

































 Биология
БиологияПохожие презентации:
 наследование")









Нехромосомная наследственность
1. Нехромосомная наследственность
2.
Известно, что главными носителяминаследственности во всех известных к
настоящему времени живых системах
являются гены, локализованные в
хромосомах. Наследование этих генов
подчиняется менделевским законам
расщепления признаков в потомстве.
3.
наряду с этим объективно существует т.н. внеядерная или цитоплазматическая
наследственность, которая
обеспечивается репликацией и
транскрипцией нуклеиновых кислот,
располагающихся либо автономно в
цитоплазме клетки, либо в составе
некоторых органелл цитоплазмы (в
частности – в митохондриях и в
пластидах).
4. Нехромосомная наследственность
Основоположникомизучения внеядерной
наследственности
принято считать К.
Корренса, который в
1908 году описал
явление
пестролистности у
ночной красавицы –
признака, который
наследуется через
пластиды.
Карл Корренс
5.
Совокупность внеядерных генетическихэлементов клетки принято называть
плазмоном, а отдельные элементы этой
системы – плазмогенами. Существует
несколько основных критериев, которые
позволяют отличить
цитоплазматическую наследственность
от хромосомной
6. Критерии внеядерного наследования
Первым критерием являются различия врезультатах реципрокных по полу скрещиваний в
случаях, когда имеется явление гетерогаметности.
Такие различия обусловлены тем, что
сперматозоиды практически лишены цитоплазмы,
поэтому признаки, детерминируемые внеядерными
генами, будут наследоваться исключительно по
материнской линии. Этот критерий
цитоплазматической наследственности не
применим для тех организмов, для которых
характерно явление изогамии (грибы, водоросли),
так как количество цитоплазмы обоих родителей в
зиготе в этих случаях будет одинаково.
7. Критерии внеядерного наследования
Хромосомные гены расположены в определенныхлокусах хромосом, обнаруживают сцепление с
другими генами этой же хромосомы; их возможно
картировать. Невозможность выявить подобные
сцепления генов может свидетельствовать об их
внеядерной локализации.
Отсутствие типичного менделевского
количественного расщепления в потомстве также
может свидетельствовать о том, что данный
(рассматриваемый) признак детерминирован
внехромосомным геном.
8. Критерии внеядерного наследования
Следует подчеркнуть, что вышеперечисленныекритерии цитоплазматической наследственности,
каждый по отдельности, не являются
абсолютными, то есть наличие положительного
ответа только по одному из них не дает
возможности говорить «априори» о нехромосомной
детерминации того или иного признака.
Лишь совокупный анализ всех этих критериев
может дать утвердительный или, наоборот,
отрицательный результат.
9. Критерии внеядерного наследования
Кроме того, существует целый ряд явлений,которые имитируют цитоплазматическую
наследственность, однако не относятся к таковой.
Это случаи наследования признаков через
различные инфекции цитоплазмы, варианты
длительных модификаций и, наконец, явление
предетерминации цитоплазмы. Таким образом,
лишь полный генетический анализ во всех его
вариантах, помогает исследователям установить –
осуществляется наследование либо ядерными
генами, либо плазмагенами, либо имеет место
имитация действия плазмагенов.
10. Пластидная наследственность
Пластидная наследственность была впервыепродемонстрирована К. Корренсом в 1908 году на
примере наследования признака пестролистности у
растения ночная красавица.
Известно, что у этого растения имеется
разновидность, обладающая признаком
пестролистность (то есть на одном растении
одновременно находятся листья полностью зеленые
и листья, в которых зеленая ткань чередуется с
бесцветными полосами и пятнами – участками
тканей, лишенных хлорофилла).
11. Пестролистность у фиалки
из www.donnaflora.ru12. Пластидная наследственность
Оказалось, что если цветки у ночной красавицыобразуются на зеленых ветвях такого
пестролистного растения, то независимо от того,
какой пыльцой их опылять, они дадут семена, из
которых всегда вырастают нормальные зеленые
растения. Если же семена завязываются на
пестролистных побегах, то образуется потомство,
смешенное в различном соотношении и состоящее
из зеленых, пестролистных и неокрашенных
растений (как правило, нежизнеспособных).
13. Пластидная наследственность
Этот факт можно объяснить, предположив,что у пестролистных растений имеются два
типа пластид: нормальные и аномальные,
не способные образовывать хлорофилл. При
размножении из нормальных происходят
нормальные, а из аномальных –
аномальные пластиды. Если семяпочка
образуется на пестролистном побеге, то в ее
клетках содержится два типа пластид
(белые и зеленые).
14. Пластидная наследственность
Из такой семяпочкиобразуются яйцеклетки,
содержащие в цитоплазме
либо зеленые пластиды,
либо белые пластиды, либо
те и другие одновременно.
При этом механизм
распределения зеленых и
белых пластид является
совершенно случайным.
Соответственно, случайным
будет и распределения
признаков окрашенности,
неокрашенности и
пестролистности в
потомстве.
(из И.Ф.Жимулева, 2003)
15. Пластидная наследственность
Передача пластидной наследственностипроисходит исключительно по материнской
линии, так как пластиды находятся
исключительно в яйцеклетках, но не в
спермиях.
16. Пластидная наследственность
Это положение хорошодемонстрируется на примере
реципрокных скрещиваний
пестролистных и нормальных
зеленых растений. Если в
таком скрещивании в качестве
материнского берется
пестролистная форма, то она
будет образовывать три типа
яйцеклеток и, соответственно,
в потомстве будет наблюдаться
расщепление. Если же в
качестве материнского
растения берется нормальная
(зеленая) форма, то в этом
случае тип образующихся
яйцеклеток будет всегда один –
все с пластидами.
(из И.Ф.Жимулева, 2003)
17. Цитоплазматическая мужская стерильность
Другим примером цитоплазматической(также пластидной) наследственности
считают явление цитоплазматической
мужской стерильности (ЦМС), которая
была обнаружена у многих растений –
кукурузы, лука, льна, свеклы.
18. ЦМС у кукурузы
Кукуруза – однодомное растение; женскиецветки у нее собраны в початок, мужские –
в метелку. Иногда в метелке встречаются
недоразвитые пыльники, содержащие
стерильную пыльцу. Выяснилось, что
стерильность пыльцы определяется
некоторыми особенностями цитоплазмы.
19. ЦМС у кукурузы
Если растение, обладающее ЦМС, опылитьпыльцой взятой от других растений, то в
потомстве будут наблюдаться формы со
стерильной пыльцой. Следовательно,
признак мужской стерильности передается
по материнской линии. Даже когда все 10
пар хромосом стерильного по пыльце
растения замещали хромосомами растений с
нормальной пыльцой, то мужская
стерильность сохранялась.
20. ЦМС у кукурузы
Цитоплазму, которая обуславливает мужскуюстерильность, принято обозначать как цит(s),
соответственно, нормальную цитоплазму
обозначают как цит(n). Генетический анализ
показал, что на явление мужской стерильности,
помимо цитоплазматических факторов, влияет
также и ядерный генотип растения. Так,
цитоплазма цитs может обусловить стерильность
только при наличии в генотипе рецессивного гена
rf в гомозиготном состоянии. Если же в генотипе
присутствует доминантный ген RF (даже в
гетерозиготном состоянии), растение имеет
нормальную фертильную пыльцу. Следовательно,
ядерный ген RF способен восстанавливать
фертильность пыльцы.
21. Взаимодействие ядерных и цитоплазматических генов в формировании ЦМC у кукурузы
22. Использование явления ЦМC
Такие взаимоотношения между цитоплазмой и ядернымгенотипом позволили разработать методику и составить схему
получения межлинейных гибридов кукурузы с
использованием явления ЦМС.
Использование явления ЦМС при создании межлинейных
гибридов кукурузы имеет определенные причины. В свое
время было выяснено, что при скрещивании специально
подобранных линий можно получать гибриды, которые на 25
– 30 % превышают по урожайности лучшие сорта. Такие
линии высевают чередующимися рядами на участках
гибридизации. Однако для получения гибридных семян
необходимо было вручную удалять на растениях материнской
формы все метелки, что требовало больших затрат.
23. Использование явления ЦМC
Открытие и использованиецитоплазматической мужской стерильности
позволило коренным образом решить
проблему получения гибридной кукурузы.
В частности, путем возвратных
насыщающих скрещиваний были получены
стерильные аналоги материнских линий,
т.о. гибриды перевели на стерильную
основу, что позволило возделывать их без
затрат ручного труда на обрывание метелок.
24. Использование явления ЦМC
У многих сортов культурных растенийиспользование в сельскохозяйственной
практике гибридов стало возможным
только после открытия ЦМС, так как
ручная кастрация цветков у таких видов
как лук, огурцы, помидоры, сорго и др.
затруднена или даже невозможна.
25. Геном митохондрий человека
- представлен однойкольцевой молекулой
ДНК размером 16 569
пар нуклеотидов. Он
кодирует 13 белков, 22
(все) транспортные
РНК, две рибосомные
РНК.
из chemistry.umeche.maine.edu
26. Геном митохондрий человека
60% генов, кодирующих белки, приходится на семьсубъединиц комплекса, окисляющего НАДН;
остальные гены кодируют две субъединицы АТФсинтетазы, три субъединицы цитохромоксидазы,
одну субъединицу цитохрома b.
В транскрибируемых и транслируемых областях
цепей митохондриальной ДНК выявлено меньшее
число некодирующих участков по сравнению с
ядерной ДНК. Установлено также, что по целому
ряду характеристик генетический код
митохондриальной ДНК отличается от
универсального.
27. Дефекты митохондриального генома и патология
Поскольку митохондрии находятся в центреэнергетического обмена клетки, любое нарушение в
них может негативно воздействовать на основные
функции организма. В настоящее время выявлен
ряд генетических заболеваний, связанных с
нарушениями митохондриальных процессов,
многие из которых обусловлены дефектами
митохондриального генома. В частности, некоторые
митохондриальные дефекты приводят к
нарушению работы сердечной мышцы. Поскольку
миокард зависит от окислительного метаболизма
митохондрий, неудивительно, что генетические
нарушения митохондриальной функции приводят к
кардиомиопатиям.
28. Дефекты митохондриального генома и патология
Известны также заболевания, связанные сдефектами структурных митохондриальных
белков, импорта белков в митохондрии,
передачи сигналов между
митохондриальным и ядерным геномом и
другие формы патологий. В этой связи
возникла и интенсивно развивается целая
отрасль медицинской генетики человека –
митохондриальная медицина.
29. Особенности митохондриальной ДНК
Наследование строго по материнской линии.Отсутствие рекомбинации
Отсутствие интронов, защитных гистонов и
эффективной ДНК-репарационной системы.
Высокая скорость спонтанного
мутирования.
30. Эти свойства сделали мтДНК неоценимым инструментом для врачей, судебных медиков, биологов-эволюционистов и историков
В целом скоростьнакопления мутаций в
мтДНК в 10 раз больше, чем
в ядерной. В ней есть один
интересный участок
примерно в 300
нуклеотидов (показан
сверху на рисунке),
называемый
гипервариабельным, в
котором скорость
накопления мутаций в 10
раз выше, чем в остальной
части мтДНК.
Из bio.fizteh.ru/student/files/biology/biolections/
31. Эти свойства сделали мтДНК неоценимым инструментом для врачей, судебных медиков, биологов-эволюционистов и историков
Скорость накопления нуклеотидных заменв гипервариабельном участке мтДНК
человека: 1 нуклеотид в 18 - 20 тысяч лет.
Зная скорость накопления мутаций можно
проводить сравнения по мтДНК между
представителями ныне существующих
народов и рас, а также современными
человекообразными обезьянами и
образцами мтДНК, полученными из
ископаемых остатков.
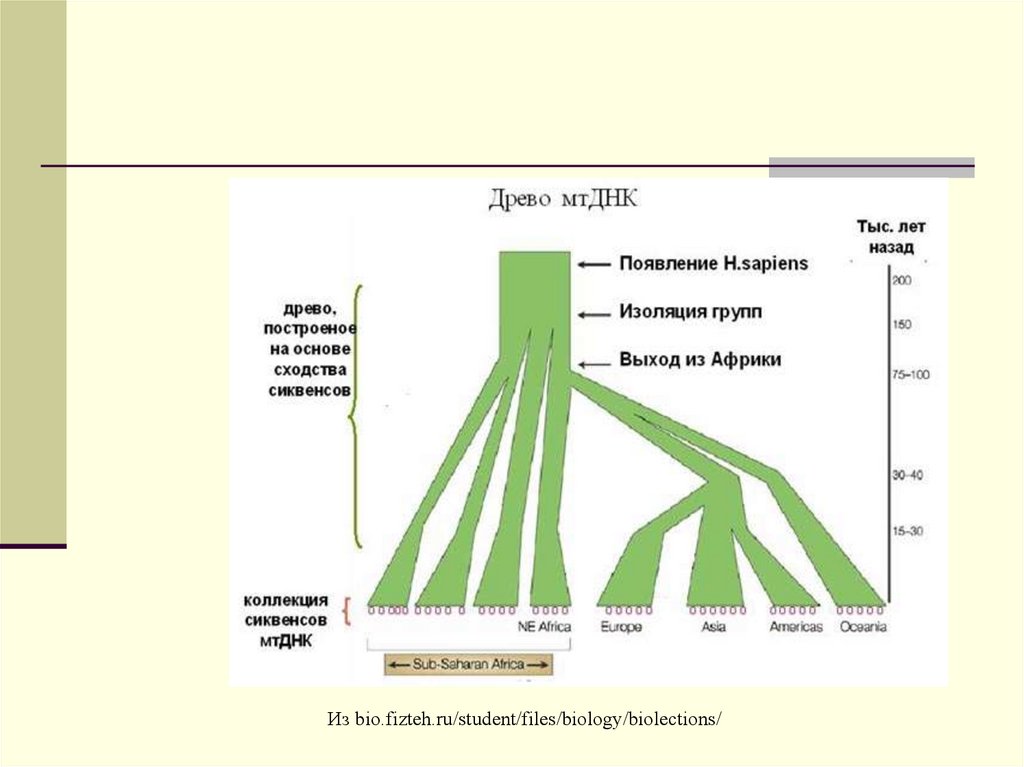
32.
Т.о. высокая скоростьмутирования делает
мтДНК наиболее
подходящей для
выяснения вопросов
эволюции человека.
Из bio.fizteh.ru/student/files/biology/biolections/
33.
Из bio.fizteh.ru/student/files/biology/biolections/34. Причины существования цитоплазматической наследственности
- как полагают, кроются в необходимостиобеспечения большей гибкости в процессе
развития клетки. Известно, что репликация
хромосом всегда тесно связана с циклом
клеточного деления, в то же время
органоиды, в том числе пластиды и
митохондрии, развиваются относительно
независимо от клеточного деления, при этом
непосредственно реагируют на воздействия
окружающей среды.
35. Причины существования цитоплазматической наследственности
Поэтому, с точки зрения природнойцелесообразности желательно, чтобы часть
генетической информации, могла бы
претерпевать репликацию независимо от
ДНК хромосом. По-видимому,
внехромосомные генетические системы
имеют определенное значение для более
быстрой реакции на меняющиеся условия
среды.