




















































")


























")
")














")




























 Биология
БиологияПохожие презентации:

")








Углеводы и их метаболизм
1. Углеводы и их метаболизм
1. Классификация и строение углеводов.2. Основные моносахариды.
3. Основные дисахариды.
4. Основные полисахариды.
5. Переваривание углеводов. Особенности у жвачных.
6. Перенос глюкозы в клетки.
7. Метаболизм глюкозы в клетке.
8. Фосфорилирование и дефосфорилирование глюкозы.
9. Метаболизм гликогена.
10. Регуляция уровня глюкозы.
11. Гликолиз
2. Определение и организация углеводов
Термин «углеводы», предложенный в XIX столетии, былоснован на предположении, что все углеводы содержат 2
компонента — углерод и воду, и их элементарный состав
можно выразить общей формулой Cm(H2O)n. Хотя из этого
правила есть исключения и оно не абсолютно точно, тем
не менее указанное определение позволяет наиболее,
просто характеризовать класс углеводов в целом.
Углеводы можно разделить на 3 основные группы в
зависимости от количества составляющих их мономеров:
моносахариды, олигосахариды и полисахариды.
3. Функции углеводов
1. Энергетическая2. Структурная
3. Защитная
4. Входят в состав НК и коферментов
5. Образуют смешанные биополимеры
У животных – до 2, у растений – до 80%
сухой массы
4. Классификация углеводов
5. Моносахариды
Моносахариды — производные многоатомных спиртов,содержащие карбонильную группу. В зависимости от
положения
в
молекуле
карбонильной
группы
моносахариды подразделяют на альдозы и кетозы.
Альдозы содержат функциональную альдегидную группу
—НС=О, тогда как кетозы содержат кетонную группу
>С=О. Название моносахарида зависит от числа
составляющих его углеродных атомов, например
альдотриозы, кетотриозы, альдогексозы, кетогексозы и
т.д.
6. Альдозы и кетозы
7. Триозы, тетрозы, пентозы, гексозы
8. Триозы: альдозы и кетозы
9. Изомеры
10. Гексозы и пентозы
11. Основные моносахариды
12. Химические свойства моносахаридов
13. Аминосахара
14. Фосфаты сахаров
15. глюкоза
Глюкоза является альдогексозой. Она может существоватьв линейной и циклической формах. Циклическая форма
глюкозы, предпочтительная в термодинамическом
отношении, обусловливает химические свойства глюкозы.
Как и все гексозы, глюкоза имеет 4 асимметричных
углеродных атома, обусловливающих наличие стереоизомеров. Возможно образование 16 стереоизомеров,
наиболее важные из которых D- и L-глюкоза. Эти типы
изомеров зеркально отображают друг друга.
16. Изомеры глюкозы
17. Семейство глюкозы
18. Образование гликозидов.
Гликозидная связь имеет важное биологическое значение,потому что именно с помощью этой связи осуществляется
ковалентное связывание моносахаридов в составе олиго- и
полисахаридов. При образовании гликозидной связи ОНгруппа одного моносахарида взаимодействует с ОНгруппой другого моносахарида или спирта. При этом
происходят отщепление молекулы воды и образование
гликозидной связи.
19. Этерификация.
Это реакция образования эфирной связи междуОН-группами моносахаридов и различными
кислотами. В метаболизме углеводов важную
роль
играют
фосфоэфиры
—
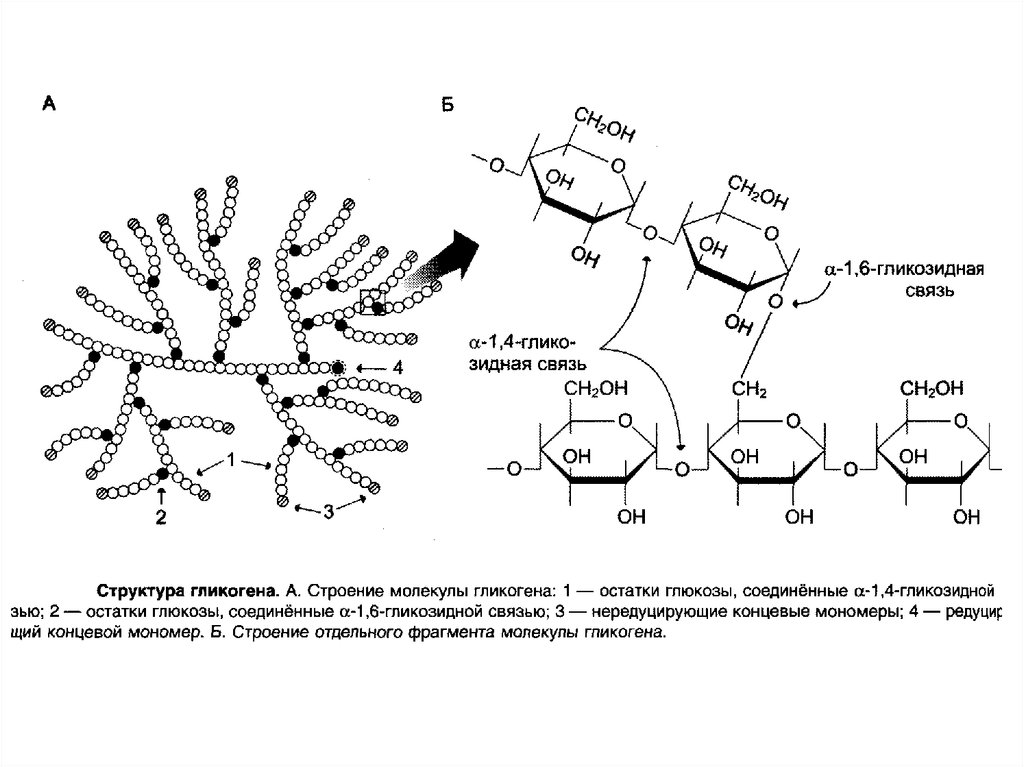
эфиры
моносахаридов и фосфорной кислоты. В
метаболизме глюкозы особое место занимает
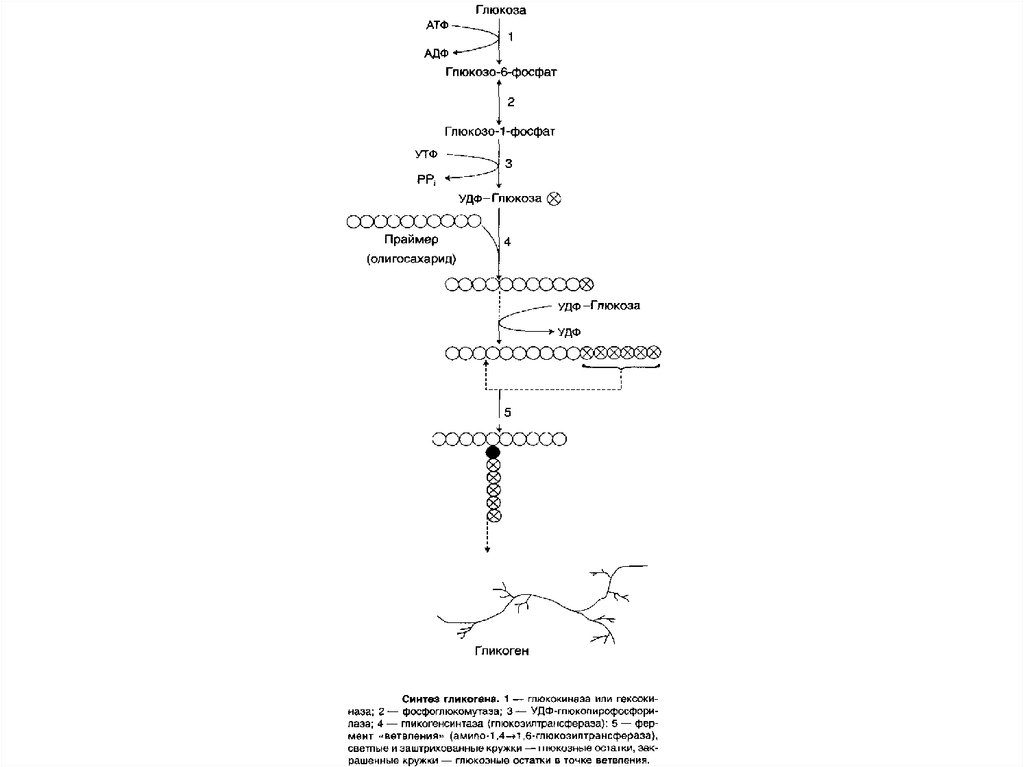
глюкозо-6-фосфат.
Образование
глюкозо-6фосфата происходит в ходе АТФ-зависимой
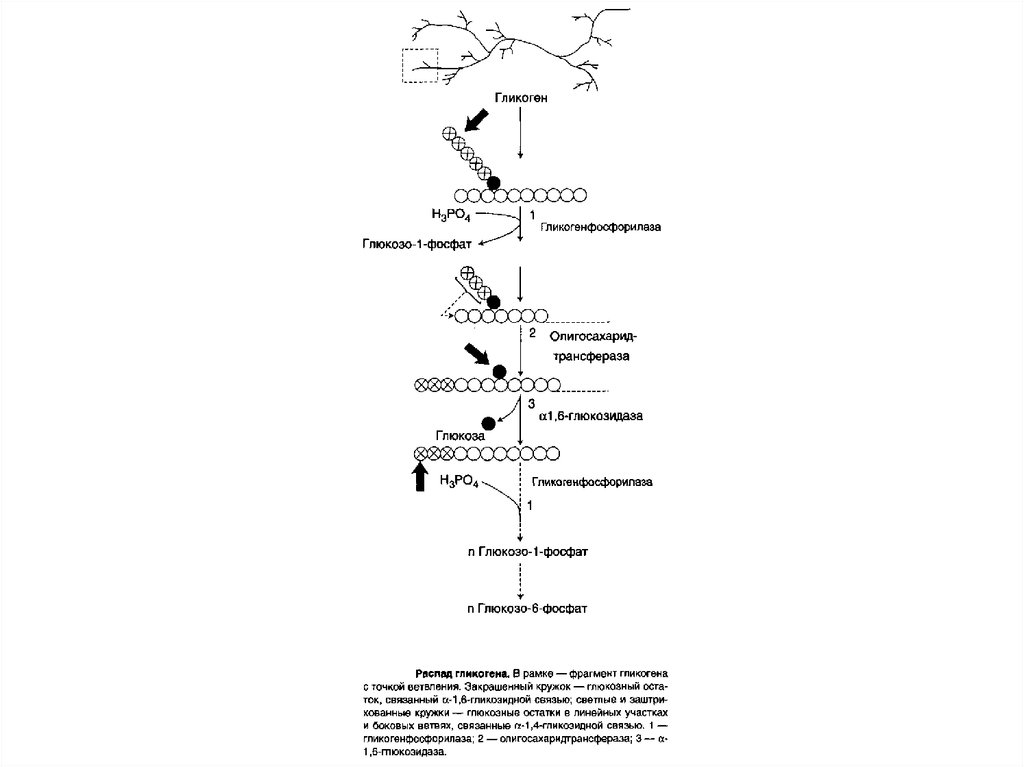
реакции при участии ферментов, относящихся к
группе киназ. АТФ в данной реакции выступает
как донор фосфатной группы.
20. Окисление и восстановление.
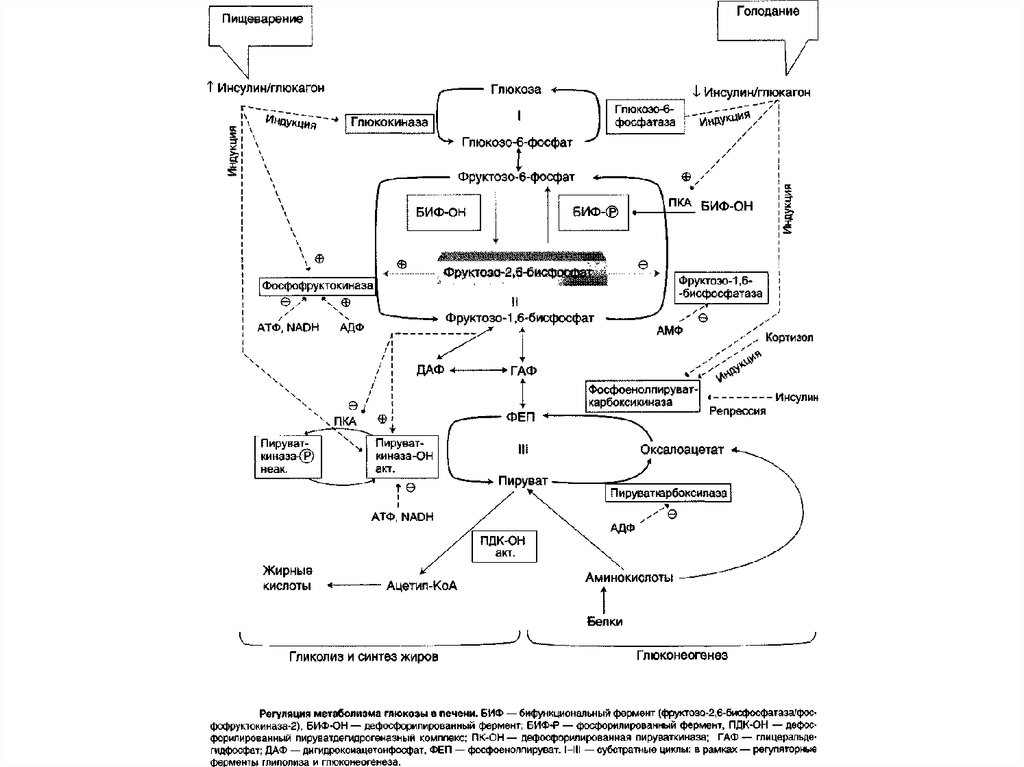
При окислении (концевых групп глюкозы -СНО и -СН2ОНобразуются 3 различных производных. При окислении
группы -СНО образуется глюконовая кислота. Если
окислению подвергается концевая группа -СН2ОН,
образуется глюкуроновая кислота. А если окисляются обе
концевые группы, то образуется сахарная кислота,
содержащая 2 карбоксильные группы. Восстановление
первого углерода приводит к образованию сахароспирта
— сорбитола.
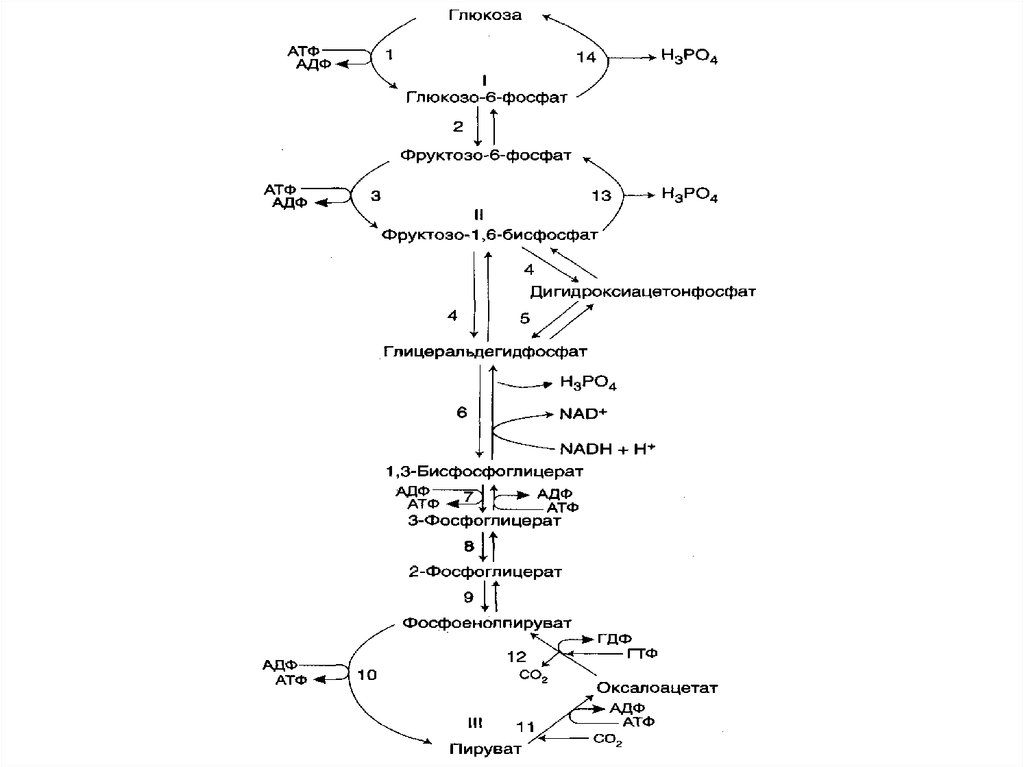
21. фруктоза
Фруктоза является кетогексозой (кетогруппа находится увторого углеродного атома). Фруктоза так же, как и
глюкоза, существует в циклической форме, образуя - и аномеры.
22. ОЛИГОСАХАРИДЫ
Олигосахариды содержат несколько (от двух до десяти)остатков моносахаридов, соединённых гликозидной
связью. Дисахариды — наиболее распространённые
олигомерные углеводы, встречающиеся в свободной
форме, т.е. не связанной с другими соединениями. По
химической природе дисахариды представляют собой
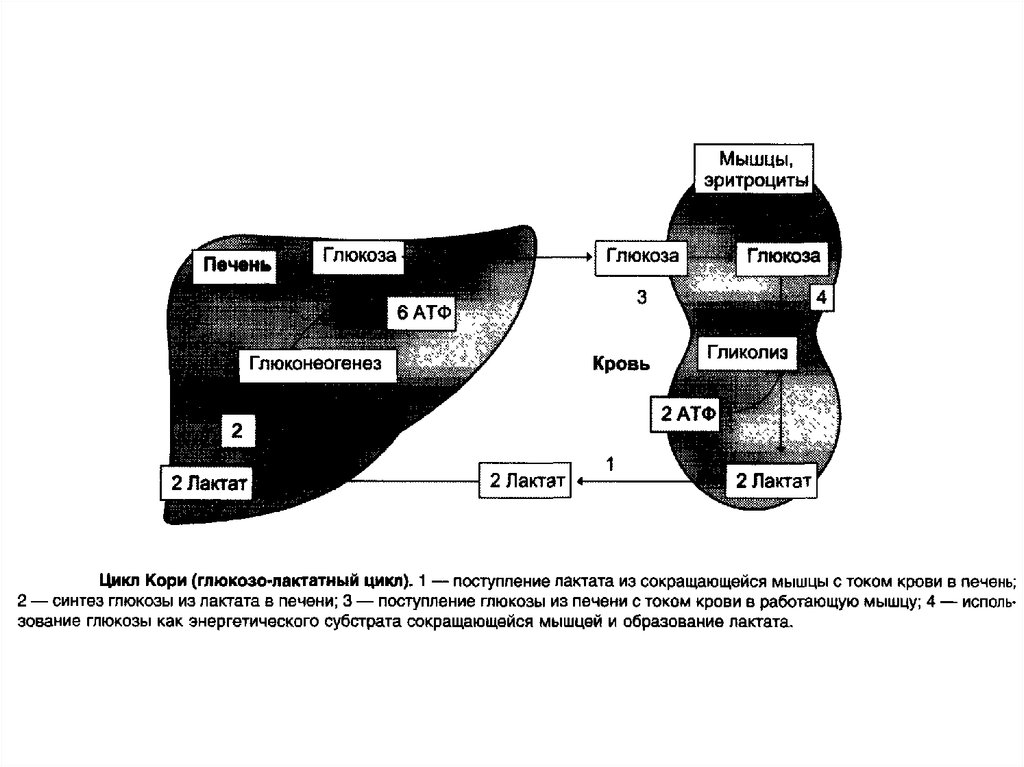
гликозиды,
которые
содержат
2
моносахарида,
соединённые гликозидной связью в - или конфигурации. В пище содержатся в основном такие
дисахариды, как сахароза, лактоза и мальтоза.
Среди трисахаридов наиболее известна рафиноза,
содержащая остатки фруктозы, глюкозы и галактозы.
Содержится в сахарной свекле и многих других растениях.
23. Дисахариды
24. Мальтоза
25. мальтоза
Мальтоза поступает с продуктами, содержащимичастично гидролизованный крахмал, например, солод,
пиво. Мальтоза также образуется при расщеплении
крахмала в кишечнике. Мальтоза состоит из двух остатков
D-глюкозы, соединённых -1,4-гликозидной связью.
26. Сахароза
27. сахароза
Сахароза — дисахарид, состоящий из -D-глюкозы и D-фруктозы, соединённых , -1,2-гликозидной связью.В сахарозе обе аномерные ОН-группы остатков глюкозы и
фруктозы участвуют в образовании гликозидной связи.
Следовательно,
сахароза
не
относится
к
восстанавливающим сахарам.
Сахароза — растворимый дисахарид со сладким вкусом.
Источником сахарозы служат растения, особенно сахарная
свёкла, сахарный тростник. Последнее объясняет
возникновение тривиального названия сахарозы —
«тростниковый сахар».
28. Лактоза
29. лактоза
Лактоза — молочный сахар; важнейшийдисахарид молока млекопитающих. В коровьем
молоке содержится до 5% лактозы, в женском
молоке — до 8%. В лактозе аномерная ОН-группа
первого углеродного атома остатка D-галактозы
связана -гликозидной связью с четвёртым
углеродным атомом D-глюкозы ( -1,4-связь).
Поскольку аномерный атом углерода остатка
глюкозы не участвует в образовании гликозидной
связи, следовательно, лактоза относится к
восстанавливающим сахарам.
30. Дисахариды пищи
31. Полисахариды
Структурные различия междуполисахаридами определяются:
строением моносахаридов, составляющих цепь;
типом гликозидных связей, соединяющих
мономеры в цепи;
последовательностью остатков моносахаридов в
цепи.
В зависимости от строения остатков моносахаридов
полисахариды можно разделить на
гомополисахариды (все мономеры идентичны) и
гетерополисахариды (мономеры различны). Оба типа
полисахаридов могут иметь как линейное
расположение мономеров, так и разветвлённое.
32. Гомо- и гетерополисахариды
33. Строение полисахаридов
34. резервные полисахариды
В зависимости от выполняемых ими функций полисахаридыможно разделить на 2 основные группы:
резервные
полисахариды,
выполняющие
энергетическую функцию. Эти полисахариды служат
источником глюкозы, используемым организмом по мере
необходимости.
Резервная функция этих углеводов обеспечивается их
полимерной природой. Полисахариды менее растворимы,
чем моносахариды, следовательно они не влияют на
осмотическое давление и поэтому могут накапливаться в
клетке, например, крахмал — в клетках растений,
гликоген — в клетках животных;
Структурные – выполняющие опорную функцию.
Целлюлоза растений.
35. крахмал
Крахмал — разветвлённый полисахарид (м.м. 105-107),состоящий из остатков глюкозы (гомогликан). Он
находится в клетках растений в виде гранул, практически
нерастворим в воде. В муке – 80%, в картофеле – 25%.
Крахмал состоит из амилозы (10-30%) и амилопектина
(70-90%). Амилоза — неразветвлённый полисахарид,
включающий 200—300 остатков глюкозы, связанных 1,4-гликозидной связью. Благодаря -конфигурации
глюкозного остатка, полисахаридная цепь имеет
конформацию спирали. Амилопектин – разветвленный
через каждые 20-30 единиц.
Декстрины – образуются при частичном расщеплении
полисахаридов – крахмала и гликогена.
36. Строение крахмала
37. Амилоза и амилопектин
38. Амилопектин
39. гликоген
Гликоген — полисахарид животных и человека. Так же,как крахмал в растениях, гликоген в клетках животных
выполняет резервную функцию, но, так как в пище
содержится лишь небольшое количество гликогена, он не
имеет пищевого значения. Много в печени и мышцах.
М.м. – 105 – 108 Д.
Гликоген представляет собой структурный аналог
крахмала, но имеет большую степень ветвления:
примерно на каждые 10 остатков глюкозы приходится
одна -1,6-гликозидная связь.
40. Гликоген
41. структурные полисахариды
структурные полисахариды, обеспечивающие клеткам иорганам механическую прочность;
полисахариды, входящие в состав межклеточного
матрикса, принимают участие в образовании тканей, а
также в пролиферации и дифференцировке клеток.
Полисахариды межклеточного матрикса водорастворимы
и сильно гидратированы.
42. Целлюлоза
43. целлюлоза
Целлюлоза (клетчатка) — основной структурныйполисахарид растений. Это самое распространённое
органическое соединение на земле. Доля целлюлозы в
клеточных стенках растений составляет 40—50%.
Целлюлоза имеет молекулярную массу порядка 106 Д,
длина молекулы может доходить до 6-8 мкм.
Целлюлоза — линейный полисахарид гомогликан,
построенный из остатков глюкозы, соединённых между
собой -1,4-гликозидными связями. Пищеварительная
система человека не имеет ферментов, гидролизующих связи в полисахаридах. Поэтому целлюлоза —
неиспользуемый углевод, но этот пищевой компонент
необходим для нормального протекания переваривания.
44. Образование гликопротеинов
45. Протеогликаны
Протеогликаны (мукополисахариды,гликозаминогликаны) – высокомолекулярные
углеводно-белковые соединения. Образуют
основную массу межклеточного матрикса
соединительной ткани. Составляют до 30% сухой
массы.
Линейные полимеры, построенные из повторяющихся
дисахаридных единиц. Всегда связаны с белками.
Состоят из остатков глюкозамина (либо галактозамина)
и остатков D-глюкуроновай (либо L-идуроновой)
кислот. Например, гиалуроновая кислота
(образование геля), хондроитинсульфат,
дерматансульфат, кератинсульфат, гепарин и др.
46. Протеогликан
47. Строение экстрацеллюлярного матрикса
48. Переваривание углеводов
49. Переваривание углеводов в ротовой полости
Вротовой
полости
пища
измельчается
при
пережёвывании, смачиваясь при этом слюной. Слюна на
99% состоит из воды и обычно имеет рН 6,8. В слюне
присутствует гидролитический фермент -амилаза
( -1,4-гликозидаза), расщепляющая в крахмале
1,4-гликозидные связи. В ротовой полости не может
происходить полное расщепление крахмала, так как
действие фермента на крахмал кратковременно.
50. Переваривание углеводов в кишечнике
Последующиеэтапы
переваривания
нерасщеплённого
или
частично
расщеплённого крахмала, а также других
углеводов пищи происходит в тонком
кишечнике в разных его отделах под
действием гидролитических ферментов —
гликозидаз.
51. Панкреатическая -амилаза
Панкреатическая -амилазаВ двенадцатиперстной кишке рН среды
желудочного содержимого нейтрализуется, так
как секрет поджелудочной железы имеет рН 7,5—
8,0 и содержит бикарбонаты (НСО3~). С секретом
поджелудочной железы в кишечник поступает
панкреатическая -амилаза. Этот фермент
гидролизует -1,4-гликозидные связи в крахмале
и декстринах.
Продукты переваривания крахмала на этом этапе
— дисахарид мальтоза, содержащая 2 остатка
глюкозы, связанные -1,4-связью.
52. Гидролиз крахмала панкреатической -амилазой
Гидролиз крахмала панкреатической-амилазой
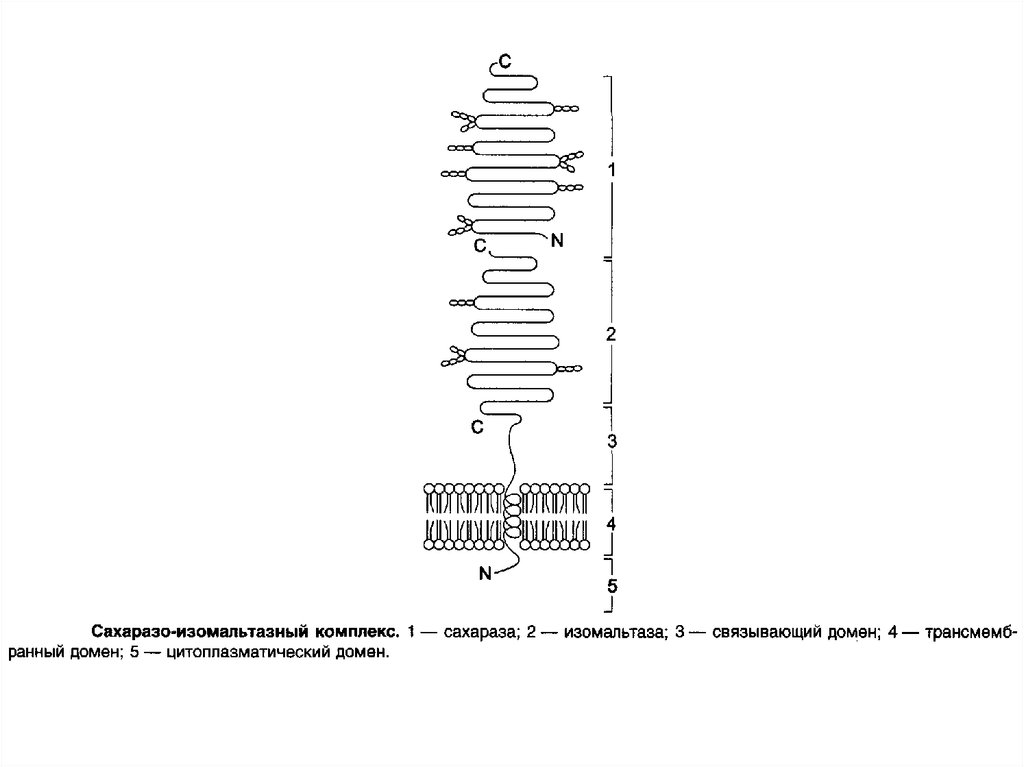
53. Сахаразо-изомальтазный комплекс
Этот ферментативный комплекс состоит из двухполипептидных цепей и имеет доменное строение.
Сахаразо-изомальтазный комплекс прикрепляется к
мембране микроворсинок кишечника с помощью
гидрофобного (трансмембранного) домена, образованного
N-концевой частью полипептида. Каталитический центр
выступает в просвет кишечника. Связь этого
пищеварительного фермента с мембраной способствует
эффективному поглощению продуктов гидролиза клеткой.
Сахаразо-изомальтазный комплекс гидролизует сахарозу и
изомальтозу, расщепляя -1,2- и -1,6-гликозидные связи.
54.
55. -Гликозидазный комплекс (лактаза)
-Гликозидазный комплекс(лактаза)
Лактаза расщепляет -1,4-гликозидные связи
между галактозой и глюкозой в лактозе.
Этот ферментативный комплекс по химической
природе является гликопротеином. Лактоза, как и
другие гликозидазные комплексы, связана с
щёточной каемкой и распределена неравномерно
по всему тонкому кишечнику. Активность
лактазы колеблется в зависимости от возраста.
56. Переваривание углеводов
+НОНА) крахмал
декстрины
амилаза
гликоген
(слюни, панкреатическая,
+НОН
амилаза
мальтоза
кишечная)
Б) мальтоза
+НОН
глюкоза
мальтаза
(слюни, панкреатическая,
кишечная)
В) сахароза
Г) лактоза
+НОН
сахараза
(кишечная)
+НОН
лактаза
(кишечная)
кровь
глюкоза
фруктоза
кровь
глюкоза
галактоза
кровь
57.
Переваривание углеводов у жвачныхСложные
углеводы:
+НОН
бактериальные
ферменты
Глюкоза бактериальные
ферменты
брожения
энергия
для + летучие карбоновые кислоты
бактерий
кровь
58.
За сутки образуется 3 – 5 кг летучих карбоновыхкислот, среди которых:
Уксусная – 70 - 75%
Пропионовая – 15 - 20%
Масляная – 5 -10%
Молочная – 3 - 5%
Прочие – 0,1- 1%
59. Всасывание моносахаридов в кишечнике
Транспорт моносахаридов в клетки слизистойоболочки кишечника может осуществляться
разными
способами:
путём
облегчённой
диффузии и активного транспорта. В случае
активного транспорта глюкоза и Na+ проходят
через мембраны с люминальной стороны,
связываясь с разными участками белкапереносчика. При этом Na+ поступает в клетку по
градиенту концентрации, и одновременно
глюкоза транспортируется против градиента
концентрации.
60.
61.
62. Содержание моносахаридов в крови, мг%
Животные% глюкозы
% фруктозы
% пентозы
Лошадь
74
11
15
Корова
54
16
30
Овца
83
5
12
Свинья
57
16
27
Курица
85
4
11
Собака
79
7
14
Человек
68
15
17
Рыбы
86
4
10
63.
Гипергликемия – повышенный уровень глюкозы вкрови
а) Алиментарная
б) Патологическая
Гипогликемия – пониженный уровень глюкозы в крови
Глюкозурия – наличие глюкозы в моче
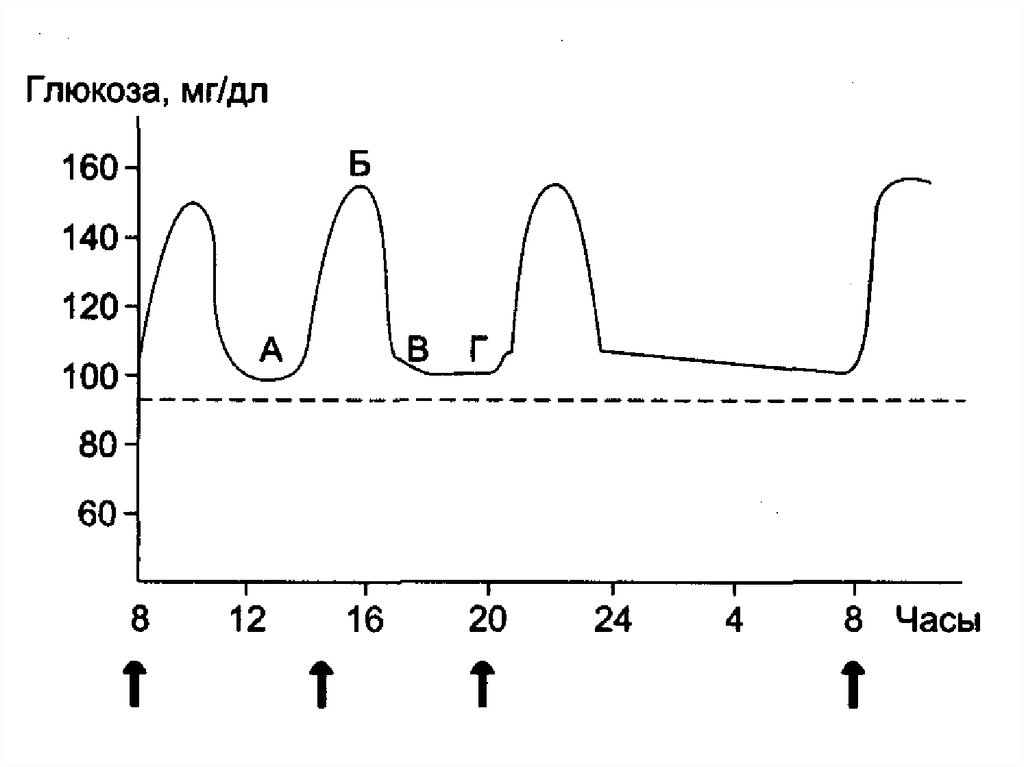
64. Гормоны, регуляторы уровня глюкозы
а) гипогликемического действия: инсулинб) гипергликемического действия:
Основные: адреналин – во всех тканях,
глюкагон – только в печени.
А также: тироксин и
гликокортикоиды.
через фосфорилазу
65. Регуляция уровня сахара в крови
Центр углеводного обмена гипоталамусагипофиз
АКТГ,ТТГ
железы внутренней секреции
ГОРМОНЫ
инсулин
адреналин
глюкагон
тироксин
гликокортикоид
66.
ГликогенезИНСУЛИН
синтез гликогена из глюкозы
ГЛИКОКОРТИКОИДЫ
Гликонеогенез
синтез гликогена через
глюкозу из других веществ
(молочная и пропионовая
кислоты, безазотистые остатки
аминокислот)
67. Адреналин
Адреналин стимулирует выведение глюкозы изпечени в кровь, для того чтобы снабдить ткани (в
основном мозг и мышцы) «топливом» в
экстремальной ситуации. Эффект адреналина в
печени
обусловлен
фосфорилированием
(и активацией) гликогенфосфорилазы. Адреналин
имеет сходный с глюкагоном механизм действия.
Но возможно включение и другой эффекторной
системы передачи сигнала в клетку печени
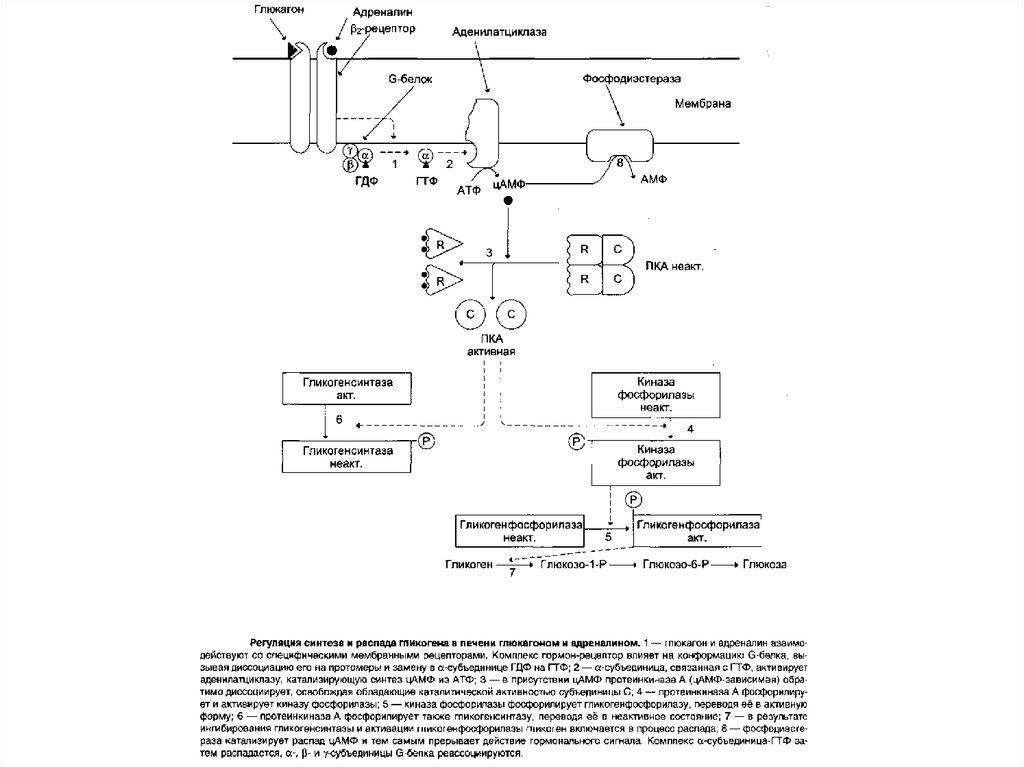
68. Глюкагон
Глюкагон для гепатоцитов служит внешнимсигналом о необходимости выделения в кровь
глюкозы
за
счёт
распада
гликогена
(гликогенолиза) или синтеза глюкозы из других
веществ — глюконеогенеза. Гормон связывается с
рецептором на плазматической мембране и
активирует
при
посредничестве
G-белка
аденилатциклазу,
которая
катализирует
образование цАМФ из АТФ. Далее следует каскад
реакций, приводящий в печени к активации
гликоген-фосфорилазы
и
ингибированию
гликогенсинтазы.
69.
70.
71. Транспорт глюкозы из крови в клетки
Потребление глюкозы клетками из кровотока происходиттакже путём облегчённой диффузии. Следовательно,
скорость трансмембранного потока глюкозы зависит
только от градиента её концентрации. Исключение
составляют клетки мышц и жировой ткани, где
облегчённая диффузия регулируется инсулином (гормон
поджелудочной железы). В отсутствие инсулина
плазматическая мембрана этих клеток непроницаема для
глюкозы, так как она не содержит белки-переносчики
(транспортёры) глюкозы.
72. Переносчики глюкозы
Все типы ГЛЮТ могут находиться как вплазматической мембране, так и в цитозольных
везикулах. ГЛЮТ-4 (и в меньшей мере ГЛЮТ-1)
почти полностью находятся в цитоплазме клеток.
Влияние инсулина на такие клетки приводит к
перемещению везикул, содержащих ГЛЮТ, к
плазматической мембране, слиянию с ней и
встраиванию транспортёров в мембрану. После
чего возможен облегчённый транспорт глюкозы в
эти клетки. После снижения концентрации
инсулина в крови транспортёры глюкозы снова
перемещаются в цитоплазму, и поступление
глюкозы в клетку прекращается
73.
74.
75.
76. МЕТАБОЛИЗМ ГЛЮКОЗЫ В КЛЕТКЕ
После всасывания в кишечнике моносахаридыпоступают
в
воротную
вену
и
далее
преимущественно в печень. Поскольку в составе
основных углеводов пищи преобладает глюкоза,
её можно считать основным продуктом
переваривания углеводов. Другие моносахариды,
поступающие из кишечника в процессе
метаболизма, могут превращаться в глюкозу или
продукты её метаболизма.
77. ФОСФОРИЛИРОВАНИЕ ГЛЮКОЗЫ
В дальнейших превращениях в клетках глюкоза идругие моносахариды участвуют только в виде
фосфорных
эфиров.
Фосфорилирование
свободных моносахаридов — обязательная
реакция на пути их использования, она приводит
к образованию более реакционно-способных
соединений и поэтому может рассматриваться как
реакция активации.
78. глюкозо-6-фосфат
Образование глюкозо-6-фосфата в клетке— своеобразная «ловушка» для глюкозы,
так как мембрана клетки непроницаема для
фосфорилированной
глюкозы
(нет
соответствующих транспортных белков).
Кроме того, фосфорилирование уменьшает
концентрацию свободной глюкозы в
цитоплазме. В результате создаются
благоприятные условия для облегчённой
диффузии глюкозы в клетки из крови.
79. Роль глюкозо-6-фосфата
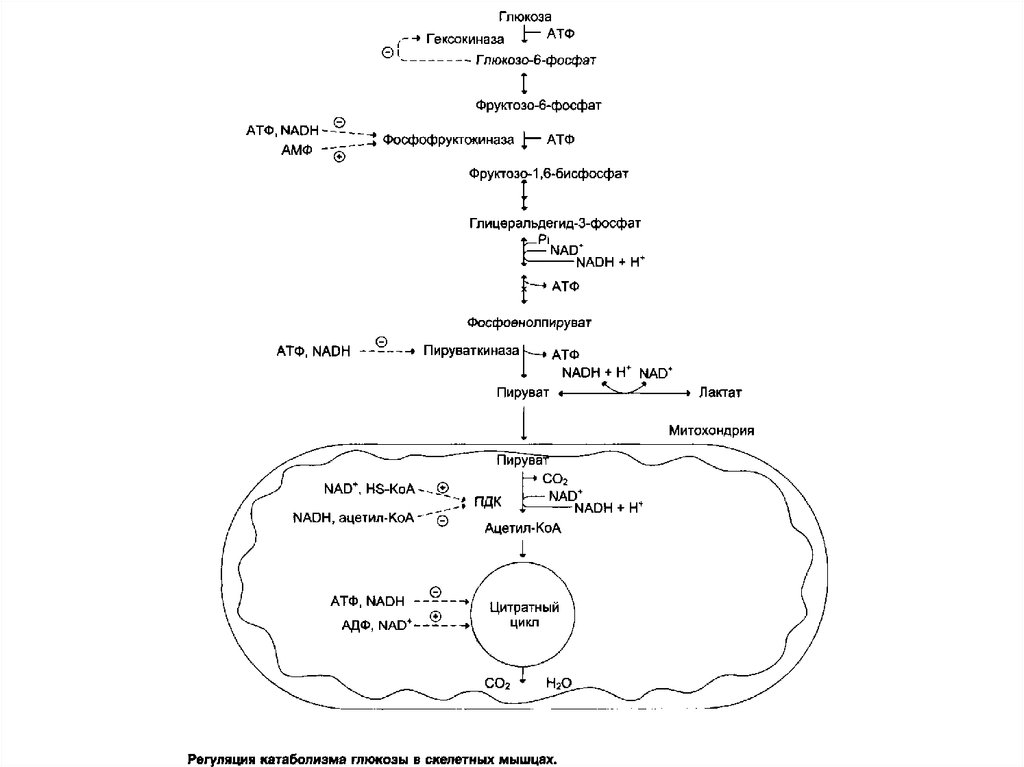
80. Глюкокиназа
Глюкокиназа. Фосфорилирование глюкозы вгепатоцитах
в
период
пищеварения
обеспечивается свойствами глюкокиназы, которая
имеет не высокое сродство к глюкозе. В этот
период концентрация глюкозы в воротной вене
больше, чем в других отделах кровяного русла и
может превышать 10 ммоль/л, а следовательно,
активность
глюкокиназы
в
гепатоцитах
повышается.
81. Гексокиназа
Гексокиназаотличается
от
глюкокиназы
высоким сродством к глюкозе (Кm <0,1 ммоль/л).
Следовательно, этот фермент, в отличие от
глюкокиназы, активен при низкой концентрации
глюкозы в крови, что характерно для
постабсорбтивного состояния. Печень в этот
период поглощает гораздо меньше глюкозы, так
как
скорость
её
внутриклеточного
фосфорилирования
глюкокиназой
резко
снижается. Тогда как потребление глюкозы
мозгом, эритроцитами и другими тканями
обеспечивается активной в этих условиях
гексокиназой.
82. Дефосфорилирование глюкозо-6-фосфата
Превращение глюкозо-6-фосфата в глюкозу возможно впечени, почках и клетках эпителия кишечника. В клетках
этих органов имеется фермент глюкозо-6-фосфатаза,
катализирующая
отщепление
фосфатной
группы
гидролитическим путём: Глюкозо-6-фосфат +Н2О -»
Глюкоза + Н3РО4
Образовавшаяся
свободная
глюкоза
способна
диффундировать из этих органов в кровь. В других
органах и тканях глюкозо-6-фосфатазы нет, и поэтому
дефосфорилирование глюкозо-6-фосфата невозможно.
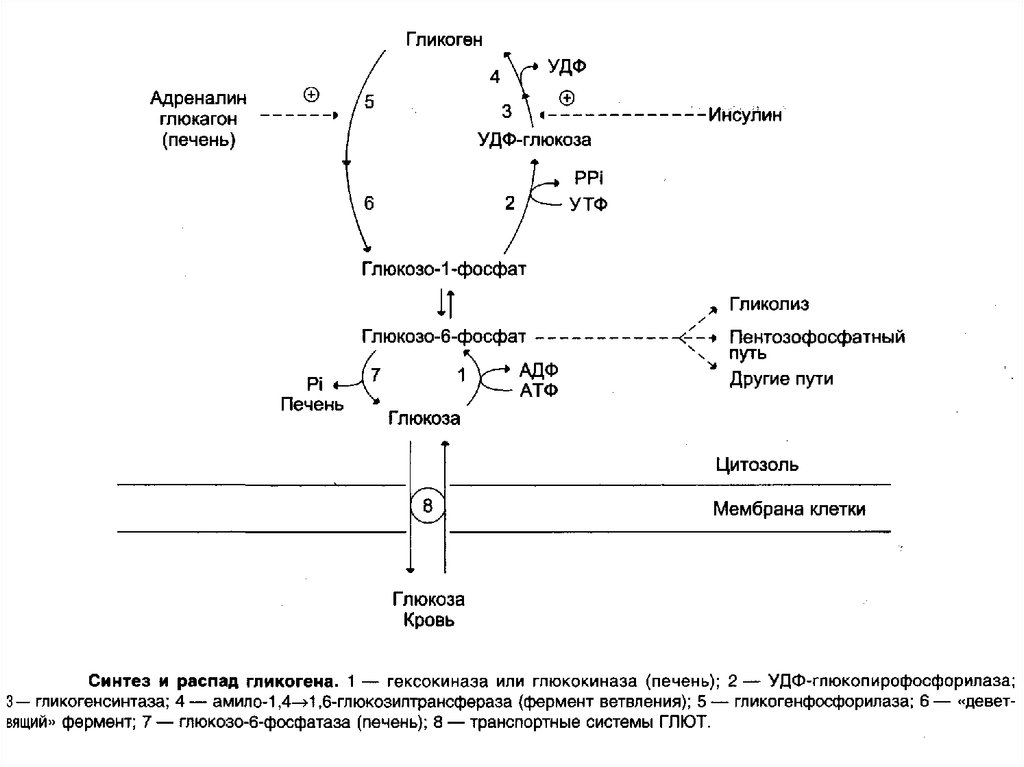
83. Строение и функции гликогена.Метаболизм.
Гликоген — разветвлённый гомополимер глюкозы, вкотором остатки глюкозы соединены в линейных участках
-1,4-гликозидной связью. В точках ветвления мономеры
соединены -1,6-гликозидными связями. Эти связи
образуются примерно с каждым десятым остатком
глюкозы. Следовательно, точки ветвления в гликогене
встречаются примерно через каждые десять остатков
глюкозы. Так возникает древообразная структура с
молекулярной массой >107Д, что соответствует
приблизительно 50 000 остатков глюкозы
84.
85. Функции гликогена в печени и мышцах
86. СИНТЕЗ ГЛИКОГЕНА (ГЛИКОГЕНОГЕНЕЗ)
Гликогенсинтезируется
в
период
пищеварения (через 1—2 ч после приёма
углеводной пищи). Следует отметить, что
синтез гликогена из глюкозы, как и любой
анаболический
процесс,
является
эндергоническим, т.е. требующим затрат
энергии.
87.
88.
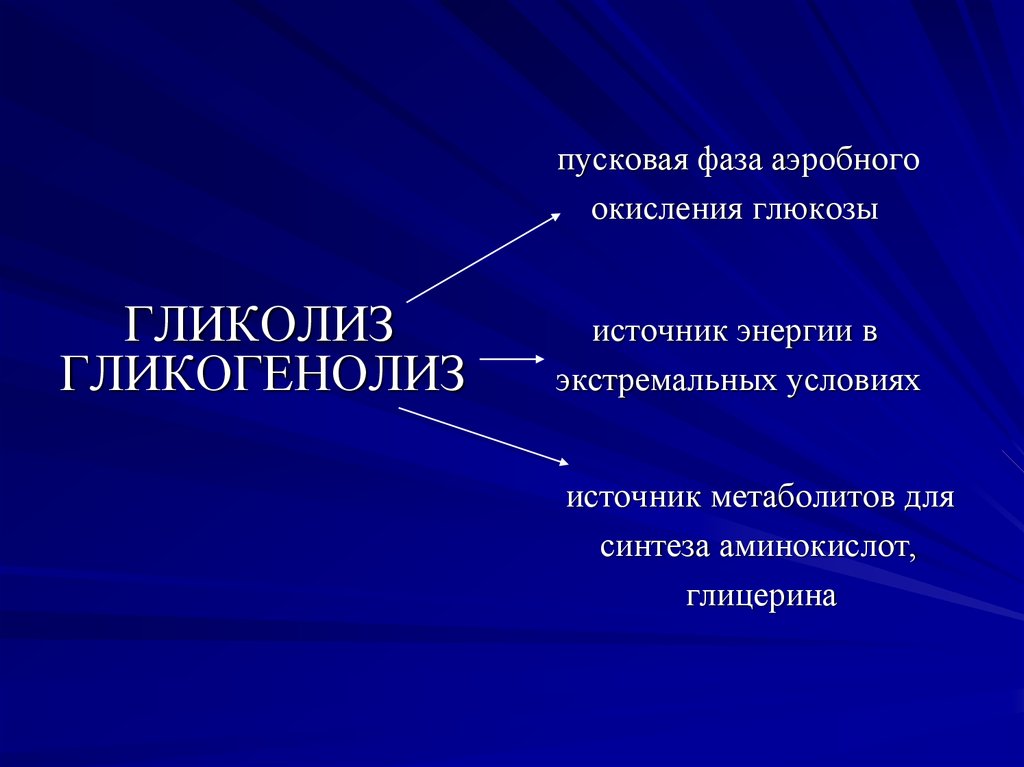
пусковая фаза аэробногоокисления глюкозы
ГЛИКОЛИЗ
ГЛИКОГЕНОЛИЗ
источник энергии в
экстремальных условиях
источник метаболитов для
синтеза аминокислот,
глицерина
89. Распад гликогена (гликогенолиз)
Распад гликогена или его мобилизация происходят в ответна повышение потребности организма в глюкозе.
Гликоген печени распадается в основном в интервалах
между приёмами пищи, кроме того, этот процесс в печени
и мышцах ускоряется во время физической работы.
Распад гликогена происходит путём последовательного
отщепления остатков глюкозы в виде глюкозо-1-фосфата.
Гликозидная связь расщепляется с использованием
неорганического фосфата, поэтому процесс называется
фосфоролизом, а фермент гликоген-фосфорилазой.
90.
ГЛИКОГЕНфосфорилаза А
ГЛЮКОЗО – 1 – ФОСФАТ
фосфоглюкомутаза
ГЛЮКОЗО – 6 – ФОСФАТ
глюкозо – 6 – фосфатаза
ГЛЮКОЗА
мембрана
(кровь)
91.
92.
93. РЕГУЛЯЦИЯ МЕТАБОЛИЗМА ГЛИКОГЕНА В ПЕЧЕНИ
Как уже отмечалось, первичный сигнал длясинтеза инсулина и глюкагона — изменение
концентрации глюкозы в крови. Инсулин и
глюкагон постоянно присутствуют в крови, но
при смене абсорбтивного периода на постабсорбтивный изменяется их относительная
концентрация, что является главным фактором,
переключающим метаболизм гликогена в печени.
Отношение концентрации инсулина в крови к
концентрации глюкагона называют «инсулинглюкагоновый индекс».
94. Метаболическая роль углеводов
1. Источник энергии ≈ 16 кДж/г.2. Пластический материал.
3. «В пламени» углеводов окисляются жиры и белки.
4. Исходные продукты для построения аминокислот.
5. Исходные продукты для образования витамина С.
6. Участвуют в детоксикации ядов экзо-и эндогенного
происхождения.
95. Основные пути использования углеводов
1. Окисление.а) анаэробное - гликолиз и гликогенолиз;
б) аэробное - через ЦТК, пентозный путь,
глюкуронидный путь
2. Синтез гликогена.
3. Синтез других веществ
96. Анаэробное окисление
а) ГликолизГлюкоза ---> энергия (2 АТФ) + молочная кислота
б) Гликогенолиз
Гликоген ---> Глюкоза ---> энергия (З АТФ) +
молочная кислота
Аэробное окисление:
Глюкоза -------- энергия + пировиноградная кислота
97.
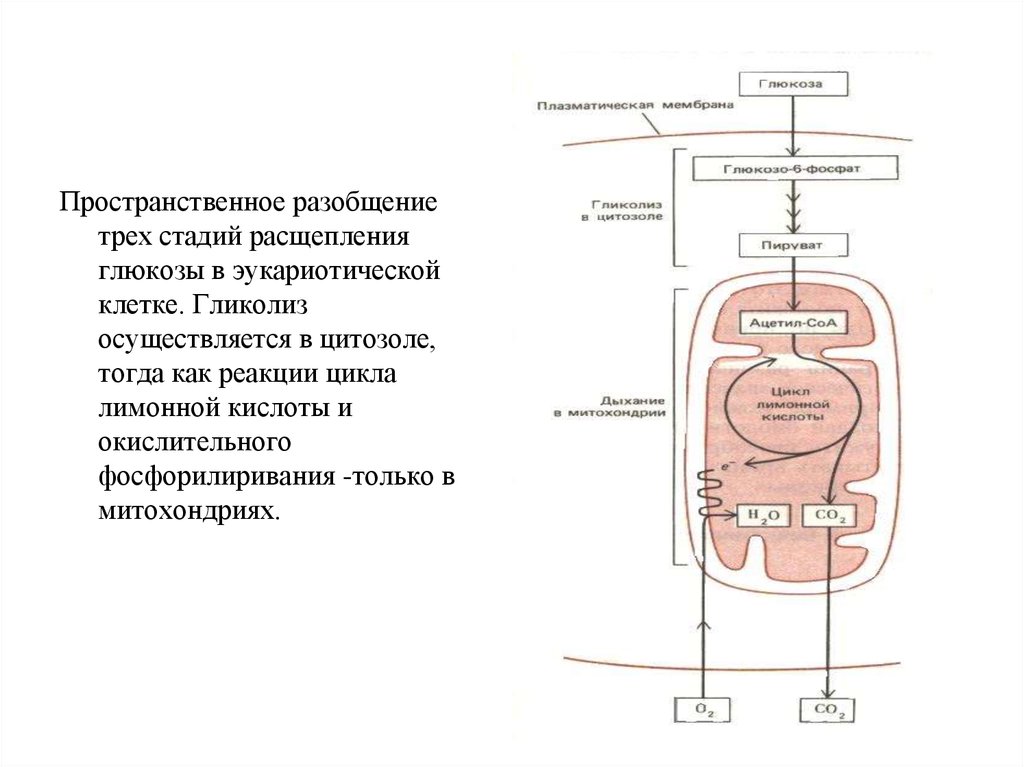
Пространственное разобщениетрех стадий расщепления
глюкозы в эукариотической
клетке. Гликолиз
осуществляется в цитозоле,
тогда как реакции цикла
лимонной кислоты и
окислительного
фосфорилиривания -только в
митохондриях.
98.
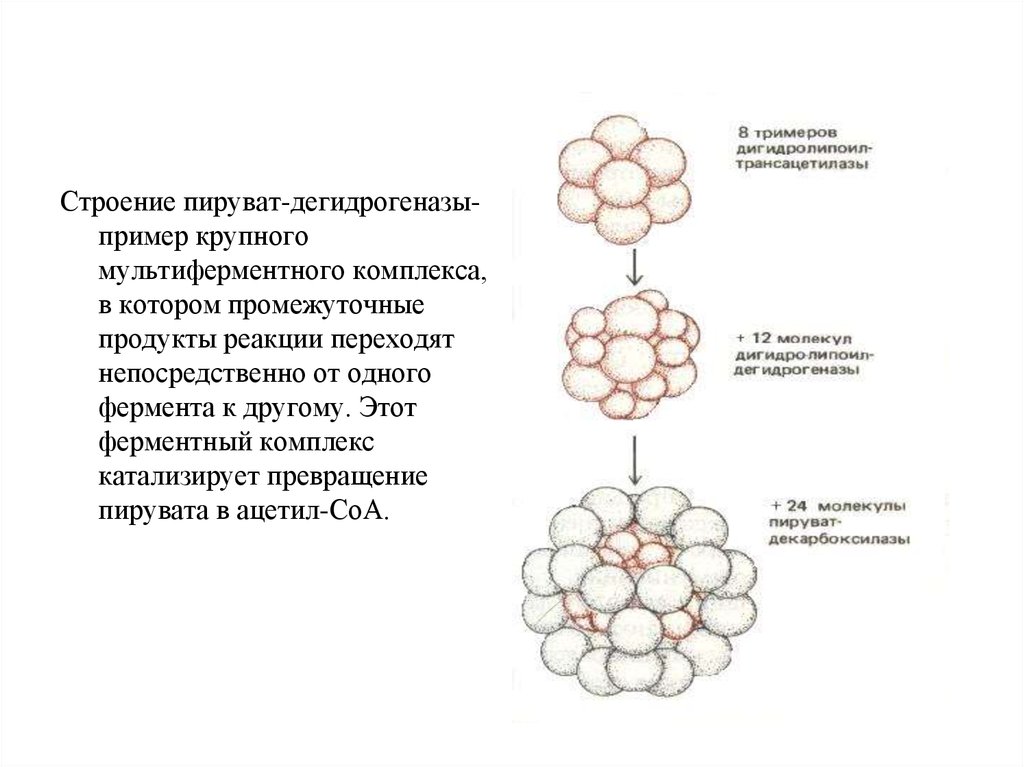
Строение пируват-дегидрогеназыпример крупногомультиферментного комплекса,
в котором промежуточные
продукты реакции переходят
непосредственно от одного
фермента к другому. Этот
ферментный комплекс
катализирует превращение
пирувата в ацетил-СоА.
99. Катаболизм глюкозы
Окисление глюкозы до СО2 и Н2О (аэробный распад).Аэробный распад глюкозы можно выразить суммарным
уравнением:
С6Н12О6 + 6О2 → 6 СО2 + 6Н2О + 2820 кДж/моль.
Этот процесс включает несколько стадий.
1. Процесс окисления глюкозы с образованием двух молекул
пирувата;
2.Общий путь катаболизма, включающий превращение
пирувата в ацетил-КоА, его дальнейшее окисление
(дегидрирование) в цитратном цикле;
3.Перенос протонов и электронов по дыхательной цепи
митохондрий на кислород, образование воды и
сопряженный синтез АТФ.
100. Этапы аэробного гликолиза
В аэробном гликолизе можно выделить 2этапа.
1.Подготовительный этап, в ходе которого
глюкоза фосфорилируется и расщепляется
на две молекулы фосфотриоз. Эта серия
реакций протекает с использованием 2
молекул АТФ.
2.Этап, сопряжённый с синтезом АТФ. В
результате
этой
серии
реакций
фосфотриозы
превращаются
в
пируват.
Энергия,
высвобождающаяся
на
этом
этапе,
используется для синтеза 10 моль АТФ.
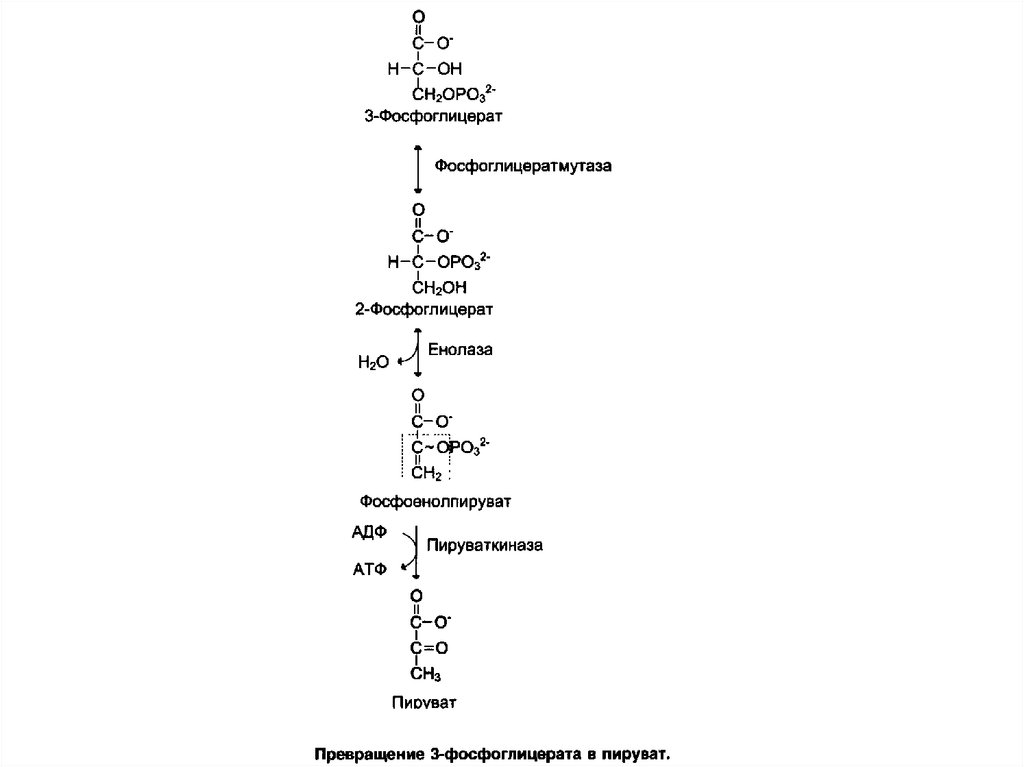
101. Превращение глюкозо-6-фосфата в триозофосфаты
102.
103.
104. Анаэробный гликолиз
Анаэробным гликолизом называют процесс расщепленияглюкозы с образованием в качестве конечного продукта
лактата. Этот процесс протекает без использования
кислорода и поэтому не зависит от работы
митохондриальной дыхательной цепи. АТФ образуется за
счёт
реакций
субстратного
фосфорилирования.
Суммарное уравнение процесса:
С6Н12О6 + 2 Н3Р04 + 2 АДФ = 2 С3Н6О3 + 2 АТФ + 2 Н2О.
105.
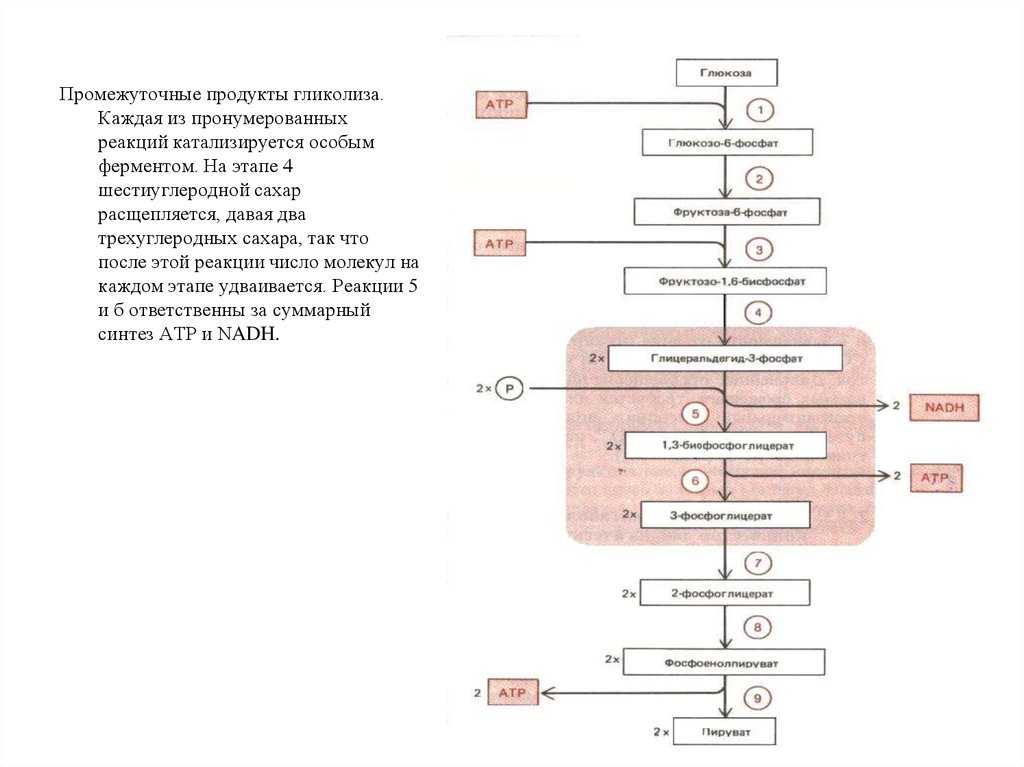
Промежуточные продукты гликолиза.Каждая из пронумерованных
реакций катализируется особым
ферментом. На этапе 4
шестиуглеродной сахар
расщепляется, давая два
трехуглеродных сахара, так что
после этой реакции число молекул на
каждом этапе удваивается. Реакции 5
и б ответственны за суммарный
синтез АТР и NADH.
106.
107.
108.
109.
110.
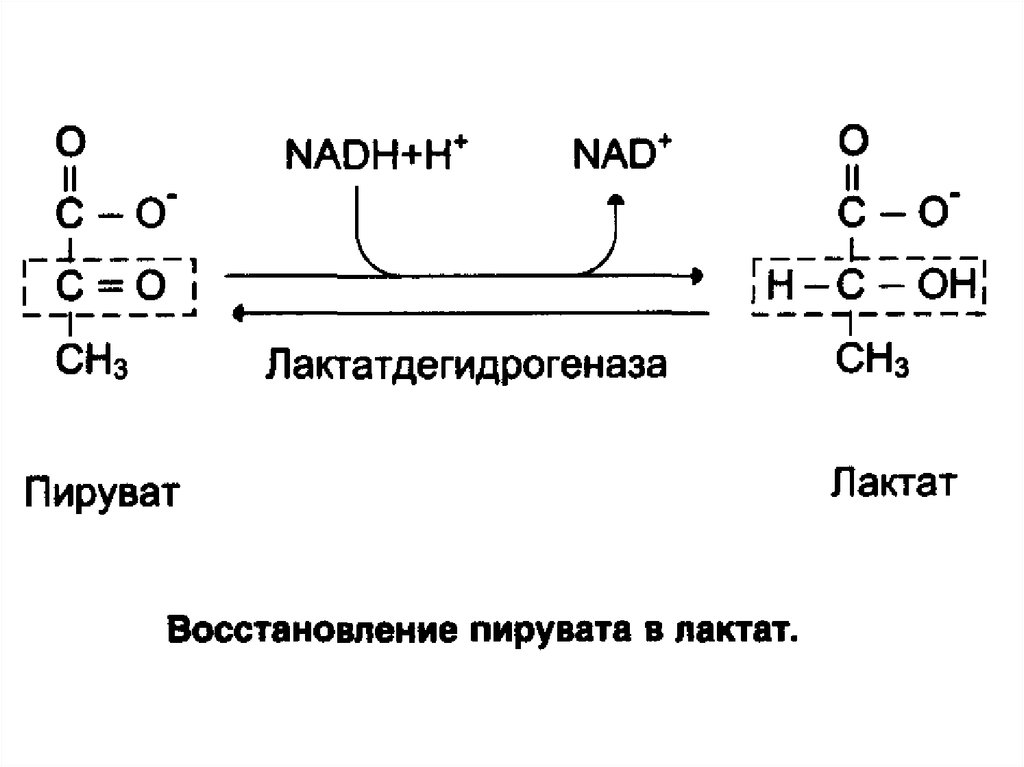
Лактат, образовавшийся в интенсивно работающихмышцах или в клетках с преобладающим анаэробным
способом катаболизма глюкозы, поступает в кровь, а
затем в печень. В печени отношение NADH/NAD+
ниже, чем в сокращающейся мышце, поэтому лактатдегидрогеназная реакция протекает в обратном
направлении, т.е. в сторону образования пирувата из
лактата. Далее пируват включается в глюконеогенез, а
образовавшаяся глюкоза поступает в кровь и
поглощается скелетными мышцами.
111.
Цикл Кори выполняет 2 важнейшие функции:1 — обеспечивает утилизацию лактата;
2
— предотвращает накопление лактата и, как
следствие этого, опасное снижение рН (лактоацидоз).
Часть пирувата, образованного из лактата, окисляется
печенью до СО2 и Н2О. Энергия окисления может
использоваться для синтеза АТФ, необходимого для
реакций глюконеогенеза.
112.
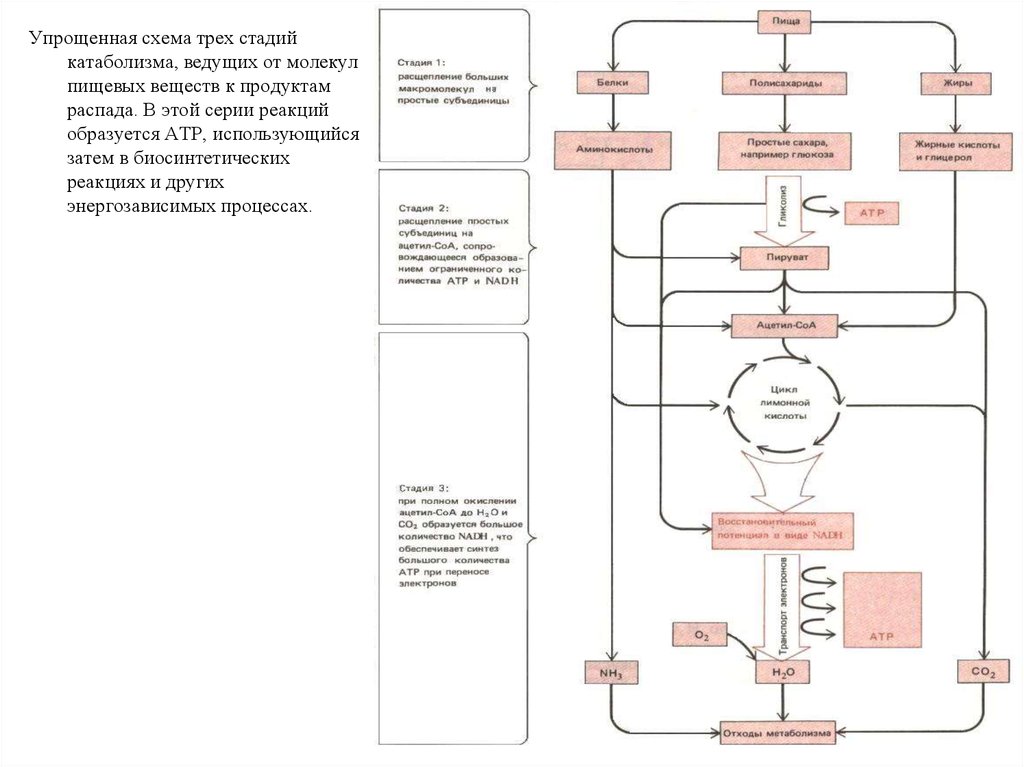
Упрощенная схема трех стадийкатаболизма, ведущих от молекул
пищевых веществ к продуктам
распада. В этой серии реакций
образуется АТР, использующийся
затем в биосинтетических
реакциях и других
энергозависимых процессах.
113. Центральным процессом катаболизма является цикл лимонной кислоты
Главная функция цикла лимонной кислоты окисление ацетогруппы, включающейся в этотцикл в форме молекул ацетил-СоА. Процесс этот
носит циклический характер, поскольку
ацетогруппа окисляется не сразу, а лишь после
того, как она ковалентно присоединится к более
крупной молекуле - оксалоацетату, которая
регенерируется после каждого оборота цикла.
114.
ЦТК – последовательные превращения ди- итрикарбоновых кислот, в ходе которых
вовлекаемая в этот цикл уксусная кислота
окисляется до СО2 и Н2О с выделением 12
АТФ/М окисленной кислоты.
ЦТК – универсальный заключительный
этап окисления углеводов, жирных
кислот, глицерина, аминокислот.
115.
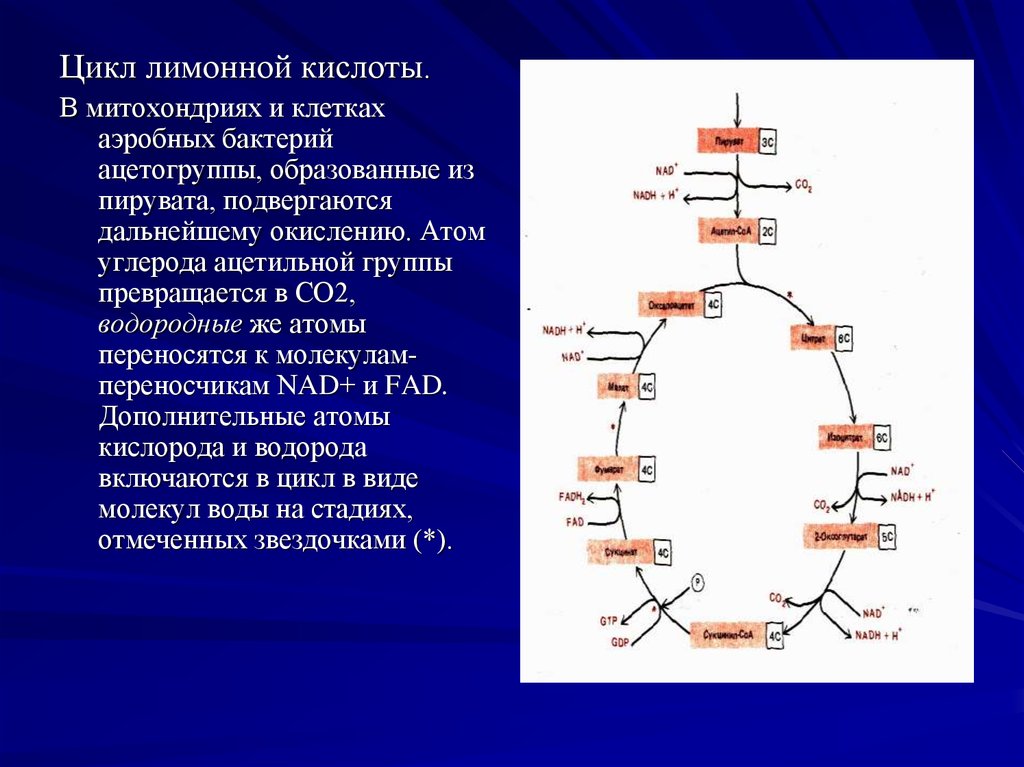
Цикл лимонной кислоты.В митохондриях и клетках
аэробных бактерий
ацетогруппы, образованные из
пирувата, подвергаются
дальнейшему окислению. Атом
углерода ацетильной группы
превращается в СО2,
водородные же атомы
переносятся к молекулампереносчикам NAD+ и FAD.
Дополнительные атомы
кислорода и водорода
включаются в цикл в виде
молекул воды на стадиях,
отмеченных звездочками (*).
116. Цикл лимонной кислоты (трикарбоновых кислот, Кребса)
117. Общая схема цитратного цикла
118.
Энергетическая эффективность окисления1М глюкозы
38 АТФ
в том числе:
А) гликолиз -- 2 МК + 2 АТФ
Б) 2 МК О 2 ПВК + 6 АТФ
О
В)2 ПВК
2СН3СО.SКоА + 6 АТФ
Г) 2СН3СО.SКоА ЦТК Н2О + СО2 + 24 АТФ
38 АТФ
2
2
119. ПЕНТОЗОФОСФАТНЫЙ ПУТЬ ПРЕВРАЩЕНИЯ ГЛЮКОЗЫ
Пентозофосфатныйпуть,
называемый
также
гексомонофосфатным шунтом, служит альтернативным
путём окисления глюкозо-6-фосфата. Пентозофосфатный
путь состоит из 2 фаз (частей) — окислительной и
неокислительной.
В окислительной фазе глюкозо-6-фосфат необратимо
окисляется в пентозу — рибулозо-5-фосфат, и образуется
восстановленный NADPH.
В неокислительной фазе рибулозо-5-фосфат обратимо
превращается в рибозо-5-фосфат и метаболиты гликолиза.
120.
Пентозофосфатный путь обеспечивает клетки рибозой длясинтеза пуриновых и пиримидиновых нуклеотидов и
гидрированным
коферментом
NADPH,
который
используется в восстановительных процессах.
Суммарное
уравнение
пентозофосфатного
пути
выражается следующим образом:
3 Глюкозо-6-фосфат + 6 NADP+ → 3 СО2 +6 (NADPH +
Н+) + 2 Фруктозо-6-фосфат + Глицеральдегид-3-фосфат.
Ферменты пентозофосфатного пути, так же, как и
ферменты гликолиза, локализованы в цитозоле.
121. Окислительный этап
В окислительной части пентозофосфатного пути глюкозо6-фосфатподвергается
окислительному
декарбоксилированию, в результате которого образуются
пентозы. Этот этап включает 2 реакции дегидрирования.
122. Окислительный этап пентозофосфатного пути
123.
Суммарноеуравнение
окислительного
этапа
пентозофосфатного пути можно представить в виде:
Реакции окислительного этапа служат основным
источником NADPH в клетках. Гидрированные
коферменты снабжают водородом биосинтетические
процессы, окислительно-восстановительные реакции,
включающие защиту клеток от активных форм кислорода.
NADPH как донор водорода участвует в анаболических
процессах, например в синтезе холестерина. Это источник
восстановительных эквивалентов для цитохрома Р450,
катализирующего образование гидроксильных групп при
синтезе стероидных гормонов, жёлчных кислот, при
катаболизме
лекарственных
веществ
и
других
чужеродных соединений
124. Неокислительный этап
Неокислительный этап пентозофосфатного пути включаетсерию обратимых реакций, в результате которых
рибулозо-5-фосфат превращается в рибозо-5-фосфат и
ксилулозо-5-фосфат, и далее за счёт переноса углеродных
фрагментов в метаболиты гликолиза — фруктозо-6фосфат и глицеральдегид-3-фосфат. В этих превращениях
принимают участие ферменты: эпимераза, изомераза,
транскетолаза и трансальдолаза.
125.
Суммарный результат метаболизма 3 молекул рибулозо-5фосфата в неокислительной фазе пентозофосфатного пути— образование 2 молекул фруктозо-6-фосфата и 1
молекулы глицеральдегид-3-фосфата. Далее фруктозо-6фосфат и глицеральдегид-3-фосфат могут превратиться в
глюкозу. С учётом стехиометрического коэффициента,
равного 2, для образования 5 молекул глюкозы
(содержащих 30 атомов углерода) потребуются 4
молекулы
фруктозо-6-фосфата
и
2
молекулы
глицеральдегид-3-фосфата (в сумме содержащие также 30
атомов углерода) или, соответственно, 6 молекул
рибулозо-5-фосфата. Таким образом, неокислительный
путь можно представить как процесс возвращения пентоз
в фонд гексоз.
126. Пентозофосфатный цикл
Окислительныйэтап
образования
пентоз
и
неокислительный этап (путь возвращения пентоз в
гексозы) составляют вместе циклический процесс.
Такой процесс можно описать общим уравнением:
6 Глюкозо-6-фосфат + 12 NADP+ + 2 Н2О → 6 Рибулозо-5фосфат + 12 NADPH +12 Н+ + 6СО2.
Это означает, что из 6 молекул глюкозы образуются 6
молекул рибулозо-5-фосфат (пентозы) и 6 молекул СО2.
127. Превращения рибулозо-5-фосфата.
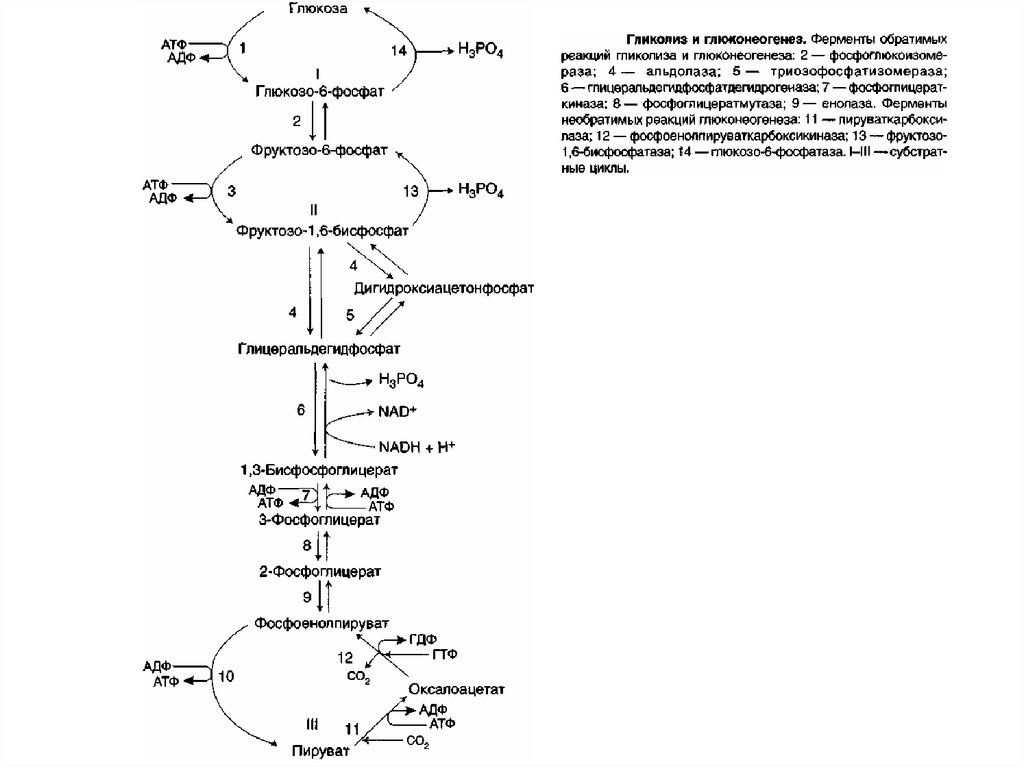
128. Реакции глюконеогенеза
Большинство реакций глюконеогенеза протекает засчёт обратимых реакций гликолиза и катализируется
теми же ферментами. Однако 3 реакции гликолиза
термодинамически необратимы. На этих стадиях
реакции глюконеогенеза протекают другими путями.
Необходимо отметить, что гликолиз протекает в
цитозоле, а часть реакций глюконеогенеза происходит
в митохондриях.
129. Образование фосфоенолпирувата из пирувата — первая из необратимых стадий глюконеогенеза
Образованиефосфоенолпирувата
из
пирувата
происходит в ходе двух реакций, первая из которых
протекает в митохондриях. Пируват, образующийся из
лактата
или
из
некоторых
аминокислот,
транспортируется в матрикс митохондрий и там
карбоксилируется с образованием оксалоацетата.
Пируваткарбоксилаза, катализирующая данную
реакцию,
—
митохондриальный
фермент,
коферментом которого является биотин. Реакция
протекает с использованием АТФ.
130. Включение субстратов в глюконеогенез
131.
132. Образование оксалоацетата из пирувата
133. Превращение оксалоацетата в малат
134. Превращение оксалоацетата в фосфоенолпируват
135.
136. Гидролиз фруктоза-1,6-бисфосфата и глюкоза-6-фосфата
Отщепление фосфатной группы из фруктозо-1,6бисфосфатаи
глюкозо-6-фосфата
—
также
необратимые реакции глюконеогенеза. В ходе
гликолиза эти реакции катализируют специфические
киназы с использованием энергии АТФ. В
глюконеогенезе они протекают без участия АТФ и
АДФ и ускоряются не киназами, а фосфатазами —
ферментами, принадлежащими к классу гидролаз.
Ферменты фруктозо-1,6-бисфосфатаза и глюкозо-6фосфатаза катализируют отщепление фосфатной
группы от фруктозо-1,6-бисфосфата и глюкозо-6фосфата. После чего свободная глюкоза выходит из
клетки в кровь.
137. Энергетический баланс глюконеогенеза из пирувата
Суммарный результат глюконеогенеза из пируватавыражается следующим уравнением:
2 Пируват + 4 АТФ + 2 ГТФ + 2 (NADH+Н+) + 4 Н2О →
Глюкоза + 4 АДФ + 2 ГДФ +6 Н3РО4 + 2 NAD+.
138. Синтез глюкозы из лактата
Лактат, образованный в анаэробном гликолизе, неявляется
конечным
продуктом
метаболизма.
Использование лактата связано с его превращением в
печени в пируват. Лактат как источник пирувата важен
не столько при голодании, сколько при нормальной
жизнедеятельности организма. Его превращение в
пируват и дальнейшее использование последнего
являются способом утилизации лактата.
139.
Лактат, образовавшийся в интенсивно работающихмышцах или в клетках с преобладающим анаэробным
способом катаболизма глюкозы, поступает в кровь, а
затем в печень. В печени отношение NADH/NAD+
ниже, чем в сокращающейся мышце, поэтому лактатдегидрогеназная реакция протекает в обратном
направлении, т.е. в сторону образования пирувата из
лактата. Далее пируват включается в глюконеогенез, а
образовавшаяся глюкоза поступает в кровь и
поглощается скелетными мышцами.
140.
Цикл Кори выполняет 2 важнейшие функции:1 — обеспечивает утилизацию лактата;
2
— предотвращает накопление лактата и, как
следствие этого, опасное снижение рН (лактоацидоз).
Часть пирувата, образованного из лактата, окисляется
печенью до СО2 и Н2О. Энергия окисления может
использоваться для синтеза АТФ, необходимого для
реакций глюконеогенеза.
141. Синтез глюкозы из аминокислот
В условиях голодания часть белков мышечной тканираспадается до аминокислот,
которые далее
включаются в процесс катаболизма. Аминокислоты,
которые при катаболизме превращаются в пируват или
метаболиты цитратного цикла, могут рассматриваться
как потенциальные предшественники глюкозы и
гликогена и носят название гликогенных. Например,
оксалоацетат, образующийся из аспарагиновой
кислоты, является промежуточным продуктом как
цитратного цикла, так и глюконеогенеза.
142.
Из всех аминокислот, поступающих в печень,примерно 30% приходится на долю аланина. Это
обуславливает
следующая
последовательность
событий (глюкозо-аланиновый цикл): глюкоза в
мышцах → пируват в мышцах → аланин в мышцах →
аланин в печени → глюкоза в печени → глюкоза в
мышцах. Весь цикл не приводит к увеличению
количества глюкозы в мышцах, но он решает
проблемы транспорта аминного азота из мышц в
печень и предотвращает лактоацидоз.
143. Синтез глюкозы из глицерола
Глицерол образуется при гидролизе триацил-глицеролов,главным образом в жировой ткани. Использовать его могут
только те ткани, в которых имеется фермент глицерол
киназа, например печень, почки. Этот АТФ-зависимый
фермент катализирует превращение глицерола в αглицерофосфат (глицерол-3-фосфат). При включении
глицерол-3-фосфата в глюконеогенез происходит его
дегидрирование
NAD-зависимой
дегидрогеназой
с
образованием дигидроксиацетонфосфата, который далее
превращается в глюкозу.