







































 Химия
ХимияПохожие презентации:

")

")






Обмен липидов
1. Обмен липидов
2.
МОБИЛИЗАЦИЯ ЖИРОВМобилизации жиров (липолиз) – это гидролиз
собственных ТАГ до глицерола и жирных кислот.
Гидролиз внутриклеточного жира осуществляется под действием
гормончувствительной липазы - ТАГ-липазы.
Образовавшийся диацилглицерол другие тканевые липазы
(диацилглицероллипаза, моноацилглицероллипаза) гидролизуют до
глицерола и жирных кислот.
3. Катаболизм липидов. Внутриклеточный липолиз.
Catecholaminesкатехоламины
Клеточная
Cell
membrane
мембрана
АденилатAdenylate
cyclase
циклаза
АТФ
ATP
Жирные
и
Plasmaкислоты
fatty acids
глицерин
плазмы
& glycerol
цАМФ
cAMP
Гормон-чувствит.
липаза
Протеинкиназа
Protein kinase
Гормон-чувствит.
липаза
HSL = Hormone Sensitive Lipase
Гормон-чувствит.
липаза Р
Triglycerides
Триглицериды
4. Катаболизм липидов. Окисление глицерина.
Глицеринглицеролкиназа
АТФ
АДФ
Глицерол-3фосфат
глицерол-3-фосфат
дегидрогеназа
НАД
НАДН2
Дигидроксиацетонфосфат
триозофосфатизомераза
Глицеральдегид-3фосфат
Гликолиз
5. Окисление глицерола
При окислении глицерола до СО2 иН2О выделяется 22 молекулы АТФ
1 НАДН2 = 3 АТФ
Гликолиз от ГАФ до ПВК = 5 АТФ
(2СФ+ НАДН2–3)
ПВК переходит в ацетил КоА – НАДН2
=3 АТФ
Ацетил КоА идет в ЦТК = 12 АТФ
- 1 АТФ на глицеролкиназу
6.
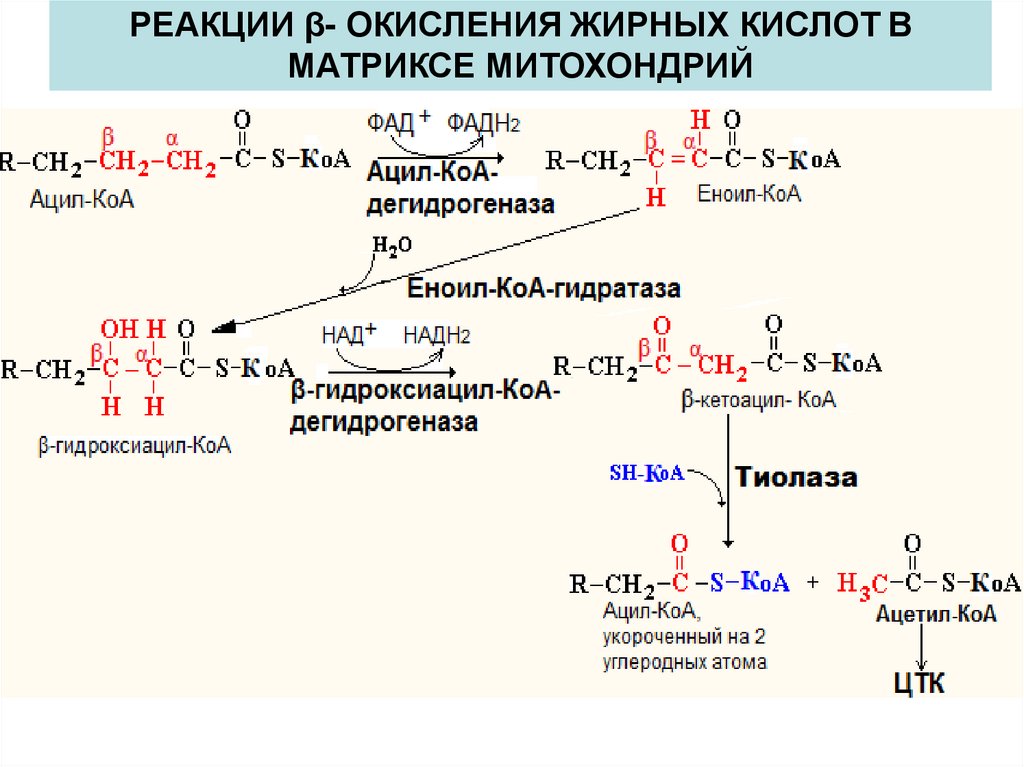
β- ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТβ-Окисление - специфический путь катаболизма жирных
кислот, при котором от карбоксильного конца жирной
кислоты последовательно отделяется по 2 атома
углерода в виде ацетил-КоА.
Реакции β-окисления и окисления ацетил-КоА в ЦТК и
дыхательной цепи служат одним из основных источников
энергии для синтеза АТФ.
β-Окисление жирных кислот происходит только в
аэробных условиях!!!
Протекает в матриксе митохондрий клеток многих тканей:
печени, почках, сердечной и скелетной мышцах.
7. Бета –окисление ЖК
• β-окисление включает следующиеосновные этапы:
• 1) активация жирной кислоты в
цитоплазме клетки
• 2) транспорт активированной ЖК в
митохондрии
• 3) последовательность реакций βокисления
8. Активация жирной кислоты в цитоплазме
АЦИЛ + НSКоА + АТФ АЦИЛ КоА + АМФ + РРнЕ : ацил КоА синтетаза
9. Транспорт в митохондрию
+H2C-N
(CH3)3
O
HC-O C
R
CH2
COOH
ацил-карнитин
В.С.Гулевич
10. Lipid Catabolism. Oxidation of Fatty Acids.
Транспортжирных
кислот
матрикс
Lipid
Catabolism.
Oxidation
of вFatty
Acids.
митохондрий.
Наружная мембрана
митохондрии
Цитозоль
Внутренняя мембрана
митохондрии
Межмембранное
пространство
Матрикс
Карнитин:
ацилтрансфераза II
Карнитин
Карнитин
АЦИЛКарнитин
АЦИЛКарнитин
Карнитин:
ацилтрансфераза I
Транспортер
11.
РЕАКЦИИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ ВМАТРИКСЕ МИТОХОНДРИЙ
12.
ЭНЕРГЕТИЧЕСКИЙ ЭФФЕКТ β- ОКИСЛЕНИЯ ЖИРНЫХКИСЛОТ
1 виток β-окисления жирных кислот дает 17 АТФ:
1 НАДН2→3 АТФ
1 ФАДН2→2 АТФ
1 ацетил-КоА→12 АТФ
n
число атомов углерода в жирной кислоте
n/2 кол-во образовавшихся молекул ацетил-КоА
n/2-1 кол-во циклов в спирали β-окисления ЖК
12
кол-во молекул АТФ, образующихся при окислении
ацетил-КоА в ЦТК
5 кол-во молекул АТФ, которые дают НАДН2 и ФАДН2 при окислении в ЭТЦ
- 1 затраты АТФ на активацию ЖК
13. Регуляция β- ОКИСЛЕНИЯ
Основная регуляция происходитчерез КАТ 1 с помощью ингибитора
малонилКоА.
При высокоуглеводной диете
активность КАТ 1 снижена, при
голодании активность высокая.
АДФ +
АТФ -
14. Связь β- ОКИСЛЕНИЯ с ЦТК и ЦПЭ
β- ОКИСЛЕНИЕ жирных кислотпоставляет в ЦТК, ацетил КоА.
β- ОКИСЛЕНИЕ жирных кислот дает
для дыхательной цепи НАДН2 и
протоны и электроны с ФАДН2.
Таким образом, β- ОКИСЛЕНИЕ идет
только в присутствии кислорода!
15.
ОСОБЕННОСТИ β- ОКИСЛЕНИЯНЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ
16. Карнитин содержащие препараты
L – карнитин – сжигатель жира, БАД дляспортивного питания.
Карнитон - препарат для лечения
сердечно- сосудистой патологии
ЛВ карнитин - стимулирующий препарат
17. Препарат коррекции катаболизма жирных кислот
Милдронат – (содержит мильдоний –ингибитор карнитина) При недостатке
кислорода Бета – окисление
невозможно. Поэтому при ишемических
состояниях отключают этот процесс и
переводят метаболизм на др.
энергетические потоки – гликолиз.
18. Кетоновые тела
АцетонАцетоацетат
β-гидроксибутират
19. Значение кетоновых тел
• Кетоновые тела образуются впечени и экспортируются
в другие ткани в качестве
метаболического топлива.
20. Свойства кетоновых тел
• Высокая растворимость в воде• Высокая доступность в ткани для
энергетического пополнения
Избыточный синтез КТ приводит к
ацидозу в крови.
Ацетон компенсаторно выделяется
при дыхании.
21. Синтез кетоновых тел
Синтез КТ происходит ВМИТОХОНДРИЯХ ПЕЧЕНИ:
1. При голодании
2. При углеводном голодании (диета)
3. При сахарном диабете
4. При напряженной мышечной
работе
22. Кетоновые тела
Причина избыточного синтеза КТ:НЕДОСТАТОЧНОСТЬ ЩУК, который
печень использует для
ГЛЮКОНЕОГЕНЕЗА, поэтому Ацетил
КоА в полном объеме не может идти
в ЦТК и идет на синтез КТ.
23. Синтез и окисление кетоновых тел
SynthesisСинтез
Oxidation
Окисление
(Liver)
(печень)
(Nonhepatic cells)
(внепеченочные
ткани)
ACETYL CoA
22Ацетил
КоА
Тиолаза
Tiolase
ACETYL CoA
22 Ацетил
КоА
-HS-CoA
-HS-КоА
Tiolase
Тиолаза
ACETOACETYL CoA
АцетоацтеилКоА
ГМГКоА –
синтаза
Acetyl
CoAКоА
Ацетил
HS-CoA
-HS-КоА
HS-CoA
-HS-КоА
β-HYDROXYβ –METHYLGLUTARYL КоА
CoA
β-гидроксиметилглутарил
ACETOACETYL CoA
АцетоацетилКоА
Succinate
Сукцинат
HMG CoA lyase
ГМГ КоА - лиаза
-Acetyl
CoAКоА
-Ацетил
NADH++H+
Succinyl CoA
Сукцинил
КоА
NAD
CO2
ACETONE
АЦЕТОН
ACETOACETATE
АЦЕТОАЦЕТАТ
β-HYDROXYBUTYRATE
β-ГИДРОКСИБУТИРАТ
24. Синтез липидов
25.
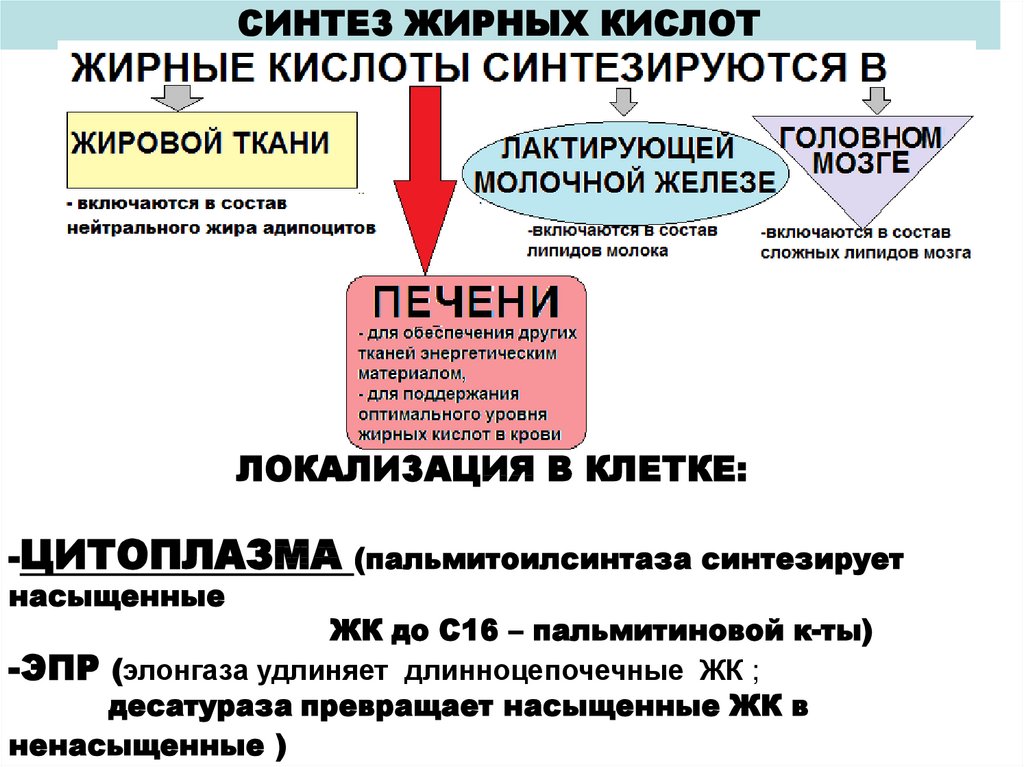
СИНТЕЗ ЖИРНЫХ КИСЛОТЛОКАЛИЗАЦИЯ В КЛЕТКЕ:
-ЦИТОПЛАЗМА (пальмитоилсинтаза синтезирует
насыщенные
ЖК до С16 – пальмитиновой к-ты)
-ЭПР (элонгаза удлиняет длинноцепочечные ЖК ;
десатураза превращает насыщенные ЖК в
ненасыщенные )
26. Синтез жирных кислот
• Синтез жирных кислот происходит послеприема пищи богатой углеводами.
• Субстрат для синтеза ЖК – Ацетил КоА
Транспорт Ацетил КоА в цитоплазму
происходит с помощью цитрата.
• Цитрат имеет возможность выходить из
митохондрии, так как ЦТК ингибирован
избытком АТФ ( регуляторный фермент
изоцитрат ДГ)
27. Транспорт ацетил КоА в цитоплазму
МИТОХОНДРИЯАцетил КоА + ЩУК + Н2О ЦИТРАТ + НSКоА
Е: цитратсинтаза
Так как, регуляторная реакция ЦТК изоцитратдегидрогеназа
аллостерически ингибирована избытком АТФ цитрат с
помощью переносчиков переходит в цитоплазму.
ЦИТОПЛАЗМА
ЦИТРАТ + АТФ + НSКоА Ацетил КоА + ЩУК + АДФ
Е: цитратлиаза
28.
СИНТЕЗ ЖИРНЫХ КИСЛОТ (ЦП)Карбоксилирование ацетил-КоА с
образованием малонил-КоА
Активность фермента ацетил-КоА-карбоксилазы
определяет скорость всех последующих реакций синтеза
жирных кислот.
Регуляция: ИНСУЛИН, ЦИТРАТ – активируют, ГЛЮКАГОН,
ВЖК – ингибируют.
29.
СИНТЕЗ ЖИРНЫХ КИСЛОТСтроение мульферментного комплекса- ПАЛЬМИТОИЛСИНТАЗЫ
Пальмитоилсинтаза - димер из двух полипептидных цепей. Каждая
субъединица содержит 8 доменов: 7 активных центров и ацилпереносящий
белок. Каждая субъединица имеет две SH-группы: одна SH-группа
принадлежит цистеину, другая - остатку фосфопантотеновой кислоты
(производное вит. В5).
30.
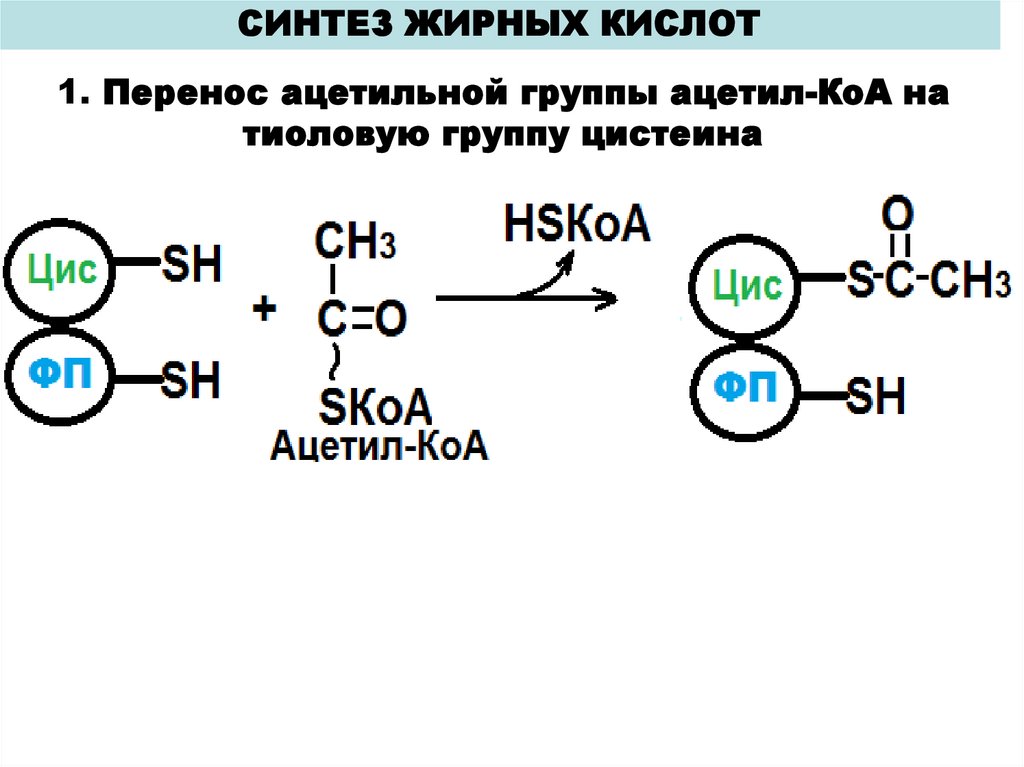
СИНТЕЗ ЖИРНЫХ КИСЛОТ1. Перенос ацетильной группы ацетил-КоА на
тиоловую группу цистеина
31.
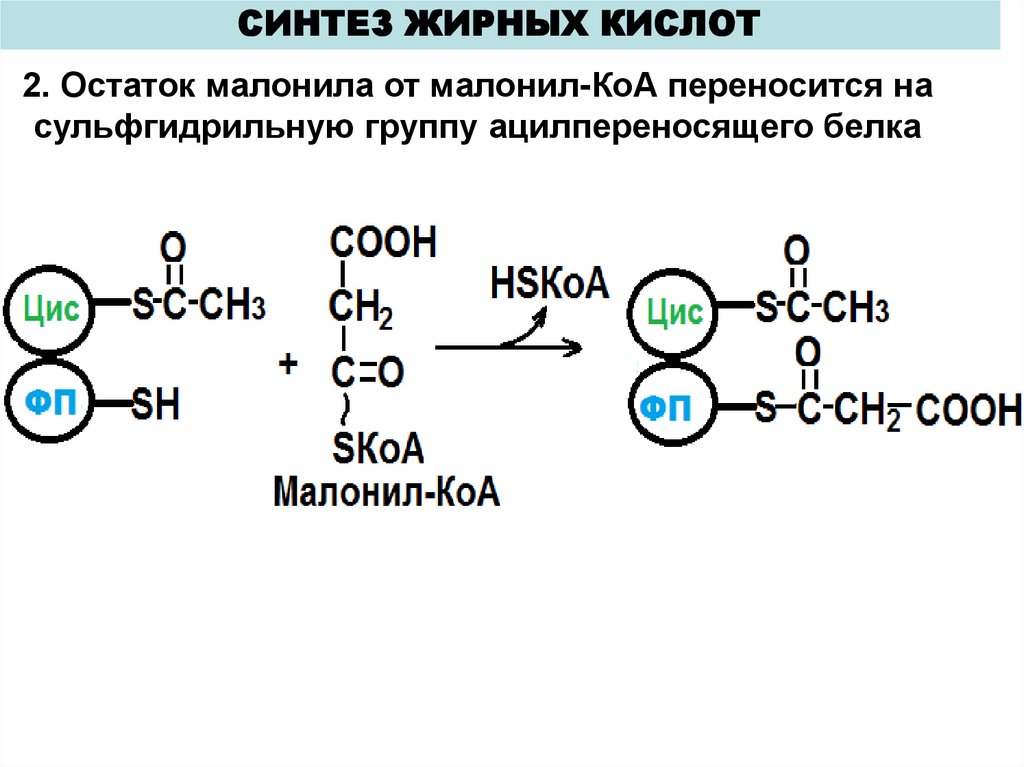
СИНТЕЗ ЖИРНЫХ КИСЛОТ2. Остаток малонила от малонил-КоА переносится на
сульфгидрильную группу ацилпереносящего белка
32.
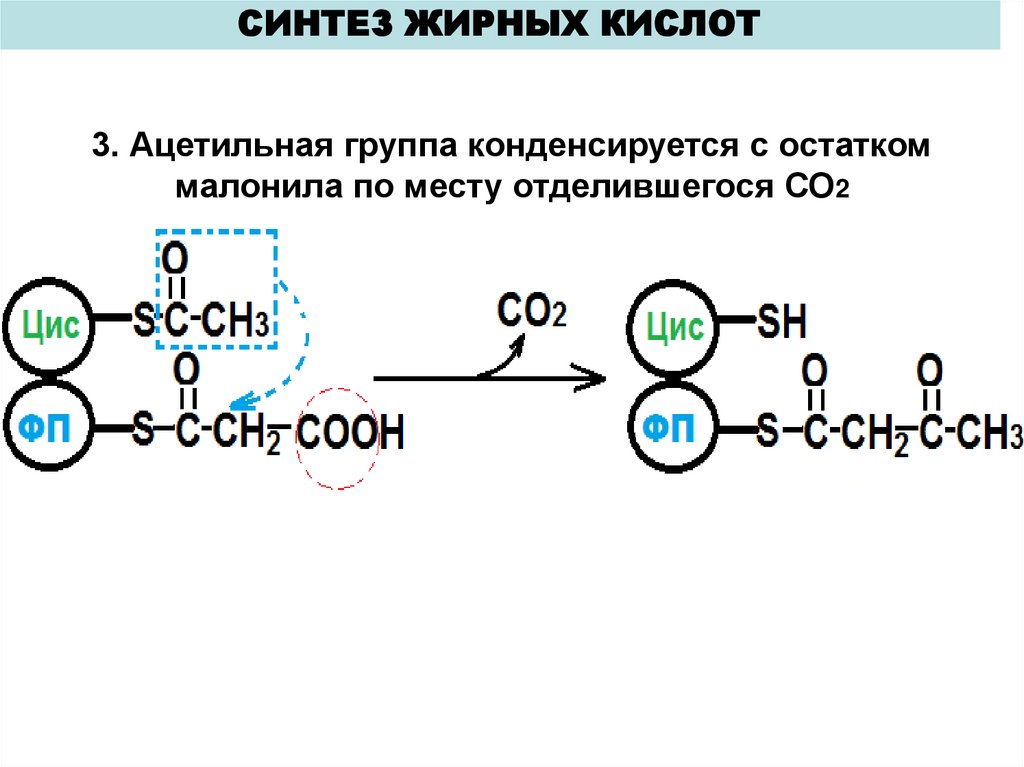
СИНТЕЗ ЖИРНЫХ КИСЛОТ3. Ацетильная группа конденсируется с остатком
малонила по месту отделившегося СО2
33.
СИНТЕЗ ЖИРНЫХ КИСЛОТ4. Кетоацильный остаток восстанавливается, донор
атомов водорода - НАДФН2
34.
СИНТЕЗ ЖИРНЫХ КИСЛОТ5. Дегидратация β-гидроксиацильного остатка
35.
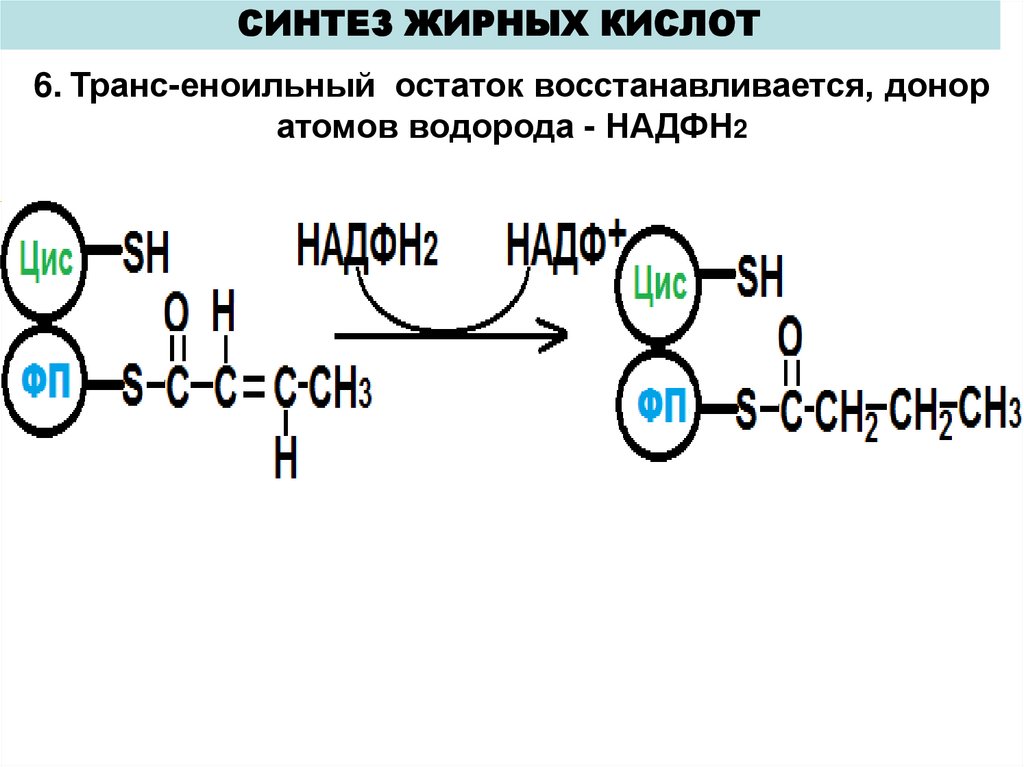
СИНТЕЗ ЖИРНЫХ КИСЛОТ6. Транс-еноильный остаток восстанавливается, донор
атомов водорода - НАДФН2
36.
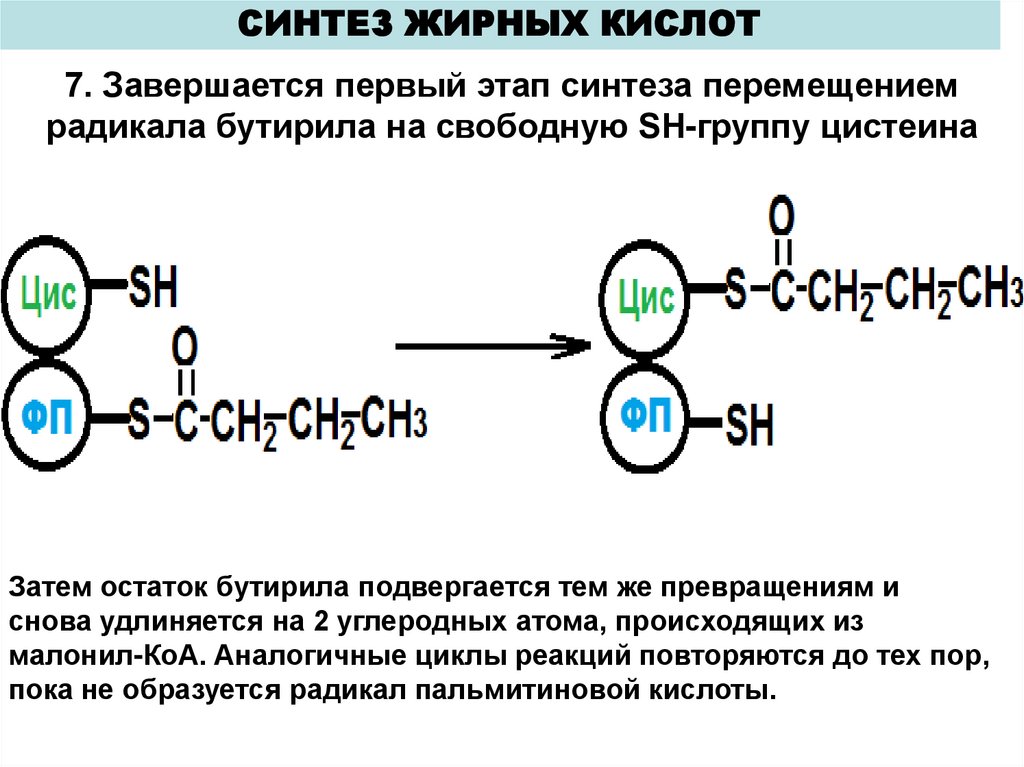
СИНТЕЗ ЖИРНЫХ КИСЛОТ7. Завершается первый этап синтеза перемещением
радикала бутирила на свободную SH-группу цистеина
Затем остаток бутирила подвергается тем же превращениям и
снова удлиняется на 2 углеродных атома, происходящих из
малонил-КоА. Аналогичные циклы реакций повторяются до тех пор,
пока не образуется радикал пальмитиновой кислоты.
37.
ЭЛОНГАЦИЯ ЖИРНЫХ КИСЛОТЭлонгаза использует малонил-КоА в качестве донора
углеродных атомов, и НАДФН2- в качестве
восстановителя.
38.
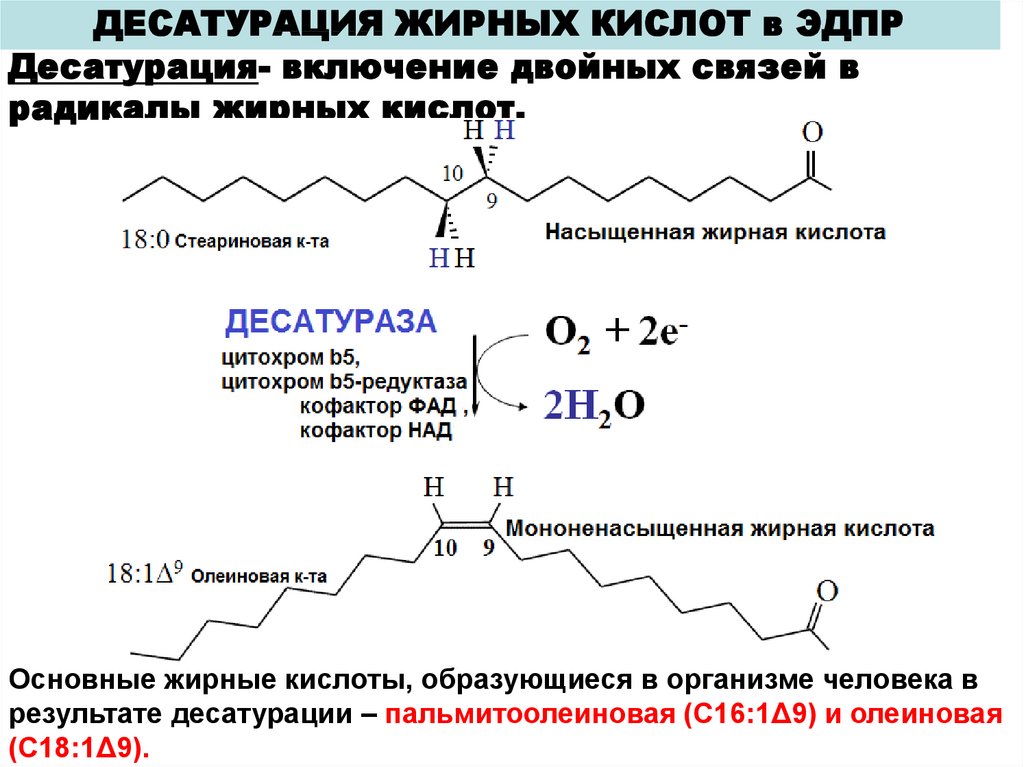
ДЕСАТУРАЦИЯ ЖИРНЫХ КИСЛОТ в ЭДПРДесатурация- включение двойных связей в
радикалы жирных кислот.
Основные жирные кислоты, образующиеся в организме человека в
результате десатурации – пальмитоолеиновая (С16:1Δ9) и олеиновая
(С18:1Δ9).
39. Синтез ТАГ
ПеченьГЛИЦЕРОЛ + АТФ ГЛИЦЕРОЛ-3-Ф + АДФ
Е: глицеролкиназа
Адипоциты
глюкоза ДАФ + НАДН+Н ГЛИЦЕРОЛ-3-ф + НАД
Е: 3-ф-глицерол ДГ
3-ф-глицерол + 2 ацилКоА фосфатидная к-та + 2 НSКоА
Е: ацилтрансфераза
40. Синтез ТАГ и фосфолипидов
Фосфатидная кислотаФосфатидат
фосфатаза
+НОН
-Н3РО4
ЦТФ
ЦДФ + Фн
Присоединение
органического
основания
(этаноламина,
холина, серина,
инозитола и др.
1,2-диглицерид
Ацилтрансфераза
Органич.
основание
Глицерофосфолипид
Нейтральный жир
41. Липотропные и антилипотропные вещества.
ЛИПОТРОПНЫЕ ВЕЩЕСТВАВЕЩЕСТВА
(защищают печень от
Жировой дистрофии )
АНТИЛИПОТРОПНЫЕ
(вызывают жировую дистрофию
печени )
-холин,
- С-четыреххлористый
метионин,
-хлороформ,
-серин,
-этанол,
-витамин B5 (пантотеновая кислота), -фосфор (P)
-витамин B6 (пиридоксин),
-мышьяк (As),
-витамин B12 (кобаламин)
-свинец (Pb),
ГЕПАТОПРОТЕКТОРЫ
-оротовая кислота , В3
(мегадозы),
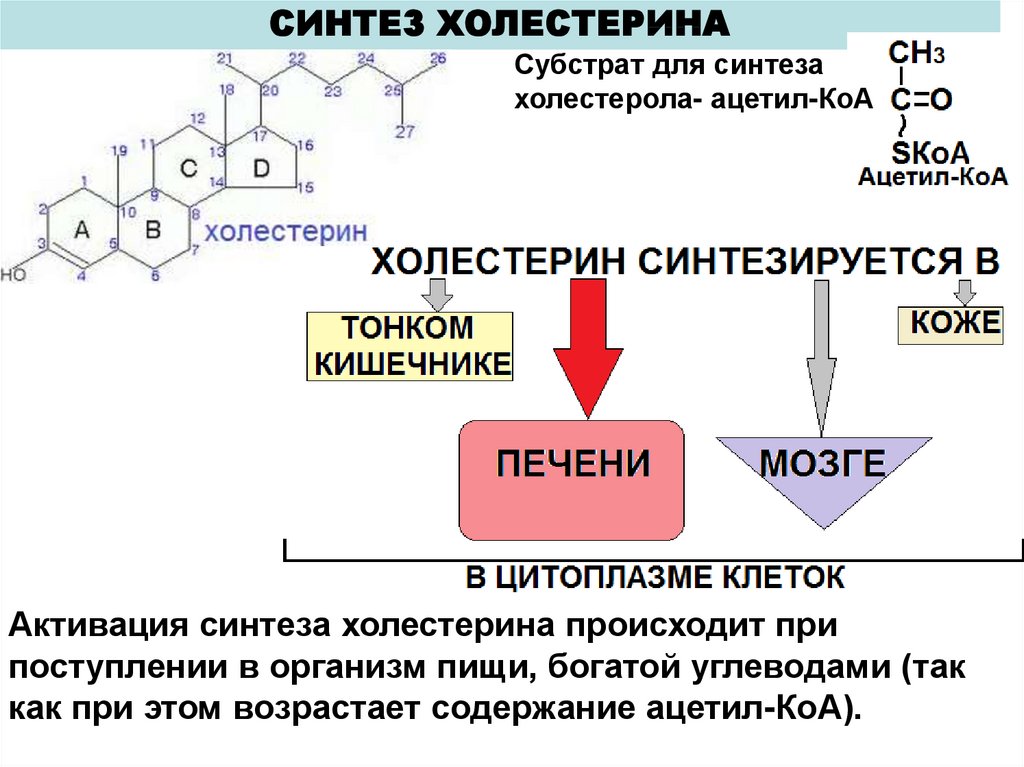
42.
СИНТЕЗ ХОЛЕСТЕРИНАСубстрат для синтеза
холестерола- ацетил-КоА
Активация синтеза холестерина происходит при
поступлении в организм пищи, богатой углеводами (так
как при этом возрастает содержание ацетил-КоА).
43.
СИНТЕЗ ХОЛЕСТЕРИНАОСНОВНЫЕ СТАДИИ СИНТЕЗА ХОЛЕСТЕРИНА:
• синтез мевалоната (С6)
• синтез сквалена из мевалоната (С30)
• циклизация сквалена и образование
холестерина (С27)
44.
СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕМЕВАЛОНАТА
Регуляторная
реакция синтеза
холестерина
45. Регуляция синтеза холестерина
Гидроксиметилглутарил КоАредуктаза (ГМГ редуктаза)
Дефосфорилированная форма –
активная (гормон инсулин)
Фосфорилированная форма –
неактивная (гормон глюкагон)
Экзогенный холестерин - ингибитор
46. Анаболизм липидов. Синтез холестерина
2 ацетил-КоAАцетоацетил-КоA
Ацетил-КoA
-HS-КoA
ГМГ-КоА
Mevalonic
acid
Мевалоновая
кислота
2 ATP
2ADP
CO2
Изомеризация,
конденсация
Сквален (C30 )
Окисление
циклизация
Ланостерол
-3CH3(деметилирование)
Холестерол
47. Лекарственные препараты
С возрастом снижается регуляцияГМГредуктазы экзогенным ХС,
начинает появляться
ГИПЕРХОЛЕСТЕРОЛЕМИЯ
приводящая к атеросклерозу.
Статины – ингибируют ГМГ редуктазу
(МЕВАКОР, МЕВАСТАТИН,
ФЛУВАСТАТИН)
48. Центральная роль ацетилКоА в липидном обмене
49. Схема превращения глюкозы в нейтральный жир
ГЛЮКОЗА -гликолизДАФ
Глицерол-3-фосфат
Пируват
Ацетил-КоА
Жирные
кислоты
Фосфатидная кислота
+НОН
-H3PO4
1,2-диглицерид
НЕЙТРАЛЬНЫЙ ЖИР
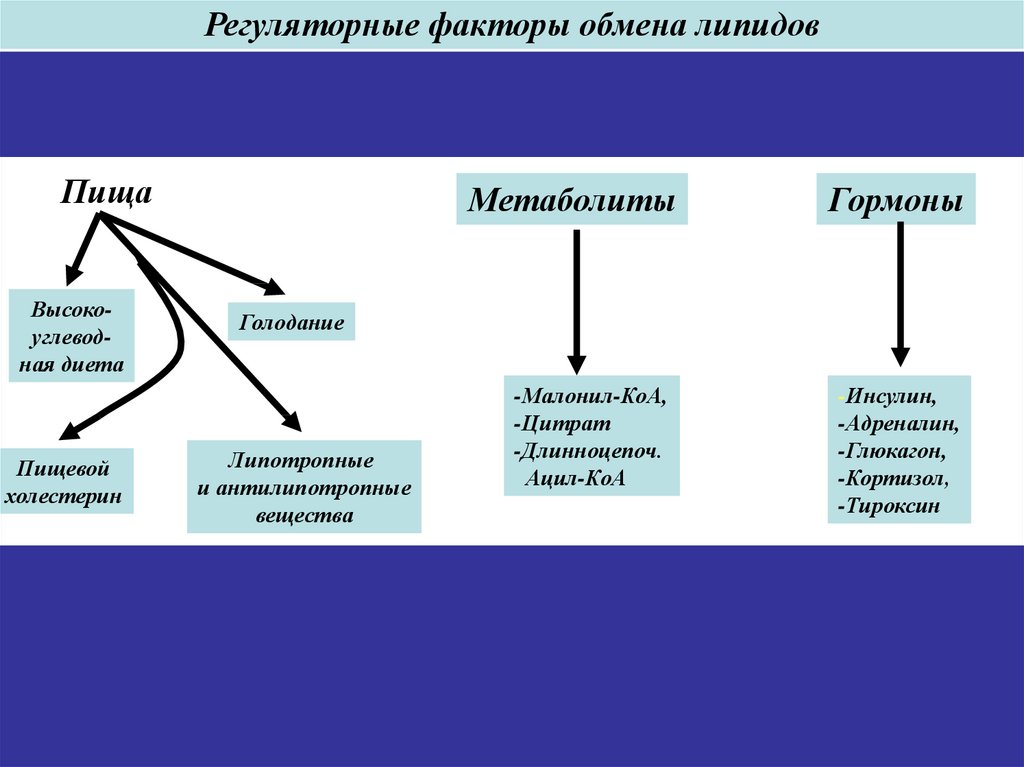
50.
Регуляторные факторы обмена липидовПища
Высокоуглеводная диета
Пищевой
холестерин
Метаболиты
Гормоны
Голодание
Липотропные
и антилипотропные
вещества
-Малонил-КоА,
-Цитрат
-Длинноцепоч.
Ацил-КоА
-Инсулин,
-Адреналин,
-Глюкагон,
-Кортизол,
-Тироксин