






































 Биология
БиологияПохожие презентации:









 1728 ж. итальяндық ғалым Якопо Бартоломео Беккари (1682-1766) бидай ұнынан бөліп алған")
Зат алмасуға кіріспе. Жасуша биоэнергетикасы және катаболизмнің ортақ жолдары
1. Зат алмасуға кіріспе. Жасуша БИОЭНЕРГЕТИКАсы және КАТАБОЛИЗМнің ортақ жолдары.
• Негізгі сұрақтары:1. Пирожүзім қышқылының Қышқылды
декарбоксилдену.
2. Үшкарбонды қышқыл циклі
3. Қайықша механизмдер
4. Макроэргиялық байланыс.Ұлпаның тыныс
алуы туралы түсінік және биологиялық
қышқылдану.
5. Биологиялық тотығу
6. Митохондриялар құрылысы
7. Тыныс алу тізбегі, тіндік тыныс алу
8. Тотығып фосфорлану,ажыруы мен
қабысуы
2.
Спецификалық және жалпы катаболизмжолдары
• . Катаболитті процесті екі бөлімге бөлуге болады:
- катаболизмнің спецификалық жолы, Түрлі класс
байланыстары үшін әр түрлі (белок, май, көмірсу и др.),
- катаболизмнің жалпы жолы - Әр түрлі класстағы
заттардың катоболизмінің біртекті жолдары,
спецификалық жолдың жалғасы болып табылады.
катаболизмнің жалпы жолдары негізінен,негізгі
биоэнергетикалық процеспен байланысқан, клеткада
энергияның жинақталуы,босауы жүреді.
• Катаболизмнің спецификалық жолына нәруыз
гидролизі,көмірсу,липид реакциясы жатады,ол асқазанішек жолдарында түзіледі немесе ұлпада
түзіледі,моносахаридтердің,аминқышқылдарының,май
қышқылдарының,спирттің бұзылу реакциясы жасушада
жүзеге асады және соңында пирожүзім қышқылы немесе
ацетил радикалы ацетил КоА формасы түрінде пайда
болуымен аяқталады.
3. Спецификалық және жалпы катаболизм жолдары
Цикл Кребса:1, 6 – система окисления и декарбоксилирования; 2 – цитрат-синтеза: кофермент А; 3, 4 –
аконитат-гидратаза; 5 – изоцитратдегидрогеназа; 7 – сукцинатдегидрогеназа; 8 – фумаратгидрогеназа; 9 – малатдегидрогеназа; 10 – спонтанное превращение; 11 –
пируваткарбоксилаза
4.
Пирожүзім қышқылының қышқылдыдекарбоксилденуі.
• Пируваттың қышқылды декорбоксилденуі матрикс
жағындағы митохондрияның ішкі мембранасында
мультиэнзим пируват-дегидрогеназды комплекс
арқылы катализденеді.
• Пируват-дегидрогеназды комплекс (пирожүзім
қышқылының қышқылдық декарбоксилаза ) үш
ферменттен тұрады (Е1,Е2,Е3):
• Е1-пируватдекарбоксилаза коферментпен ТПФ
(тиаминпирофосфат),
• Е2-дигидролипоил-трансацетилаза коферментпен
липоевая кислота (ЛК) и кофактором - НS-КоА,
• Е3-дигидролипоил-дегидрогеназа с коферментом
ФАД и кофактор – НАД.
5. Пирожүзім қышқылының қышқылды декарбоксилденуі.
• Пирожүзім қышқылы (пируват) және ацетилКоА - катаболизмнің жалпы жолы қосылғанкезде көмірқышқыл газ бен суға дейін
ыдырайтын,митохондрияда АТФ синтезі жүру
үшін энергия түзетін метоболиттер.
• Катаболизмнің жалпы жолына келесі
метоболиттік процесстер жатады (бұл екі
процесс митохондрияда локализденген):
• - Пирожүзім қышқылының қышқылды
декарбоксилденуі,
• - Ацетил-КоАның Кребс циклінде
үшкарбонды қышқылға айналуы (Ц Т К ).
6.
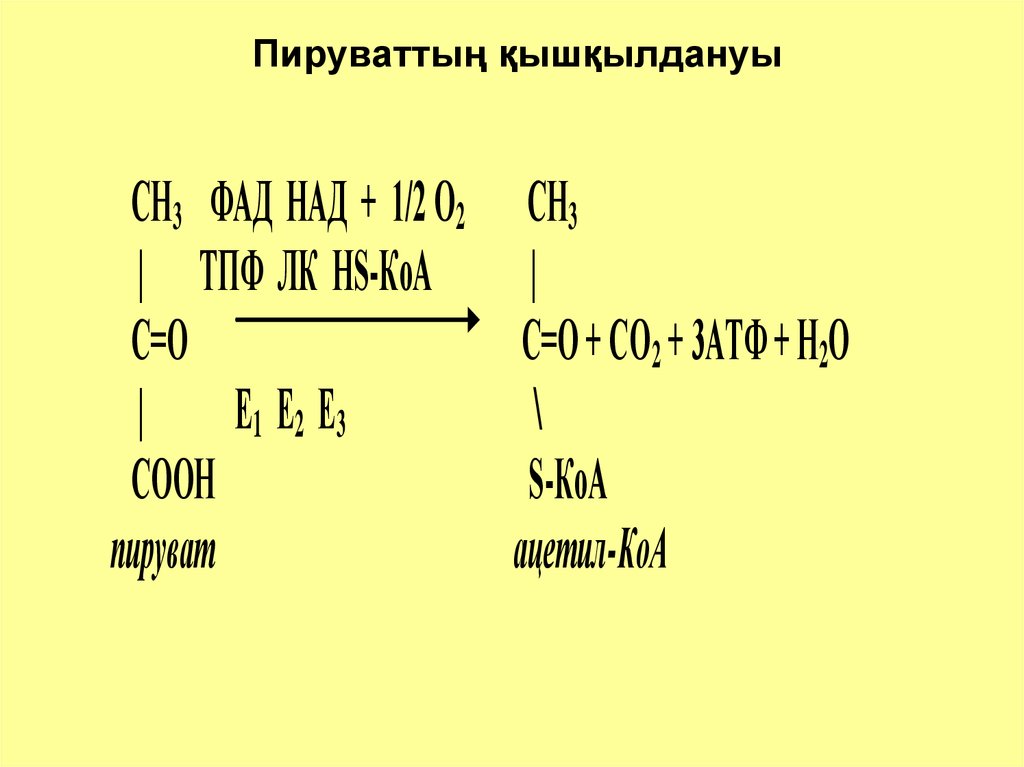
Пируваттың қышқылдануыСН3 ФАД НАД + 1/2 О2 СН3
| ТПФ ЛК НS-КоА
|
С=О
С=О + СО2 + 3АТФ + Н2О
|
Е 1 Е2 Е3
\
СООН
S-КоА
пируват
ацетил-КоА
7. Пируваттың қышқылдануы
Пируватдекарбоксилаза әсерінің нәтижесінде тиаминопирофосфаттың қатысуыменпируваттан карбоксильді топ бөлініп шығады да,нәтижесінде көмірқыышқыл газы пайда
болады.Ал ацетильді қалдық күкірт атомына қосылады:
S-S
СН3
CН3
||
|
| ТПФ Л-К
С=О
С=О
\
+ СО2
|
E1
S SH
СООН
||
пируват
Л-К
8.
Екінші энзим Е2 - дигидролипоилтрансацетилаза - ацетильді радикалдың НS-КоАға ауысуын катализдейді. Осының нәтижесінде дигидролипоевая кислота босап шығадыжәне ацетил-КоА пайда болады :
СН3
|
СН3
С=О НS-КоА |
НS SH
\
С=О + | |
S SH E2
\
Л-К
||
S-КоА дигидролипоевая
Л-К
ацетил-КоА кислота
ацетил-дигидролипоевая
кислота
9.
Үшінші сатыда Е3ферментінің әсерлесуінен - дигидролипоилдегидрогеназ ФАД пенНАДқатысында дигидролипоевойқышқылдың қышқылдануы және НАДтың қалыпқа келуі жүреді:
НS SH ФАД НАД S-S
| |
| | + НАДН2
Л-К
Е3 Л-К
Содан соң, қалпына келген НАД митохондриядағы протондар мен электрондар мен протондар
тізбегінде қышқылданады да су пайда болады және қышқылды фосфорилдену реакциясында 3
молекулалы АТФ синтезделеді.
10.
Үшкарбонды қышқыл цикліСН3
|
С=О +
\
S-КоА
ацетил-КоА
СООН
|
С=О
|
СН2 цитратсинтетаза
|
СООН
оксалоацетат
( ЩУК)
S-КоА
/
С=О
|
HS-КоА
СН 2
+ НОН
|
НО-С-СООН цитратсинтетаза
|
СН2
|
СООН цитрил-КоА
СООН
СООН
|
|
СН2
СН2
|
-НОН |
+НОН
НО-С-СООН
С-СООН
|
||
СН2
аконитаза СН
аконитаза
|
( Fe++ )
|
( Fe++ )
СООН
СООН
цитрат
цис-аконитат
СООН
|
СН2
|
Н-С-СООН
|
Н-С-ОН
|
СООН
изоцитрат
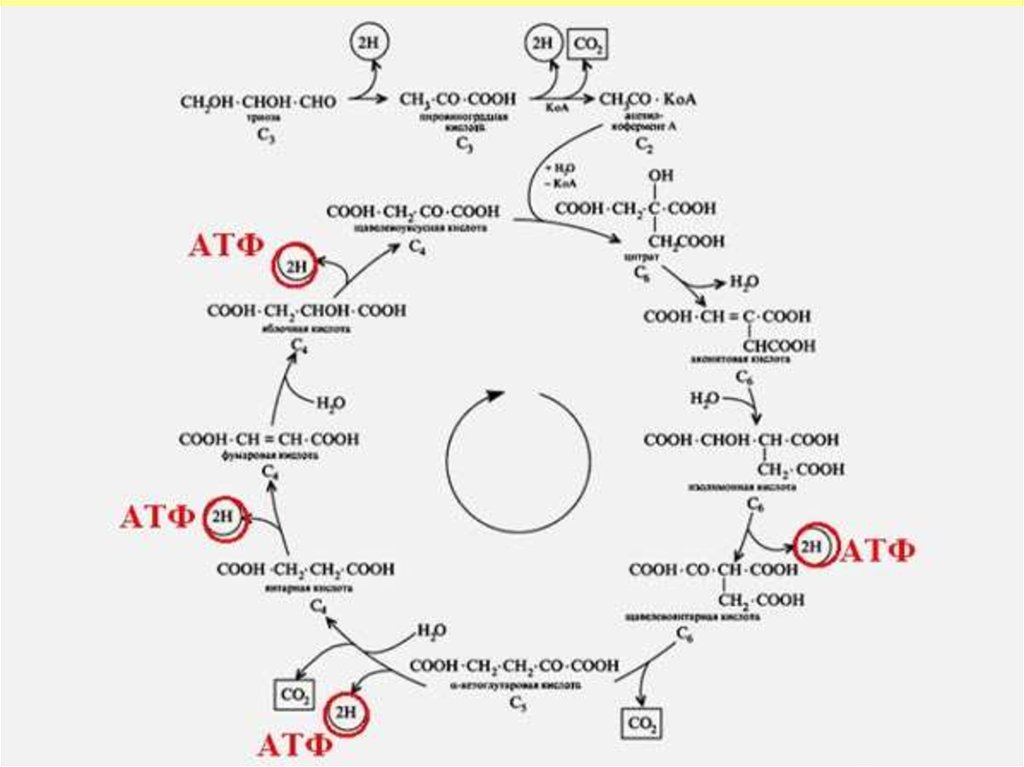
11. Үшкарбонды қышқыл циклі
Үшкарбон қышқыл цикліСООН
СООН
СООН
|
НАДН2
|
|
СН2
СН2
CO2
СН2
СО2
|
НАД
|
|
Н-С-СООН
Н-С-СООН
СН2
|
( Мn++) изоцитрат
|
изоцитрат
|
( Е 1 ` Е2 Е 3 )
Н-C-ОН
дегидрогеназа
С=О дегидрогеназа С=О
|
|
|
СООН
СООН
СООН
изоцитрат
оксалосукцинат
альфа-кетоглутарат
Содан соң -кетоглутаратТЫҢ ТҮЗІЛУІМЕН ИЗОЦИТРАТТЫҢ ҚЫШҚЫЛДАНУ РЕАКЦИЯСЫ ЖҮРЕДІ, кетоглутарат дегидрогеназнды мультиэнзимді комплекстің қышқылдануына әкелетін (Е1,Е2,Е3),
Е1`- -кетоглутаратдекарбоксилаза, Е2-дигидролипоилтрансацилаза,
Е3- дигидролипоилдегидрогеназа:
НАДН2
ФАД НАД
ТПФ ЛК НS-КоА
СООН
ГТФ
|
HS-КоА
СН2 Рн + ГДФ
|
СН2 сукцинаттиокиназа
|
С=О
\
S-КоА
сукцинил-КоА
СООН
|
СН2
|
СН2
|
СООН
сукцинат
12. Үшкарбон қышқыл циклі
Кребс цикліСукцинил-КоА
макроэрг
секілді,
сукцинаттың
түзілуімен
сукцинаттиокиназаға дейін ыдырайды. Энергия байланысы ГДФтың ГТФ
ға дейінгі субстратты фосфорленуінде қолданылады. Нуклеозид
дифосфаткиназа әсерінен фосфор қышқылының қалдығы ГТФ тың АДФқа
ауысуы жүреді және АТФсинтезделеді.
СООН ФАДН2
СООН
СООН НАДН 2 СООН
|
|
|
|
СН2 ФАД
СН + НОН
НО-С-Н НАД
С=О
|
||
|
|
СН2 СДГ(Fe)
СН фумараза
СН 2 МДГ
СН2
|
|
|
|
СООН
СООН
СООН
СООН
сукцинат
фумарат
малат
оксалоацетат
(ЩУК)
Кребс циклі оксалоацетаттың регенерациясымен және ацетильді
радикалдың көмірқышқыл газ бен суды қышқылдатуымен аяқталады.
13. Кребс циклі
Энергияның шығуы• Сент-Дьёрди – Кребс үшкарбон қышқыл
циклінде субстратты сәйкес
дегидрогеназамен дегидрлейтін 4 реакция
орны бар:
• изоцитратдегидрогеназа, кофермент НАД,
• альфа-кетоглутаратдегидрогеназы, кофактор
НАД
• сукцинатдегидрогеназа, кофермент ФАД,
• малатдегидрогеназа, кофермент НАД.
• Қышқылдану кезінде энергияның шығуы ЦТК - 12 АТФтағы КоА-ның 1 молекуласы
• Барлық реакцияларда катоболизмнің
жалпы жолы АТФ тың 15 молекуласы
арқылы синтезделеді.
14. Энергияның шығуы
15.
16.
17.
Транспорттың қайықшамеханизмдері
• Топтық транслокацияның өзіндік нұсқасы
болып протондардың (Н+) митохондриялы
мембрана арқылы көшу механизмі саналады.
• Митохондрия мембранасы НАД Н+
молекуласының қышқылдануынан цитозолда
түзілетін әртүрлі субстарттар үшін не
проницаемы.
• НА Д Н+ эквивалентінің қалпына келе
отырып көшуі қосалқы жолмен жүзеге асады
яғни,
• - глицерофосфатты және малатты
челночными механизмами транспорта
18. Транспорттың қайықша механизмдері
19.
Малатный челнокцитоплазма
мембрана Митохондриальды матрикс
малат
Малат
НАД
(Т1)
α-кетоглутарат
(Е1)
α-кетоглутарат
(E3)
глутамат
НАДН2
Оксалоацетат
НАД
(E4)
глутамат
(Т2)
аспартат
(Е2)
Аспартат
НАДН2
оксалоацетат
20. Малатный челнок
Тотығудың негізгі түрлері• Органикалық байланыстың
(субстрат)катоболизм реакциясы
биосистемада бос энергияның
босатылуымен байланысты, негізінде
қышқылды-қалпына келгіш реакция болып
табылады және оксидоредуктаз класының
ферментімен катализденеді және клетканың
әр түрлі компартментінде локализденеді:
• Оксидазды - митохондрияда
• Микросомальды - эндоплазматикалық
ретикулум мембранасында
• Пероксидті- пероксиомада
21. Тотығудың негізгі түрлері
Маркоэргтер• Жануар организмінің энергетикалық субсттратына
көмірсу,липид,аминоқышқылдар жатады. көмірсудың
калориясы 4,1 ккал/г, липидтікі - 9,3 ккал/г және
нәруыз(аминқышқыл) - 4,1 ккал/г.
• Бос энергияның негізгі энергетикалық субстраттан босап
шығуы катаболит процессі кезінде жүреді. Сонымен
қатар,бос энергия кейбір фосфоорганикалық
қосылыстардың макроэгиялық
байланыстарында(макроэргада) жинақтала алады.
• Макроэргиялық байланысқа АТФжәне басқа
нуклеотидтер;-5`-үшфосфат (ГТФ, УТФ, ЦТФ),
креатинфосфат, 1,3- дифосфоглицерат,
фосфоенолпируват жатады.
• Барлық макроэргалардың ішінде АТФ - жасушаның
негізгі химиялық тасымалдаушысы, энергияның шығуы
мен жұту процесстерін өзара байланыстыратын
(катаболизм және анаболизм), фосфатты топтардың
энергия тасымалдау реакциясындажалпы ортақ өнім
қызметін атқарады.
22. Маркоэргтер
Клеткада энергияның трансформация механизміКөмірсу катоболизмі кезінде бос энергияның,липид
және аминқышқылдың босап шығуы, аэробты және
анаэробты жағдайда жүзеге асады.
• Энергиялық жол аэробты жағдайда қолайлы жол болып
табылады, яғни,ұлпаның ауаны жұту және көмірқышқыл
газын шығару сияқты міндетті қызметтерді орындайд.Ол
ұлпалық тыныс алу,ішкі тыныс алу немесе жасушалық
тыныс алу деп аталады.
• Органикалық байланыстың (субстрат)катоболизм
реакциясы биосистемада бос энергияның босатылуымен
байланысты, негізінде қышқылды-қалпына келгіш
реакция болып табылады және оксидоредуктаз
класының ферментімен катализденеді және клетканың
әр түрлі компартментінде локализденеді.Осыған
байланысты бұл процесс- биологиялық қышқылдану деп
аталады.
23. Клеткада энергияның трансформация механизмі
24.
Биологиялық тотығумеханизмі
Биологиялық қышқылдану негізіне субстрат дегидрлену реакциясы
жатады,протондардың және электрондардың су түзу арқылы кислородқа
ауысуы және бос энергияның бөлінуі:
+
2Н +2еSН2
2Н
S
__
½О2 O
(ΔG)
Н 2О
25. Биологиялық тотығу механизмі
Бұл процесстің негізгі қызметі организмді
энергиямен қамтамасыз ету. (АҮФ түрінде)
Субстраттың молекулалы кислородқа ауысу
процессіне мыналар қатысады:
1.Коферменттері НАД немесе НАДФ болып
табылатын-Пиридинзависимые анаэробные
дегидрогеназы.
2.Коферменттері ФАД немесе ФМН болып
табылатын-Флавинзависимые аэробные
дегидрогеназы;
3.Цитохромдар, кофермент ретінде
железопорфинді қолданады.
4.Убихинон (коэнзим Q) және ақуыздар,
құрамына негемовое железо кіреді.
26.
Анаэробты дегидрогеназалар• Бұл ферменттер сутегі атомдарының(протондар және
электрондардың) оттектебасқа кез-келген акцепторға
ауысуын катализдейді
• Представители:
• альфа-кетоглутарат дегидрогеназа
• Пируват дегирогеназа
• Изоцитратдегирогеназа
• Малатдегирогеназа
• Бұл ферменттердің коферменті НАД, НАДФ болып
табылады.
27. Анаэробты дегидрогеназалар
CONH2CONH2
N
CH2
O
O
P
O
o
OH
H
P
O
H
O
H
NH2
OH
N
O
P
O
OH
H H
OH OH
OH
N
N
O
N
CH2
OH
( НАД+)
H
OH
N
O
N
O
H
NH2
N
N
CH2
CH2
O
H OH OH
O
O
N
H
OH O
O
H
P
OH
OH
( НАДФ+)
28.
АЭРОБТЫ ДЕГИДРОГЕНАЗАЛАР• Бұл ферменттер сутегі атомдарының(протондар және
электрондардың) кез-келген акцепторға,сонымен қоса
оттекке де ауысуын катализдейді
• Представители:
• сукцинатдегидрогеназа
• глицероосфатдегирогеназа
• Ацил-КоА-дегирогеназа
• Бұл ферменттердің коферменті болып витамин В2
өндіретін ФАД,ФМН болып табылады.
29. АЭРОБТЫ ДЕГИДРОГЕНАЗАЛАР
HС
N
С
С
С
NH
H3 С
С
С
С
С
N
N
С
С
С
NH
Н3 С
С
С
С
С
N
С
H
CH2
Н
ОН
Н
ОН
Н
H C OH
OH
О
N
CH
OH
О
NH2
H C OH
O P
ОН
СН2
H C OH
O
N
СН2
O
N
CH2
С
Н3 С
С
H3 С
С
H
H
С
C
C
N
C
N
О
N
Н
Р
ОН
О
О
Н
ОН
О
CH
ФМН
Р
Н
СН2
Н
ФАД
O
30.
Кофермент QТыныс алу тізбегінің компонентіубихинон,бензохиноннан өндіріледі және
ФАДН2 –дегидрогеназ коферменті болып
табылады, протондар мен электрондардың
цитихромды жүйедегі ферменттерге
тасымалдануын катализдейді.
Убихинон қышқылдану процессіне де,қалпына
О
ОH
келу процессіне
де қатысады.
С
H3 С
С
С СН3
H3 С
С
С R
С
O
+
2H+
+
2е-
С
H3 С
С
С СН3
H3 С
С
С R
С
OH
31. Кофермент Q
ЦитохромдарЭлектрондардың убихиноннан кислородқа тасымалдануын цитохромдар
жүйесі жүзеге асырады :
b, c1, c,
a- a3.
Олардың барлығы кофермент ретінде кездесетін геминовты группаға ие
Катализдік процесс барысында цитохром құрамындағы темір
валенттілігін өзгертеді:
Fe++
Fe+++
Әр цитохромда қышқылды-қалпына келу потенциалының мөлшері әр
түрлі.Қышқылдану тізбегінде олар убихинон мен кислородтың
арасына орналаады:
b
С1
С
aa
3
b, c1, c цитохромдары электрондарды аралық тасымалдау қызметін
атқарады, ал аа3 цитохромы тыныс алу ферменті болып
табылады,оттекпен әрекеттеседі.
32. Цитохромдар
Цитохромның қышқылдану-қалпына келуреакциясына әсер ету механизмі
Fe3+ + eFe2+
Fe2+ - e Fe3+
Cu2+ + e Cu+
Cu+ - e Cu2+
Цитохром b және с1 КоQH2-дегидрогеназа ферменті ретінде әсер етеді және
реакция катализдейді:
КоQH2 + 2C1(Fe3+)
КоQ + 2H+ + 2C1 (Fe2+)
Аа3 цитохромдары цитохромоксидаза ретіне әсер етеді. Бұл соңғы өнім:
2аа3 (Fe2+) + О2
2аа3 (Fe3+) + 2O-2O-- + 4e- + 4H+
2H2O
Представители цитохромов:
аскорбат ДГ,
адреналин ДГ,
норадреналин ДГ.
33. Цитохромның қышқылдану-қалпына келу реакциясына әсер ету механизмі
Тыныс алу тізбегінің құрылымы• Электрондар мен протондардың митохондриальды тыныс
алу тізбегіне тасымалдануы қышқылдану-қайта қалпына
келу потенциалы бойынша жүзеге асады.
• Тыныс алу тізбегінің әрбір ферментінің өзіне тән редокспотенциалы бар.Оның мөлшері НАД (-0,32в) ден
О2(+0,82в) дейін өседі,және олардың редокс
потенциалының айырымы 1,14 Вольт тең.
• НАДН2 субстратының 1 молекуласы қышқылдануынан
босап шығатын энергия мөлшері 3 АТФ синтездеуге
жеткілікті.
• Тыныс алу тізбегінде фосфорилдеуге жеткілікті 3 бөлім
бар.
34. Тыныс алу тізбегінің құрылымы
ТЫНЫС АЛУ ТІЗБЕГІ2н+2е
SH2
НАД
2Н+
ФАД
2Н+
КоQ
2е
a
2
1
3
b c1 c
2е
(ФАДН2)
(НАДН2)
a3
2е
1/2O2
O+H
Q + H 2O
2е
35. ТЫНЫС АЛУ ТІЗБЕГІ
ЖӘНЕ РЕДОКСПОТЕНЦИАЛ
Митохондриальді тыныс алу тізбегінің ферменттері :
SmH2---->ФАД
SnH2
- 0.05в
SхH2-->НАД-->ФМН--->KoQH2---->b----->c1------>c------->a------->a3------->½O2
- 0,41в - 0,32в - 0,11в
+ 0,02в + 0,04в+ 0,23в + 0,25в + 0,29в + 0,55в
/\
АДФ+Рн=АТФ
/\
АДФ+Рн=АТФ
+ 0,82в
/\
АДФ+Рн=АТФ
36. ТЫНЫС АЛУ ТІЗБЕГІ ЖӘНЕ РЕДОКС ПОТЕНЦИАЛ
ФОСФОРИЛДЕНУ ЖОЛДАРЫФосфорилденудің бірінші жолы НАД және ФАД
ферменттерінің арасында жүзеге асады,және олардың
потенциалдарының мәні 0,246 тең
АДФ + Н3РО4
АТФ
Екінші жолы В және С1 цитохромдарының арасында жүзеге
асады, олардың арасындағы потенциал мәні 0,19
АДФ + Н3РО4
АТФ
Үшінші жолы аа3 және 1/2О2 цитохромдарының арасында
жүзеге асады, олардың арасындағы потенциал мәні 0,30
Вольт
АДФ + Н3РО4
АТФ
37. ФОСФОРИЛДЕНУ ЖОЛДАРЫ
Глицерол-3-фосфатКо-А-производные
Май қышқылдары
ФП4
ФП3
Изоцитрат
сукцинат
ФП2
Малат
Аскорбат
Пируват
АТФ
Глутамат
НАД
АДФ
1
-
ФП
АТФ
КоQ
b
АДФ
2
-
АТФ
C1
a3
c
a
АДФ
3
-
Аминобарбитал, ротенон
Антимицин
Цианид
Локализация трех пунктов фосфорилирования в цепи (1,2,3)
О2
38.
Тотығып фосфорлану• Бір субстрат молекуласының қышқылдануынан
митохондрияға бір атом оттегі енгізіліп, бір молекуладан
үш молекулалы фосфорлы қышқылға дейін қолданыла
алады және 1,2 және 3 молекулалы АТФсинтезделеді.
• Бұл процесс - АТФ синтеаза процессі субстраттың
биологиялық қышқылдану реакцияларында қышқылды фосфорилену атына ие болды.
• Қышқылды фосфорилденудің сандық көрсеткішін
белгілеу үшін - коэффициент Р/О қолданылады.
• Коэффициент Р/О (АДФ/О) - фосфорлы қышқылдың
молекула санына қатынасы (АДФ), митохондрияда белгілі
бір субстраттың қышқылдануы оттек атомының санына
қатынасы.
39. Тотығып фосфорлану
40.
П. Митчелдің гипотеза ұсынысына орай, фосфорилденетінАДФ қозғалыс күші әр түрлі редокс-потенциалдар
энергиясында қолданылады және ол электрондарды
қышқылдайтын субстраттан тыныс алу тізбегінің бойымен
оттекке тасымаладау барысында пайда болады.
(+)
2(Н)+
(-)
A--
SхН2
Sх
2 е( H+)
+
Н АТФ-аза
АДФ
+ Н3РО4
AН2
A
(2Н+)+ 2ОНАТФ
2(Н2О)
41. П. Митчелдің гипотеза ұсынысына орай, фосфорилденетін АДФ қозғалыс күші әр түрлі редокс-потенциалдар энергиясында қолданылады
Протонды (трансмембранды )электрохимиялық потенциал ( H+)
• Трансмембранды электрохимиялық
потенциал, мембранада Н+-градиент
үлесінен пайда болған,жасушада энергия
универсальды форма болып табылады және
мыналарда қолданыла алады:
- АТФ синтезі үшін
- ион транспорты үшін Са және басқа
иондар транспорты үшін
- фосфаттар транспорты үшін
- АДФ және АТФ транспорты үшін
• - жылутүзуүшін
• - қозғалмалы белсенділік үшін.