



















































































")
 Биология
БиологияПохожие презентации:










Темы семинарских занятий по курсу Молекулярная биология
1.
1.Темы семинарских занятий по курсу Молекулярная биологияОсновы молекулярной биологии.
Вопросы к зачету по Основам молекулярной биологии.
1 Семинар. Как и почему появились в клетках нуклеиновые кислоты
1.Локализация и функция нуклеиновых кислот.
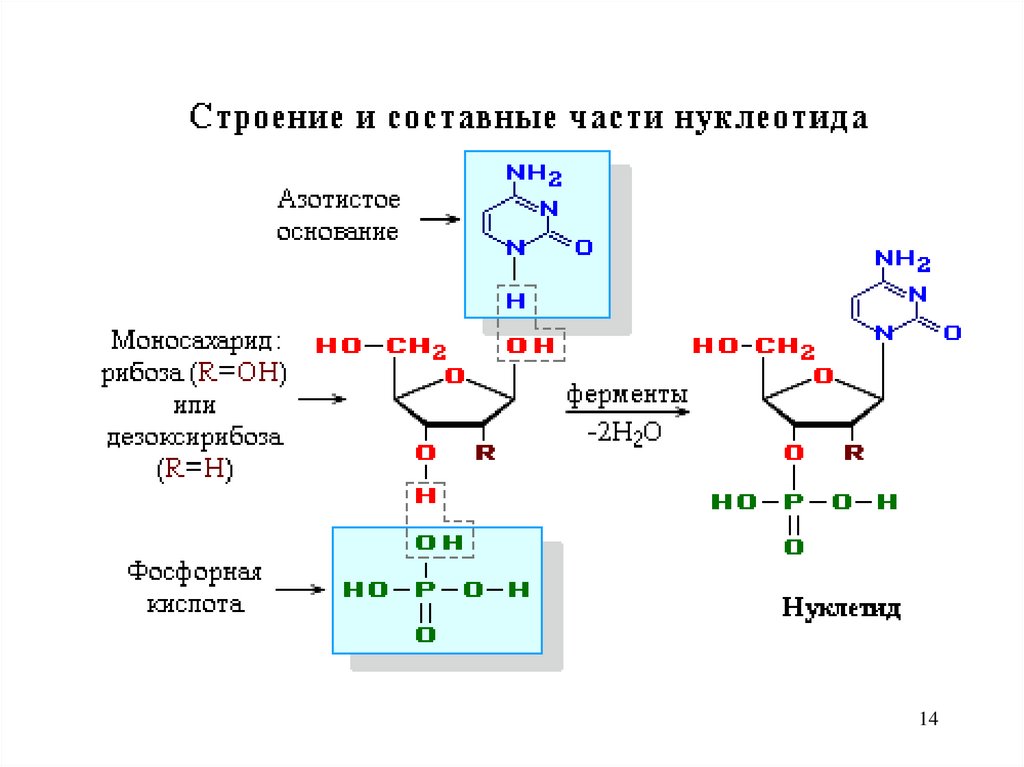
2.Строение и составные части нуклеотида.
3.Основания, нуклеозиды, нуклеотиды.
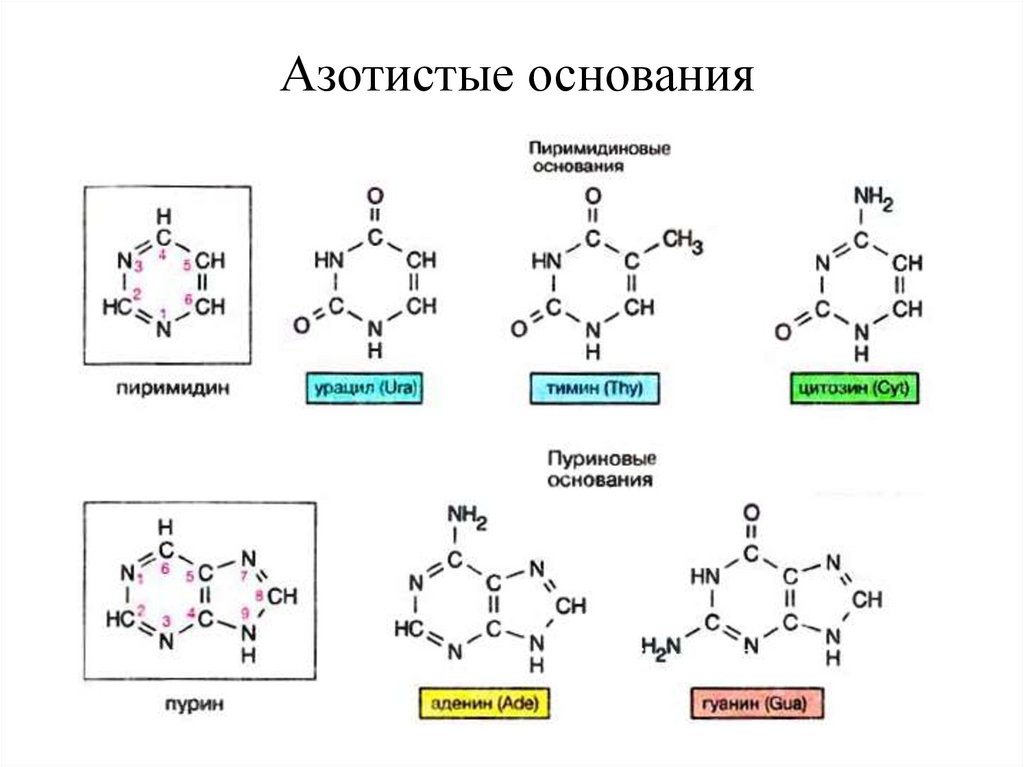
4.Азотистые основания
5.Химическая характеристика нуклеозидов и нуклеотидов
6. Таутомерия оснований: лактамная и лактимная форма.
7.Структура двойной спирали ДНК. Предподсылки к созданию модели ДНК
Уотсона и Крика. 1. Правила Чаргоффа. 2.Данные рентгеноструктурного
анализа Розалин Франклин и М. Уилкинса
2.
8.Биологическое значение двуспирального строения ДНК.9.Почему ДНК двойная спираль, а не лестница,
10.В чем причина плоской структуры и комплементарности пар оснований и размера в
3.4 ангстрема между ними?
11.Почему структура моносахаридных остатков не плоская, как у оснований, а имеет
цис- гош конформацию в цепи ДНК?
12.Почему диаметр спирали ДНК около18-20 ангстрем?
13.Почему цепь ДНК не меняет свой диаметр по всей длине цепи
14.Почему в ДНК комплементарны пурин-пиримидиновые основания, а не пуринпурин или пиримидин-пиримидин?
15. Какие формы двойной спирали ДНК вы знаете и в чем причина таких изменений?
16.. Почему ДНК двойная спираль , а РНК одинарная спираль.?
3.
17. Расшифровка первичной структуры ДНК18. Организация генома
19. 4 типа структурно-функциональной организации генов в геноме.
20. Две важные особенности в организации эукариотических генов
21. Интроны в генах рРНК и тРНК-
22. % состав информативной или ген-содержащей части ДНК
23. Физико-химические свойства ДНК
24. Третичная структура ДНК бактерий, вирусов и эукариот
25. 5 уровней организации структурыа ДНК
26. Структура нуклеосом
27. Цитоплазматическая, митохондриальная и хлоропластная ДНК
28. Бактериальные плазмиды. Мигрирующие элементы плазмид и хромосом
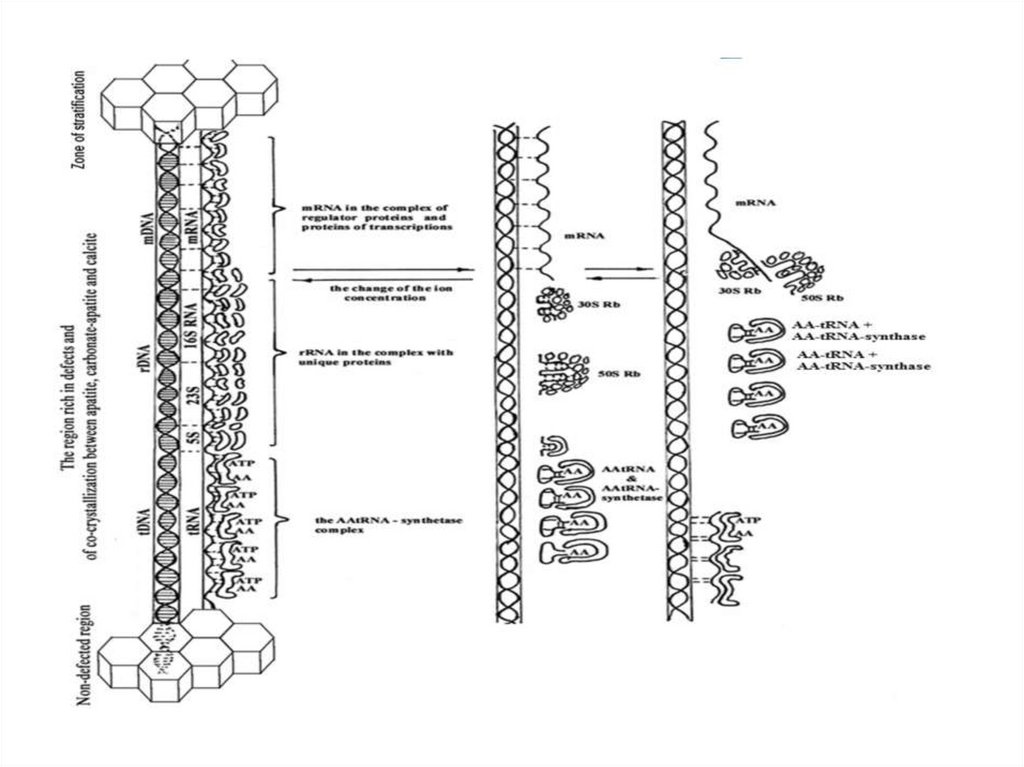
29. РНК и место их формирования на апатите и карбонат-апатите
30. Структура рРНК (80%),
31. Структура мРНК (5%),
32. Структура тРНК (15%),
4.
5.
• Mолекулярная биология - это наука о структуре ифункциях
нерегулярных
биополимеров
нуклеиновых кислот и белков, механизмах
хранения, воспроизведения, передачи и реализации
генетической информации.
6.
История Н.К.1868 гной, выделил из ядер лейкоцитов – нуклеины, в состав которых входит фосфор,
нуклеиновую кислоту и белок
1880 в хромосомах найдена НК
1928 Гриффитс Убитый штамм Diplococcus pneumonia + живой (не патогенный) заразили
мышей (см.сл)
1943 Эвери определил природу трнсформирующего агента – ДНК убитых клеток
D.pneucoccus (см.сл)
•1952 Херши и Чейз Фаг Т2 запускал в E.coli свою РНК, но не белок (см.сл.)
•1949 Рис и Мирский ДНК в клетках константна
•1912 отец и сын Брэгги изобрели метод рентгеновской кристаллографии основанный на
том, что пучок параллельных рентгеновских лучей, падающих на регулярное скопление
атомов, образует так называемую дифракционную картину, которая зависит от атомной
массы атомов и их пространственного расположения.
•1940 г.г. Астбюри использовал данный метод для определения пространственной
структуры ДНК. И нашел, что ДНК представляет собой стопку из уложенных один над
други1953. Эрвин Чаргафф.м нуклеотидов. В виде плоских дисков.
•С помощью кислотного гидролиза ДНК с последующей хроматографией и количественным
анализом установлены закономерности: А/Т=1; Г/Ц=1; (Г+Ц)/(А+Т)=К - коэффициент
специфичности, постоянен для каждого вида.
7.
1. 1928г. Опыты Фредерика Гриффита.Гриффит работал с пневмококками - бактериями,
вызывающими пневмонию. Он брал два штамма
пневмококков: капсульный и бескапсульный.
Капсульный - патогенный (вирулентный), при
инфицировании таким штаммом мыши погибают,
бескапсульный - непатогенный. При введении мышам
смеси убитых нагреванием (и, следовательно,
потерявших вирулентность) капсульных
пневмококков и живых бескапсульных
невирулентных бактерий, животные погибали в
результате размножения капсульных вирулентных
форм. Обнаруженное явление Гриффит
интерпретировал как трансформацию. Определение:
Трансформация - это приобретение одним
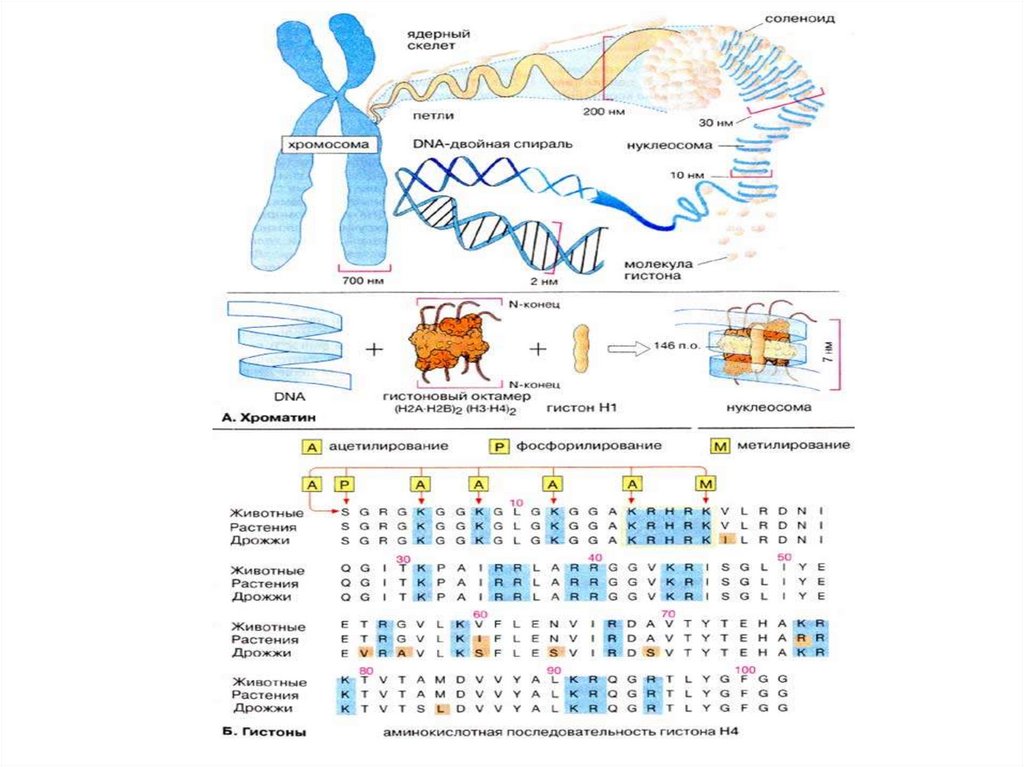
организмом некоторых признаков другого
организма за счет захвата части его
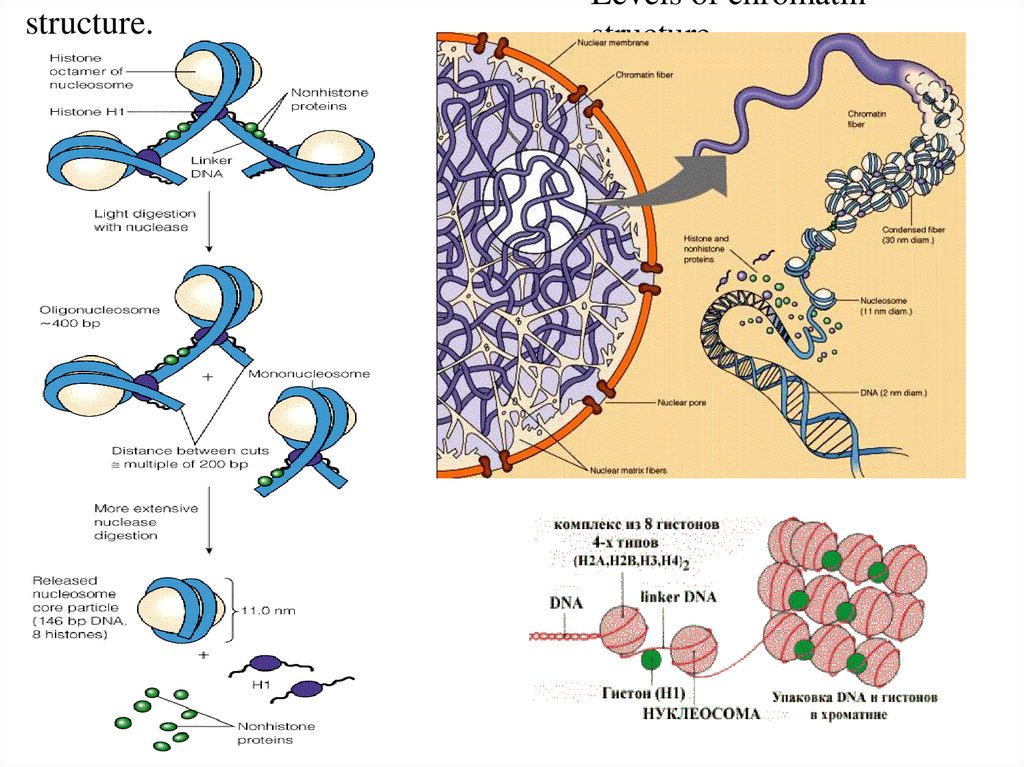
генетической информации.
В 1944г. этот эксперимент был повторен Освальдом
Эйвери, Колином Мак-Леодом и Маклином МакКарти в варианте смешивания бескапсульных
пневмококков с взятыми от капсульных белками,
полисахаридами или ДНК. В результате этого
эксперимента была выявлена природа
трансформирующего фактора.
Трансформирующим фактором оказалась ДНК.
8.
2. 1952г. Эксперимент Альфреда Херши иМарты Чейз. Фаги (бактериофаги) - это
вирусы, размножающиеся в бактериях. E. сoli
- кишечная палочка (эубактерия).
• Суть опыта: фаги, у которых белковая
оболочка была мечена радиоактивной серой
(S35), а ДНК - радиоактивным фосфором
(Р32), инкубировали с бактериями. Затем
бактерии отмывали. В смывных водах не
обнаруживали Р32, а в бактериях - S35
Следовательно, внутрь попала только
ДНК. Через несколько минут из бактерии
выходили десятки полноценных фагов,
содержащих и белковую оболочку, и
ДНК.Отсюда следовал однозначный вывод о
том, что именно ДНК выполняет
генетическую функцию - несет
информацию как о создании новых копий
ДНК, так и о синтезе фаговых белков.
9.
3. 1957г. Опыты Френкеля - Конрата• Френкель-Конрат работал с вирусом табачной
мозаики (ВТМ). В этом вирусе содержится РНК,
а не ДНК. Было известно, что разные штаммы
вируса вызывают разную картину поражения
листьев табака. После смены белковой оболочки
"переодетые" вирусы вызывали картину
поражения, характерную для того штамма, чья
РНК была покрыта чужим белком.
Следовательно, не только ДНК, но и РНК
может служить носителем
генетической информации.
• На сегодняшний день существуют сотни
тысяч доказательств генетической роли
нуклеиновых кислот. Приведенные три
являются классическими
10.
•В 50 годы исследования продолжили 3группы:1 я Лайус Полинг
2я физик Морис Уилкинс
3я Джеймс Уотсон и Френсис Крик.
Первую модель ДНК представил Полинг в1953 г – она не получила признания.
Уилкинс получил очень четкие рентгенограммы ДНК, на которых отчетливо было видно,
что молекула НК состоит из 2х нитей и была подтверждена гипотеза Астбюри о
межнуклеотидном расстоянии равном 0.34нм. Уотсон и Крик обратили внимание на эту
публикацию . Они дополнили свои предположения правилами Эрвина Чаргаффа и в апреле
1953 г опубликовали свою модель ДНК.
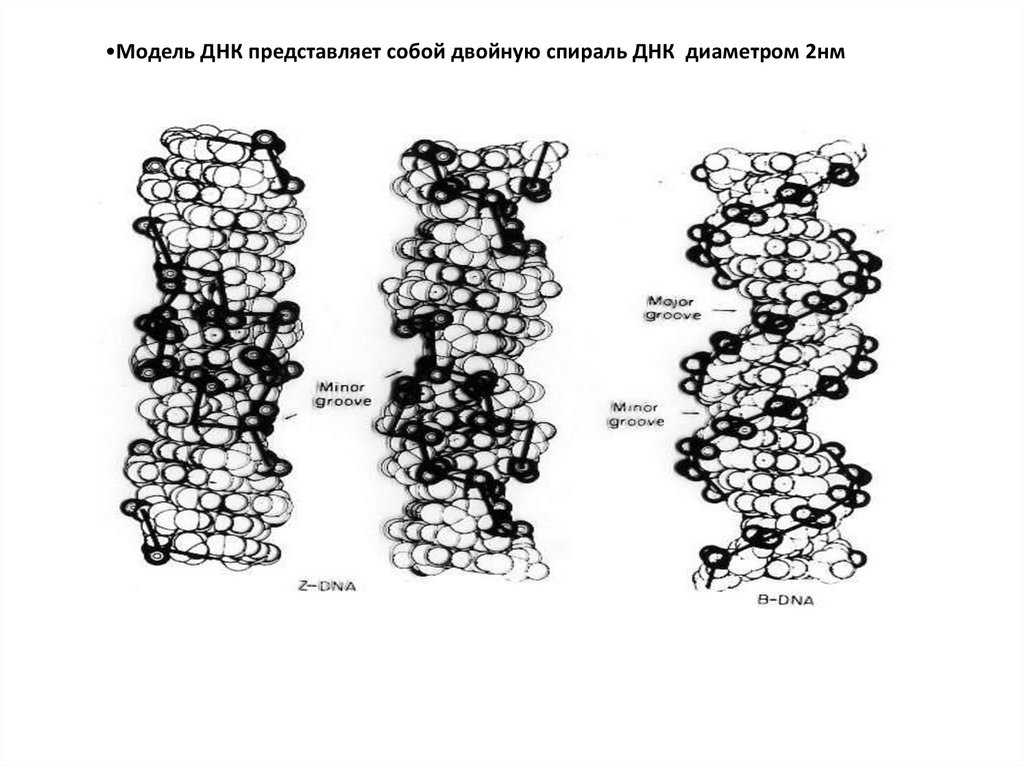
•Модель ДНК представляет собой двойную спираль ДНК ? диаметром 2нм
•Вдоль оси соседние пары оснований распологаются на расстоянии 0.34нм ? одна от другой
•Полный оборот спирали приходиться на 3.4нм т.е. на 10 пар оснований
Последовательность нуклеотидов в цепи отражает генетическую информацию ? и должна
соответствовать ее молекулярной структуре
Чтобы спираль имела одинаковый диаметр вдоль цепи они предположили в парах
оснований правило комплементарности пурин-пиримидиновых оснований. т.е. А=Т и Г=Ц
? из правил Чаргаффа
11.
Основное значение правила комплементарности состоит в признании ДНКсамокомплементарной молекулой. Обе цепи антипараллельны.
1956г Значительный вклад в понимание механизмов самоудвоения внес
Артур Корнберг, который открыл ДНК-полимеразы.
1969 –синтезирован ген тРНК
Работы выше названных ученых заложили фундамент в понимание
процессов
наследственности,
наследственной информации.
структуры
и
роли
ДНК
в
передаче
12.
В70-80
годы
исследования
продолжила
группа
профессора
Э.Я.Костецкого, которая предложила новую теорию происхождения жизни
на комплексе минеральных кристаллов. Обосновала и предложила ответ
на
вопрос,
как
комплементарными
и
почему
возникла
двойная
пурин-пиримидиновыми
спираль
парами
ДНК
с
оснований
и
едиными параметрами ДНК во всех таксономических группах, почему
возникли тРНК, рРНК, мРНК, как и почему возникла рибосома в готовом
виде со своими белками, как возник матричный механизм, универсальный
генетический код с вырожденностью по третьему нуклеотиду, как
возникла структуная ассиметрия молекул, как возникла живая система
без нарушения второго закона термодинамики, была сформулирована
новая
идея
об
истинном
носителе
генетической
информации
-
суперспецифической АА-тРНК-синтетазе и поставлена под сомнение
ключевая наследственная роль ДНК, опровергнута природа и другие
вопросы молекулярной биологии.
13.
II. Функция НКХранение и передача информации
Энергетическая АТФ, ГТФ, ЦТФ, УТФ
Кофакторы ферментов НАД, НАДФ, ФАД
Регуляция метаболизма ЦДФ (синтез ФЛ – ЦДФ ДГ, ЦДФ-холин, ЦДФ –
этаноламин, ЦДФ-инозитол), УДФ (синтез гликолипидов –УДФ Гл, УДФ-Гал),
АТФ-АДФ, ГТФ- синтез белка, цАМФ, перенос сигналов в межнейрональном
пространстве
III - Структура НК
•Модель ДНК представляет собой двойную спираль ДНК диаметром 2нм
Основания - лактамная формма - C-N=H , -C=O,
лактимная форма - C –NH2, -C-OH
нуклеотиды, нуклеозиды
14.
.14
15.
Азотистые основания16.
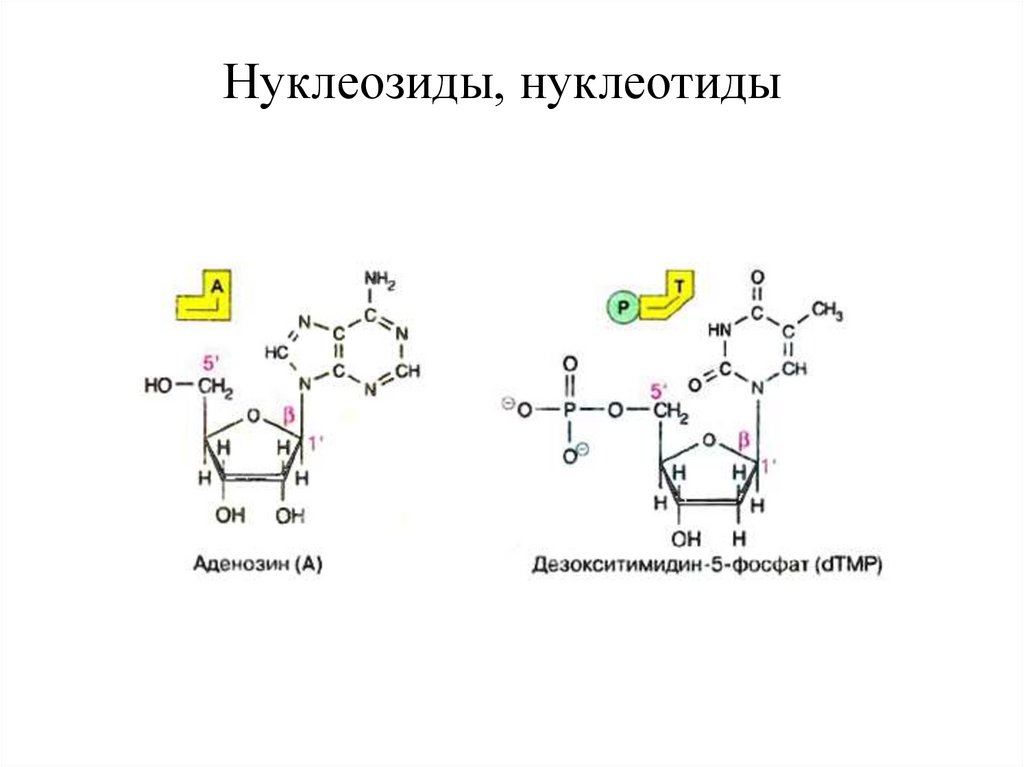
Нуклеозиды, нуклеотиды17. НУКЛЕОЗИДЫ
•Нуклеозиды - это N-гликозиды пиримидиновых или пуриновыхоснований, в которых 1-й углеродный атом сахара (пентозы) связан
гликозидной связью с атомом N1 пиримидина или атомом N9 пурина.
Пентозы всегда присутствуют в фуранозной форме.
Существует два ряда нуклеозидов: рибонуклеозиды, которые содержат в
качестве сахарного компонента D-рибозу, и 2'-дезоксирибонуклеотиды,
содержащие 2-дезокси-D-рибозу.
•Гликозидная связь. В нуклеозидах и нуклеотидах N-гликозидная связь
- это связь между 1-м углеродным атомом сахара - пентозы и атомом N1
пиримидина или N9 пурина. N-гликозидная связь имеет бетаконфигурацию.
•Для пиримидиновых нуклеозидов длина связи С1'-N максимальна и
составляет 1.52 А при торсионном угле вращения, определяемом через
атомы - (О4'- С1'- N1 - С2), и равным примерно 180 град. По мере его
приближения к -140 град. длина N-гликозидной связи уменьшается до 1.48
А практически линейно.
•В пуриновых нуклеозидах длина гликозидной связи менее вариабельна и
составляет примерно 1.46 А.
17
18. НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
Нуклеотиды представляют собой фосфорнокислые эфирынуклеозидов, в которых фосфорная кислота связана
сложноэфирной связью с одной из свободных гидроксильных
групп сахарного кольца.
•Фосфорилирование дезоксирибонуклеозида может
осуществляться по положениям С3' и С5', так как атомы С1'
и С4' включены в фуранозное кольцо, а атом С2' не имеет
гидроксильной группы. В РНК только положения 1'и
4'недоступны для этерификации, зaмещение по С2', С3' и С5'
возможно.
•Нуклеотиды являются сильными кислотами. Они
называются, адениловой, гуаниловой, тимидиловой,
цитидиловой и уридиловой кислотами.
18
19.
Основания - лактамная формма - C-N=H , -C=O, лактимная форма - C –NH2, -C-OH• Tаутомерия оснований
В гетороциклах протоны, связанные с азотом, могут переходить
на другие атомы азота или на атомы кислорода кетогруппы.
Поэтому в растворах будет существовать равновесие различных
таутомерных структур, быстро переходящих друг в друга.
В результате таутомерых превращений U и G в енольной форме при
спаривании могут имитировать С и А, а С и А в иминоформе - U и G,
что может привести к мутациям в ДНК.
• В растворе обычные кето- и аминоформы всегда превалируют
над енольной и имино-формами, доля которых, как правило, не
превышает 0,1 %. Эта величина может сильно меняться для
модифицированных оснований или для оснований в составе
комплементарной пары.
19
20. Олигонуклеотиды, полинуклеотиды
21. Нуклеиновые кислоты являются нерегулярными полимерами, мономеры которых - нуклеотиды
• Нуклеотид = нуклеозид +фосфорная кислота =
азотистое основание+
пентоза+фосфорная
кислота.
• В РНК пентоза - рибоза.
• В ДНК - дезоксирибоза.
21
22.
23. Спаривание оснований в ДНК
24.
25.
Вторичная структура ДНКМодели предшествовали
1Правила Чаргоффа (установлены закономерности: А/Т=1; Г/Ц=1; (Г+Ц)/(А+Т)=К коэффициент специфичности, постоянен для каждого вида.
2.Данные рентгеноструктурного анализа Розалин Франклин и М. Уилкинса
Уилкинс получил очень четкие рентгенограммы ДНК, на которых отчетливо было видно, что
молекула НК состоит из 2х нитей и была подтверждена гипотеза Астбюри о
межнуклеотидном расстоянии равном 0.34нм.
3. 1953 модель Уотсона и Крика . Они дополнили свои предположения словом спираль и в
апреле 1953 г опубликовали свою модель ДНК.
Модель ДНК представляет собой двойную спираль ДНК диаметром 2нм. Две цепи
обвивают друг-друга, образуя правую спираль, углевод-фосфатные цепи снаружи, а
основания внутри и расположены стопкой, обуславливая стекинг взаимодействия, причем
плоскости оснований параллельны друг другу и перпендикулярны длинной оси спирали.
Они не доступны для воды. Каждое основание одной цепи комплементарно спарено с
основаниями другой цепи водородными связями (пур=пир. Они близки даже по
расстоянию гликозидных центров (11А) Разрешенными являются А=Т, Г=Ц пары. Хотя в
принципе , спариваться могут любые основания, но без порядка в цепи.
26.
Вдоль оси соседние пары оснований распологаются на расстоянии 0.34нм одна от другой.Полный оборот спирали приходиться на 3.4нм т.е. на 10 пар оснований
Последовательность нуклеотидов в цепи отражает генетическую информацию и должна соответствовать ее
молекулярной структуре. Двойная спираль обеспечивает максимальную стабильность за счет : 1. Н- связей, 2.
Гидрофобного взаимодействия между основаниями.
На виток приходится 10 нук.пар (34А), шаг 3.4А, 36 0
Формы ДНК: В форма in vivo. Есть А, С, Z. Z форма появляется в учас тке ДНК с чередованием Г-Ц пар в
кросинговере. У нее на виток 12 пар, поворот 300, зигзаг.
Чтобы спираль имела одинаковый диаметр вдоль цени они предположили в парах оснований
комплементарность пурин-пиримидиновых оснований. т.е. А=Т и Г=Ц
Правило комплементарности взаимосвязано и подтверждается правилами Чаргоффа о эквивалентности.
Основное значение правила комплементарности состоит в признании ДНК самокомплементарной
молекулой. Обе цепи антипараллельны.
1я структура обращенные повторы,
Si, Sr, Mg, Ca
Правило Чаргоффа ∑пур=∑пир, А=Т, Г=Ц, А/Т=1, Г/Ц=1 А+Ц=Г+Т
Соотношение нуклеотидов из разных источников различно
27.
•Модель ДНК представляет собой двойную спираль ДНК диаметром 2нм28.
При исследовании ДНК различными методами обнаружено наличие различных
форм ДНК образуемые при различных условиях (концентрации солей,
влажность), некоторые из которых способны существовать в живых
организмах.
Существуют A, B, C, D, T-семейства форм ДНК, которые могут быть
подразделены на различные подтипы (C', C'').
В-ДНК - основное состояние ДНК показанное на кристаллах и в водных растворах.
С-ДНК - форма существующая при пониженной концентрации Na и влажности 44-
66%, если GC=31-72%.
А-ДНК - такая форма образуется у гибридов ДНК-РНК, следовательно при
транскрипции ДНК переходит в А-форму, в месте контакта RNA-pol. Для этой формы
характерно наличие внутренней пустоты диаметром 5Å.
Z-ДНК - левозакрученная форма Переходу B-->Z способстует наличие GC-5'
последовательности являющейся местом метилирования у организмов. Такие
последовательности в плазмидах при сверхспирализации переходят из B в Z форму.
При B-->Z переходе участок в 11 пн имеет переходную форму между левой и правой
спиралью.
Z-ДНК обнаружена в междисках политенных хромосом D. melanogaster.
Полинуклеотид GC-5' находясь в В-форме при низкой концентрации соли образует
нуклеосомы. При высоких концентрациях соли полинуклеотид GC-5' переходит в Zформу которая не образует нуклеосом.
А-, Z- формы не могут существовать в водном растворе без дополнительных
воздействий (белки, суперспирализация).
D-DNA - AT-богатые участки ДНК фага Т2, у которого цитозин заменен на 5'гидроксиметилцитозин, единственная из известных природных ДНК находится в Dформе. Кроме того ДНК фага гликозилирована более чем на 70%.
Двойная спираль D-ДНК закручена сильнее чем B-ДНК и имеет глубокий малый желоб
28
- удобнуюю полость для размещения воды и катионов.
29. Формы двойной спирали ДНК
•Существуют несколько форм двойнойспирали ДНК.
•В основной - В-форме (паракристаллическая,
Н2О>40%) на виток приходится 10
комплементарных пар. Плоскости азотистых
оснований перпендикулярны оси спирали.
Соседние комплементарные пары повернуты
друг относительно друга на 360. Диаметр
спирали 20Å, причем пуриновый нуклеотид
занимает 12Å, а пиримидиновый - 8Å.
•А-форма (кристаллическая, Н2О<40%) - 11
пар азотистых оснований на виток. Плоскости
азотистых оснований отклонены от нормали к
оси спирали на 200. Отсюда следует наличие
внутренней пустоты диаметром 5Å. Высота
витка 28Å. Такие же параметры у гибрида из
одной цепи ДНК и одной цепи РНК.
•С-форма - шаг спирали 31Å, 9.3 пар
оснований на виток, угол наклона к
перпендикуляру 60.
•Все три формы - правозакрученные спирали.
•Есть еще несколько форм правых спиралей и
всего одна левая спираль (Z -форма). Высота
витка в Z-форме -44.5 Å, на виток приходится
12 пар нуклеотидов. Ни А-, ни Z- формы не
могут существовать в водном растворе без
дополнительных воздействий (белки или
29
суперспирализация).
30. A, B и Z –формы ДНК
31.
32.
Пространственная модельДНК.
Fundamental elements of
structure in the DNA double
helix
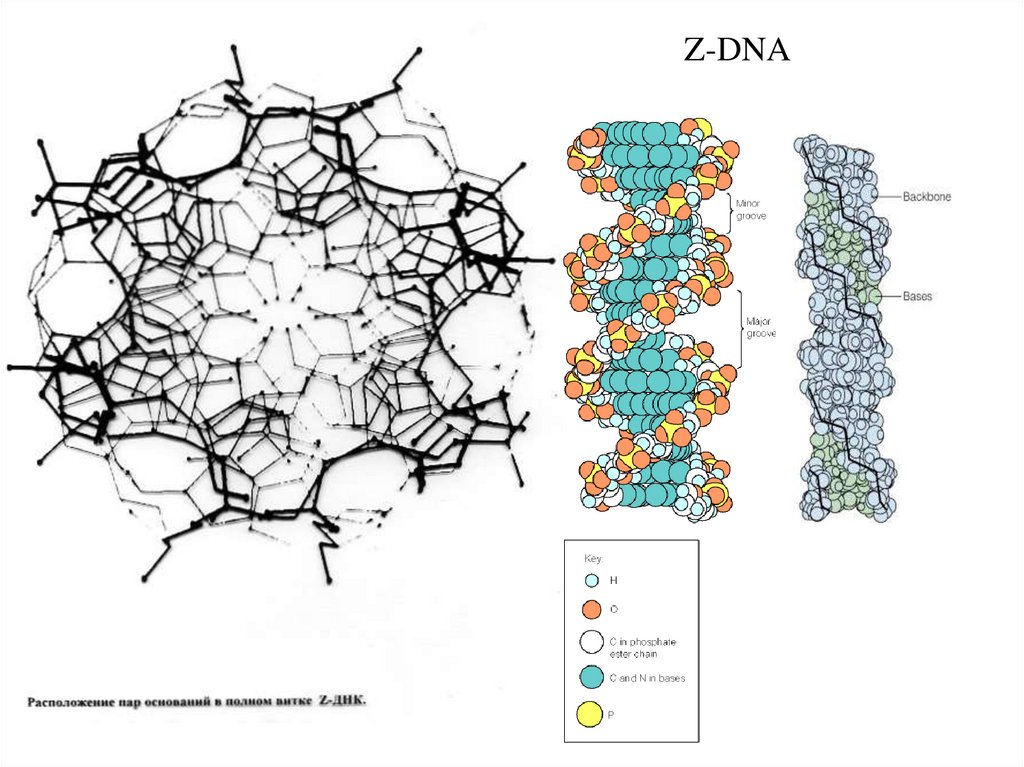
Z-DNA
33.
Вначале постараемся дать общую информацию о достоинствах апатита, а затемостановимся на его физико-химических свойствах, как предшественника будущих
протоклеток.
Наше внимание было обращено на следующие свойства апати-та, позволившие
предположить его особую роль в процессе абио-генного синтеза биомолекул. На рис. 7-9
приведены кристаллы апа-тита (рис. 9) и кристаллы апатита с сопутствующими минералами
(рис. 7-8).
ВАЖНЫЕ КЛЮЧЕВЫЕ МОМЕНТЫ по апатиту
34.
Апатит с нефелином и сфеном Кольский полуостров35.
Апатит в диопсиде Ковдор36.
Апатит кристаллы с кальцитом. Слюдянка.Сопутствующие минералы. Сфен, циркон, пироксен,
амфибол, шпинель, везувиан, флогопит, нефелин, магнезит, ква
рц, полевой шпат, слюды, кальцит.
37.
38.
39.
40.
Апатит впороде
41.
42.
ВАЖНЫЕ КЛЮЧЕВЫЕ МОМЕНТЫ по апатитуСлово апатит греческого происхождения и обозначает «обманываю», так как
апатит очень часто путали со многими минералами. Относится к группе
полудрагоценных минералов.
1. Апатит единственный из 1500 минералов - источник фосфора на поверхности
Земли, без которого
живые системы не бывают. Типичные представители:
фторапатит
хлорапатит
Ca5(PO4)3F,
Ca5(PO4)3Cl,
гидроксиапатит
Ca5(PO4)3OH,
карбонатапатит
Ca5(PO4,CO3OH)F,OH..
2. Апатит занимает особое место в огромном минеральном мире, ибо находится
на грани живой и неживой природы.
3. Проблема включения фосфора в органические соединения живых систем
(нуклеиновые кислоты, фосфорсодержащие белки и другие соединения) связана с
его пребиотической локализацией в апатите.
4. Апатит в составе позвоночника это место где формируются стволовые клетки.
43.
5. Апатит является минеральной составляющей костей и зубов позвоночныхживотных и человека, а также основой многих патологических обызвествлений.
6. Современные обитатели Земли наряду с апатитом включают в состав
минерализованных структур кальцит, арагонит и кристобалит, которые имеют с
ним систему согласованных пропорций.
7. Апатит активно используется в биохимии и молекулярной биологии для
разделения белков и нуклеиновых кислот в силу близости их структурных
особенностей. Хроматография на гидроксилапатите (ГА) показала, что адсорбция
органических веществ носит стереоспецифический характер
8. Элементарная ячейка апатита (с параметром по с = 6,879Å) и двойная спираль
ДНК имеют сходную периодичность в 3,4 А°.
9. Спиральное расположение фосфатных групп вдоль гексагональной оси в
элементарной ячейке апатита обеспечивает спиральность будущей ДНК.
44.
10. Кристаллохимические особенности апатита по своим параметрам допускаютодновременный синтез на своей матрице цепей ДНК в комплексе с основными
белками (колаген, протамин и др.) или их раздельный синтез.
11. Для живых систем апатит может быть источником РО4, Са, Si, Sr, Mg, Ge, Fe,
Mn, Zn, Cu, K, Na и др.
12. Апатит- это кристаллы способные к изоморфизму, без изменения, основных
параметров своей кристаллической структуры, что обеспечивает единство
метаболизма и будущее разнообразие живых систем сформированных на их основе.
Для живых систем апатит может быть источником РО4, Са, Si, Sr, Mg, Ge, Fe, Mn,
Zn, Cu, K, Na и др
13.
Для
обоснования
предложенной
теории
были
сопоставленны
данные
рентгеноструктурного анализа апатита с одной стороны [54,88],цепей ДНК Zформы [90–91], цепей коллагена, полиглицина II и полипролина II [76–79] – с
другой;построена и проанализирована модель апатита с фрагментом ДНК Zформы, в том числе компьютерная.
45.
46.
ДБ
В
Г
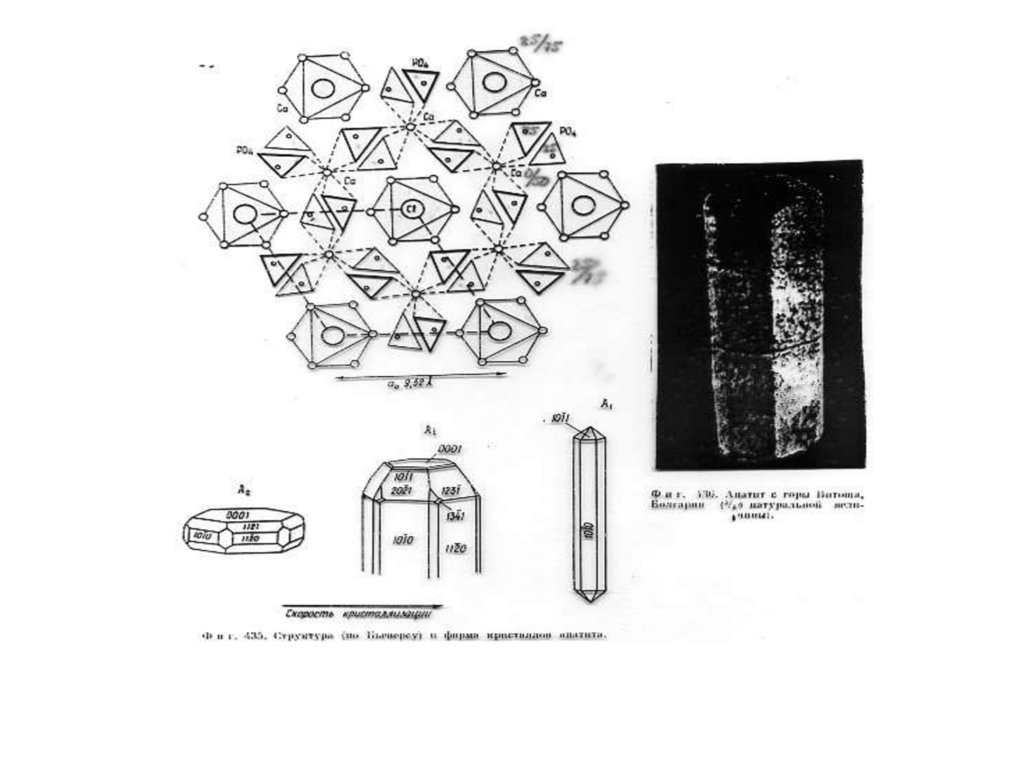
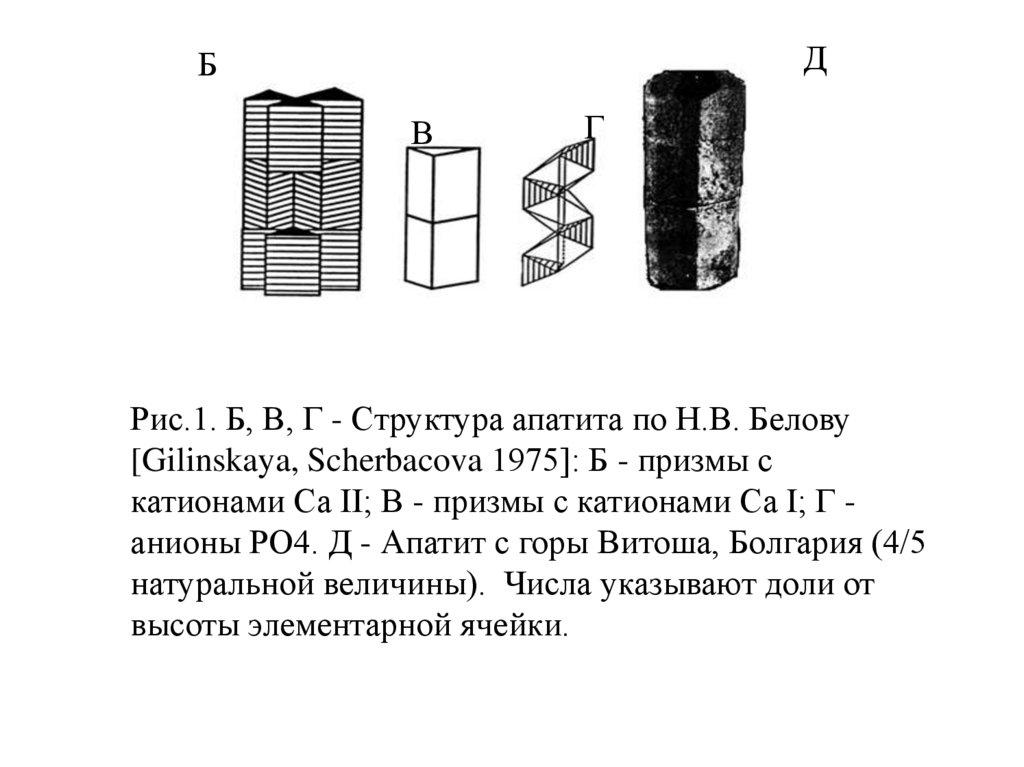
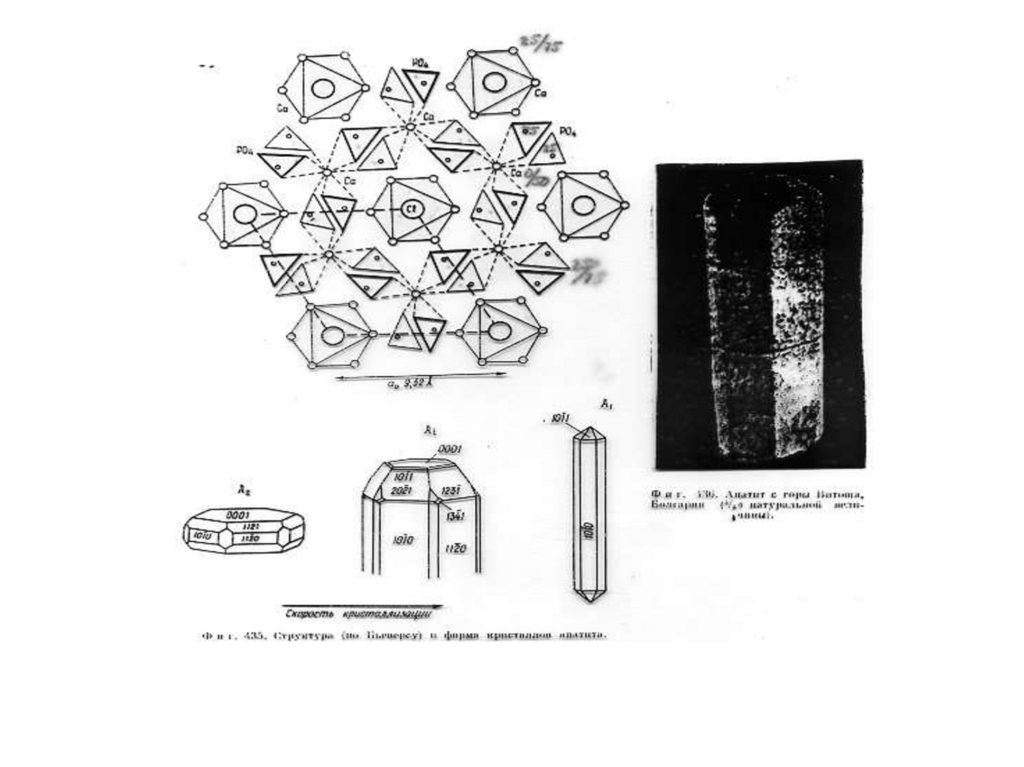
Рис.1. Б, В, Г - Cтруктура апатита по Н.В. Белову
[Gilinskaya, Scherbacova 1975]: Б - призмы с
катионами Са II; В - призмы с катионами Са I; Г анионы РО4. Д - Апатит с горы Витоша, Болгария (4/5
натуральной величины). Числа указывают доли от
высоты элементарной ячейки.
47.
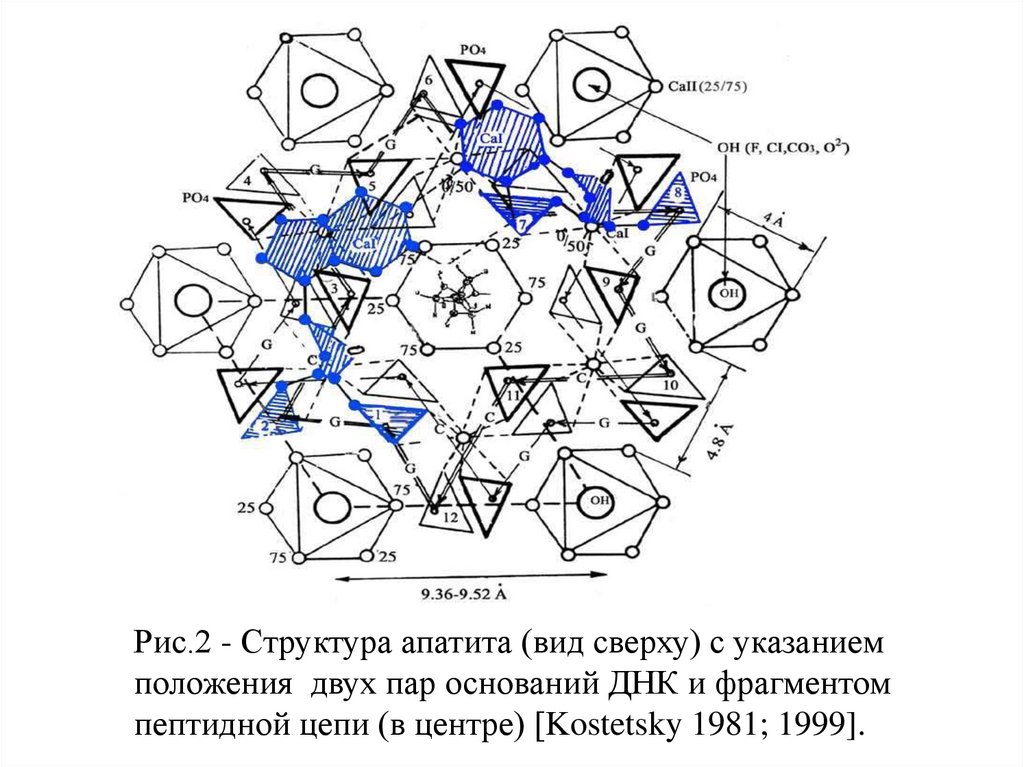
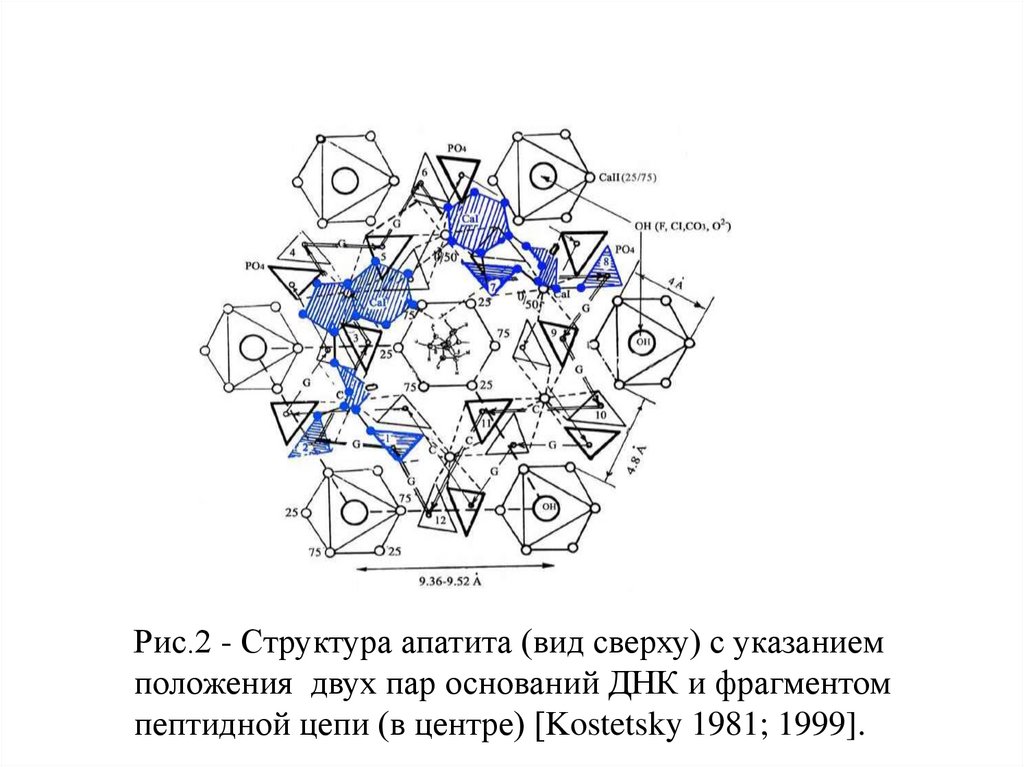
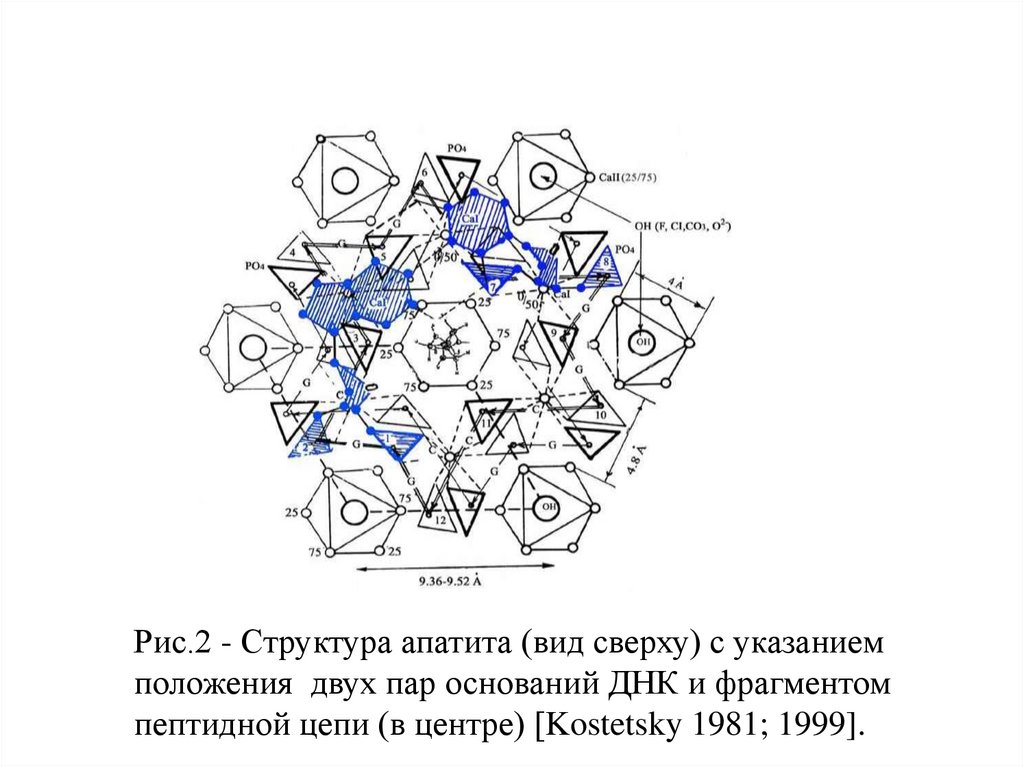
Рис.2 - Структура апатита (вид сверху) с указаниемположения двух пар оснований ДНК и фрагментом
пептидной цепи (в центре) [Kostetsky 1981; 1999].
48.
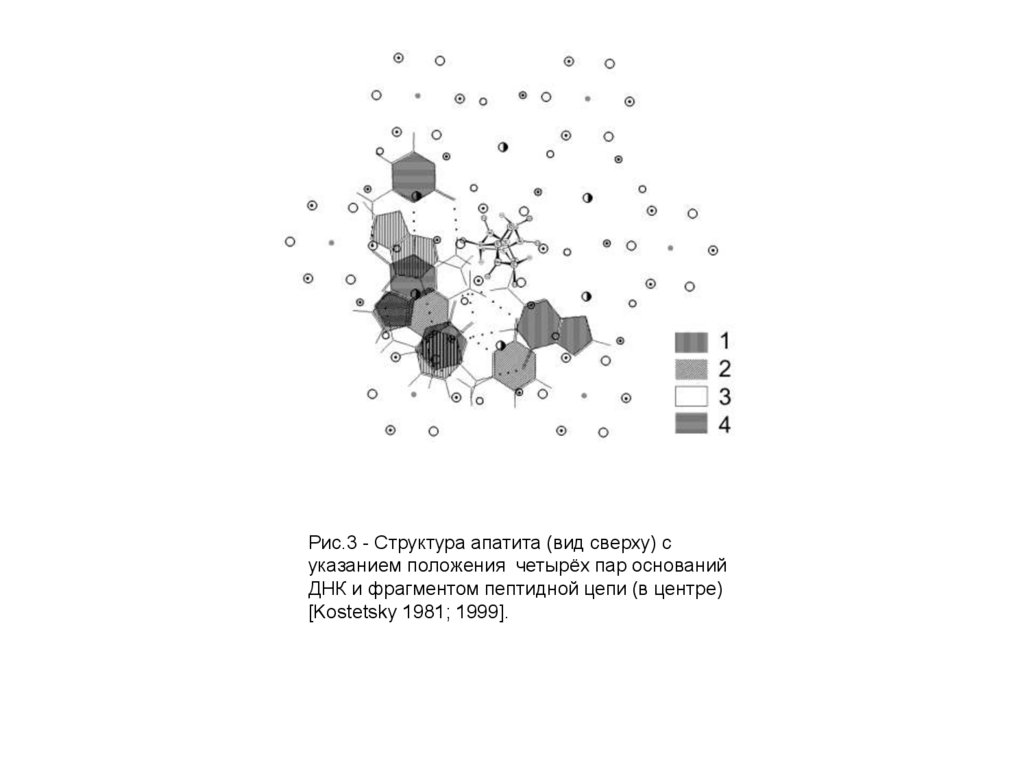
Рис.3 - Структура апатита (вид сверху) суказанием положения четырёх пар оснований
ДНК и фрагментом пептидной цепи (в центре)
[Kostetsky 1981; 1999].
49.
Z-DNA50.
АБ
В
Рис.6. А - Структура апатита с расположенными в ней
цепями полиглицина II(полипролина II, коллагена). В
зоне формирования белковых цепей в ячейке апатита
анионы РО43- замещены на CО32-. Б,В - Электронная
микрофотография гексагональной упаковки молекулы
белка в кристалле каталазы из печени быка
(увеличение Х5.105) (Б) и кристаллической
лейцинаминопептидазы (В) [Вайнштейн 1986, с.156].
51.
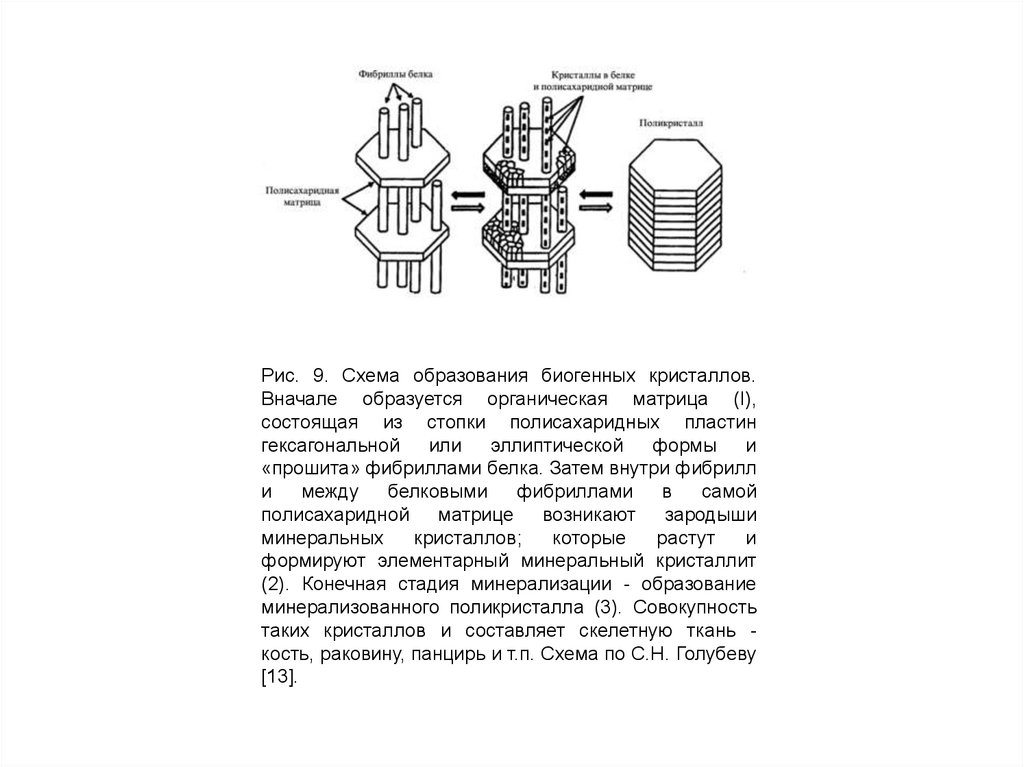
Рис. 9. Схема образования биогенных кристаллов.Вначале образуется органическая матрица (I),
состоящая из стопки полисахаридных пластин
гексагональной или эллиптической
формы и
«прошита» фибриллами белка. Затем внутри фибрилл
и
между
белковыми
фибриллами
в
самой
полисахаридной матрице возникают зародыши
минеральных
кристаллов;
которые
растут
и
формируют элементарный минеральный кристаллит
(2). Конечная стадия минерализации - образование
минерализованного поликристалла (3). Совокупность
таких кристаллов и составляет скелетную ткань кость, раковину, панцирь и т.п. Схема по С.Н. Голубеву
[13].
52.
АБ
компьютерная графика. Рис.4. А – Структура апатита (вид сбоку) построена
по данным рентгеноструктурного анализа [Kay et al. 1964; Sudarsanan, Young 1969]; Б –
Структура апатита (вид сбоку) с цепями ДНК введёнными в его состав. А, Б –
53.
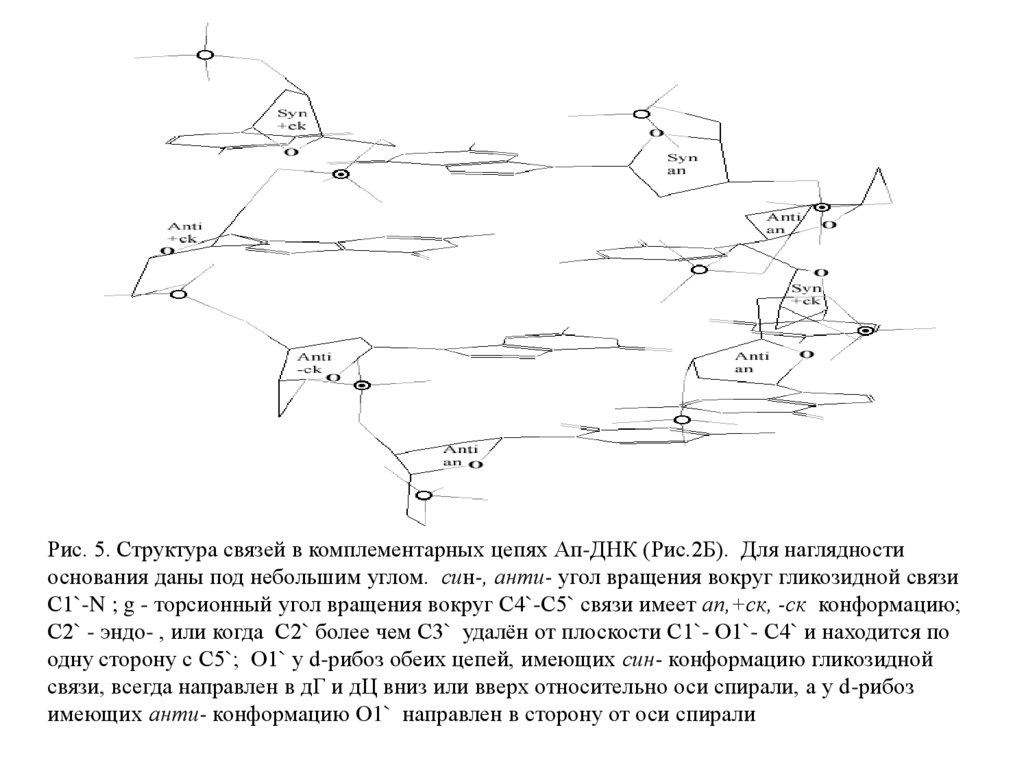
Рис. 5. Структура связей в комплементарных цепях Ап-ДНК (Рис.2Б). Для наглядностиоснования даны под небольшим углом. син-, анти- угол вращения вокруг гликозидной связи
С1`-N ; g - торсионный угол вращения вокруг С4`-С5` связи имеет an,+ск, -ск конформацию;
С2` - эндо- , или когда С2` более чем С3` удалён от плоскости С1`- О1`- С4` и находится по
одну сторону с С5`; О1` у d-рибоз обеих цепей, имеющих син- конформацию гликозидной
связи, всегда направлен в дГ и дЦ вниз или вверх относительно оси спирали, а у d-рибоз
имеющих анти- конформацию О1` направлен в сторону от оси спирали
54. Различные конформации сахара
Пятичленное фуранозное кольцоможет принимать конформацию,
подобную конверту, в которой
четыре атома из пяти находятся
в одной плоскости, а один атом
выходит из нее на расстояние до
0.5 Å, либо изогнутую твистконформацию, в которой любые
два соседних атома смещены в
разные стороны от плоскости,
построенной по трем
оставшимся атомам сахарного
кольца.
В обоих случаях атомы,
выходящие из плоскости в ту
же сторону, что и атом С5',
называют эндо-атомами, а
атомы, смещенные в
противоположную сторону,
экзо-атомами.
Фуранозное кольцо в составе нуклеозидов замещено несимметрично, поэтому в нуклеозидах и
нуклеотидах, фазовые углы псевдовращения Р распределены по циклу неравномерно. Они
образуют две группы, концентрируясь в интервалах от -1 до 34 градусов (С3'-эндо) и от 137 до
194 градусов (С2'-эндо). В соответствии с расположением в цикле псевдовращения эти
предпочтительные конформации называют северными (N) и южными (S).
•Для ДНК характерны две основные конформации сахарного кольца: С3'-эндо и С2'-эндо (или
ее разновидность, С3'-экзо - конформация), переход между которыми определяется активностью
молекул воды окружающей среды, а для РНК всегда наблюдается С3'-эндо-конформация.
54
55.
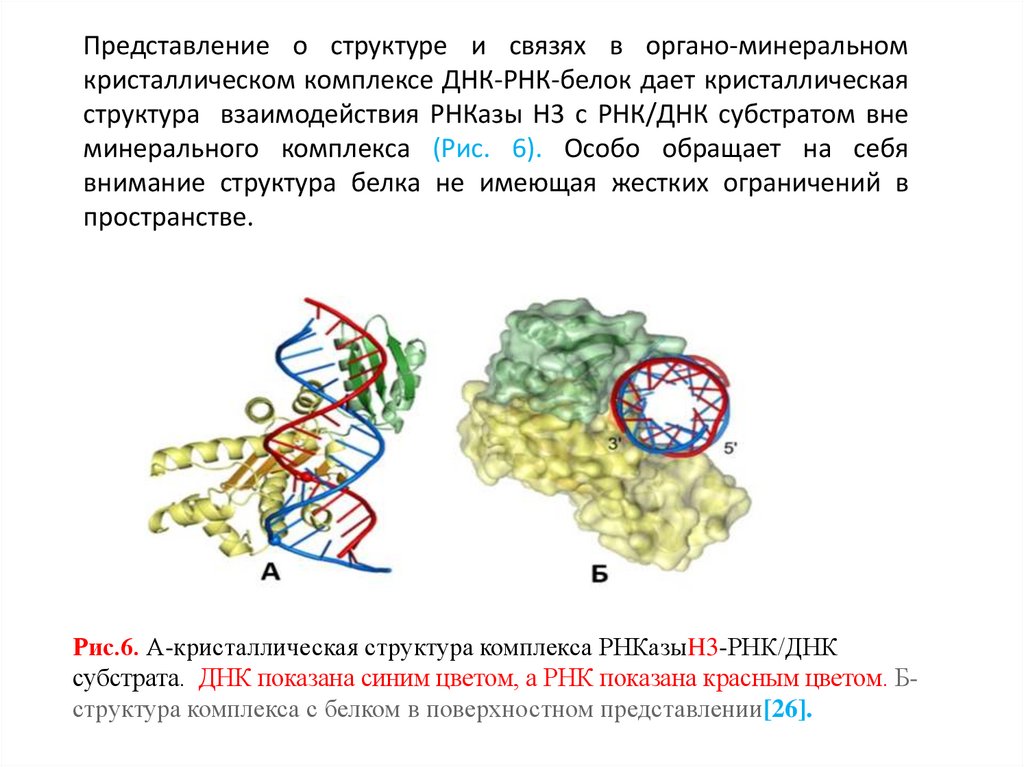
Представление о структуре и связях в органо-минеральномкристаллическом комплексе ДНК-РНК-белок дает кристаллическая
структура взаимодействия РНКазы Н3 с РНК/ДНК субстратом вне
минерального комплекса (Рис. 6). Особо обращает на себя
внимание структура белка не имеющая жестких ограничений в
пространстве.
Рис.6. А-кристаллическая структура комплекса РНКазыH3-РНК/ДНК
субстрата. ДНК показана синим цветом, а РНК показана красным цветом. Бструктура комплекса с белком в поверхностном представлении[26].
56.
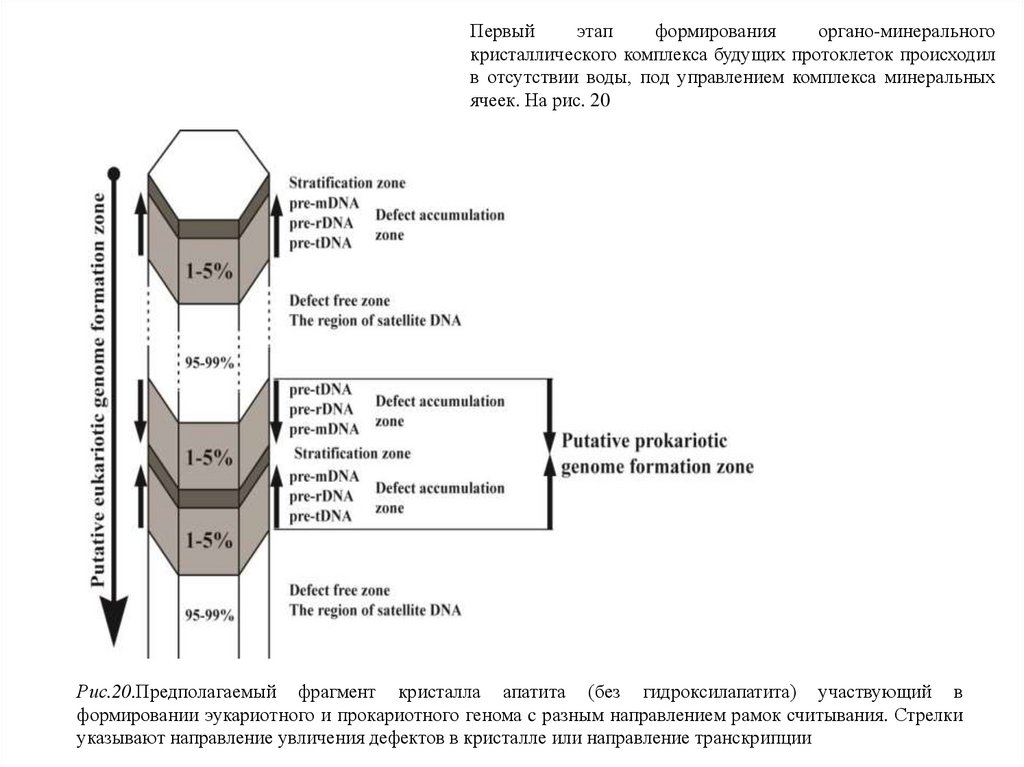
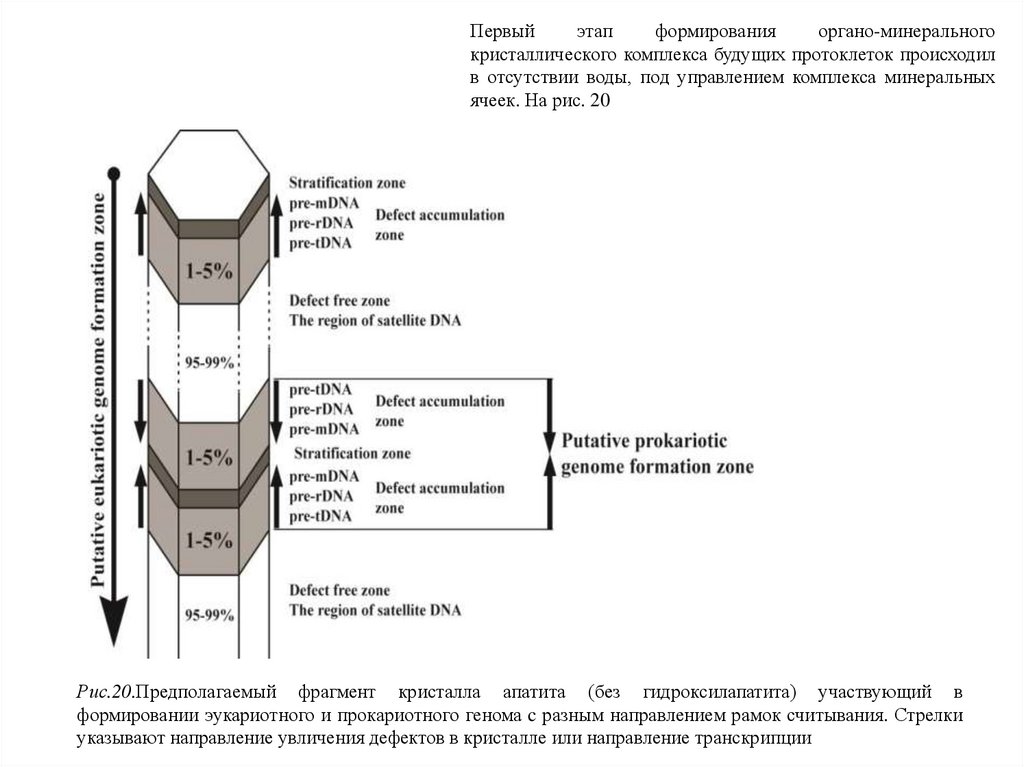
Первыйэтап
формирования
органо-минерального
кристаллического комплекса будущих протоклеток происходил
в отсутствии воды, под управлением комплекса минеральных
ячеек. На рис. 20
Рис.20.Предполагаемый фрагмент кристалла апатита (без гидроксилапатита) участвующий в
формировании эукариотного и прокариотного генома c разным направлением рамок считывания. Стрелки
указывают направление увличения дефектов в кристалле или направление транскрипции
57.
58.
Структуру и размеры формирующихся органических молекул обусловливалиструктура, параметры и расположение организующих центров (РО43ˉ-групп )
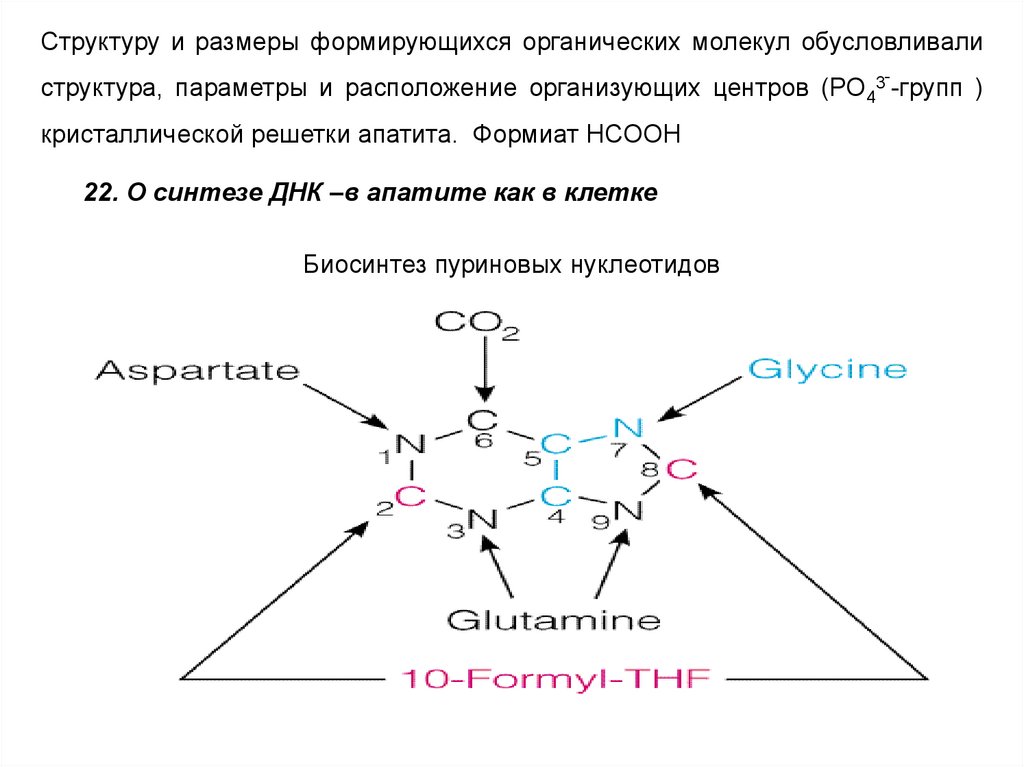
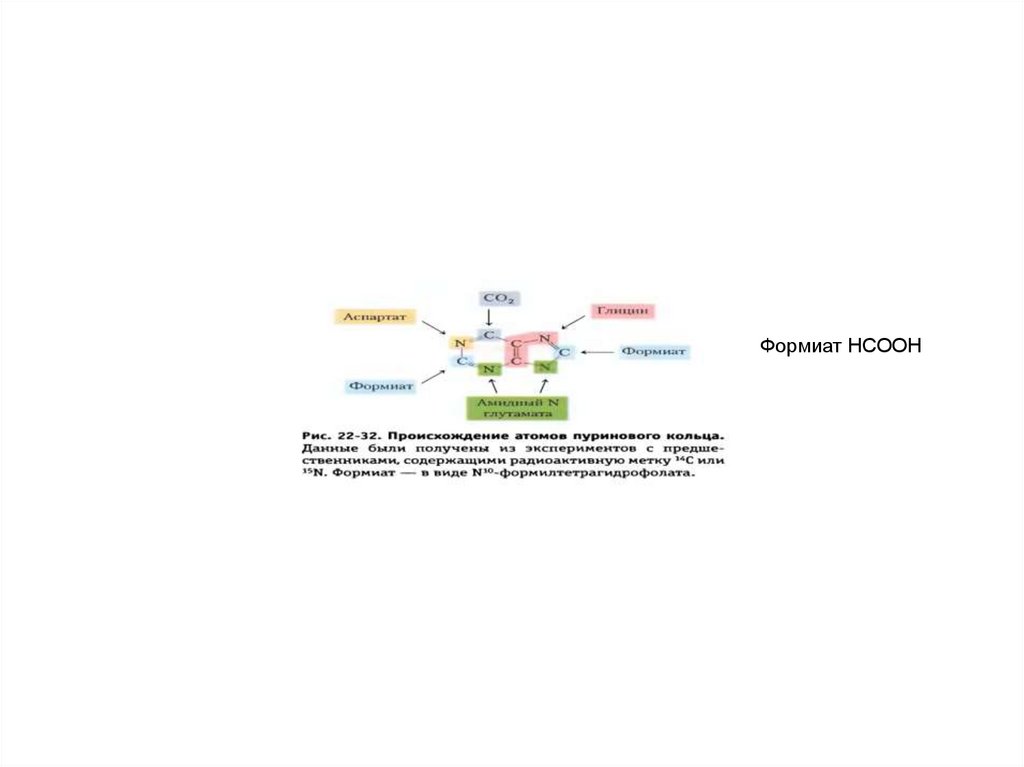
кристаллической решетки апатита. Формиат HCOOH
22. О синтезе ДНК –в апатите как в клетке
Биосинтез пуриновых нуклеотидов
59.
Формиат HCOOH60.
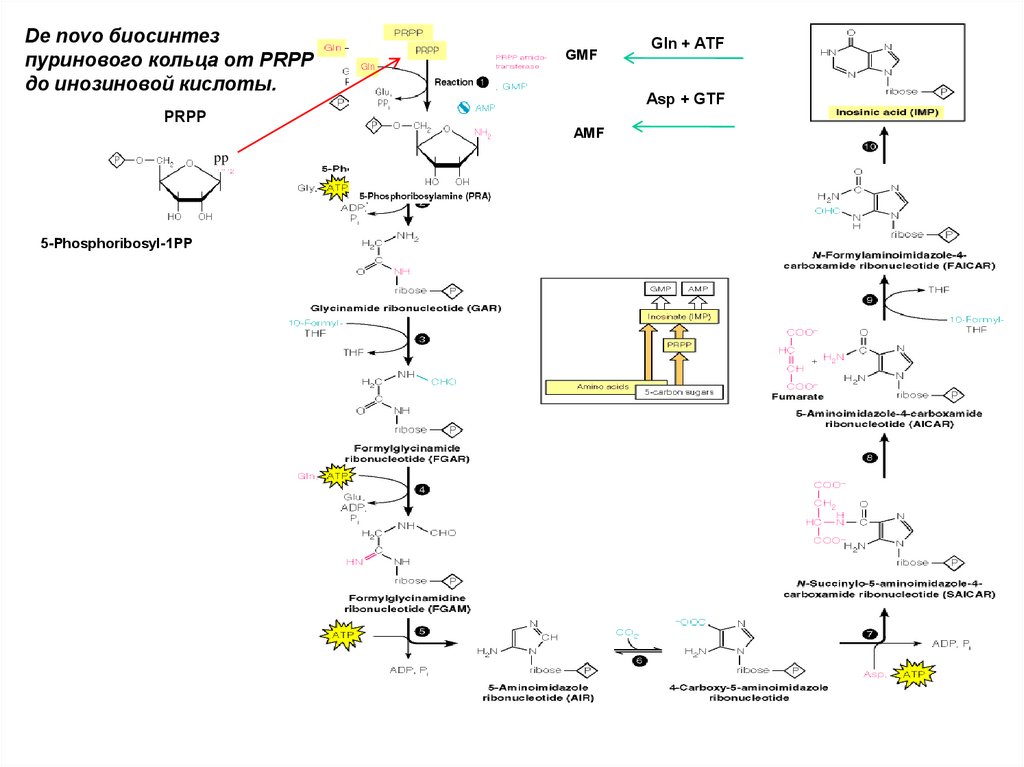
De novo биосинтезпуринового кольца от PRPP
до инозиновой кислоты.
GMF
Asp + GTF
PRPP
AMF
pp
5-Phosphoribosyl-1PP
Gln + ATF
61.
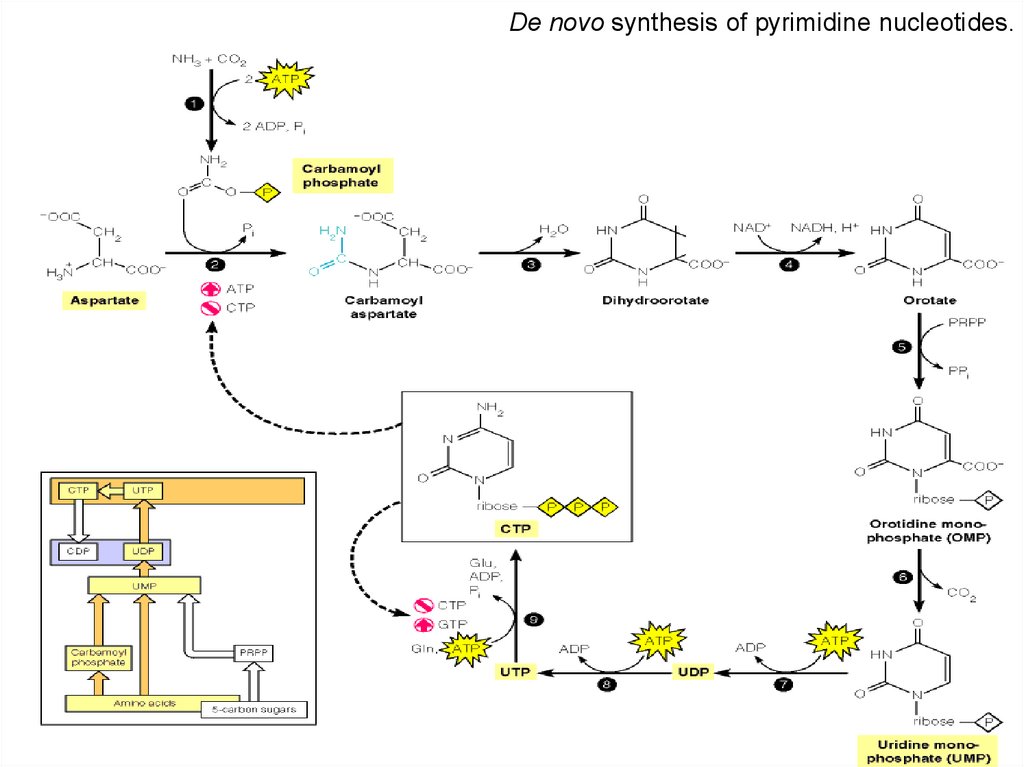
De novo synthesis of pyrimidine nucleotides.62.
Пути от инозиновой кислоты к GMP и AMP.63. Нуклеосома
64.
65. Гистоновый октамер
66.
structure.Levels of chromatin
structure.
67.
Представление о структуре и связях в органо-минеральномкристаллическом комплексе ДНК-РНК-белок дает кристаллическая
структура взаимодействия РНКазы Н3 с РНК/ДНК субстратом вне
минерального комплекса (Рис. 6). Особо обращает на себя
внимание структура белка не имеющая жестких ограничений в
пространстве.
Рис.6. А-кристаллическая структура комплекса РНКазыH3-РНК/ДНК
субстрата. ДНК показана синим цветом, а РНК показана красным цветом. Бструктура комплекса с белком в поверхностном представлении[26].
68.
Первыйэтап
формирования
органо-минерального
кристаллического комплекса будущих протоклеток происходил
в отсутствии воды, под управлением комплекса минеральных
ячеек. На рис. 20
Рис.20.Предполагаемый фрагмент кристалла апатита (без гидроксилапатита) участвующий в
формировании эукариотного и прокариотного генома c разным направлением рамок считывания. Стрелки
указывают направление увличения дефектов в кристалле или направление транскрипции
69.
70.
Comparison of the twomajor forms of DNA.
71.
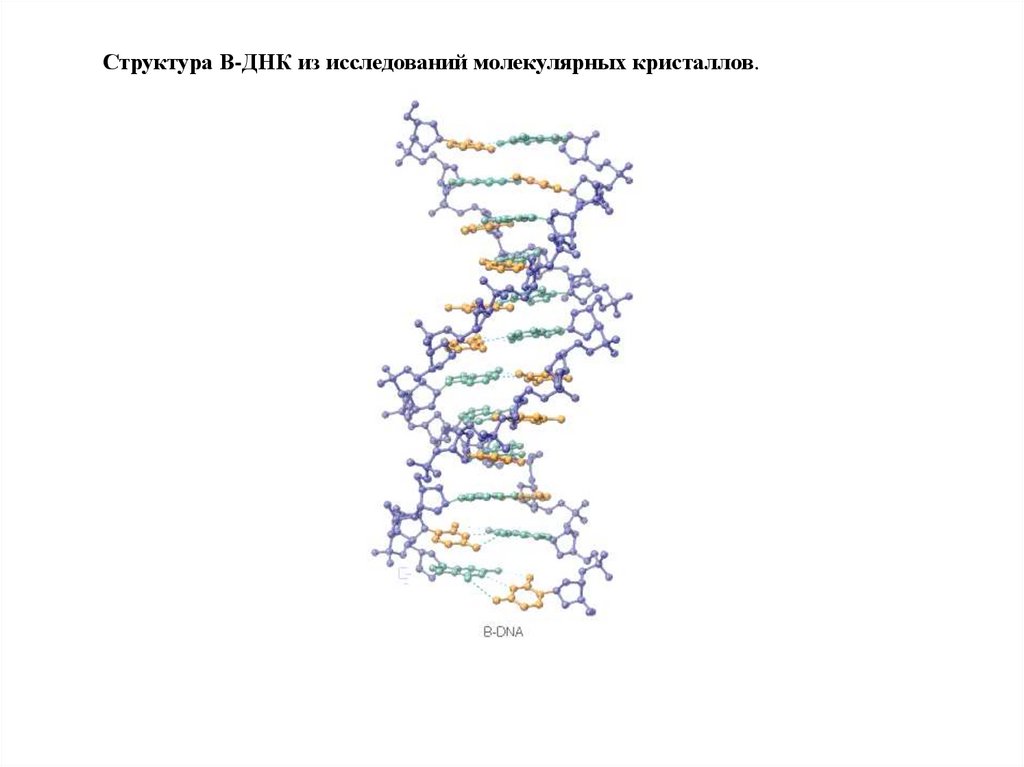
Структура B-ДНК из исследований молекулярных кристаллов.72.
Расшифровка первичной структуры:1.клонирование
2.рестриктазы
3.химические реагенты специфичные на каждый нуклеотид
4.электрофорез в ПААГ
На 5* конец вводят Р32* делят на 4 части, щепят на специфические нуклеотиды, все 4 образца
делят в ПААГ, гели наносят на рентгеновскую пленку и получают полоски с Р32.
Для получения расположения фрагментов ДНК их получают с разными рестриктазами.
Организация генома
У прокариот безядерные хромосомы
ДНК бактериофагов – все последовательности генов уникальны, без повторов
ДНК бактерий большинство генов уникальны. Несколько раз повторяются у E.coli только гены
т.РНК и р.РНК.
В геноме высших эукариот ДНК на 3-4 порядка больше, а белков на 1 порядок чем у бактерий
В геноме человека 2млрд.300мил. нуклеотидов, из них 93-95% приходится на сателитную, не
информативную часть ДНК. Это часто повторяющиеся дуплеты, триплеты или тетроплеты
нуклеотидов ( сотни тысяч или миллионы раз.)
73.
4 типа структурно-функциональной организации генов в геноме.1.Уникальные гены, имеющие специализированные функции: глобин, альбумин,
церуллоплазмин, овальбумин – представленные одной или несколькими копиями на
гаплоидный геном. Эти гены содержат интронные последовательности.
2.Уникальные гены обладающие общими функциями и активностью в большинстве
клеток. Это гены вовлеченные в регуляцию транскрипции или процессинг пре-мРНК.
3. множественные сгруппированные гены- обычно они имеют фиксированное положение
на хромасомах.
Это рибосомные гены 5S, 18S, 28S РНК, часть генов тРНК и гены
кодирующие гистоны.Отвечают за общеклеточные функции. Высоко повторенные. Гены
весьма консервативные, невелики по сравнению с конечным продуктом транскрипции,
чередуются
со
спейсерами.
Последние
обычно
гетерогенны
по
длинне
и
последовательности, часто даже в одном и том же организме, но консервативны в местах
стыковки с геном.
Предполагается, что зоны спейсеров прилегающие к генам, участвуют в регуляции
транскрипции.
74.
Траанскрипция: РНКп-за I (гены 18S,28S RNA)РНКп-за II (гистоновые гены)
РНКп-за III (гены 5SРНК и тРНК)
4. Множественные гены рассеянные по геному - это гены имеющие принципиально
разную организацию. Это часть рибосомных и гистоновых генов, рассеянных в
геноме в виде одиночных последовательностей (орфонов); часть генов тРНК
(сериновых и тирозиновых тРНК дрожжей); гены различных типов
низкомолекулярных ядерных РНК (няРНК), мобильные диспергированные гены и
«вездесущие» повторяющиеся элементы Alu, , которые существует в виде не
одинаковых фрагментов с общей длинной порядка 300 п.н., повторенных более чем
миллионом копий и занимают около 10% всей ДНК.
В целом есть две важные особенности в организации эукариотических генов:
Во-первых, большинство структурных генов эукариот присутствует в виде
мультигенных семейств, члены которых кодируют структурно- и функционально
родственные белки, экспресирующиеся на разных стадиях развития организма или
тканей.
75.
Во-вторых, они в отличие от генов прокариот в большинстве своем организованыкак отдельные транскрипционные единицы.
Интроны в генах рРНК и тРНК- как правило это один интрон; Это гены
ф.а.тРНК, сер.тРНК,лейц.тРНК – дрожжей, митохондриальные гены
рРНК
дрожжей и нейроспор, ядерные гены р.РНК тетрахимены и дрозофилы.
Гены рРНК позвоночных непрерывны.
У генов тРНК интрон всегда расположен рядом с антикодоновой петлей и частично
гомологичен антикодону.
Некоторые гены у эукариот вообще без интронов. Большинство прерывисты.
Общие черты прерывистых генов:
1.они разорваны таким образом, что всегда порядок чередования экзонов в геноме,
тот же, что и в зрелой мРНК т.е. нет хаотического перемешивания экзонов.
2.интроны ядерных генов обычно содержат большое число терминирующих
кодонов и не могут кодировать полипептид .
3.структура прерывистого гена одинакова во всех тканях независимо от того
работает он или нет.
4.прерывистыми
могут
быть
все
митохондриальные, хлоропластные.
классы
генов:
ядерные,
ядрышковые,
76.
В число генов не имеющих интронов входят: гистоновые и интерфероновые, ген тепловогошока дрозофилы, актиновый ген гриба диктиостелиум, большинство ядерных генов дрожжей.
Интроны больше распространены у высших эукариот.
У коллагена больше 50 интронов. Размер интронов от десятков до тысяч пар оснований.
Пример: мРНК коллагена – 5000п.о., а сам ген 40000п.о. По нуклеотидной последовательности
интроны сильно различаются, как внутри одного гена, так и у разных. Как правило они
относятся к фракции уникальных последовательностей ДНК. Внутри интронов встречаются
повторяющиеся элементы генома. Интроны начинаются нуклеотидами ГТ и кончаются АГ.
Очередность
вырезания
интронов
не
всегда
соответствует
последовательности
их
расположения в молекуле РНК. Иногда при сплайсинге образуются разные мРНК, но всегда у
них одинакова кодирующая зона, но отличны по 3 и 5 концам.
Структура пре-мРНК - 2-4 основных глобулярных белка и далее сидят энзимы процессинга.
77.
В информативной или ген-содержащей части ДНК64% уникальные (структурные ) гены
36% повторяющиеся до
1/4 умеренноповторяющиеся (от10 до 104 копий) на гаплоидный геном. Так рРНК до
600 штук у человека и до 1000 у животных.
Многократноповторяющиеся – тРНК, гены гистоны, гены кератины, гены
иммуноглобулинов. Такие гены обычно идут тандемом , отделяясь друг от друга
спейсером (промежутком)
В виде нескольких копий у мышей, дрожжей, дрозофил найдены мобильные
диспергированные гены МДГ
78.
4.Физико-химические свойства ДНКВ молекуле ДНК между амино- и кетогруппами азотистых оснований образуются Н- связи.
Огромная длинна по сравнению с размером ядра: онковирус SV 40 – 1.7мкм ;
хромосома мыши 3.4x104мкм; хромосома E.coli 1400мкм диаметр клетки 2мкм
Денатурация ДНК при повышении Т плавления – изменение пространственного
расположения цепей без разрыва ковалентных связей. За изменением следят с пом. УФ
260 нм. При нагревании ДНК ведет себя, как кристалл: расплетается в пределах
небольшого температ. интервала (1-20)). S образные профили Т плавления
свидетельствуют о кооперативности процесса. При отжиге (медленном обратном
охлаждении) структура ДНК восстанавливается- ренатурация. Скорость восстановления 2х
спиральных цепей ДНК высших организмов показала их высокую гетерогенность.
79.
Третичная структура ДНК бактерий, вирусов и эукариотОдно и двухцепочечные ДНК бактерий и вирусов всегда плотно упакованы,
суперспирализованны и существуют в кольцевой ковалентно-замкнутой форме. В такой
ДНК легче чем в кольцевой разводятся комплементарные цепи перед репликацией и
транскрипцией. ДНК бактерий прикреплена к внутренней стороне мембраны.
80.
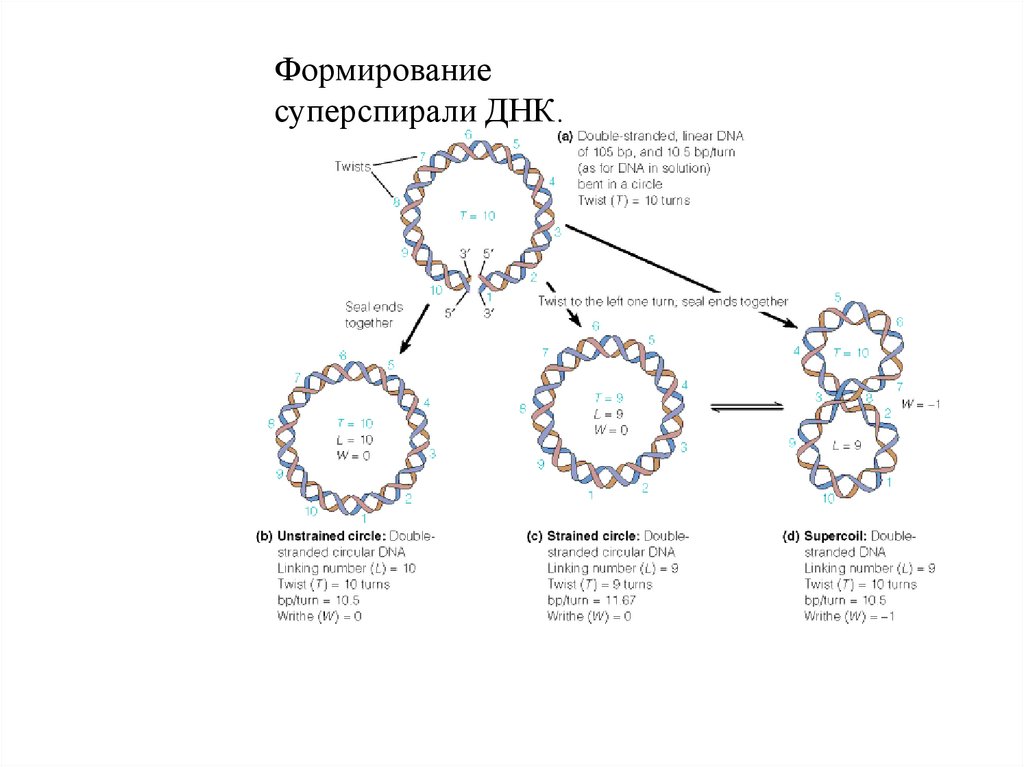
Формированиесуперспирали ДНК.
81.
ДНКэукариот
имеет
линейную
структуру,
многократно
суперспирализованную, но в отличии от прокариот в комплексе с белками.
ДНК эукариот почти вся находится в хромосомах ядер. Лишь небольшая
часть в митохондриях и хлоропластах. (cм.ниже слайд).
Хроматин содержит ДНК, гистоны, не гистоновые белки, РНК.
Гистонов 5 - Н1, Н2а, Н2в, Н3, Н4. – лизин и аргинин богатые. В одном
гистоне одна полипептидная цепь в средней части спирализованная и
скрученная в глобулу. Гистоны – ДНК ионно связаные.
Н1 - лизина больше 29%
Н2а и Н2в – 20-22% лиз, арг
Н3 и Н4 – у всех видов животных и растений имеют сходный АК состав,
содержат много аргинина и лизина.
82. Суперспирализация ДНК
83.
Рис.2 - Структура апатита (вид сверху) с указаниемположения двух пар оснований ДНК и фрагментом
пептидной цепи (в центре) [Kostetsky 1981; 1999].
84.
85. Гистоновый октамер
86.
87.
Структура ДНК имеет 5 уровней организации:1.нуклеосомный, 2.Нуклеомерный, 3.хромомерный
(петлевой домен), 4.хромонемный, 5.хроматидный
88.
89.
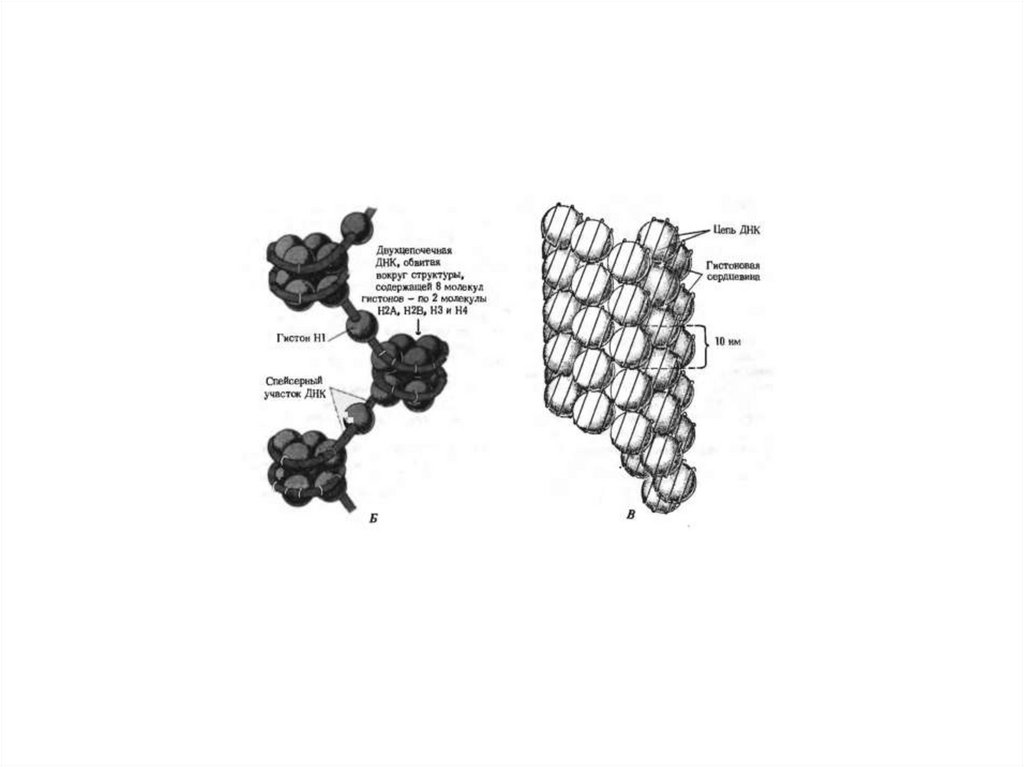
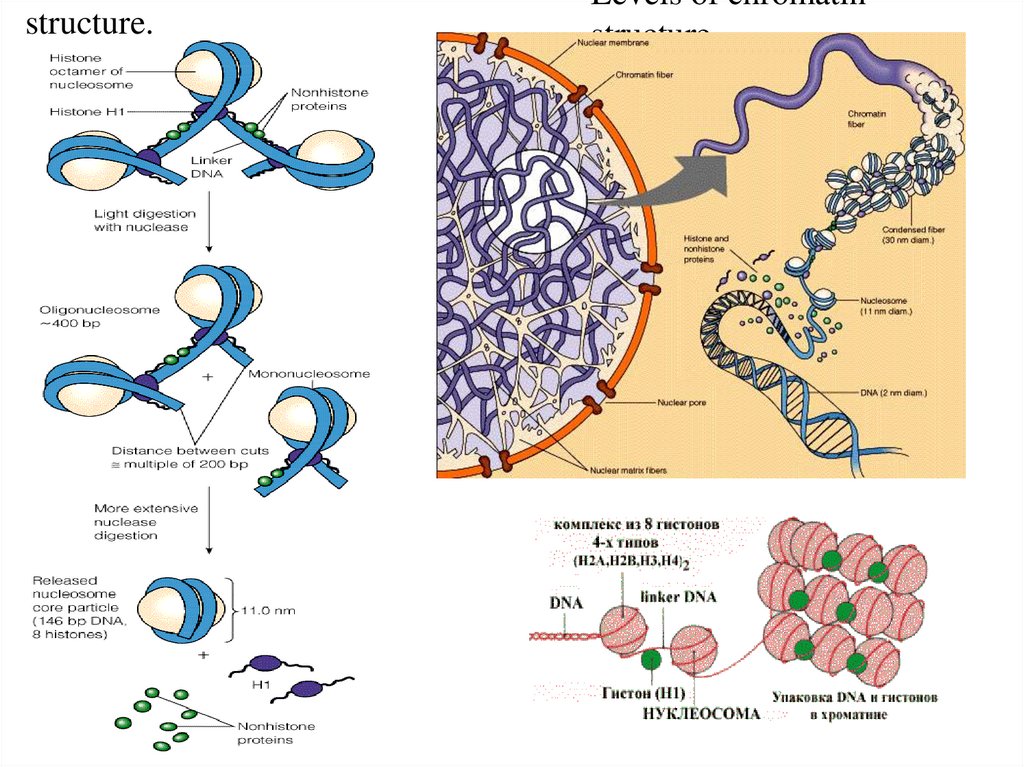
1. Уровень Нуклеосомный – исходный –Цепочка бусин с Н1 гистоном между бусин, обеспечиваетскручивание ДНК по поверхности гистоновой сердцевины.
Нуклеосомы – ДНК (160-240 пар н.)+1 Н1+(2Н2а,+2Н2в+2Н3+2Н4). На одну нуклеосому приходится два
витка ДНК из 160-240 н.п. В нуклеосоме можно выделить ядро или кор.
2й уровень. – нуклеомерный (сверхбусина) где идет объединение 8-10 нуклеосом в виде глобулы. Это
образование из нуклеосомной нити более толстых фибрилл (20-35нм). Образуются в результате
скручивания нуклеосомной нити по форме соленоида. Шаг соленоида 11нм, на один виток 8-10 штук
нуклеосом.
В сумме 1 и 2й уровень обеспечивают уменьшение линейных размеров ДНК в 40-50 раз.
3 уровень хромомерный, суперспирализованные петли , образующие розетко-видные домены ДНК,
спирализованные в структуры более высоких порядков.
4 уровень – хромонемный: сближенные в линейном порядке хромомеры образуют толстые (0.10.2мкм) нити, которые уже можно наблюдать в световом микроскопе.
5 уровень хроматида.
90. Нуклеосома
91.
92.
structure.Levels of chromatin
structure.
93.
94. Нуклеокапсид вируса табачной мозаики
1 — РНК,2 — белковая субъединица,
3 — упакованный нуклеокапсид
95.
Цитоплазматическая ДНК составляет менее 1% от суммы ДНК в митохондрияхи
хлоропластах.
Заключенная
в
ней
информация
обеспечивает
цитоплазматическую наследственность.
мтДНК – 2х цепочечная, кольцевая. В одной митохондрии от 2 до 10 молекул
ДНК в матриксе.
Все гены в мтДНК расположены компактно, почти без промежуточных
нуклеотидов. Есть исключение.
мтДНК - кодирует 2рРНК митохондриальных рибосом, все тРНК, ограниченное
число белков. Это субъединицы ферментативных комплексов внутренней
митохондриальной мембраны.
ДНК хлоропластов в 10 раз больше мтДНК животных.
96.
Бактериальные плазмидыКольцевые
молекулы
ДНК,
автономно
размножаются,
стабильно
наследуются.
Некоторые плазмиды могут включаться в хромосомы бактерий в виде эписом.
Плазмидная ДНК – 1-15% от массы хромосом ДНК бактерий. Могу нести информацию от
2 до 200 белков. Мелких плазмид в бактериях может быть десятки, крупных 1-2. Две
близкородственные плазмиды не могут существовать в одной клетке.
Многие плазмиды обеспечивают устойчивость бактерий к
антибиотикам. Передаваясь
от клетки к клетке они обеспечивают их устойчивость к действию лекарств:
пенициллину, тетрациклину и т.д.
Есть плазмиды отвечающие за способность расщеплять алифатические и ароматические
углеводороды – плазмиды биодеградации.
Способность к переносу и автономной
репликации делают плазмиды важным инструментом в генной инженерии.
встречаются в синезеленых водорослях (бактериях) и дрожжах.
Плазмиды
97.
Мигрирующие элементы ДНК бактериальных плазмид и хромосом: IS и Транспозоны.IS – или вставочные последовательности содержат только те гены, которые необходимы для
внедрения в ДНК (800-1400 н.п.) На концах содержат прямые или обращенные повторы. IS элементы
обнаружены в плазмидах, хромасомах бактерий и бактериофагов в виде повторяющихся нуклеотидных
последовательностей. Встраиваясь IS элементы могут или способствовать проявлению гена или его
блокировать.
Транспозоны – более сложные мигртрующие элементы состоящие из 3000-25000 н.п. Они ведут себя
как IS эл. часто включая их в свой состав , но содержат кроме генов, ответственных за способность
перемещаться , также дополнительные гены (например гены устойчивости к антибиотикам,
сульфаниламидам). Концевые повторяющиеся нуклеотидные последовательности – это типичный
признак мигрирующих элементов , структура необходимая для перемещения (транспозиции).
Встраивание и перемещение «прыгающих генов» контролируют специальные белки.
IS эл. и транспозоны входящие в состав плазмид или бактериофагов обуславливают встраивание этих
структур в бактериальную хромосому.
Мобильные диспергированные гены (МДГ) есть и у эукариот (5000-10000н.п.). На их концах есть
повторы по 200-500п.н. По мнению Темина МГДГ вступая в контакт с некоторыми клеточными генами
приводят к раковой трансформации клетки.
98.
Структуры РНК и место их формированияна апатите и карбонат-апатите
87-101слайд
99.
100.
Рис.2 - Структура апатита (вид сверху) с указаниемположения двух пар оснований ДНК и фрагментом
пептидной цепи (в центре) [Kostetsky 1981; 1999].
101.
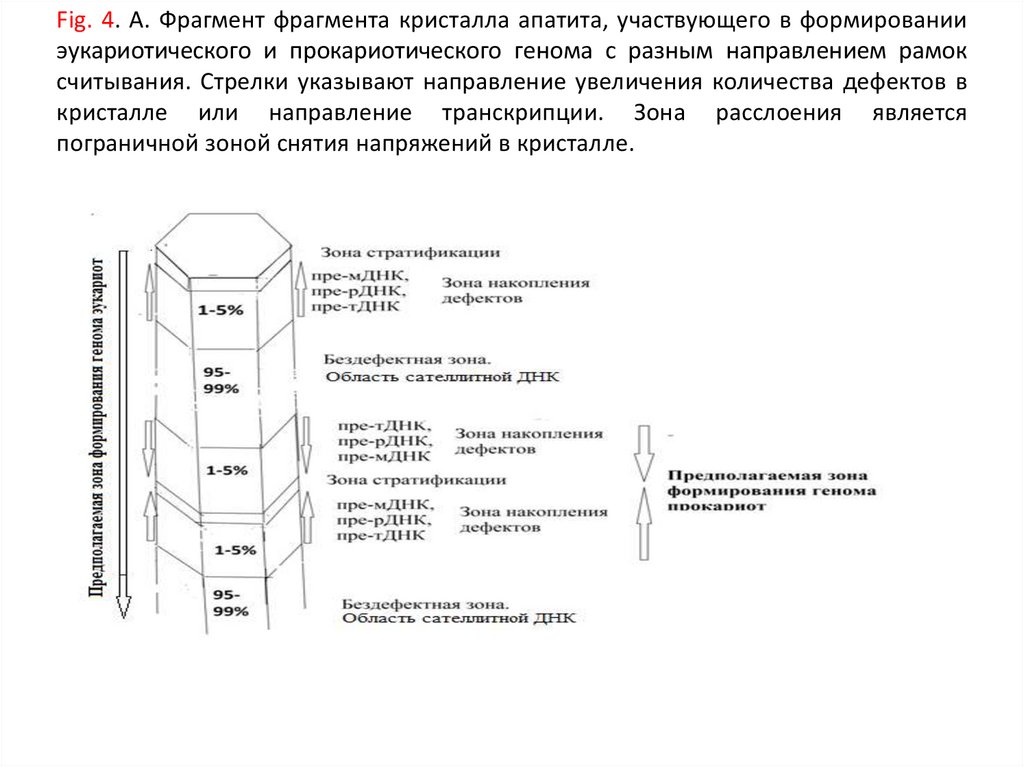
Fig. 4. А. Фрагмент фрагмента кристалла апатита, участвующего в формированииэукариотического и прокариотического генома c разным направлением рамок
считывания. Стрелки указывают направление увеличения количества дефектов в
кристалле или направление транскрипции. Зона расслоения является
пограничной зоной снятия напряжений в кристалле.
102.
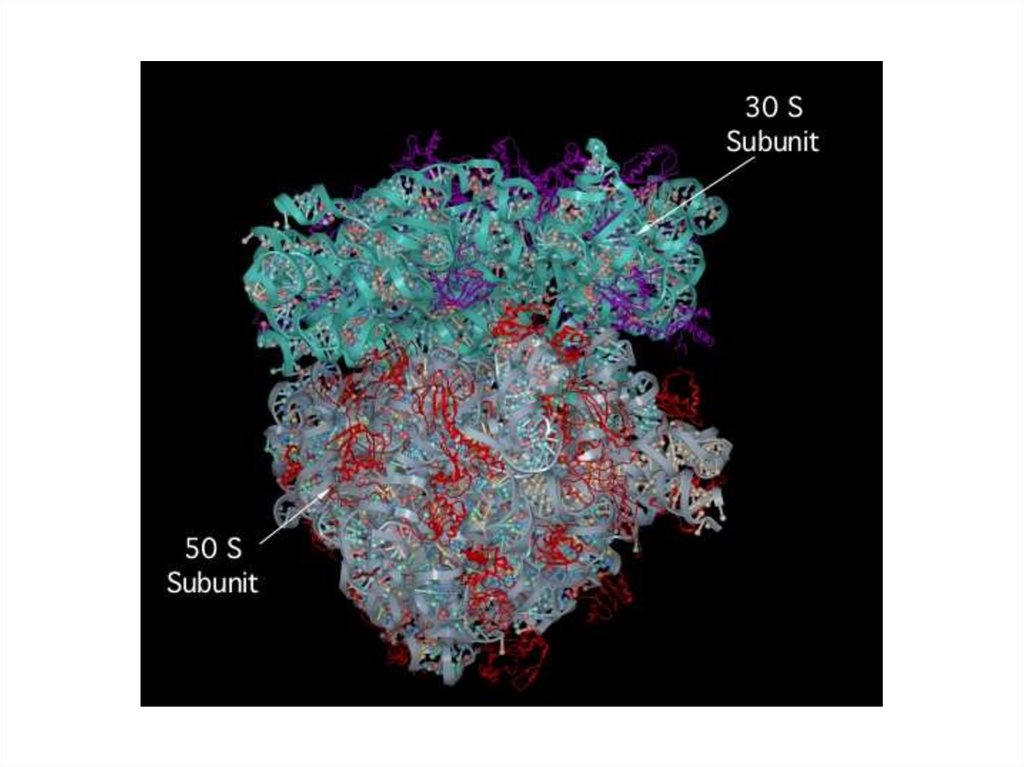
Структура рРНК (80%),70S Rb 104 (100.000 Rb в клетке)
– 50S Rb субчастица состоит из – 23SрРНК (2000-4000п.н.) +5SрРНК (60-90п.н.) +34
белка
- 30S Rb субчастица состоит из - 16SрРНК (1500-2000 п.н.) + 21
белок .
∑ индивидуальных белков в 70S Rb 55штук c м.м. от 5000 до 70000
80S Rb 106 (1.000.000 Rb в клетке)
– 60S Rb субчастица состоит из – 28SрРНК (2000-4000п.н.) + кодирующ
5SрРНК+5.8SрРНК (60-90п.н.)
+41 белок
- 40SRb субчастица состоит из - 18SрРНК (1500-2000 п.н.) + 30 белков. ∑ белков
в Rb 71 c м.м. от 107 - 10 8
103.
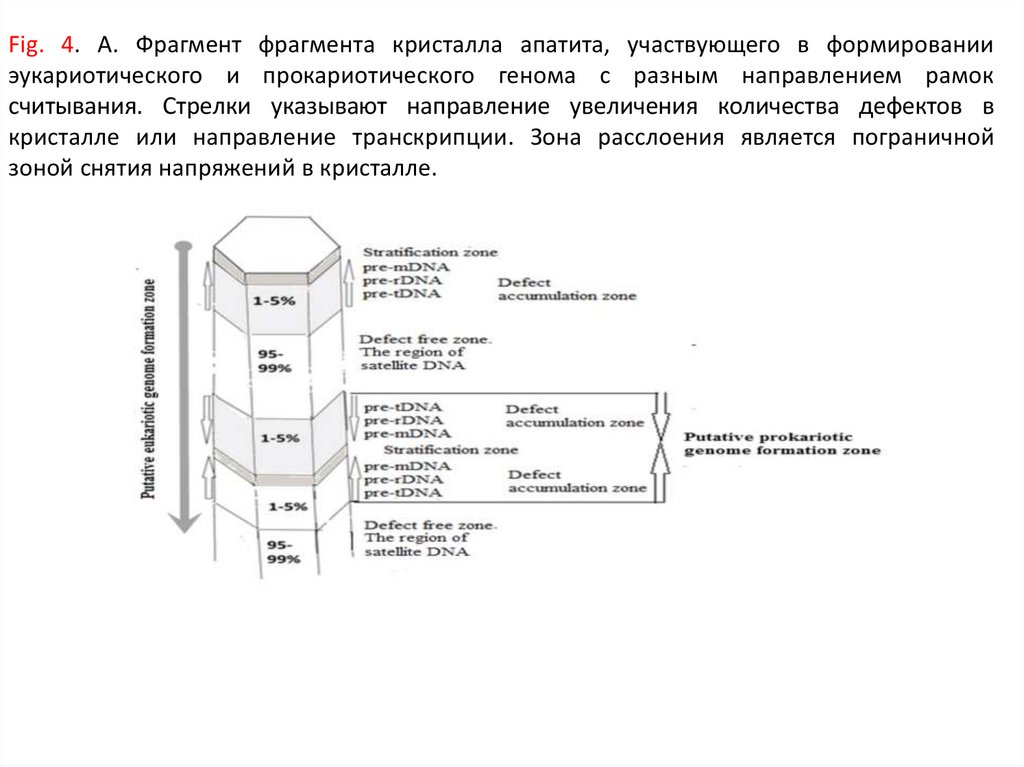
Fig. 4. А. Фрагмент фрагмента кристалла апатита, участвующего в формированииэукариотического и прокариотического генома c разным направлением рамок
считывания. Стрелки указывают направление увеличения количества дефектов в
кристалле или направление транскрипции. Зона расслоения является пограничной
зоной снятия напряжений в кристалле.
104.
105.
106.
Components of a 70S prokaryoticribosome
107.
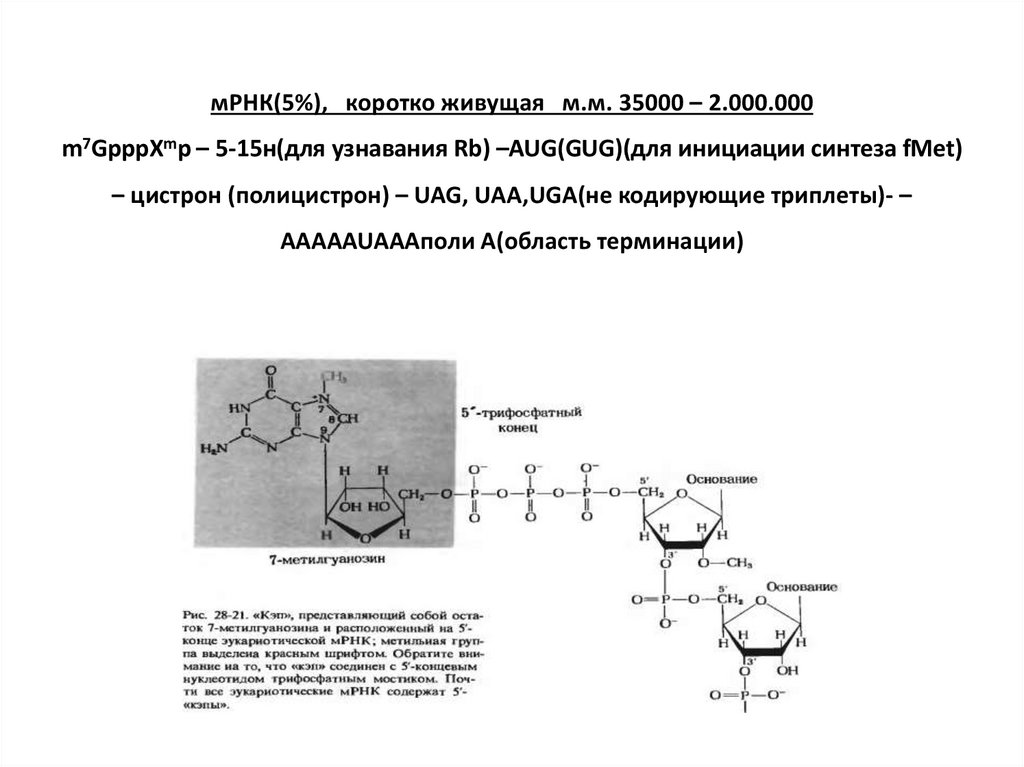
мРНК(5%), коротко живущая м.м. 35000 – 2.000.000m7GpppXmp – 5-15н(для узнавания Rb) –AUG(GUG)(для инициации синтеза fMet)
– цистрон (полицистрон) – UAG, UAA,UGA(не кодирующие триплеты)- –
AAAAAUAAAполи A(область терминации)
108.
тРНК(15%), м.м.23000-30000тРНК – 70-90нп, 60-80 шт., смесь тРНК в клетке кристаллы
ACC рецепторная ножка, ΨU петля или Т стебель (7
нуклеотидов) для связи с Rb, антикодоновая
петля(стебель)(7 нуклеотидов), UH2 петля или D стебель
(8-12 нуклеотидов) для связи с ААтРНК-синтетазой.
109.
110.
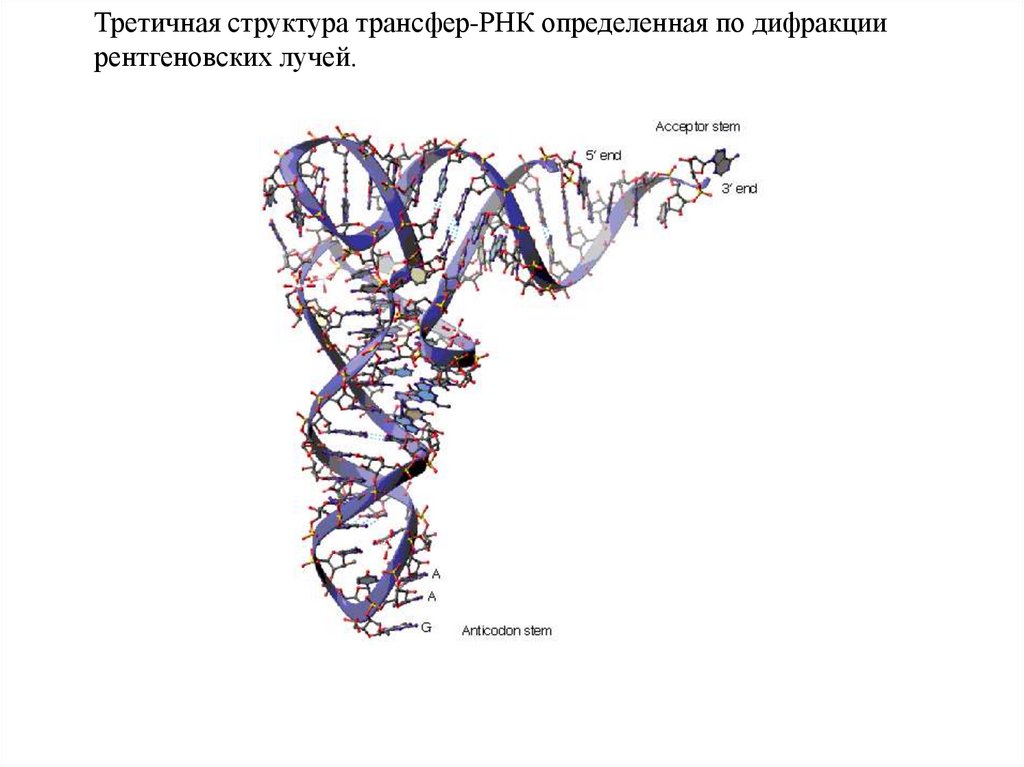
Третичная структура трансфер-РНК определенная по дифракциирентгеновских лучей.
111.
Выборкамодифицированных и
необычных оснований,
обнаруженных в тРНК
112.
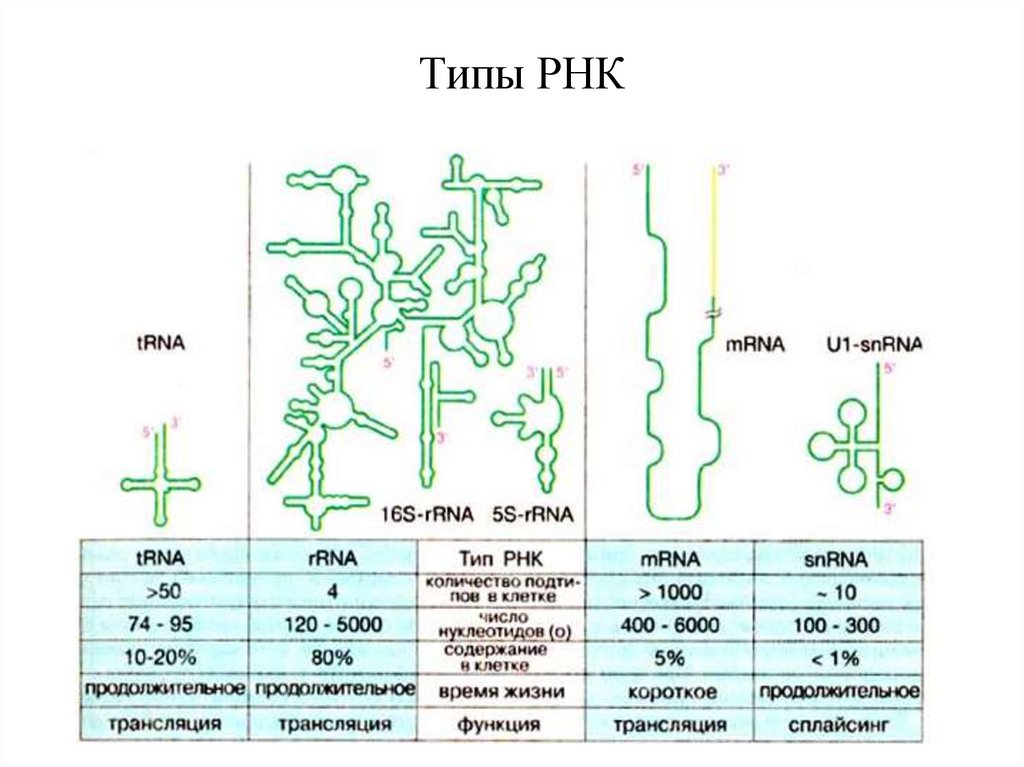
Типы РНК113.
Конец показа слайдов114.
Activation of the lac operon.Structure of E. coli 30S pre-rRNA
115.
116.
117. Разветвленные цепи ДНК
118. Метилирование ДНК
Бактериальный фермент ДНК-метилтрансферазы HhaI вместе с узнаваемым участком ДНК(фиолетовая спираль). Домен, распознающий мишень фермента образует специфический
контакт с краями оснований в большой канавке молекулы ДНК. Большой каталитический
домен (голубой) отвечает за перенос метильной группы от S-аденозил-L-метионина
(AdoMet) на цитозин. Мишень — цитозин — полностью выворачивается из двойной
спирали, глубоко погружаясь в активный центр фермента.
119. Теломеры
В клетках человека теломерыобычно представлены
одноцепочечной ДНК и состоят
из несколько тысяч
повторяющихся единиц
последовательности ТТАГГГ.
Эти последовательности
стабилизируют концы хромосом,
формируя очень необычные
структуры, называемые Gквадруплексами и состоящие из
четырёх взаимодействующих
оснований. Четыре гуаниновых
основания, все атомы которых
находятся в одной плоскости,
образуют пластинку,
стабилизированную
водородными связями между
основаниями и хелатированием в
центре неё иона металла (чаще
всего калия). Эти пластинки
располагаются стопкой друг над
другом.
120. Кроссинговер. Структура Холлидея (Holliday junction)
121.
122.
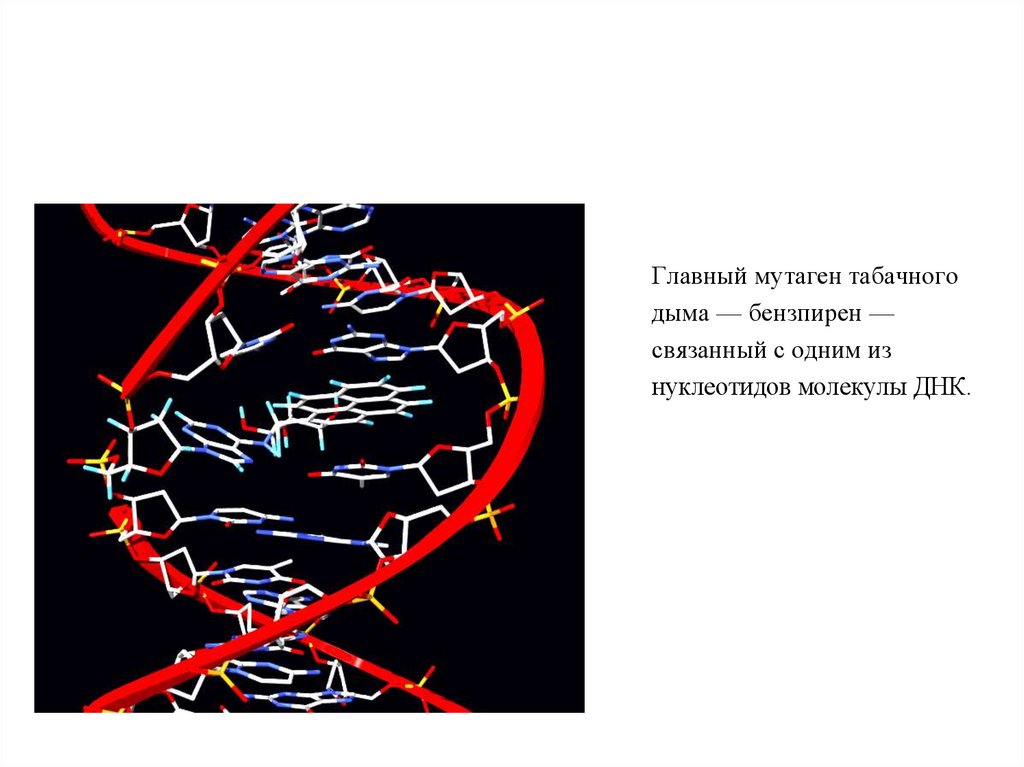
Главный мутаген табачногодыма — бензпирен —
связанный с одним из
нуклеотидов молекулы ДНК.