла "сонбодрствование":
• Фаза медленного сна: ГАМК, серотонин, норадреналин,
ацетилхолин, глутамат,
• Бодрствование: норадреналин, глутамат, ацетилхолин,
гистамин, серотонин.
Новые нейрохимические агенты, имеющие значение в
организации цикла "сон-бодрствование":
орексин
мелатонин,
дельта-сон-индуцирующий пептид,
аденозин,
простагландины (PGD2),
интерлейкины, мурамилпептид, цитокины.
71
72. Однако
• многие факты и, в частности, то, что можнопреодолеть сон интересным занятием или
то, что у сросшихся близнецов, имеющих
общее кровообращение, одна голова
может спать, а другая бодрствовать, делало
эту гипотезу не вполне состоятельной.
72
73. 2. Теория центров сна
Фон Экономо (1930); Гесс (1933)• передний гипоталамус
73
74. Однако
• в дальнейшем было показано, что состояние снаможно было получить при электростимуляции в
определенном ритме и других структур
головного мозга, что противоречит
представлениям о локальном нервном центре
сна.
• синхронизирующий центр Моруцци – область
средней части варолиевого моста в РФ,
• дорсальное ядро шва – медленный сон,
• голубое пятно - БДГ-сон, орбито-фронтальная
область коры – передний базальный мозг
• …….. ?
74
75. 3. Теория условного торможения
• И.П. Павлов даже сделал вывод о том, чтоусловное торможение и сон - один и тот же
процесс и считал, что торможение
развивается, охватывая основные корковые и
подкорковые структуры головного мозга
(1922).
• Диссертация "Материалы к физиологии сна",
написанная Рожанским Н. А. в 1913 г., была
выполнена под руководством И. П. Павлова.
75
76. 4. Теории деафферентации сенсорных систем
• Бремер, конец 1930-х годов• Для бодрствования необходим хотя бы
минимальный уровень активности коры,
поддерживаемый сенсорными стимулами,
а сон – состояние, обусловленное
снижением эффективности сенсорной
стимуляции мозга, т.е. деафферентацией.
Его опыты стали ключевым в пользу теории
пассивного засыпания.
76
77. Однако
• даже в изолированном переднем мозгепоявляются ритмичные колебания,
характерные для цикла сна/бодрствования.
• Кроме того, лишение человека сенсорных
стимулов постепенно уменьшают
длительность сна.
• Перерезка только сенсорных трактов,
проходящих через ствол мозга, не вызывает
постоянного сна
77
78. 5. Теория неспецифических регуляторов сна
Активирующие структуры:• ВАРС
• Неспецифическая и специфическая таламическая
система
• Ядра заднего гипоталамуса
• Структуры базального переднего мозга
• Структуры каудального ядра
• Структуры серого околоводопроводного вещества
• Черная субстанция
Гипоталамические центры за счет связей с лимбическими структурами
мозга могут оказывать восходящие активирующие влияния на кору мозга
при отсутствии влияний ретикулярной формации ствола мозга
78
79. Однако
• в ретикулярной формации существуют не толькоцентры бодрствования, но и сна. Каудальные отделы
оказывают тормозное влияние на ростральные.
• Кроме того, во время сна нейронная активность
ретикулярной формации (особенно в фазу БДГ-сна) не
отличается от бодрствования.
• Кроме того, в изолированном переднем мозгу
наблюдается чередование нейрофизиологических
процессов, характерных для сна/бодрствования, и
обусловленных структурами промежуточного мозга –
медиальным таламусом и передним гипоталамусом….
79
80. 6.Серотонинергическая теория
• М. Жуве - конец 60-х годов• В верхних отделах ствола мозга есть две области – ядра
шва и голубое пятно, у нейронов которых такие же
обширные проекции, как и у нейронов ретикулярной
формации, достигающие многие области ЦНС.
• Медиатором в клетках ядер шва служит серотонин, в
голубом пятне – норадреналин.
• Истощение запасов серотонина и НА и вызывает
бессонницу. Медленный сон регулируется
серотонинергической системой ядер шва, а БДГ-сон норадренергической системой нейронов голубого
пятна и РФ среднего мозга (центры быстрых
движений глаз). Активность ядер голубого пятна
приводит к торможению ядер шва и пробуждению.
80
81. Однако
• сейчас доказано, что нейроны ядер шванаиболее активны при бодрствовании и
серотонин большую роль играет не
только в регуляции цикла
сон/бодрствование
Фармакологический сон
неадекватен по своим
механизмам
естественному сну
81