
















 Биология
БиологияПохожие презентации:










Оплодотворение и дробление
1.
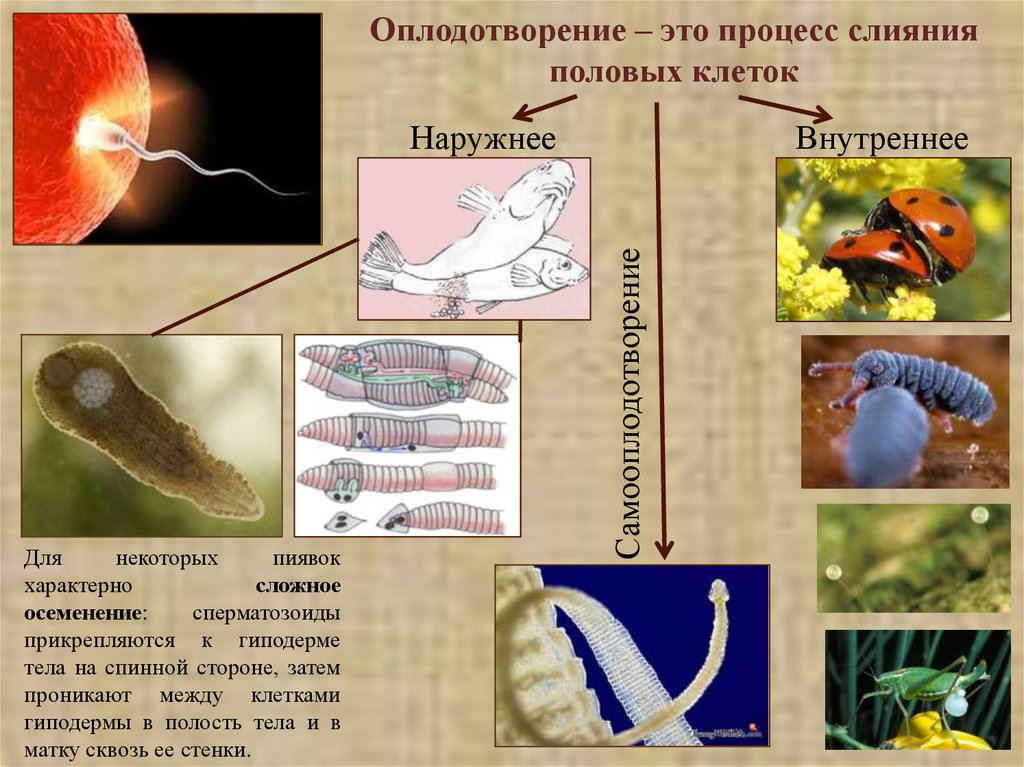
Оплодотворение – это процесс слиянияполовых клеток
Для
некоторых
пиявок
характерно
сложное
осеменение:
сперматозоиды
прикрепляются к гиподерме
тела на спинной стороне, затем
проникают между клетками
гиподермы в полость тела и в
матку сквозь ее стенки.
Внутреннее
Самооплодотворение
Наружнее
2.
Схематическое изображение процесса оплодотворения (В.Г.Елисеев, 1983)1 – цитоплазма яйцеклетки; 2 – ядро яйцеклетки; 3 – блестящая оболочка; 4 –
фолликулярный эпителий; 5 – головка сперматозоида; 6 – шейка сперматозоида; 7 – хвост
сперматозоида; 8 – воспринимающий бугорок; 9 – оболочка оплодотворения; 10 – женский
пронуклеус; 11 – мужской пронуклеус; 12 – веретено между центриолями; 13 – синкарион
3.
1 фаза – СБЛИЖЕНИЕСперматозоиды двигаются благодаря:
- Хемотаксису (pH=7-7,5 – быстро перемещаются по направлению к
яйцеклеткам, а рН=5,8-6,5 парализует их);
- Гамонам (М. Гартман, 1940),
веществам яйцеклеток, возбуждающе
влияющим на активность сперматозоидов, они видоспецифичны;
- Реотаксису, т.е. способности двигаться против тока жидкости,
направленного из яйцевода;
- Перистальтике маточных труб;
- Мерцательному движению ресничек эпителия маточных труб.
- Разность потенциалов между
электрозарядностью
семенной
жидкости (+) и яйцеклетки (-).
Сперматозоид млекопитающего:
А – общий вид; Б – в возбужденном состоянии:
1 – акросома; 2 – ядро; 3–4 – центриоли;
5 – митохондриальная спираль;
6 – осевая нить (цитоскелет шейки и хвоста)
4.
2 фаза – ПРОНИКНОВЕНИЕУ млекопитающих в яйцеклетку проникает лишь один сперматозоид моноспермия. У беспозвоночных животных, рыб, амфибий, рептилий и птиц
возможна полиспермия (проникает несколько сперматозоидов), но в слиянии
ядер (оплодотворении) принимает участие только один.
Полиспермия
Физиологическая
Патологическая
При патологической в яйцо проникают «лишние» спермии, возникают
уродства и ранняя гибель зародыша («лишние» центросомы образуют
«лишние» центры дробления и идет «уродливое» дробление).
5.
Взаимодействию гамет предшествует «узнавание» и акросомная реакция.«Узнавание» происходит по принципу «ключ-замок» (фертилизинантифертилизин). Ферменты, выделяемые из акросом разрушают лучистый
венец, расщепляют гликозаминогликаны вторичной (блестящей) оболочки
яйцеклетки (трипсин – разрушает фолликулярные клетки и блестящую
облочку, гиалуронидаза – разжижает вещества склеивающие клетки
лучистого венца и способствует проникновению сквозь него).
Акросомная реакция сперматозоида
А-В – слияние наружной мембраны акросомы и мембраны сперматозоида;
Г-Д – полимеризация актина и образование акросомного выроста.
Излияние содержимого акросомного пузырька 1 – мембрана акросомы; 2 - мембрана
сперматозоида; 3 – глобулярный актин; 4 – ферменты акросомы;; 5 – байндин;
6 – вырост акросомы; 7 – актиновые микрофиламенты; 8 – ядро сперматозоида.
6.
В цитоплазму яйцеклетки проникает головка, шейка и часть хвостового отдела(митохондриальное влагалище). Проникновение сперматозоида значительно
усиливает процессы внутриклеточного обмена, что связано с повышением
дыхания и активизацией ферментативных систем яйцеклетки.
Успех определяется условиями
жидкой среды (особенно при
наружном
оплодотворении).
Реакция среды должна быть
слабо щелочной. Непременное
условие – наличие солей NaCl,
KCl,
CaCl2,
MgCl2
и
гидроксильных ионов.
После проникновения спермия
через студенистую оболочку
начинаются
изменения
поверхностных слоев яйца –
кортикальная реакция (распад
кортикальных
гранул,
их
содержимое
сливается
с
желточной
оболочкой
с
образованием
оболочки
оплодотворения ).
7.
Медленный блокСутью этой реакции является зкзоцитоз содержимого кортикальных гранул, которое
оказывается между мембраной яйцеклетки и ее первичной оболочкой. Реакция
запускается с увеличением концентрации Са+2 в ооплазме (запасы в ЭПС). Механизм
кортикальной реакции сходен с таковым акросомальной реакции и происходит в
течение 1-2 минут после проникновения.
Содержимое кортикальных гранул включает ферменты, растворяющие белки,
которые соединяют вителиновую оболочку с мембраной, и мукополисахариды,
создающие осмотический градиент а, следовательно, привлекающие воду.
В результате первичная оболочка с оставшимися на ней сперматозоидами отделяется от
мембраны яйцеклетки. Вместе со слоем мукополисахаридов она образует оболочку
оплодотворения.
У
морского
ежа,
наблюдается
изменение
байндин-рецепторов
и
выделение
гиалина,
одевающего
яйцеклетку
сплошным
слоем.
У млекопитающих не наблюдается
образования оболочки оплодотворения,
однако выделение ферментов из
кортикальных гранул приводит к
изменению
ZРЗ-рецепторов
и
отсоединению от них сперматозоидов. Кортикальная реакция и
Это явление называется реакцией образование оболочки
оплодотворения
прозрачной оболочки.
8.
Быстрый блок (ББ описан у морских ежей и у амфибий)Сразу после того, как первый сперматозоид вступает в контакт с яйцом, путем
изменения электрического потенциала мембраны (К+-Nа+-насос), в течение 0,1 секунды
мембрана яйцеклетки деполяризуется (величина потенциала становится равной
значению от -10 до +20 мВ, в норме U≈-70 мВ). Через 1 минуту величина потенциала
возвращается к исходному значению. Спермии способны сливаться только с
мембранами, обладающими потенциалом покоя,
У млекопитающих ББ, по-видимому, нет, так как он не играет существенной роли
из-за малого числа сперматозоидов, достигающих места оплодотворения. У рептилий и
птиц (яйца богаты желтком, в яйцеклетку проникает несколько сперматозоидов, но все
они, кроме одного разрушаются. Факторы разрушения сперматозоидов пока остаются
неизвестными.
Деполяризация мембраны яйцеклетки
9.
3 фаза – СИНКАРИОНОбразуются ♂ и ♀ пронуклеусы с последующим их слиянием. У многих
видов животных ядра ♂ и ♀ клеток во время сближения переходят в состояние
метафазы, затем их хромосомы образуют единую материнскую «звезду», но
уже с 2n набором. В других случаях ядра вначале сливаются и затем переходят
в состояние кариокинеза. Одновременно внесенные сперматозоидом
центриоли расходятся к полюсам клетки, и этот одноклеточный зародыш –
зигота вступает во второй период эмбрионального развития – период
дробления.
Начало дробления
Зигота
♀ пронуклеус
♂ пронуклеус
10.
Метаболические события в яйце:1. Увеличивается вязкость цитоплазмы и ее проницаемость;
2. Интенсивно перемещаются составные части ооплазмы, иногда
наблюдается их расслоение – ооплазматическая сегрегация, что определяет
пространственную организацию будущего зародыша.
3. Резко изменяется обмен аминокислот, увеличивается содержание
свободных
аминокислот,
что
является
показателем
усиления
диссимиляторных процессов;
4. Усиливается обмен веществ, в первые 10 минут отмечено усиление
углеводного обмена, заметное потребление гликогена;
5. Резко возрастает (в 100 и более раз) фосфатный обмен, калийный (в 10 и
более раз) и кальциевый
6. Возрастает
активность
ферментов
цитоплазмы,
особенно
протеолитических;
7. Рибосомы объединяются в полисомы;
8. Активизируется синтез белков и новых и-РНК (нерибосомальных) (зрелая
яйцеклетка имеет все необходимое для синтеза белков и нуклеиновых
кислот).
11.
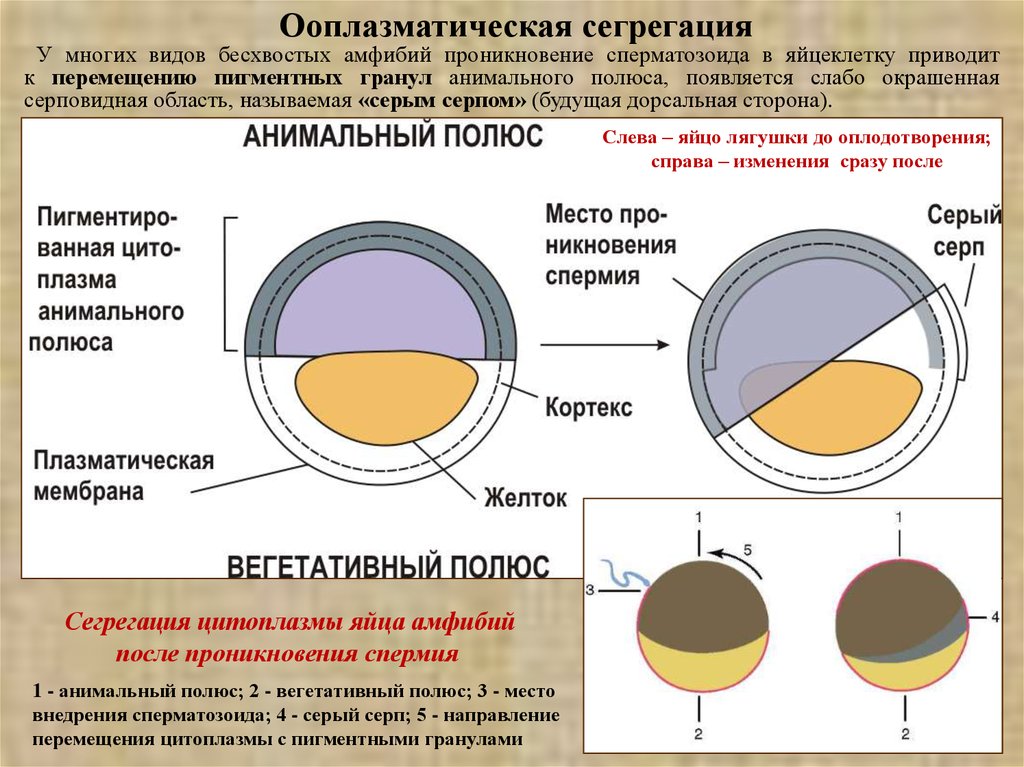
Ооплазматическая сегрегацияУ многих видов бесхвостых амфибий проникновение сперматозоида в яйцеклетку приводит
к перемещению пигментных гранул анимального полюса, появляется слабо окрашенная
серповидная область, называемая «серым серпом» (будущая дорсальная сторона).
Слева – яйцо лягушки до оплодотворения;
справа – изменения сразу после
Сегрегация цитоплазмы яйца амфибий
после проникновения спермия
1 - анимальный полюс; 2 - вегетативный полюс; 3 - место
внедрения сперматозоида; 4 - серый серп; 5 - направление
перемещения цитоплазмы с пигментными гранулами
12.
ДРОБЛЕНИЕряд последовательных митотических делений оплодотворенного или
инициированного к развитию яйца
1.
2.
3.
4.
5.
Правила:
Правило Пфлюгера: веретено всегда тянется в направлении
наименьшего сопротивления.
Правило Бальфура: скорость голобластического дробления обратно
пропорциональна количеству желтка (желток затрудняет деление как
ядра, так и цитоплазмы).
Правило Сакса: клетки обычно делятся на равные части, и плоскость
каждого нового деления пересекает плоскость предшествующего
деления под прямым углом.
Правило Гертвига: ядро и веретено обычно располагаются в центре
активной протоплазмы. Ось каждого веретена деления располагается
по длинной оси массы протоплазмы. Плоскости деления обычно
пересекают массу протоплазмы под прямым углом к ее осям.
Правило Гертвига–Сакса: клеточное ядро стремится расположиться в
центре чистой, свободной от желтка цитоплазмы; веретено клеточного
деления стремится расположиться по направлению наибольшего
протяжения свободной от желтка цитоплазмы.
13.
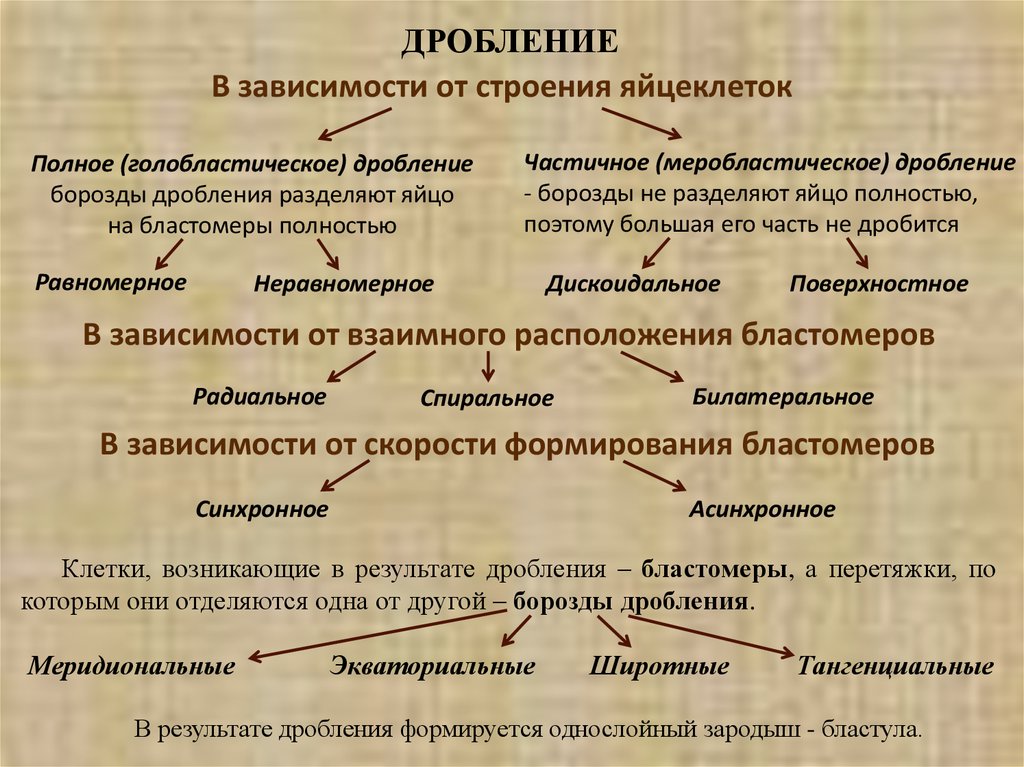
ДРОБЛЕНИЕВ зависимости от строения яйцеклеток
Полное (голобластическое) дробление
борозды дробления разделяют яйцо
на бластомеры полностью
Равномерное
Частичное (меробластическое) дробление
- борозды не разделяют яйцо полностью,
поэтому большая его часть не дробится
Неравномерное
Дискоидальное
Поверхностное
В зависимости от взаимного расположения бластомеров
Радиальное
Спиральное
Билатеральное
В зависимости от скорости формирования бластомеров
Синхронное
Асинхронное
Клетки, возникающие в результате дробления – бластомеры, а перетяжки, по
которым они отделяются одна от другой – борозды дробления.
Меридиональные
Экваториальные
Широтные
Тангенциальные
В результате дробления формируется однослойный зародыш - бластула.
14.
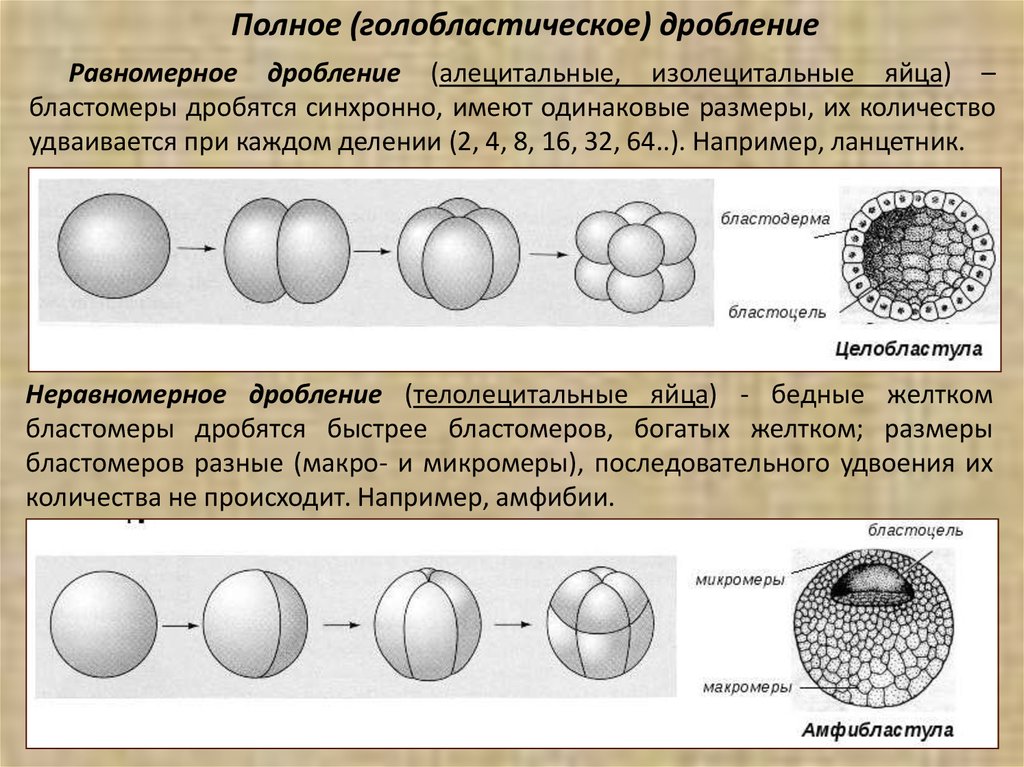
Полное (голобластическое) дроблениеРавномерное дробление (алецитальные, изолецитальные яйца) –
бластомеры дробятся синхронно, имеют одинаковые размеры, их количество
удваивается при каждом делении (2, 4, 8, 16, 32, 64..). Например, ланцетник.
Неравномерное дробление (телолецитальные яйца) - бедные желтком
бластомеры дробятся быстрее бластомеров, богатых желтком; размеры
бластомеров разные (макро- и микромеры), последовательного удвоения их
количества не происходит. Например, амфибии.
15.
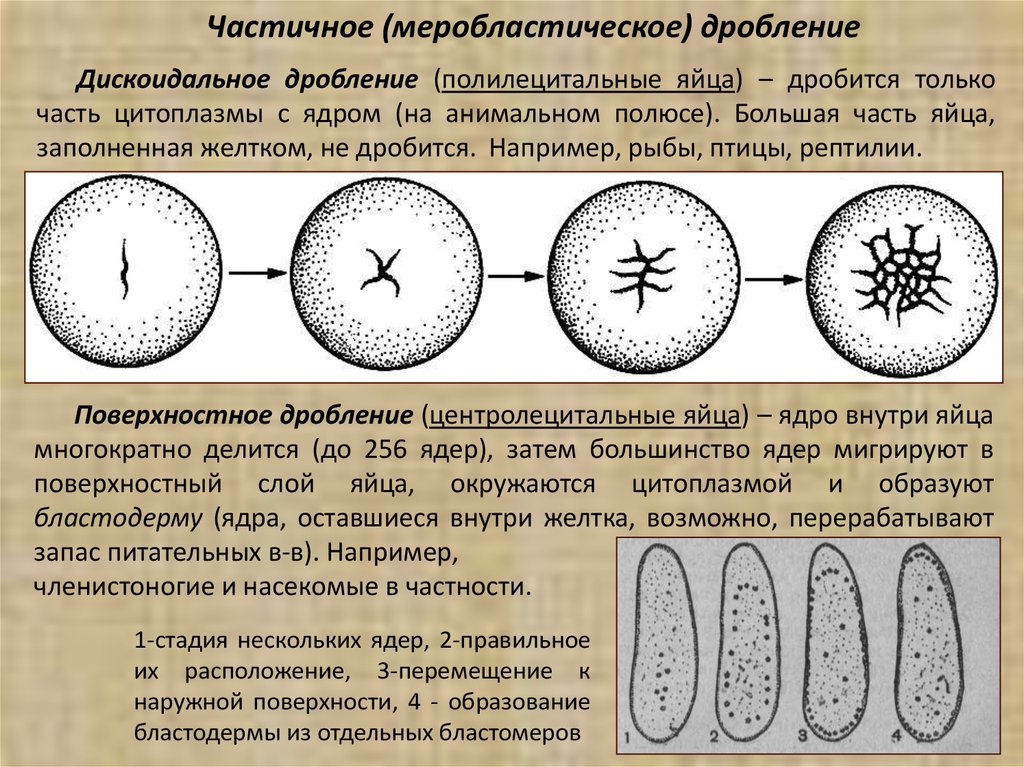
Частичное (меробластическое) дроблениеДискоидальное дробление (полилецитальные яйца) – дробится только
часть цитоплазмы с ядром (на анимальном полюсе). Большая часть яйца,
заполненная желтком, не дробится. Например, рыбы, птицы, рептилии.
Поверхностное дробление (центролецитальные яйца) – ядро внутри яйца
многократно делится (до 256 ядер), затем большинство ядер мигрируют в
поверхностный слой яйца, окружаются цитоплазмой и образуют
бластодерму (ядра, оставшиеся внутри желтка, возможно, перерабатывают
запас питательных в-в). Например,
членистоногие и насекомые в частности.
1-стадия нескольких ядер, 2-правильное
их расположение, 3-перемещение к
наружной поверхности, 4 - образование
бластодермы из отдельных бластомеров
16.
Радиальное дробление (иглокожие, хордовые) - после первых делений зиготыбластомеры анимального полушария лежат строго над бластомерами
вегетативного полушария.
Спиральное дробление (кольчатые черви, моллюски, членистоногие) бластомеры смещены друг относительно друга.
17.
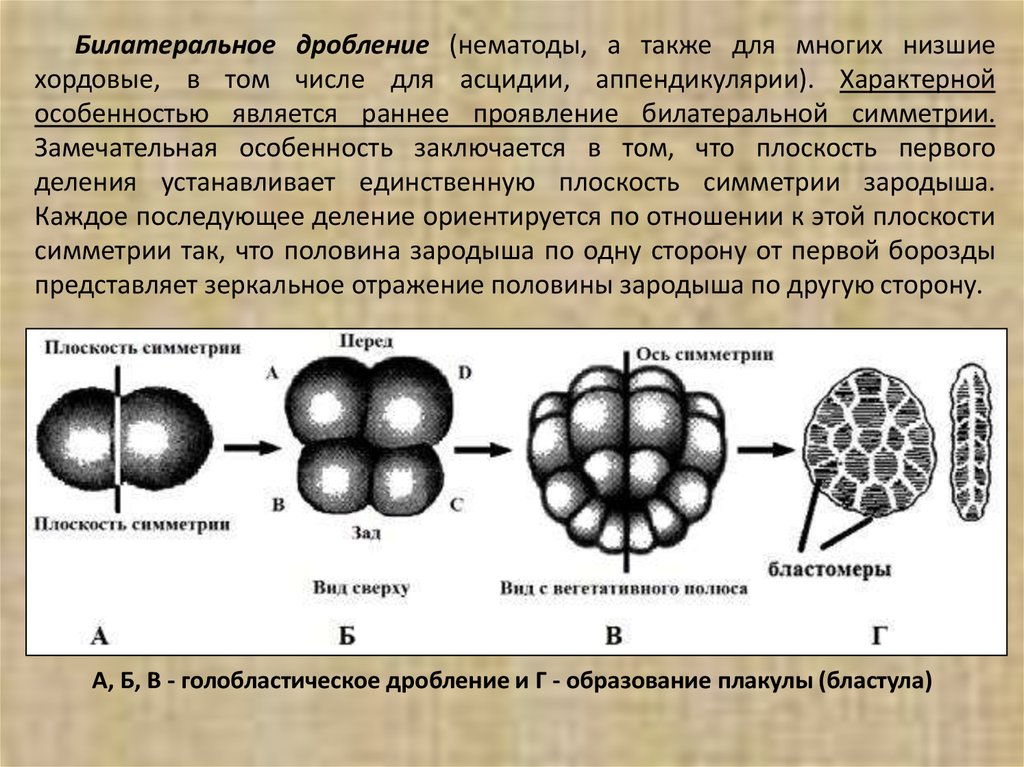
Билатеральное дробление (нематоды, а также для многих низшиехордовые, в том числе для асцидии, аппендикулярии). Характерной
особенностью является раннее проявление билатеральной симметрии.
Замечательная особенность заключается в том, что плоскость первого
деления устанавливает единственную плоскость симметрии зародыша.
Каждое последующее деление ориентируется по отношении к этой плоскости
симметрии так, что половина зародыша по одну сторону от первой борозды
представляет зеркальное отражение половины зародыша по другую сторону.
А, Б, В - голобластическое дробление и Г - образование плакулы (бластула)
18.
При радиальном и спиральном дроблениях обычноразличаются и сроки определения будущей «судьбы»
клеток, т. е. того, какая ткань разовьется в конечном счете из
той или иной их группы.
При радиальном дроблении развитие регуляционное,
«судьба» клеток определяется на относительно поздней
стадии, и разделив в экспериментальных условиях
четырехклеточный зародыш (например, морской звезды) на
отдельные клетки, можно вырастить каждую из них в
целую особь.
И наоборот, если «судьба» клеток определяется очень
рано, то экспериментальное разделение четырехклеточного
зародыша (например, кольчеца) приведет к образованию
только четырех его «четвертушек». Такое развитие
называют мозаичным; оно характерно для спирального
дробления.
19.
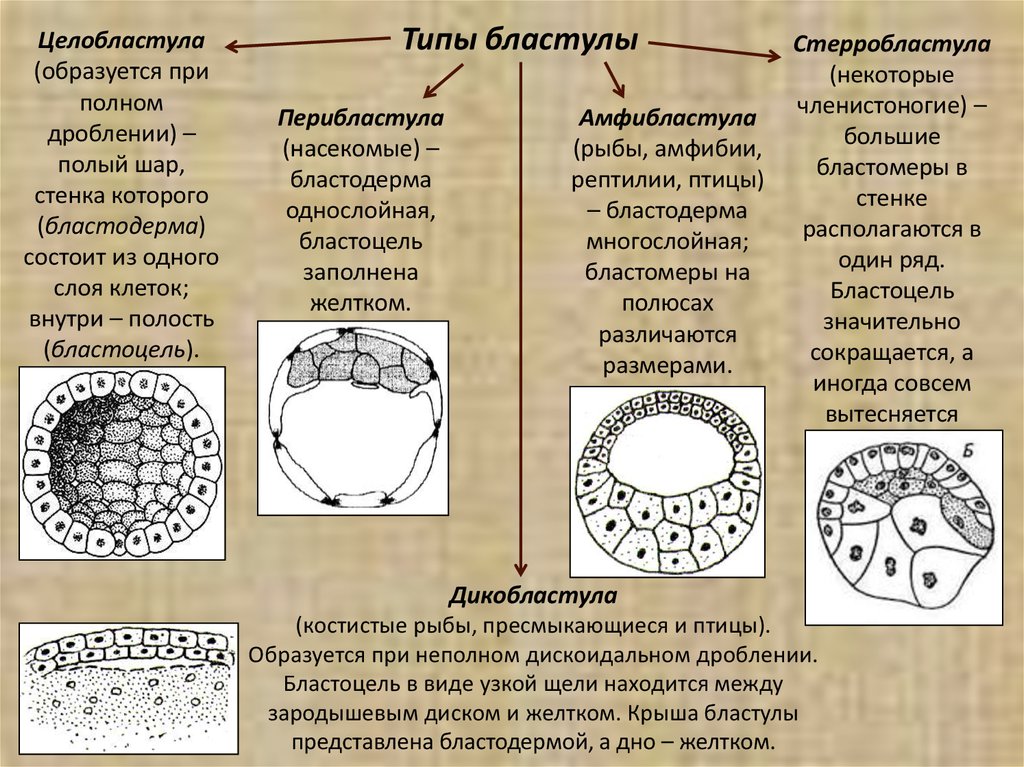
Целобластула(образуется при
полном
дроблении) –
полый шар,
стенка которого
(бластодерма)
состоит из одного
слоя клеток;
внутри – полость
(бластоцель).
Типы бластулы
Перибластула
(насекомые) –
бластодерма
однослойная,
бластоцель
заполнена
желтком.
Амфибластула
(рыбы, амфибии,
рептилии, птицы)
– бластодерма
многослойная;
бластомеры на
полюсах
различаются
размерами.
Стерробластула
(некоторые
членистоногие) –
большие
бластомеры в
стенке
располагаются в
один ряд.
Бластоцель
значительно
сокращается, а
иногда совсем
вытесняется
Дикобластула
(костистые рыбы, пресмыкающиеся и птицы).
Образуется при неполном дискоидальном дроблении.
Бластоцель в виде узкой щели находится между
зародышевым диском и желтком. Крыша бластулы
представлена бластодермой, а дно – желтком.
20.
21.
22.
Классификация иц по количеству желткаПолилецитальные
— большое
количество желтка
(членистоногие,
рептилии, птицы,
рыбы, кроме
осетровых)
Алецитальные
— не содержат желтка
(плацентарные
млекопитающие)
Мезолецитальные
— среднее
количество желтка
(осетровые,
амфибии)
Олиголецитальные
— малое количество
желтка (моллюски,
иглокожие,
млекопитающие)
23.
Классификация яиц по расположению желткаТелолецитальные
желток смещен к
вегетативному полюсу.
Противоположный полюс
- анимальный (некоторые
полилецитальные и все
мезолецитальные яйца)
Гомолецитальные
(изолецитальные) —
желток распределен
равномерно
(олиголецитальные
яйца)
Центролецитальные
—желток расположен в центре
(некоторые полилецитальные членистолногие). Анимальновегетативная полярность не
выражена, т.к. место выделения
редукционных телец м.б.
различным. По переферии
находится ободок свободной от
желтка цитоплазмы. Центр и
периферия связаны
цитоплазматическими
мостиками, все промежуточное
пространство занято желтком