

























































 Медицина
МедицинаПохожие презентации:



")






Основные формы иммунного реагирования. Понятие об иммунитете. Виды иммунитета
1.
Основные формы иммунногореагирования. Понятие об
иммунитете. Виды
иммунитета.
2.
•Антитела. Антителообразование.• Киллинг. Иммунный фагоцитоз.
• Иммунная память и толерантность.
• Характеристика видов иммунитета.
Особенности противовирусного,
антипаразитарного и других форм
иммунитета.
3.
• Для гуморального иммунитета характерна выработкаспецифических антител (иммуноглобулинов).
• Антитела - специфические белки гаммаглобулиновой природы, образующиеся в организме в
ответ на антигенную стимуляцию и способные
специфически взаимодействовать с антигеном (in
vivo, in vitro). В соответствии с международной
классификацией совокупность сывороточных белков,
обладающих свойствами антител, называют
иммуноглобулинами.
4.
• Уникальность антител заключается в том, что они способныспецифически взаимодействовать только с тем антигеном,
который вызвал их образование.
• Иммуноглобулины ( Ig ) разделены в зависимости от
локализации на три группы:
• - сывороточные (в крови);
• - секреторные ( в секретах- содержимом желудочно- кишечного
тракта, слезном секрете, слюне, особенно- в грудном молоке)
обеспечивают местный иммунитет (иммунитет слизистых);
• - поверхностные ( на поверхности иммунокомпетентных
клеток, особенно В-лимфоцитов.
5.
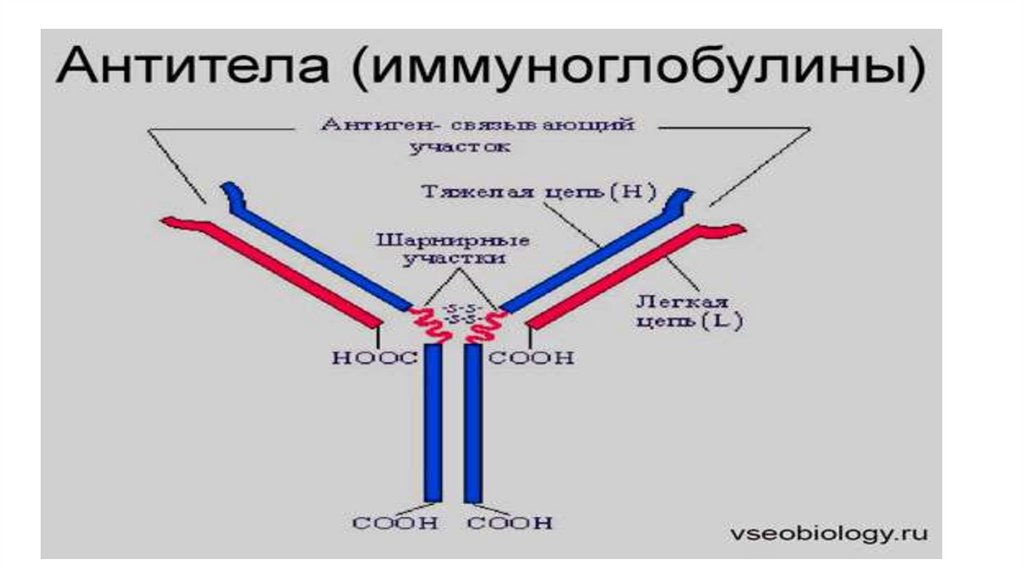
• Любая молекула антител имеет сходное строение ( Y- образнуюформу) и состоит из двух тяжелых ( Н ) и двух легких ( L )
цепей, связанных дисульфидными мостиками.
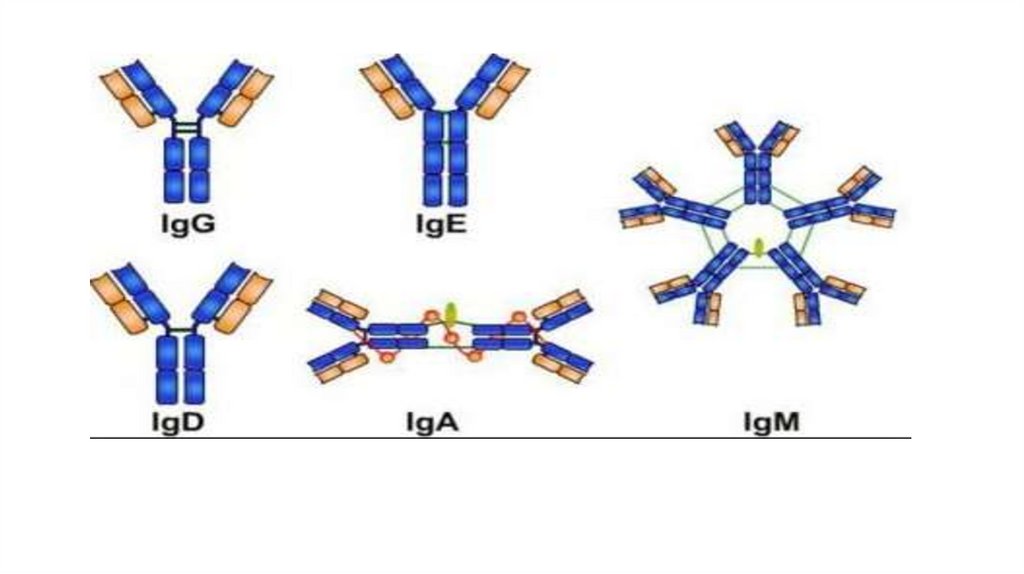
• Известно пять классов иммуноглобулинов, отличающихся по
строению тяжелых цепей, молекулярной массе, физикохимическим и биологическим характеристикам: IgG, IgM, IgA,
IgE, IgD.
• В составе IgG выделяют 4 подкласса ( IgG1, IgG2, IgG3, IgG4 ),
• в составе IgA- два подкласса (IgA1, IgA2 ).
6.
• Основные биологические характеристики антител.• 1. Специфичность - способность взаимодействия с определенным
(своим) антигеном (соответствие эпитопа антигена и активного центра
антител).
• 2. Валентность- количество способных реагировать с антигеном
активных центров
• 3. Афинность - прочность связи между эпитопом антигена и активным
центром антител, зависит от их пространственного соответствия.
• 4. Авидность - интегральная характеристика силы связи между
антигеном и антителами, с учетом взаимодействия всех активных
центров антител с эпитопами. Поскольку антигены часто поливалентны,
связь между отдельными молекулами антигена осуществляется с
помощью нескольких антител.
• 5. Гетерогенность - обусловлена антигенными свойствами антител,
наличием у них трех видов антигенных детерминант.
7.
• Характеристика основных классов иммуноглобулинов.• Ig G. Это основной класс иммуноглобулинов, защищающих
организм от бактерий, токсинов и вирусов.
• В наибольшем количестве IgG- антитела вырабатываются на
стадии выздоровления после инфекционного заболевания .
Только IgG способны транспортироваться через плаценту от
матери к плоду (проходить через плацентарный барьер) и
обеспечивать защиту материнскими антителами плода и
новорожденного.
• В отличие от IgM- антител, IgG- антитела относятся к категории
поздних- появляются позже и более длительно выявляются в
крови.
8.
• IgM.. IgM- наиболее ранний класс антител,образующихся при первичном попадании антигена в
организм. Наличие IgM- антител к соответствующему
возбудителю свидетельствует о свежем
инфицировании (текущем инфекционном процессе).
• IgM у новорожденных- это показатель
внутриутробного заражения (краснуха, ЦМВ,
токсоплазмоз и другие внутриутробные инфекции),
поскольку материнские IgM через плаценту не
проходят.
9.
• IgA. Выделяют сывороточные IgA и секреторные IgA .Секреторные IgAs находятся в слюне, пищеварительных соках,
секрете слизистой носа, в молозиве. Они являются первой
линией защиты слизистых, обеспечивая их местный иммунитет
• IgE. К IgE относятся “антитела аллергии”- реагины. Уровень
IgE повышается при аллергических состояниях, гельминтозах.
• IgD. Их биологическая роль точно не установлена. Полагают,
что IgD участвуют в дифференциации В-клеток, способствуют
развитию антиидиотипического ответа, участвуют в
аутоиммунных процессах.
10.
11.
• С целью определения концентраций иммуноглобулиновотдельных классов применяют несколько методов, чаще
используют метод радиальной иммунодиффузии в геле
(по Манчини)- разновидность реакции преципитации и
ИФА.
• Определение антител различных классов имеет важное
значение для диагностики инфекционных заболеваний.
• Обнаружение антител к антигенам микроорганизмов в
сыворотках крови- важный критерий при постановке
диагноза- серологический метод диагностики.
12.
• Антитела класса IgM появляются в остром периоде заболевания иотносительно быстро исчезают, антитела класса IgG выявляются в
более поздние сроки и более длительно (иногда- годами)
сохраняются в сыворотках крови переболевших, их в этом случае
называют анамнестическими антителами.
• Выделяют понятия: титр антител, диагностический титр,
исследования парных сывороток.
• Реакции взаимодействия антител с возбудителями и их
антигенами (реакция “антиген- антитело”) проявляется в виде
ряда феноменов- агглютинации, преципитации, нейтрализации,
лизиса, связывания комплемента, опсонизации, цитотоксичности
и могут быть выявлены различными серологическими
реакциями.
13.
Любая молекула антител имеет сходное строение ( Yобразную форму) и состоит из двух тяжелых ( Н ) и двухлегких ( L ) цепей, связанных дисульфидными
мостиками. Каждая молекула антител имеет два
одинаковых антигенсвязывающих фрагмента Fab (
fragment antigen binding ), определяющих антительную
специфичность, и один Fc ( fragment constant ) фрагмент,
который не связывает антиген, но обладает
эффекторными биологическими функциями. Он
взаимодействует со “своим” рецептором в мембране
различных типов клеток ( макрофаг, тучная клетка,
нейтрофил).
14.
15.
Антигенсвязывающий центр антителкомплементарен эпитопу антигена по
принципу “ключ - замок” и образован
гипервариабельными областями L- и Н- цепей.
Антитело свяжется антигеном (ключ попадет в
замок) только в том случае, если
детерминантная группа антигена полностью
вместится в щель активного центра антител.
16.
17.
Выявлено пять классов тяжелыхцепей- альфа ( с двумя подклассами),
гамма ( с четырьмя подклассами),
эксилон, мю и дельта. Соответственно
обозначению тяжелой цепи
обозначается и класс молекул
иммуноглобулинов- А, G, E, M и D.
18.
• Антитела относятся преимущественно к γ-глобулиновойфракции белков плазмы крови.Поэтому антитела
получили название иммуноглобулинов, и их обозначают
символом Ig.
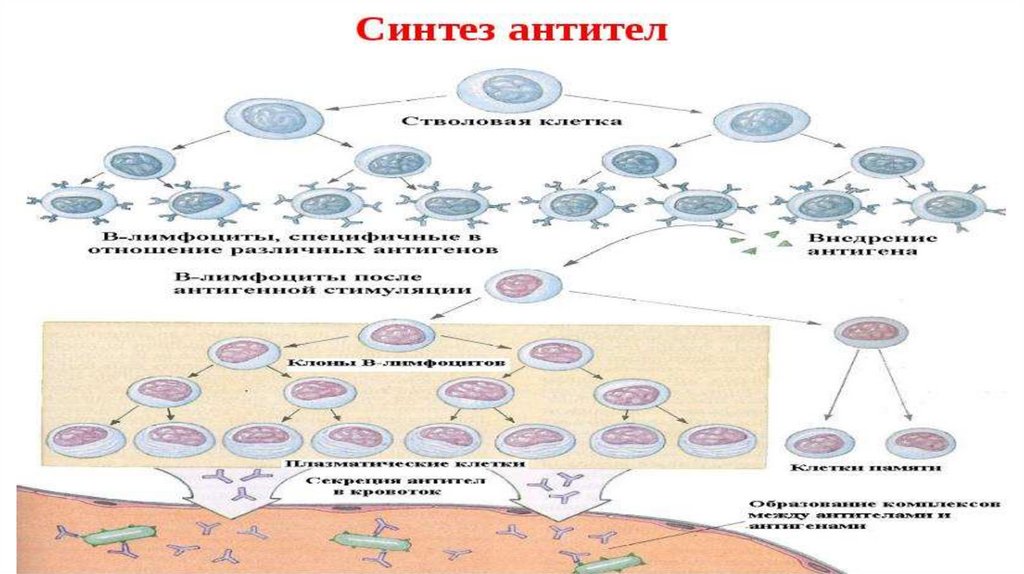
• Антитела синтезируются В-лимфоцитами и их
потомками - плазматическими клетками и в
циркулирующей форме, и в виде рецепторных молекул на
иммунокомпетентных клетках.
19.
20.
21.
• Антителопродукция в ответ на антигенный стимул имеетхарактерную динамику. Ее можно проследить на примере
сывороточных Ig .
• Выделяют
• латентную (индуктивную),
• логарифмическую,
• стационарную фазы и
• фазу снижения.
22.
• В латентную фазу антителопродукция практически неизменяется и остается на базальном уровне. В этот период
происходят переработка и представление антигена
иммунокомпетентным клеткам .
• Ввиду того что клетки делятся дихотомически (т.е. надвое),
прирост их количества происходит в логарифмической
зависимости и поэтому после первых циклов деления оно
изменяется незначительно. Параллельно происходят
дифференцировка пре-В-лимфоцитов в зрелые формы и
плазматические клетки.
23.
• Во время логарифмической фазы наблюдается интенсивный приростколичества антигенспецифических В-лимфоцитов, что находит
отражение в существенном нарастании титров специфических антител.
• В стационарной фазе количество специфических антител и
синтезирующих их клеток достигает максимума и стабилизируется.
• Освобождение макроорганизма от антигена устраняет антигенный
стимул, поэтому в фазе снижения наблюдается постепенное уменьшение
титров соответствующих антител.
24.
• Повторный контакт иммунной системы с тем же антигеном ведет кформированию вторичного иммунного ответа.
• Его латентная фаза значительно укорочена, а логарифмическая фаза
отличается более интенсивной динамикой прироста и более высокими
титрами специфических антител.
• Для стационарной фазы и фазы снижения свойственна затяжная динамика
(несколько месяцев или даже лет). При вторичном иммунном ответе
организм сразу же в подавляющем большинстве синтезирует IgG.
25.
• Это обусловлено подготовленностью иммуннойсистемы к повторной встрече с антигеном за счет
формирования иммунологической памяти :
многочисленные клоны антигенспецифических Влимфоцитов, оставшиеся после первичного
иммунного реагирования, быстро размножаются и
интенсивно включаются в процесс антителогенеза.
26.
• Для развития гуморального иммунитета слизистых оболочекхарактерны те же процессы и динамика антителообразования.
Однако в данном случае в слизистых оболочках в подавляющем
большинстве созревают и размножаются В-лимфоциты,
продуцирующие полимерные молекулы IgA.
27.
• Явление интенсивного антителообразования приповторном контакте с антигеном широко используется в
практических целях, например вакцинопрофилактике.
Для создания и поддержания иммунитета на высоком
защитном уровне схемы вакцинации предусматривают
многократное введение антигена для формирования и
поддержания иммунологической памяти.
• Этот же феномен используют при получении
высокоактивных лечебных и диагностических иммунных
сывороток (гипериммунных). Для этого животным или
донорам производят многократные введения препаратов
антигена по специальной схеме.
28.
• Динамика и интенсивностьантителообразования в значительной степени
зависят от иммуногенности антигена:
• дозы, способа и кратности его введения, а также
от состояния макроорганизма.
• Попытка повторного введения антигена в
латентной фазе может привести к
иммунологическому параличу - иммунологической
неотвечаемости на антиген в течение
определенного периода времени.
29.
• Роль антител в формировании иммунитета.• Антитела имеют важное значение в формировании
приобретенного постинфекционного и поствакцинального
иммунитета.
• 1. Связываясь с токсинами, антитела нейтрализуют их,
обеспечивая антитоксический иммунитет.
• 2. Блокируя рецепторы вирусов, антитела препятствуют
адсорбции вирусов на клетках, участвуют в противовирусном
иммунитете.
• 3. Комплекс антиген- антитело запускает классический путь
активации комплемента с его эффекторными функциями (лизис
бактерий, опсонизация, воспаление, стимуляция макрофагов).
30.
4. Антитела принимают участие в опсонизации бактерий,способствуя более эффективному фагоцитозу.
5. Антитела способствуют выведению из организма (с мочой,
желчью) растворимых антигенов в виде циркулирующих
иммунных комплексов.
IgG принадлежит наибольшая роль в антитоксическом
иммунитете,
IgM- в антимикробном иммунитете (фагоцитоз корпускулярных
антигенов), особенно в отношении грамотрицательных бактерий,
IgA- в противовирусном иммунитете (нейтрализация вирусов),
IgAs- в местном иммунитете слизистых оболочек,
IgE- в реакциях гиперчувствительности немедленного типа.
31.
Иммунный фагоцитоз• Феномен иммунного фагоцитоза основан на поглощении
фагоцитами антигенов, входящих в состав иммунных
комплексов. При этом антигенами могут быть как отдельные
молекулы или их агрегаты, так и цельные клетки или их
обломки.
• Рецепторы обеспечивают узнавание и захват фагоцитом
иммунных комплексов или опсонизированных антигенов,
которые потом эндоцитируются.
• Таким образом фагоциты участвуют в элиминации (удалении)
антигенов из организма и восстановлении его гомеостаза.
32.
Опосредованный клетками киллинг• Иммунная система располагает независимым от системы
комплемента способом уничтожения чужеродных клеток.
• Эта форма иммунного реагирования осуществляется
непосредственно клетками-киллерами и получила название
«опосредованный клетками киллинг».
• Киллинг способны осуществлять активированные фагоциты, Ткиллеры, естественные киллеры и некоторые другие клетки.
Клетки-киллеры осуществляют санацию организма от
чужеродных, трансформированных или инфицированных
клеток.
33.
• Механизм клеточно-опосредованного киллинга достаточноуниверсален.
• Киллеры вырабатывают ряд веществ, которые вызывают
нарушение целостности клеточной мембраны (или стенки) или
индуцируют апоптоз. Они осуществляют свою функцию
дистантно (на расстоянии) или при непосредственном контакте.
• Мишенью для них являются раково-трансформированные,
мутантные или зараженные вирусами клетки, грибы,
простейшие, гельминты, некоторые бактерии и другие
чужеродные клетки.
34.
• Иммунологическая память.• При повторной встрече с антигеном организм формирует более активную
и быструю иммунную реакцию — вторичный иммунный ответ. Этот
феномен получил название иммунологической памяти.
• Иммунологическая память имеет высокую специфичность к конкретному
антигену,
• распространяется как на гуморальное, так и клеточное звено иммунитета
• и обусловлена В- и Т-лимфоцитами.
• Она образуется практически всегда и сохраняется годами и даже
десятилетиями.
• Благодаря ей наш организм надежно защищен от повторных антигенных
интервенций.
35.
• На сегодняшний день рассматривают два наиболеевероятных механизма формирования
иммунологической памяти.
• Один из них предполагает длительное сохранение
антигена в организме. Этому имеется множество
примеров: инкапсулированный возбудитель
туберкулеза, персистирующие(постоянные, упорные)
вирусы кори, полиомиелита, ветряной оспы и
некоторые другие патогены длительное время, иногда
всю жизнь, сохраняются в организме, поддерживая в
напряжении иммунную систему.
36.
• Другой механизм предусматривает, что в процессе развития ворганизме продуктивного иммунного ответа часть
антигенореактивных Т- или В-лимфоцитов дифференцируется
в малые покоящиеся клетки, или клетки иммунологической
памяти.
• Эти клетки отличаются высокой специфичностью к конкретной
антигенной детерминанте и большой продолжительностью
жизни (до 10 лет и более). Они активно рециркулируют в
организме, распределяясь в тканях и органах, но постоянно
возвращаются в места своего происхождения . Это
обеспечивает постоянную готовность иммунной системы
реагировать на повторный контакт с антигеном по вторичному
типу.
37.
• Феномен иммунологической памяти широко используется впрактике вакцинации людей для создания напряженного
иммунитета и поддержания его длительное время на
защитном уровне. Осуществляют это 2—3-кратными
прививками при первичной вакцинации и периодическими
повторными введениями вакцинного препарата —
ревакцинациями.
• Однако феномен иммунологической памяти имеет и
отрицательные стороны. Например, повторная попытка
трансплантировать уже однажды отторгнутую ткань
вызывает быструю и бурную реакцию — криз отторжения.
38.
• Иммунологическая толерантность — явление,противоположное иммунному ответу и
иммунологической памяти. Проявляется она
отсутствием специфического продуктивного иммунного
ответа организма на антиген в связи с неспособностью
его распознавания.
• В отличие от иммуносупрессии иммунологическая
толерантность предполагает изначальную
ареактивность иммунокомпетентных клеток к
определенному антигену.
39.
• Иммунологическую толерантность вызывают антигены, которыеполучили название толерогены. Ими могут быть практически все
вещества, однако наибольшей толерогенностью обладают
полисахариды.
• Иммунологическая толерантность бывает врожденной и
приобретенной. Примером врожденной толерантности является
отсутствие реакции иммунной системы на свои собственные
антигены.
• Приобретенную толерантность можно создать, вводя в организм
вещества, подавляющие иммунитет (иммунодепрессанты), или же
путем введения антигена в эмбриональном периоде или в первые
дни после рождения индивидуума.
40.
• Приобретенная толерантность может быть активной ипассивной.
• Активная толерантность создается путем введения в
организм толерогена, который формирует
специфическую толерантность.
• Пассивную толерантность можно вызвать веществами,
тормозящими биосинтетическую или пролиферативную
активность иммунокомпетентных клеток
(антилимфоцитарная сыворотка, цитостатики и пр.).
41.
• Иммунологическая толерантность отличаетсяспецифичностью — она направлена к строго
определенным антигенам. По степени распространенности различают поливалентную и
расщепленную толерантность.
• Поливалентная толерантность возникает
одновременно на все антигенные детерминанты,
входящие в состав конкретного антигена.
Для расщепленной, или моновалентной,
толерантности характерна избирательная
невосприимчивость каких-то отдельных антигенных
детерминант.
42.
• Степень проявления иммунологическойтолерантности существенно зависит от ряда свойств
макроорганизма и толерогена.
• Важное значение в индукции иммунологической толерантности
имеют доза антигена и продолжительность его воздействия.
• Различают высокодозовую и низкодозовую толерантность.
• Высокодозовую толерантность вызывают введением больших
количеств высококонцентрированного антигена.
• Низкодозовая толерантность, наоборот, вызывается очень
малым количеством высокогомогенного молекулярного
антигена.
43.
• Механизмы толерантности многообразны и до концане расшифрованы. Известно, что ее основу составляют
нормальные процессы регуляции иммунной системы.
Выделяют три наиболее вероятные причины развития
иммунологической толерантности:
• 1. Элиминация из организма антигенспецифических
клонов лимфоцитов.
• 2. Блокада биологической активности
иммунокомпетентных клеток.
• 3. Быстрая нейтрализация антигена антителами.
44.
• Феномен иммунологической толерантности имеетбольшое практическое значение. Он используется для
решения многих важных проблем медицины, таких
как пересадка органов и тканей, подавление
аутоиммунных реакций, лечение аллергий и других
патологических состояний, связанных с агрессивным
поведением иммунной системы
45.
• Исторически иммунологическую толерантностьрассматривают как защиту против
аутоиммунных заболеваний. При нарушении
толерантности к собственным антигенам могут
развиваться аутоиммунные реакции, в том числе
возникать такие аутоиммунные заболевания как
ревматоидный артрит, системная красная волчанка и
другие.
46.
47.
48.
49.
• Основные механизмы отмены толерантности и развития аутоиммунныхреакций
• 1. Изменения химической структуры аутоантигенов (например- изменение
нормальной структуры антигенов клеточных мембран при вирусных
инфекциях, появление ожоговых антигенов).
• 2. Отмена толерантности на перекрестно- реагирующие антигены
микроорганизмов и эпитопы аутоантигена.
• 3. Появление новых антигенных детерминант в результате связывания
чужеродных антигенных детерминант с клетками хозяина.
• 4. Нарушение гисто- гематических барьеров.
• 5. Действие суперантигенов.
• 6. Нарушения регуляции иммунной системы ( уменьшение количества или
функциональная недостаточность супрессирующих лимфоцитов, экспрессия
молекул МНС класса 2 на клетках, в норме их не экспрессирующих- тиреоциты
при аутоиммунном тиреоидите).
50.
• Противобактериальный иммунитет основан насочетанном воздействии на бактерии и
их токсины факторов неспецифической
резистентности (лизоцима, комплемента, β-лизинов,
фагоцитов и др.) и факторов специфического
иммунного ответа. Многие белки (опсонины) крови
вместе с антителами откладываются на поверхности
бактерий, блокируя их антигены и способствуя
иммунному прилипанию к фагоцитам — начальной
стадии фагоцитоза, а в последующем и
перевариванию бактерий в фагоцитах.
51.
52.
• Противобактериальные антитела, в частностиантитела к адгезинам бактерий, препятствуют
прикреплению бактерий к тканям организма и,
следовательно, развитию начальной стадии
инфекционного процесса.
53.
• Антитела против токсинов бактерий участвуют в такназываемом антитоксическом иммунитете.
Антитоксические антитела, не влияя на колонизацию
(заселение) бактериями слизистых оболочек,
препятствуют развитию патологического процесса.
• Лизоцим, обладая ферментативной активностью,
разрушает пептидогликан клеточной стенки бактерий и
приводит к их лизису. К лизоциму наиболее
чувствительны грамположительиые бактерии, клеточные
стенки которых в основном состоят из пептидогликана.
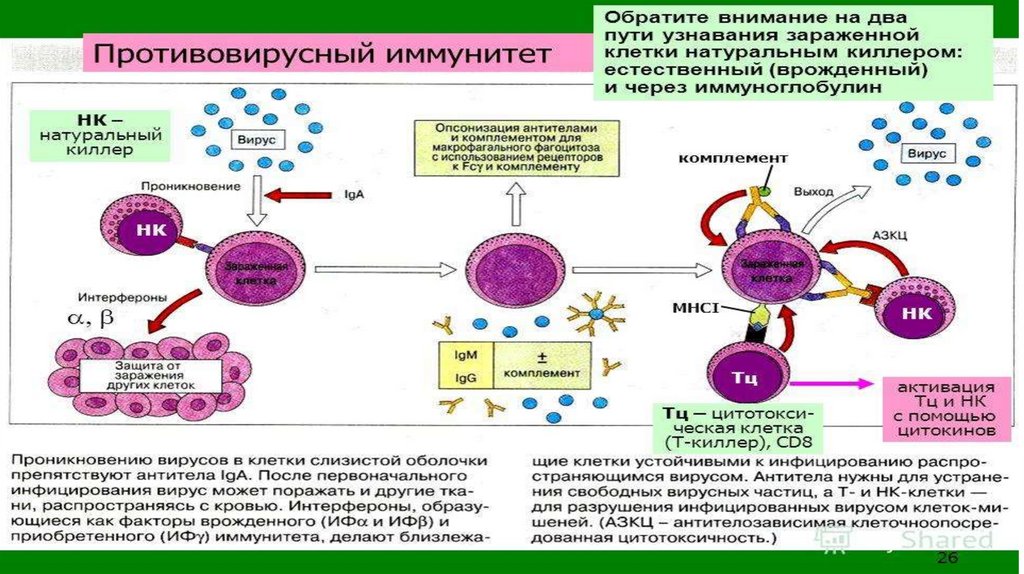
54.
• Противовирусный иммунитет. Отличиепротивовирусного И. от других видов И. (против
бактерий, простейших, грибков и т.д.) связано со
своеобразием структуры и размножения вирусов,
особенностями патогенеза вирусных инфекций.
• Видовой противовирусный И. обусловлен
отсутствием у клеток данного вида организмов
рецепторов для прикрепления (адсорбции)
соответствующих вирусов или их неспособностью
репродуцироваться после проникновения в клетку.
55.
56.
57.
• Немаловажную роль в защите от вирусов играетвоспалительная реакция, направленная на ограничение
распространения вирусов в организме и фиксацию их в
воротах инфекции. При этом помимо клеток крови
(макрофагов, естественных киллеров) противовирусный
эффект оказывают такие универсальные реакции на
внедрение вирусов, как общее или локальное повышение
температуры и увеличение кислотности среды.
• Приобретенный противовирусный И. формируется в
результате перенесенного заболевания или иммунизации
организма с помощью вакцин.
58.
59.
• Иммунитет при паразитарныхболезнях. Различают частичный видовой и
абсолютный (т.е. полную невосприимчивость
человека к данному виду паразита) иммунитет.
например, у населения влажных тропиков Африки,
в Южной Азии сформировался ряд аномальных
гемоглобинов, эритроцитарных ферментопатий,
что привело к частичному И. при тропической
малярии.
60.
• Населению определенных зон тропической Африкисвойственна полная невосприимчивость к возбудителю
трехдневной малярииPlasmodium vivax вследствие
утраты эритроцитами антигенов группы Duffy,
необходимых для проникновения плазмодия в клетку.
• Молекулярные факторы, обеспечивающие видовой И.
при паразитарных болезнях, активно изучаются, но во
многом неизвестны
61.
• Приобретенный И. при паразитарныхзаболеваниях обеспечивается теми же типами
иммунного ответа, которые характерны для
инфекций. Защитное значение имеют антитела и
клеточные факторы. Приобретенный И. может
привести к полному освобождению от возбудителя
(стерильный И.) или оказывать частичный эффект.
Последний выражается в изгнании части
популяции паразитов, обычно гельминтов,
ограничении их репродуктивной способности,
нарушении цикла развития и др.
62.
• Нередко И. выражен только при наличии паразитав организме хозяина (нестерильный И.,
премуниция). Однако даже стерильный И. не
всегда предотвращает повторное заражение.
• В целом гомеостаз организма обеспечивается
согласованной работой (взаимодействием)
иммунной, эндокринной и нервной систем.